
Abstract
We report the establishment of a novel platform to induce myogenic differentiation of human mesenchymal stem cells (hMSCs) via focal adhesion (FA) modulation, giving insights into the role of FA on stem cell differentiation. Micropatterning of collagen type I on a polyacrylamide gel with a stiffness of 10.2 kPa efficiently modulated elongated FA. This elongated FA profile preferentially recruited the β3 integrin cluster and induced specific myogenic differentiation at both transcription and translation levels with expression of myosin heavy chain and α-sarcomeric actin. This was initiated with elongation of FA complexes that triggered the RhoA downstream signaling toward a myogenic lineage commitment. This study also illustrates how one could partially control myogenic differentiation outcomes of similar-shaped hMSCs by modulating FA morphology and distribution. This technology increases our toolkit choice for controlled differentiation in muscle engineering.
Introduction
The interactions between cell and implant materials are mainly mediated by the focal adhesion (FA) complex [12]. Generally, FA of cells on a 2D matrix is normally 1.5–3-μm long with an aspect ratio (AR) of 3, whereas in a 3D matrix, this is 19-μm long with an AR of 33, indicating that the FAs in a 3D environment would adopt a more elongated shape [13]. The FA complex contains signaling components such as different types of integrins and FA kinase, which sense the microenvironment and initiate signal transduction through downstream effectors such as RhoA and Rac1 [14]. Integrins consist of 24 known subunits and serve as transmembrane mechanical links from those extracellular contacts to the cytoskeleton inside cells [15]. However, each pair of α and β of integrin plays a different role in substrate sensing. For example, α2β1-integrin modulates epithelial differentiation, and β3-integrin mediates myogenic differentiation of C2C12 cells [16,17]. In addition, FA also acts as the attachment site between a cell and a matrix, where the cell exerts forces on the matrix and transfers the feedback forces into the nucleus through the cytoskeleton [8]. However, there are limited studies using FA modulation to induce stem cell differentiation. Among these, Goffin et al. employed micropatterning to modulate FA development in myofibroblasts and found that the FA length controls the transition of smooth muscle actin (SMA) into stress fibers in a tension-dependant manner [18]. Seo et al. found that FA maturation and actin polymerization were promoted via the RhoA pathway when MSCs were cultured on microtopographical substrates [19]. However, so far, there are no reports on the synergistic study of FA modulation, matrix stiffness, and ECM ligands on stem cell differentiation; let alone the more important mechanistic studies of these interactions, which we would like to address in this article.
FA may activate the RhoA-signaling pathways and result in the synthesis and reorganization of actin filaments as well as stress fiber formation, which in turn regulates the FA assembly [20]. To create contraction in the cytoskeleton for myogenic differentiation, it is well known that myosin light-chain kinase (MLCK) is needed to phosphorylate myosin light chain (MLC) and actomyosin II [21]. It has also been shown that elongated FA would increase the expression of MLCK and cellular tension [18]. Based on the literature and above cues, we hypothesize that by generating elongated FA in hMSCs cultured on compliant surfaces, myogenic differentiation could be induced. The potential mechanism behind may be the activation of specific integrins and the corresponding downstream signaling pathways. Moreover, elongated FA may change the cytoskeleton development and arrangement, and modify cellular tension that subsequently drives hMSCs into myogenic differentiation.
Here, we report a novel bioinspired platform to combine FA modulation, matrix stiffness, and ECM ligands to synergistically influence hMSCs toward specific myogenic differentiation. Results showed that hMSCs with elongated FA would go to myogenic differentiation. This was achieved by recruitment of integrin β3 (ITB3) cluster and activation of RhoA signaling in a tension-dependent manner. To demonstrate that the myogenic differentiation is primarily driven by elongated FA and not cellular shape, we carried out an experiment to decouple the 2 effects. Similar-shaped hMSCs with a different FA morphology and distribution were induced using the micropatterning technique, and it was found that the elongated FA was more supportive of myogenic differentiation. Collectively, this study demonstrates a novel and feasible method of inducing myogenic differentiation of hMSCs and improves our understanding of the micropatterning platform in effecting the differentiation.
Materials and Methods
Substrate fabrication and surface activation
PA gel was made according to the method described by Tse and Engler [22]. Briefly, aminosilanated coverslips (diameter was 22 mm) were obtained by treating with 0.1M NaOH, 3% 3-aminopropyltriethoxysilane (Sigma) in deionized (DI) water, and 0.5% (v/v) glutaraldehyde (Sigma) in phosphate-buffered saline (PBS) as described in Supplementary Fig. S1a (Supplementary Data are available online at
Stamp fabrication and microcontact printing
The silicon master templates bearing the desired topographic features were fabricated by standard photolithography, and stamps were made according to methods previously described [24]. For patch-stamp fabrication, flat mold-casted 1:10 (crosslinker: base) polydimethylsiloxane (PDMS, Sylgard 184; Dow Corning) was cut into round shape with a diameter of 22 mm as a stamp without patterns. The surface of stamps was examined under a scanning electron microscopy (SEM; JEOL6360). To pattern FN, the human FN (50 μg/mL; BD Bioscience) solution was added onto each stamp for 1 h and then blow-dried with nitrogen gas. To pattern collagen, collagen from rat tail (COLI, 200 μg/mL; Invitrogen) was oxidized with 4 mg/mL sodium periodate in 50 mM sodium acetate buffer for 30 min according to the method described before [25]. Afterward, the COLI solution was added onto stamps for 1 h followed by blowing dry with nitrogen gas. The stamps with FN or COLI were then gently pressed on an activated PA gel for 30 min followed by incubation with 2% bovine serum albumin (Invitrogen) at 37°C for 1 h for blocking. Lastly, the PA gel was washed with PBS and sterilized by UV for 15 min before cell culture. The adsorption of COLI and FN on the PA gel with different stiffness was tested by the LUCY® 506 solution (Sigma) and was analyzed by using ImageJ. The fluorescent intensity data are shown in Supplementary Fig. S2.
