
Abstract
Neural stem cells of the subventricular zone (SVZ) represent a potentially important source of surrogate cells for the treatment of brain damage. Proper use of these cells for neuronal replacement depends on the ability to drive neuronal differentiation. Several neuromodulators stimulate neurogenesis. Here we examined the effects of the neuropeptide galanin, on neuronal differentiation in murine SVZ cultures. SVZ neurospheres obtained from early postnatal mice were treated with 10 nM to 2 μM galanin. Galanin promoted neuronal differentiation, increasing numbers of NeuN-, vesicular GABA transporter- and tyrosine hydroxylase-expressing neurons. In contrast, galanin neither affected cell proliferation assessed by BrdU incorporation nor cell death evaluated by terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL). Neuronal differentiation was further confirmed at the functional level by measuring [Ca2+]i variations in single SVZ cells after KCl and histamine stimulations to distinguish neurons from immature cells. Galanin treatment increased the numbers of neuronal-like responding cells compared to immature cells. Using selective agonists (M617, AR-M1896) and antagonists (galantide, M871) for galanin receptors 1 and 2, we showed that both galanin receptors mediated neuronal differentiation. Early proneuronal effects of galanin included positive regulation of the transcription factor neurogenin-1 (Ngn1). In addition, galanin promoted axonogenesis and dendritogenesis, increasing both the length of phosphorylated stress-activated protein kinase- and Tau-positive axons and the numbers of microtubule associated protein-2 (MAP-2)-labelled dendrites. Moreover, galanin inhibited SVZ cell migration in the transwell assay. Our results show a proneurogenic effect of galanin and open new perspectives for future applications in stem cell-based therapies for neuronal replacement.
Introduction
Materials and Methods
All experiments were performed in accordance with the European Community (2010/63/EU) guidelines for the care and use of laboratory animals.
SVZ cell cultures
SVZ cells were prepared from 1–3-days-old C57BL/6 mice either wild type (WT) or expressing the green fluorescent protein (GFP) under the control of the actin gene promoter, as previously described [29] (see Supplementary Data Supplementary Data are available online at
[125I]Galanin receptor autoradiography and galanin-stimulated [35S]GTPγS functional binding
The procedures were previously described [31 –33] and are detailed in Supplementary Data.
Self-renewal assay
SVZ cells were seeded as single cells, at 2,500 cells per well in 24-well cell culture plates in SFM containing 5 ng/mL epidermal growth factor (EGF) and 2.5 ng/mL fibroblast growth factor 2 (FGF-2) (low EGF/FGF-2) and supplemented or not (control) with 1 μM galanin. Four wells per condition were assayed. After 6 days, the numbers of primary neurospheres were quantified. Primary neurospheres were then collected by centrifugation, dissociated into single cell suspensions (Neurocult dissociation kit) and seeded in low EGF/FGF-2 medium as aforementioned. After 6 days, the numbers of secondary neurospheres were counted [34] (Supplementary Fig. S1G).
Cell proliferation studies
Proliferation was assessed using the ELISA BrdU colorimetric assay (Roche; #11 647 229 001). SVZ cells (40,000 per well) obtained from dissociated primary neurospheres (Neurocult dissociation kit), were plated in 96-well culture plates (4 wells per condition) and treated in the absence (control) or the presence of 1 μM galanin in SFM devoid of growth factors or supplemented with 5 ng/mL EGF and 2.5 ng/mL FGF-2. Galanin (10 nM, 100 nM and 1 μM, Bachem AG; #H-7450) and GALR1 and GALR2 agonists (GALR1 agonist: M617; 100 nM, #2697; GALR2 agonist: AR-M1896; 100 nM, #2699, both from Tocris) were applied for 48 h. Cell proliferation was determined by adding 10 μM BrdU for the last 4 h of the culture session. Amount of BrdU incorporation was evaluated by densitometry (at 450 nm) after the use of a peroxidase conjugated anti-BrdU antibody and reaction with a peroxidase substrate, according to the manufacturer's instructions. The absorbance measured in control (ie, SFM-treated wells) was set at 100% and values in experimental conditions were expressed as percentages of control values.