Cell seeding, real-time PCR, and immunostaining
Bone-derived hMSCs were commercialized cells from bone marrow (Lonza) and were seeded at a density of 1,000 cells/cm on PA gels with various surface treatments and were cultured as previously described [26]. Unattached cells were washed away around 45 min after cell seeding. The complete culture medium was low-glucose Dulbecco's modified Eagle medium (DMEM) (Sigma) supplemented with 10% (v/v) fetal bovine serum and 1% antibiotics (PAA Laboratories), and the medium was replaced every 2–3 days. On the seventh day, the RNA of the cell was isolated with an RNeasy mini kit (Qiagen) and was synthesized into cDNA (iScript™ cDNA Synthesis Kits; Bio-Rad). The real-time PCR was performed on a CFX96 real-time PCR detection system (Bio-Rad) with KAPA SYBR FAST master mix universal (Kapa Biosystems). Primers specific to the targeted genes were obtained from the primer bank [27] and are listed in Supplementary Table S1. GAPDH and β-actin were used as endogenous housekeeping genes, and the heat map was generated using Matrix2png [28]. The relative quantification of gene expression was analyzed with CFX manager software (Bio-Rad Laboratories, Inc.), and the Relative Expression Software Tool 2009 (REST 2009). Immunostaining was performed according to the methods described previously [26]. The primary antibodies used were mouse anti-human vinculin (1:400; Millipore), rabbit anti-rat collagen (1:200; Novus Biologicals), rabbit anti-human FN (1:1,000; Sigma), mouse anti-human cardiac myosin heavy chain (MHC, 1:200; Abcam), mouse anti-human α-sarcomeric actin (1:500; Sigma), mouse anti-human MLCK (1:100; Sigma), and mouse anti-human ITB3 (1:100; Millipore). The secondary antibodies were Alexa-Fluor® 488 goat anti-mouse IgG (1:400; Invitrogen) and Cy3 goat anti-rabbit IgG (1:400; Millipore). Cytoskeletal actin was stained with tetramethyl rhodamine-isothiocyanate (TRITC)–phalloidin (1:400; Millipore). The cell nucleus was stained with 4',6-diamidino-2-phenylindole (DAPI). C2C12 myoblasts (ATCC) were used as the positive control for MHC. The negative control was performed without a primary antibody. Images were taken using an Eclipse 80i microscope (Nikon) using 20× objective lens and were analyzed with ImageJ 1.44f software [29]. The FA size, length, and AR were analyzed using the analyze particles function by ImageJ. Briefly, the fluorescent image of vinculin was converted to 8 bits, and the background was removed by using the subtract background function at same level (rolling ball radius was set as 10). This was then followed by using the adjust threshold and then analyze particles function. The area parameter to be detected was set to 0.8–40 μm2.
GTpase-linked immunosorbent assay test and MLCK inhibition assay
hMSCs on a patterned and patch 10.2-kPa PA gel with COLI were lyzed with 50 μL of lysis buffer (Cytoskeleton, Inc.), and the cell lysate was collected on ice. Five replicates were collected for each sample type. GTpase-linked immunosorbent assay (G-LISA) (Cytoskeleton, Inc.) is a novel small G-protein activation assay, and it was used to measure the activation of RhoA and Rac1 on 10.2-kPa COLI-patterned groups in this work according to the manufacturer's instruction. All the tests were triplicate, and the absorbance was read with a microplate spectrophotometer (Tecan) at the wavelength of 490 nm. MLCK was blocked by 10 μM of 1-(5-iodonaphthalene-1-sulfonyl) homopiperazine (ML-7; Merck). Briefly, ML-7 was dissolved in dimethyl sulfoxide (DMSO) to make a 10 mM stock solution. On day 3 after cell seeding, 10 mM ML-7 was diluted in a complete DMEM to a final concentration of 10 μM to make the inhibition medium, and replace the normal DMEM in the culture plate. The blocking medium was replaced every 2 days. As for control group, cells were incubated with a complete medium supplemented with 0.1% DMSO. On day 7, cells were fixed for immunostaining.
Statistical study
All assays were triplicated for each group, and the results were expressed as means±standard deviation. Statistical analysis was performed by using a one-way ANOVA followed by Tukey's pair wise comparisons. A value of P<0.05 was considered statistically significant.
Results
Elongated FA was successfully modulated cooperatively with substrate stiffness and ECM ligands
The average shear storage moduli G′ of the PA gel fabricated were 13.7±0.93 kPa and 3.43±0.06 kPa, respectively. As the Poisson ratio of PA gel is 0.48 [30], the Young's modulus E of the PA gel was about 40.7±2.8 kPa and 10.2±0.2 kPa, respectively. The morphologies of the stamps used for patterning were shown in Fig. 1a–c. Pattern L20 (12, 6) consisting of rectangular islets of size 20×2 μm was designed to induce elongated FA according to a previous report [18]. We optimized the vertical spacing by changing it from 6 to 12 μm to guide cell growth along the direction of a higher ECM density, that is, denser islet to form elongated FA. The square islets in patterns S6.3 (12, 6) and S6.3 (6, 6) as shown in Fig. 1b and c had the same dimension of 6.3×6.3 μm while maintaining a similar area with the L20 pattern. However, the horizontal spacing between the islets was different (6 and 12 μm, respectively), and this was designed to explore the effects of spacing on cell behaviors.
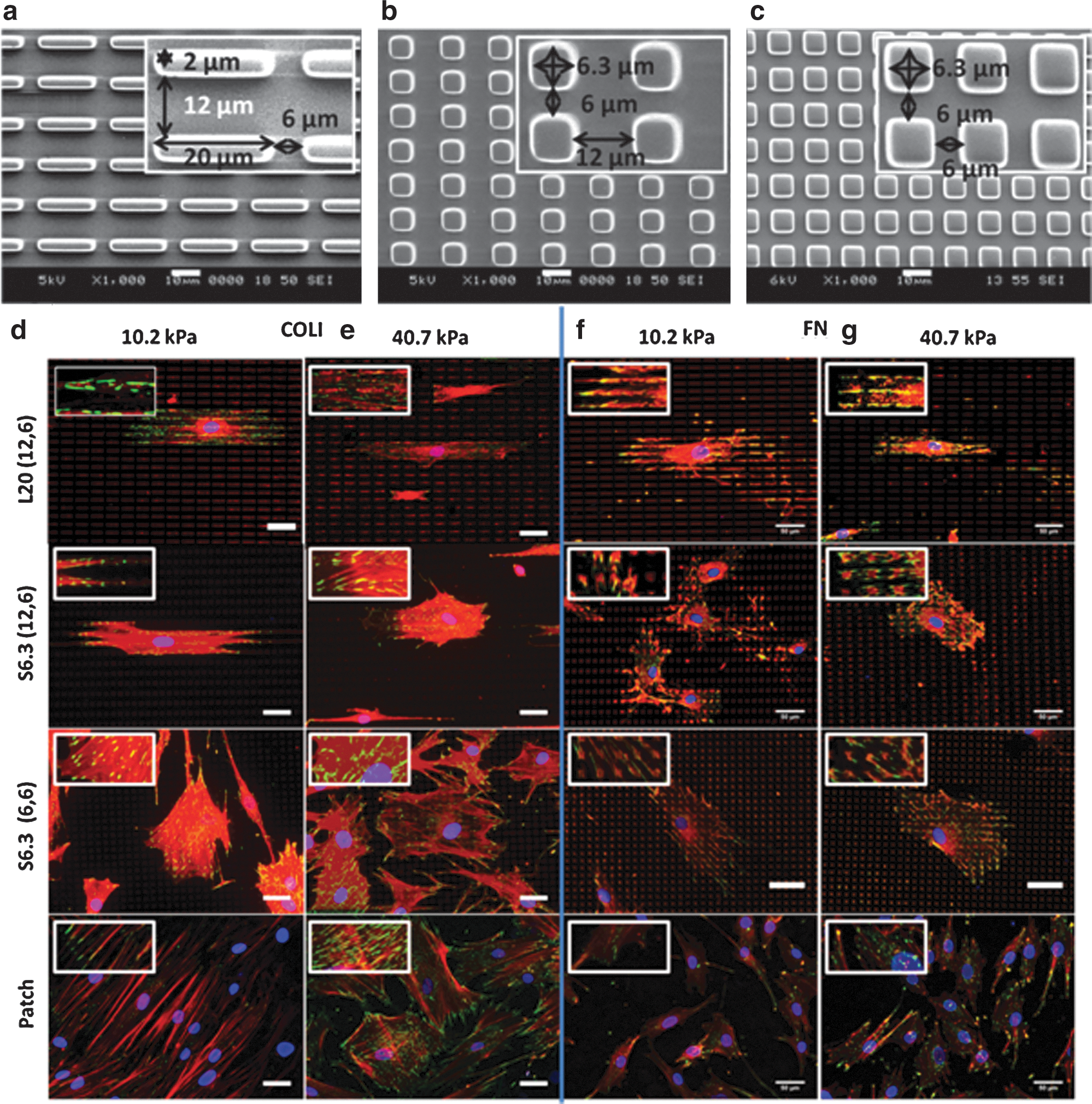
Different micropatterns and their effects on hMSC FA development.
Analysis of vinculin and F-actin expression (Fig. 1d–g) provided insights on how the patterns, stiffness, and ECM ligands affected hMSC FA and cytoskeleton development. In Fig. 1d–g, elongated FAs were successfully modulated via patterning of COLI onto a PA gel with a stiffness of 10.2 kPa [COLI-L20 (12, 6), Fig. 1d] and FN onto a PA gel with a stiffness of 40.7 kPa [FN-L20 (12, 6), Fig. 1g] respectively, by using the pattern L20 (12, 6). The elongated FA was accompanied with elongated cell shape as well. Relatively sparse FA was observed in the L20- (12, 6) patterned groups compared with the patch group (the PA gel surface was patterned with an even homogeneous layer of protein by a flat PDMS stamp) (Fig. 1d–g). As expected, densely distributed, but not elongated, FA was successfully constructed using pattern S6.3 (12, 6) and S6.3 (6, 6). Pattern S6.3 (12, 6) induced densely distributed FA, and hMSCs also have adopted elongated cell shape just as those found in cells patterned with L20 (12, 6), but lower in AR. This suggested that the spacing effects could induce elongated cell shape and align cytoskeleton over the vertical spacing (6 μm) in pattern S6.3 (12, 6), the direction which has a higher density of the ECM ligand. The observation of a similar cell shape, but different FA development, may suggest a difference in lineage commitment. Besides that, the micropatterning effect on FA development in hMSCs was observed to be dependent on the combination effect of substrate stiffness and ECM ligands. Although elongated FA was observed on a PA gel of stiffness 10.2 kPa and surface patterned with COLI-L20 (12, 6), this observation is only present on a PA gel of 40.7 kPa stiffness when FN was used for patterning, that is, FN-L20 (12, 6). However, the FA AR and the length were compromised in the FN-L20 (12, 6) group.
To better quantify the observations mentioned above, a statistical study of FA area, FA AR, FA length, and FA density was performed and is shown in Fig. 2. On the COLI-L20- (12, 6) patterned group, the FA of hMSCs on a PA gel with a stiffness of 10.2 kPa showed 40% increase in FA AR and 60% increase in length (Fig. 2b, c, left). COLI-S6.3 (12, 6) and COLI-S6.3 (6, 6) showed less influence on FA shape, but 20% increase in FA density (Fig. 2d), both on a PA gel with stiffness of 10.2 and 40.7 kPa. On the FN-L20- (12, 6) patterned group, successful elongated FA modulation could only occur on a PA gel with a stiffness of 40.7 kPa. However, it showed a relative lower FA AR and FA length compared with cells on the COLI-L20- (12, 6) patterned group.