Determination of cell apoptosis by TUNEL
Primary SVZ neurospheres were plated for 48 h on poly-D-lysine coated coverslips. Apoptosis was evaluated by the TUNEL as previously described [29] (Supplementary Fig. S1A).
Single cell calcium imaging studies
To investigate the effect of galanin on neuronal differentiation and to identify the receptor(s) mediating this effect, SVZ neurospheres were seeded on poly-D-lysine-coated coverslips. Cultures were then treated with the following drugs: galanin (10 nM, 100 nM, 1 μM), GALR1 (M617) and GALR2 (AR-M1896) agonists both at 10 and 100 nM, 1 μM galanin in combination with 100 nM of the GALR2 antagonist (M871; #2698; Tocris), 100 nM of the GALR1 agonist in combination with 100 nM of the GALR2 antagonist, 1 μM galanin with 100 nM or 1 μM of the nonselective GALR antagonist galanin (1–13)-substance P (5–11) amide (galantide, #H-1312; Bachem), and allowed to develop for 7 days at 37°C. At the end of the culture session, SCCI experiments were performed to identify functional neurons by means of their increase of free intracellular calcium levels ([Ca2+]i) specifically in response to 50 mM KCl but not in response to 100 μM histamine [29,35] (see Supplementary Data and Supplementary Fig. S1F).
Immunostaining
Cells were fixed in 4% paraformaldehyde (PFA) for 15 min at room temperature (RT). BrdU staining (10 μM; Sigma-Aldrich) was performed as previously described [29] using the Alexa fluor 594 mouse anti-BrdU (1:100; #A21304, Invitrogen). For immunocytochemistry, SVZ cells were permeabilized and nonspecific binding sites blocked using a mixture of 1% Triton X-100 and 3% bovine serum albumin (BSA) in phosphate-buffered saline (PBS) for 30 min at RT. SVZ cells were subsequently incubated overnight at 4°C with the appropriate primary antibodies as listed in Supplementary Table S1 and prepared in PBS containing 0.1% Triton X-100 and 0.3% BSA. Coverslips were rinsed in PBS and incubated for 1 h at RT with the appropriate Alexa Fluor 488 and 594 secondary antibodies produced in donkey (all from Invitrogen). SVZ cell nuclei were stained with Hoechst 33342 (2 μg/mL in PBS) for 5 min at RT and mounted in Dako fluorescent medium (Dako). For immunodetection of GALR1, GALR2, and GALR3 on brain slices, adult mice were deeply anaesthetized and transcardially perfused with 0.9% NaCl followed by 4% PFA. Brains were removed and postfixed in 4% PFA at 4°C overnight. Coronal brains sections 40 μm-thick were cut using a Leica vibratome and then placed into anti-freeze solution (ethylene glycol 30% vol/vol, glycerol 30% vol/vol, PB 0.1 M 40% vol/vol) and conserved at −20°C. Immunohistochemistry was performed as aforementioned. Fluorescent images were recorded using a confocal microscope (LSM 510 Meta; Carl Zeiss) or a fluorescent microscope (Axioskop 2 Plus; Carl Zeiss).
Migration in Boyden chambers
Cell migration was assessed using the transwell assay in which cells migrate through a microporous membrane in response to a specific molecule. The underside of the transwell membrane was coated with 10 μg/mL fibronectin 4 h before the addition of cells. The culture medium, supplemented with 2 μM galanin or 2 μM neuropeptide Y (NPY, as the positive control) or the vehicle (containing BSA, control condition), was added to each of the wells in 24-well plates. We used 2 μM galanin because a similar NPY concentration was previously shown to induce SVZ cell migration [34]. SVZ cells (6,000 cells) were added in the upper chamber of the transwell. After 6 h incubation, the cells that were present on the underside of the transwell were fixed in 4% PFA, labeled with propidium iodide (1:100; Invitrogen), and counted on 20 microscope fields (taken with a 40× objective using a Zeiss microscope) by an observer blinded to the treatments. The results are expressed as percent of the control condition set to 100%. Data were obtained from three independent experiments in which each condition was assessed in three independent wells [34].
Isolation of total RNA, cDNA synthesis and qPCR analysis: galanin and GALRs
The analysis was performed as previously described [29,31] (see Supplementary Data).