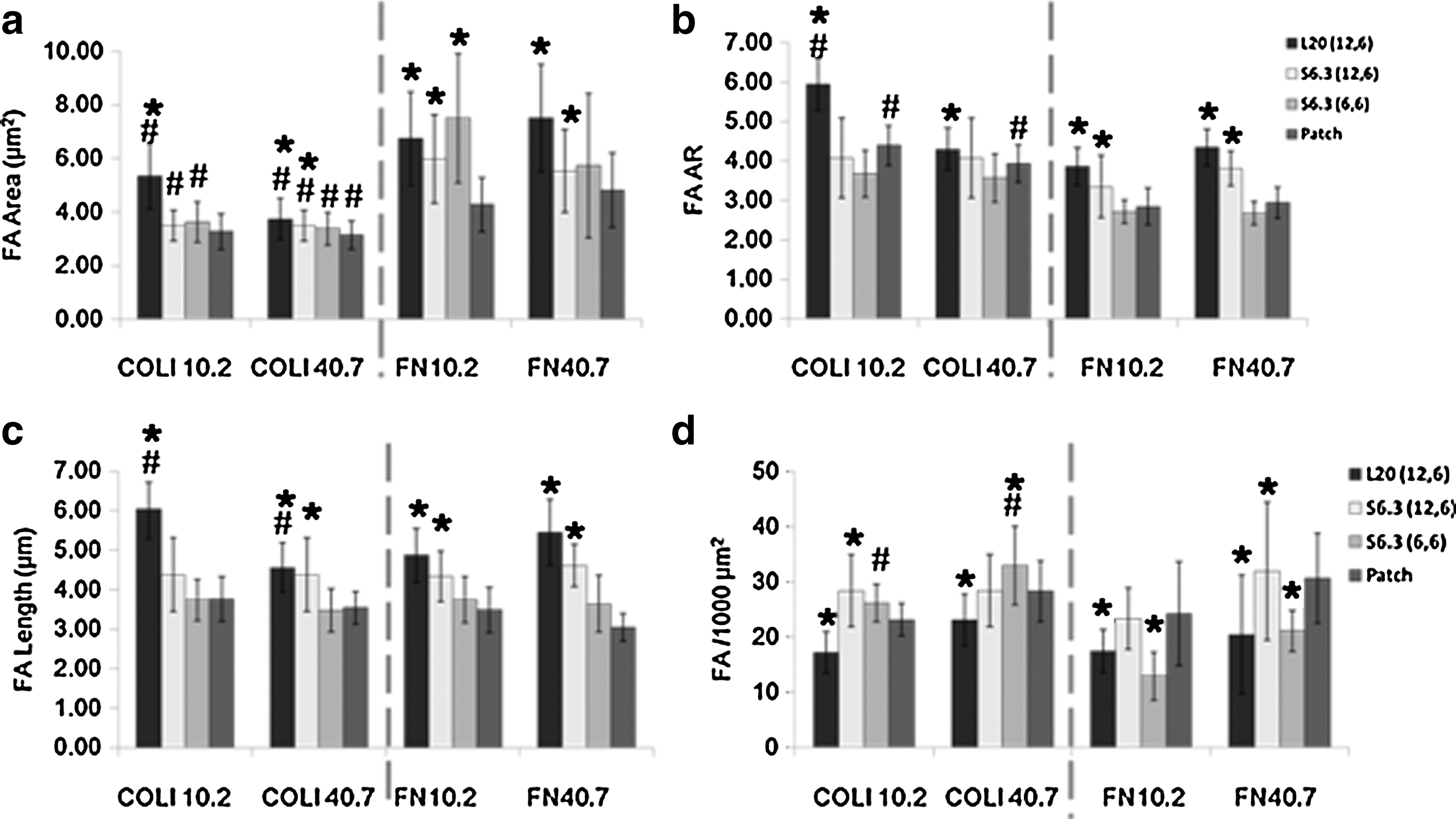
Statistical study of FA development. *P<0.05 between the pattern and patch group with the same kind of pattern protein. # P<0.05 between the COLI and FN group with a PA gel of same stiffness. Sixty cells were analyzed in each group. COLI, type-I collagen; FN, fibronectin.
The observations were in line with our previous study that was performed on PDMS. More importantly, the L20 (12, 6) pattern made significant improvement on the FA AR and length modulation. This is an important indication showing that the FA could be successfully manipulated via micropatterning with optimal matrix compliance and the ligand type.
Elongated FA drove specific myogenic differentiation of hMSCs
In Fig. 3a, the COLI-L20 (12, 6) group showed significant upregulation of myogenic markers MyoD1, GATA4, and MYH7 by 3–4-fold on a PA gel with a stiffness of 10.2 kPa, but not on the PA gel with a stiffness of 40.7 kPa. However, neither S6.3 (12, 6) nor S6.3 (6, 6) could induce specific myogenic differentiation on PA gel, regardless of the substrate stiffness. Immune detection technique was further employed to determine the above upregulated genes at translational level. As shown in Fig. 3c, MHC, which is coded by MYH7, was only significantly expressed in cells cultured on a PA gel with a stiffness of 10.2 kPa patterned with COLI-L20 (12, 6). Besides MHC, immunostaining of α-sarcomeric actin was also carried out, and the result showed a high expression in hMSCs with elongated FA (Supplementary Fig. S3).
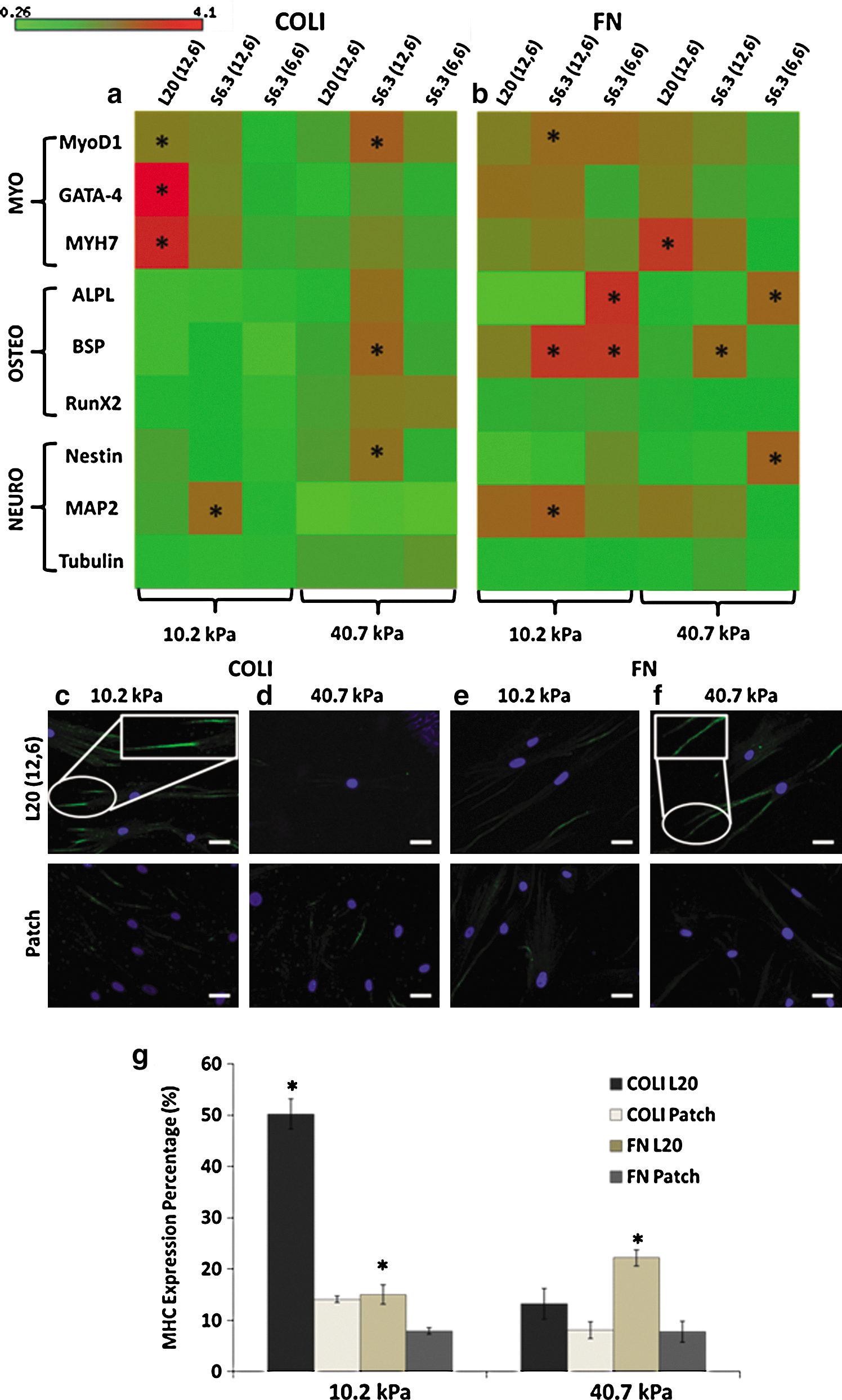
Differentiation characterization showed that elongated FA could significantly induce myogenic differentiation. Real-time PCR result showed L20- (12, 6) specific upregulated myogenic markers on a PA gel with a stiffness of 10.2 kPa in the presence of COLI
When FN was used as the ECM ligand (Fig. 3b), pattern FN-L20 (12, 6) only induced myogenic differentiation on a PA gel with a stiffness of 40.7 kPa, but not as obvious as on a PA gel with a stiffness of 10.2 kPa and in the presence of COLI in terms of expression fold as well as specificity, since upregulation of osteogenic marker BSP and neurogenic marker MAP2 was also observed. In contrast, pattern FN-S6.3 (12, 6) and FN-S6.3 (6, 6), which induced densely distributed FA, could only upregulate osteogenic, but not myogenic markers. Positive MHC staining in Fig. 3f confirmed that the above upregulated myogenic marker could functionalize at protein expression level.
Statistical study in Fig. 3g semiquantified the expression of MHC under different culture conditions. The COLI-L20- (12, 6) patterned PA gel with a stiffness of 10.2 kPa showed the highest percentage of MHC expression (cells with positive MHC staining/total cell number). It was 3.6-times higher than that in the patch group and also much higher than that in other patterns and FN groups. This further supports that the modulation of FA via micropatterning could potentially induce hMSC lineage commitment to specific myogenic differentiation at translation level.
Integrin cluster, RhoA pathway, and MLCK play key roles during myogenic differentiation
A mechanistic study has been performed to decipher the potential biomolecular basis of hMSC differentiation induced through elongated FA, which was constructed under our optimal condition, that is, a COLI-L20- (12, 6) patterned on PA gel with a stiffness of 10.2 kPa. Real-time PCR result highlighted the importance of ITB3 and MLCK through their significant upregulation in expression shown in Fig. 4a. Immunostaining images in Fig. 4c further indicated higher expression of ITB3 in the COLI-L20- (12, 6) patterned group compared with that in the patch control group. Interestingly, ITB3 was recruited to form elongated clusters when hMSCs were cultured under our optimal condition. RhoA and Rac1 were 2 of the most important signaling pathways, which would transmit the extracellular signals into the nucleus and affect hMSC lineage commitment [31]. According to the G-LISA result shown in Fig. 4b, the activation of RhoA in hMSCs on the COLI-L20- (12, 6) patterned PA gel with a stiffness of 10.2 kPa was increased by 80% compared with that in the patch group. However, there was no difference in the activation of Rac1 between them. It implied that RhoA was activated by elongated FA and might be responsible for transmitting the sensed niche signal in our optimal condition into the nucleus.
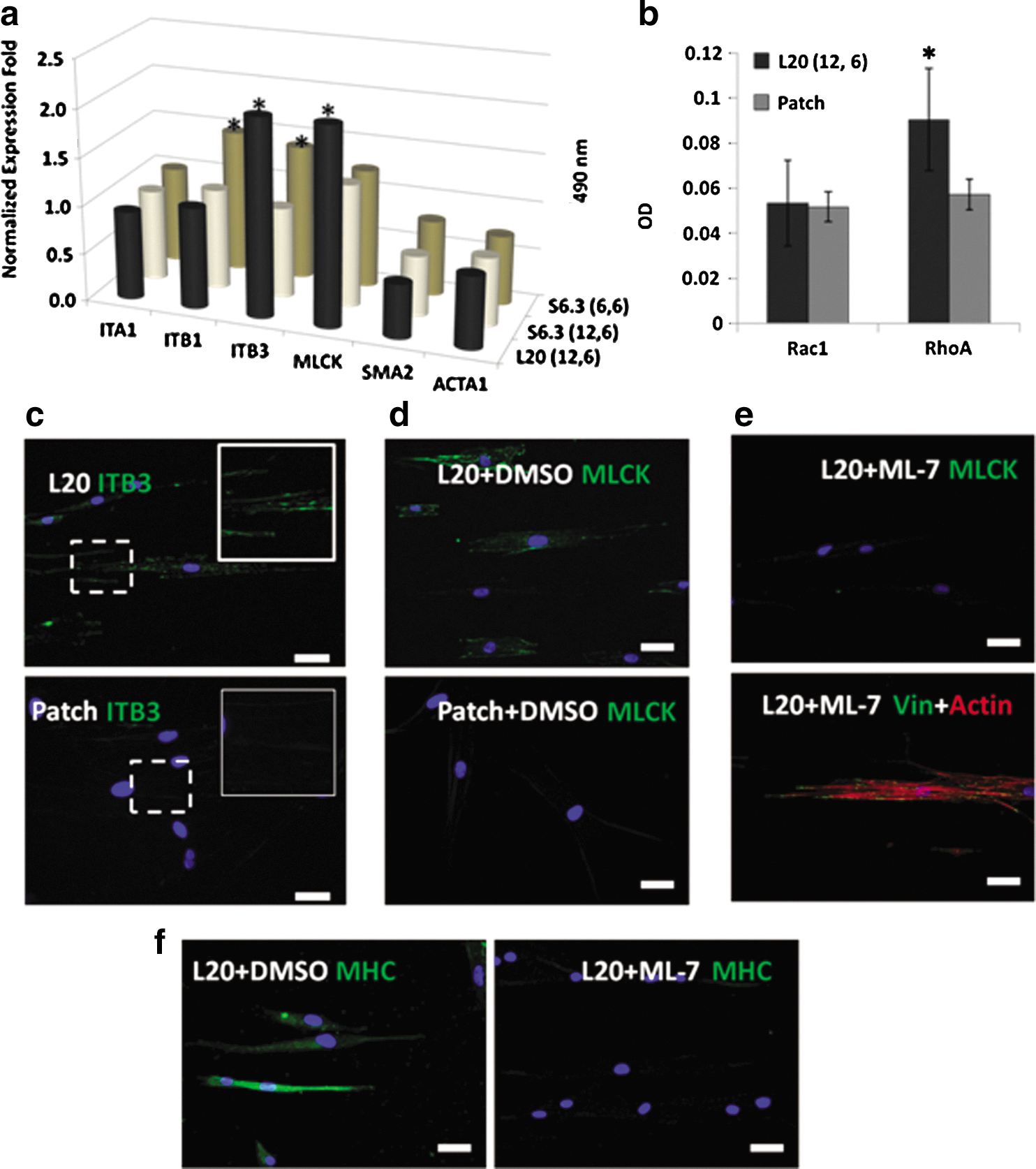
Mechanism study of cells on a 10.2-kPa PA gel showed that elongated FA-induced hMSC myogenesis was related to ITB3 and MLCK recruitment and RhoA pathway activation.
Elongated FA recruited ITB3 clusters and activated the RhoA-signaling pathway, so it was logical to follow-up the response from the cell nucleus after receiving the signal. The most obvious outcomes were the cytoskeleton development, as well as MHC maturation (shown in Figs. 1d and 3b). Under an optimal condition, which significantly showed MHC expression, the F-actin was observed to have formed stress fiber and aligned with the cell longitudinal axis, which was also in the same direction with less-spaced pattern islets. Such kind of less-compact, yet aligned and organized, F-actin formation was typically seen in fully differentiated myotubes [32]. This is in contrast with the F-actin that was randomly distributed when cultured on other patterns.
Herein, the gene expression of MLCK, α-SMA, and α-skeletal muscle actin (ACTA1) was examined. Generally, MLCK is known to phosphorylate the serine-19 residue of myosin regulatory light chain, which would enhance the actomyosin contraction via the RhoA pathway [33,34]. SMA is localized in microfilament bundles, strengthening the contraction of smooth muscle cells, but not in cardiac myocytes [35,36]. ACTA1 is a major constituent of the contractile apparatus, which can be found in skeletal muscle, but poorly expressed in cardiac myocytes [37]. However, as shown in Fig. 4a, only upregulation of MLCK was observed. SMA and ACTA1, which were smooth/skeletal muscle markers for smooth/skeletal muscle differentiation [38], were not upregulated. It corresponded well with the real-time PCR result in Fig. 3a that elongated FA could induce myogenic differentiation, and that the hMSCs have the trend to be differentiated to cardiac muscle like cells.
The FA and cytoskeleton development has a direct relationship to the cellular traction [39]. To examine our proposed idea that the elongated FA affects the cytoskeleton development and arrangement in a tension-dependant manner, we used ML-7 to inhibit MLCK function and reduce cellular tension, while maintaining the central stress fiber structure and FA development (Fig. 4e lower panel) [40]. As a consequence, MLCK expression could be efficiently inhibited by 10 μM of ML-7 compared to the control group (Fig. 4d, e). Similarly, cells cultured with ML-7 showed lower expression of MHC compared to its control group (normal MHC expression as shown in Fig. 4f). From this inhibition study, we have demonstrated that elongated FA could affect cytoskeleton MHC maturation in a cell tension-dependant manner via MLCK.
The role of elongated FA versus elongated cell shape
Many studies have extensively investigated the modulation of cell shape on hMSC differentiation, such as using strip patterns to guide myogenic differentiation of hMSCs [41,42]. It was an efficient way to induce myogenic differentiation of stem cells via manipulating the cell shape. As the cells in the L20 pattern also adopt a generally elongated cellular morphology, there is a need to demonstrate that the differentiation in this study is indeed caused by elongated FA and not cellular morphology. To decouple the 2 effects on hMSC differentiation, we designed 2 micropatterns that could induce a similar elongated cellular morphology, but with different FA development.
Both patterns could promote elongated cell shape as shown in Fig. 5a. The pattern named elongated (upper) was designed to induce elongated FA, whereas the pattern named dense (lower) was designed to induce densely distributed (dense) FA. Statistical analysis of immune-labeled vinculin of hMSCs cultured on a COLI-patterned PA gel with a stiffness of 10.2 kPa has been employed to semiquantify the distribution of FA and cell area. Figure 5b showed 72% increase in the FA length and 51% increase in the FA AR for the pattern elongated compared to the patch group. For the pattern dense, it mainly increased the FA density by 59%, but no elongated FA was observed. A similar cell AR indicated that both patterns have a similar effect on cell shape formation. Hence, cell shape would pose a minimum, if any, effect on hMSC differentiation in this study.
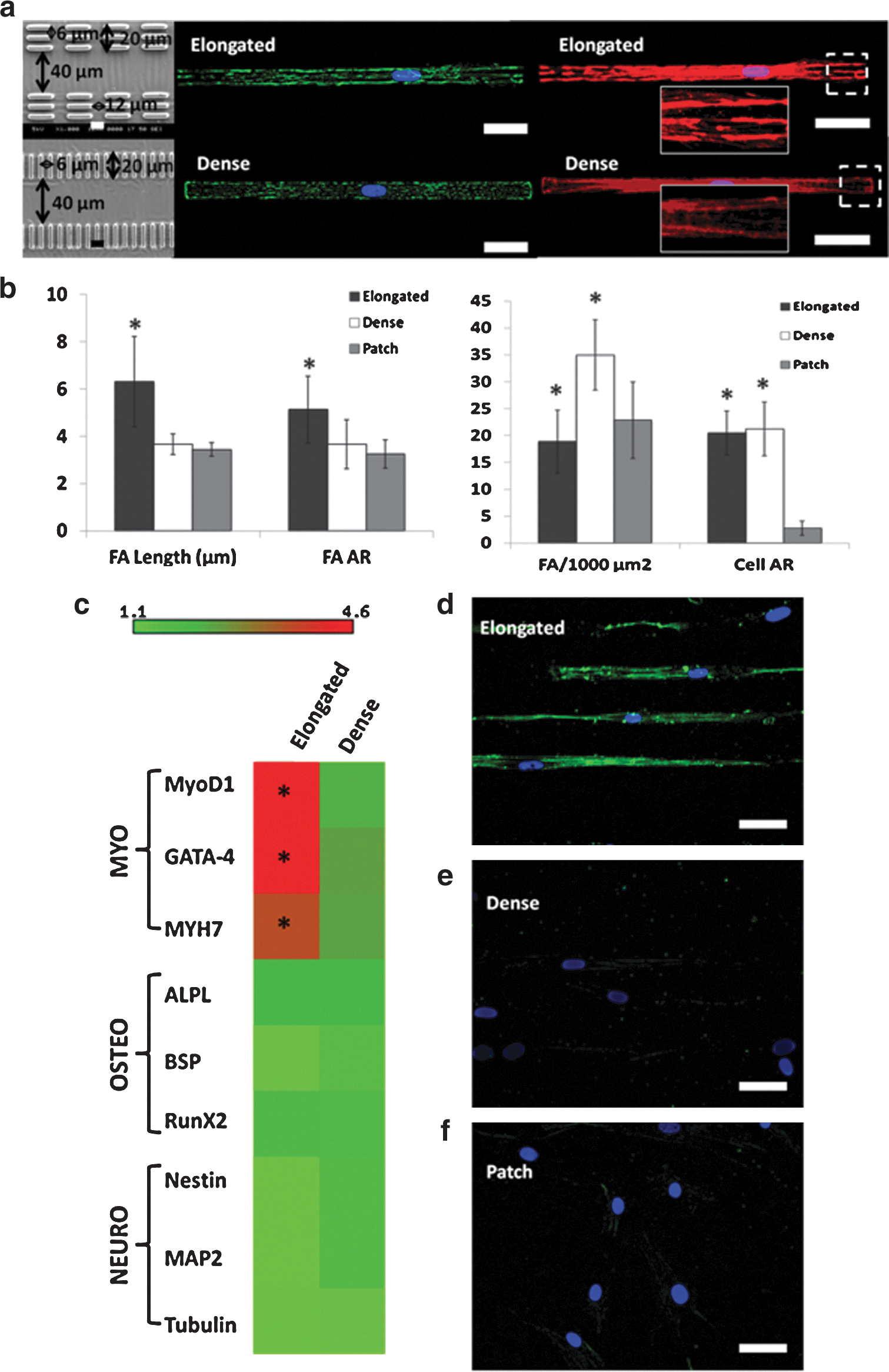
The relationship of FA and cell shape in hMSCs myogenic differentiation.
Interestingly, although hMSCs on both patterns have adopted a similar cell shape, the pattern elongated, which induced elongated FA, showed significant specific upregulation in myogenic markers over the pattern dense (Fig. 5c). This suggested that elongated FA tends to have more effect on induction of specific myogenic differentiation over elongated cell shape. The upregulated expression of MYH7 was confirmed at the translation level by immunostaining of MHC as shown in Fig. 5d. However, no obvious upregulation of MHC was observed in the pattern dense group (Fig. 5e) compared with the patch group (Fig. 5f). The expression of MLCK and the inhibition study of cells in these groups were also performed as shown in Supplementary Fig. S4. Results showed higher expression of MLCK in elongated patterned cells (Supplementary Fig. S4a), and cells with inhibited MLCK did not show expression of MHC (Supplementary Fig. S4c). Therefore, the induction of myogenic differentiation by the elongated pattern was also in a tension-dependent manner.
Discussion
Given their unique regenerative abilities, stem cells offer brilliant potentials for treating diseases such as cardiac failure, muscle damage, and neurological disorders. [43]. However, much work remains to be done at the bench before the real therapy could be realized. One of the critical issues is how to control cell differentiation. In our study, the cellular morphology was not restrained, but the FA development was manipulated using combinations of different patterns, ECM, and matrix stiffness. We have successfully modulated elongated FA via micropatterning using COLI-L20 (12, 6) on a PA gel with a stiffness of 10.2 kPa. However, this micropatterning effect was only observed on a substrate with a stiffness of 40.7 kPa in the presence of FN. A similar phenomenon was observed in a previous study showing a shift of the highest myogenic MyoD1 expression level when changing from COLI-coated PA gel (highest myogenic MyoD1 expression on 10 kPa) to FN-coated PA gel (highest myogenic MyoD1 expression on 25 kPa) [11]. Distinct surface chemistry has a different ability to resist cell traction forces, which would be associated with changes of cell behaviors such as cell attachment, spreading, and differentiation by affecting downstream signaling pathways [44,45]. Through actin stress fiber development, cells exerted higher traction force on a collagen-coated substrate than on an FN-coated substrate [46]. This helps to explain our observation as patterning effects would only be realized with the presentation of optimal stiffness and ECM protein.
The lineage commitment of hMSCs was subjected to an optimized condition [COLI-L20 (12, 6) on a PA gel with a stiffness of 10.2 kPa] corroborated our hypothesis. A major function of FA was to transmit the extracellular signal into the nucleus, resulting in a proper cellular response to the microenvironment. Elongated FA, which was found in mature human myofibroblasts, was able to increase phosphorylation of FA kinase and regulate stress fiber development in myogenic differentiation [47]. Under optimal conditions, recruited FA components such as ITB3 and vinculin formed elongated clusters along the micropatterns as the response to the modulated ECM environment. Thereafter, the elongated FA would bring about the changes in downstream signaling pathways that triggered the myogenic differentiation process [48].
Subsequently, the mechanism study revealed that the RhoA pathway, instead of Rac1, was activated by the elongated FA. Rac1 activation was responsible for the formation of lamellipodia and membrane ruffles and the loss of stress fibers [48]. In contrast, RhoA could promote the formation of integrin clusters, actin polymerization, and myosin activity [49]. More importantly, the RhoA-signaling pathway activated MLCK and nonmuscle myosin to increase the tension in cytoskeleton, which resulted in the myogenic differentiation of hMSCs [31]. The vital role of MLCK in hMSC myogenic differentiation induced through elongated FA has been determined from immunocytochemical analysis and inhibition study. MLCK could trigger MLC phosphorylation to enhance the actin–myosin interaction, which in turn increased the cell tension [50]. Without that, cells may maintain the cytoskeleton structure, but lose the cellular tension [51]. As the result, hMSCs with MLCK inhibited was unable to commit to myogenic differentiation in our study.
Apart from that, we employed another 2 micropatterns to examine the role of FA and cytoskeleton arrangement in hMSC differentiation. Even though these micropatterns could induce hMSCs to adopt a similar cell shape, the one that induced elongated FA showed higher expression in MLCK and superior myogenic differentiation over the other that induced densely distributed FA. This is the first study to decouple the cellular morphology from FA development in affecting hMSC differentiation. We understand that this observation was brought about by the insufficient cell tension generated to induce myogenic differentiation in the dense, but not elongated FA cells; this was confirmed by the MLCK mechanistic study. A latest report showed that elongated and larger FA would induce cell polarization and higher traction force compared with small FA [52]. The formation of small and densely distributed FA might not provide sufficient signals to activate the RhoA pathway, which could further trigger stress fiber formation to generate sufficient traction force, which results in insignificant myogenic differentiation.
Based on our study, we propose a model in Fig. 6a to explain the procedure of elongated FA on induction of myogenic differentiation in hMSCs. First, elongated FA would efficiently generate a sufficient traction force to recruit actin monomer and gather myosin [53]. The RhoA-signaling pathway was then activated and further promoted actin polymerization. During the polymerization, myosin II-B would act as the crosslinker to crosslink actin monomer [54]. Afterward, myosin II-A and MLCK would associate with the actin filament to create contraction along the actin and promote stress fiber formation, and further affect the biophysical cellular response to regulate cell lineage commitment [55]. Previous result showed that cells would build an arc-shape structure over the adherent sites, where there was large tension and there were lots of actomyosin complexes within [55]. In our work, we also observed the arc-like structure and well-developed stress fibers in hMSCs with elongated FA as shown in Fig. 6b. However, only a few of the similar structures and fine actin filaments were observed in other conditions (Fig. 6c, d). Based on a previous study and our observations, these arc-shape structures may be induced by elongated FA, accompanied with the actomyosin assembly and contractions. The data also supported that elongated FA regulates hMSC cytoskeleton development and myogenic differentiation in a tension-dependant manner. The tension is essential in FA-modulated myogenic differentiation; however, it should be in an optimal range. Beyond a certain range, the tension would not trigger necessary signaling pathways such as RhoA, and fails to form stress fiber to guide hMSCs to myogenic differentiation. Finally, with myosin II-A, actin and myosin would form contraction flow and assemble stress fiber as well as align the cytoskeleton. The evidence is given in Fig. 6b–d, showing only cells in Fig. 6b with elongated FA developed bundle-like stress fiber. The model explained the potential mechanism of how elongated FA would induce the cellular tension change and myosin-associated stress fiber formation, both leading to the myogenic differentiation of hMSCs. The interaction study between each factor (substrate stiffness, ECM ligand, and micropatterning in our case) suggested a potentially synergistic effect on cell lineage regulation when certain conditions could be fulfilled. The understanding of these effects working in tandem in inducing stem cell differentiation would contribute significantly to the design and progress of tissue-engineered devices.
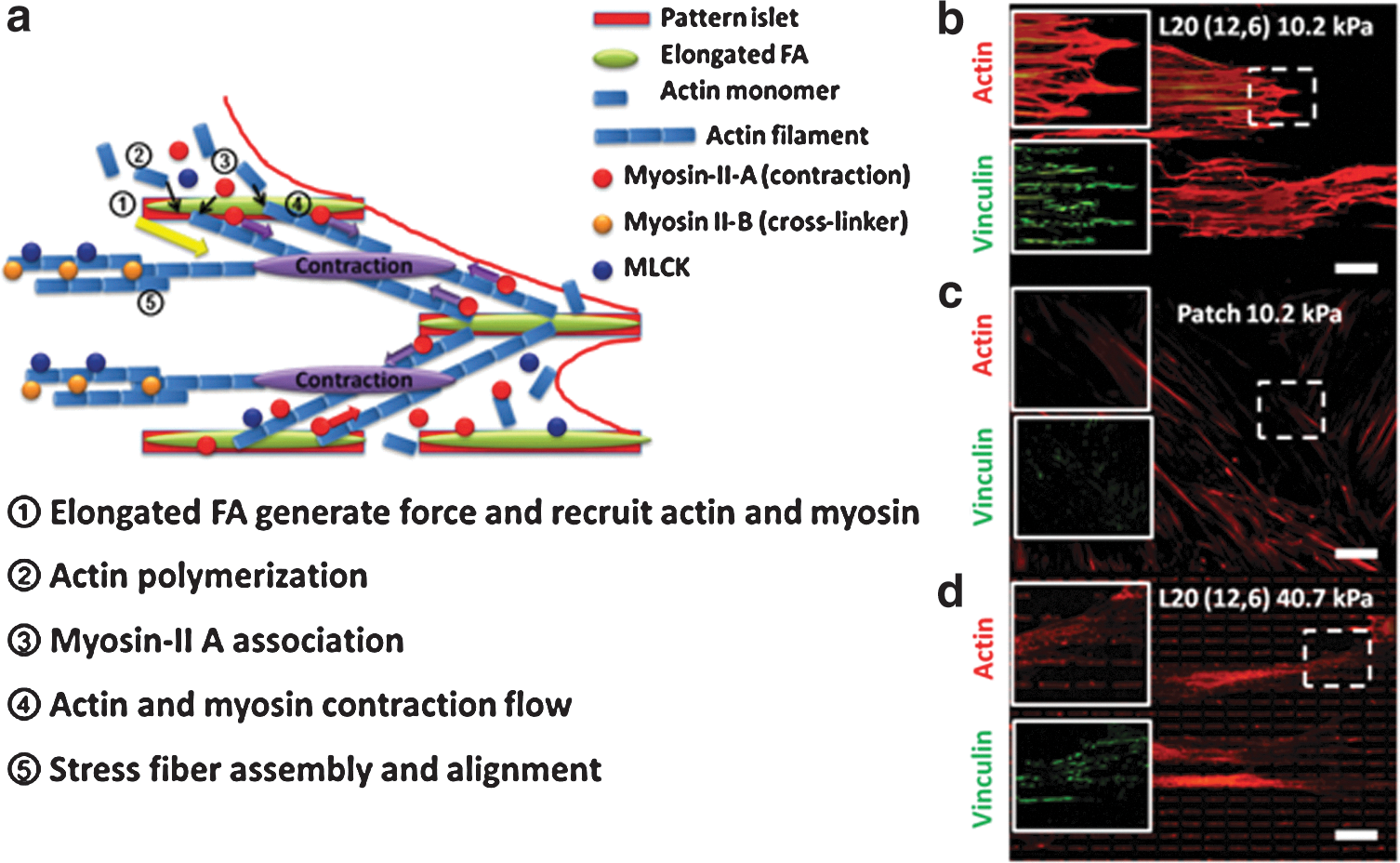
A schematic model showed the mechanism of elongated FA-induced tension-dependent myogenic differentiation.
Conclusions
In this study, a novel bioinspired micropatterning platform to influence specific myogenic differentiation via FA modulation was established. Elongated FA was engineered on a PA gel with a stiffness of 10.2 kPa using COLI as the ink for the L20 (12, 6) pattern. The elongated FA initiated significant recruitment of the ITB3 cluster and induced stress fiber alignment. In addition, a mechanistic study suggested that elongated FA would activate the RhoA-signaling pathway and recruit MLCK, which indicates an enhancement of the cellular tension. Our findings suggest a new and potential platform to manipulate stem cell differentiation via FA modulation. Just as important, through our study, we provided initial understanding of this platform in influencing stem cell lineage commitment. However, more elaborate, mechanistic studies may be required for a comprehensive understanding of the phenomena.
Footnotes
Acknowledgments
We would like to acknowledge the Singapore Stem Cell Consortium (SSCC) (grant no: SSCC/09/017) for financial support.
Author Disclosure Statement
The authors have declared that no competing interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.