Isolation of total RNA, cDNA synthesis and qPCR analysis: Mash1, Dlx2, Ngn1 transcription factors
Total RNA was isolated from SVZ neurospheres treated without (control) or with 1 μM galanin for 2, 3, and 4 days, according to illustra RNAspin Mini RNA Isolation Kit manufacturer's instructions (GE Healthcare Life Sciences) as detailed in the Supplementary Data section.
Quantitative chromatin immunoprecipitation
The protocol used was recently described [36] (see Supplementary Data).
Western blot
Detection of GALR1, GALR2 and GALR3 was performed by western blot in SVZ neurospheres and hippocampus tissue samples from newborn mice (see Supplementary Data.
Statistical analysis
For cell proliferation studies, each experimental condition was reproduced in four different wells of a 96-well plate and experiments were assayed four times. Background values obtained in the negative control wells were subtracted from all values. Within each experiment, the mean optical density evaluated at 450 nm in control wells were set to 100% and optical densities of experimental conditions are expressed as percent of the control condition. In all experiments involving the plating of neurospheres, measurements were performed at the border of SVZ neurospheres where migrating cells form pseudo-monolayers of cells. For SCCI experiments, the percentage of neuronal-like responding cells (with a histamine/KCl ratio below 0.8) was calculated on the basis of one microscopic field per coverslip, containing ∼100 cells (magnification, 40×). Except where otherwise specified, the experiments were replicated at least thrice. Three coverslips were analyzed for each condition in each experiment. The percent of NeuN, TUNEL, BrdU/NeuN, βIII tubulin/BrdU, Dlx2/BrdU, βIII tubulin/TH, VGAT positive cells in SVZ cell cultures was calculated from cell counts in five independent microscopic fields in each coverslip with a 40× objective (∼200 cells per field). Quantification of the number and the total length of newly formed neurites, positive for p-JNK and emerging from the neurospheres, was performed in two independent cultures (two coverslips for each condition) in 20 nonoverlapping fields per coverslip using digital images (magnification, 20×). The numbers of GFP/MAP-2+ dendrites per neuron, total GFP/MAP-2+ dendrite length and mean length per GFP/MAP-2+ dendrite were evaluated in three culture preparations in ∼20 nonoverlapping fields per coverslip (40× magnification). Software used was Axiovision, release 4.6 (Carl Zeiss). qChIP and qPCR experiments were performed in at least three independent cultures. Data are expressed as mean±standard error of the mean (SEM). Statistical significance was determined by using the unpaired two-tailed Student's t-test or one-way analysis of variance followed by Bonferroni or Dunnett's multiple comparison test, with P<0.05 representing statistical significance.
Results
Expression of galanin and GALRs as well as galanin binding in the SVZ
Using qPCR, the expression of galanin and GALRs was investigated in the SVZ both in vitro, in neurosphere cultures, and in vivo in tissue samples relative to that of the reference gene GAPDH. In SVZ neurospheres, the galanin gene was found not to be expressed during the 40 cycles of qPCR (Fig. 1A), but GALR1, GALR2 and GALR3 were all expressed (ΔCtGALR1: 16.00±0.56; ΔCtGALR2, 14.04±0.25; ΔCtGALR3, 18.32±0.67; Fig. 1A). Similarly, in dissected SVZ tissue samples, all three GALRs were expressed (ΔCtGALR1: 12.12±0.22; ΔCtGALR2: 16.39±0.49; ΔCtGALR3: 16.46±0.52; Fig. 1B). To identify the possible source of galanin for SVZ cells in vivo, we assessed the expression of galanin in the SVZ, the adjacent striatum and choroid plexus that produces the cerebrospinal fluid in contact with SVZ cells. We found that galanin mRNA is expressed in all studied tissues (SVZ: ΔCtgalanin=16.25±1.00; striatum: ΔCtgalanin=14.93±1.06; choroid plexus: ΔCtgalanin=15.50±0.91; Fig. 1C). Hippocampus tissue samples were previously found to express galanin, and therefore, used as a positive control for the specificity of the galanin primers [7] (ΔCtgalanin: 13.65±0.61; Fig. 1C). Expression of GALR1, GALR2, and GALR3 receptors in SVZ cultures from newborn mice was also confirmed by western blots. Hippocampus tissue samples were used as positive controls (Fig. 1D).
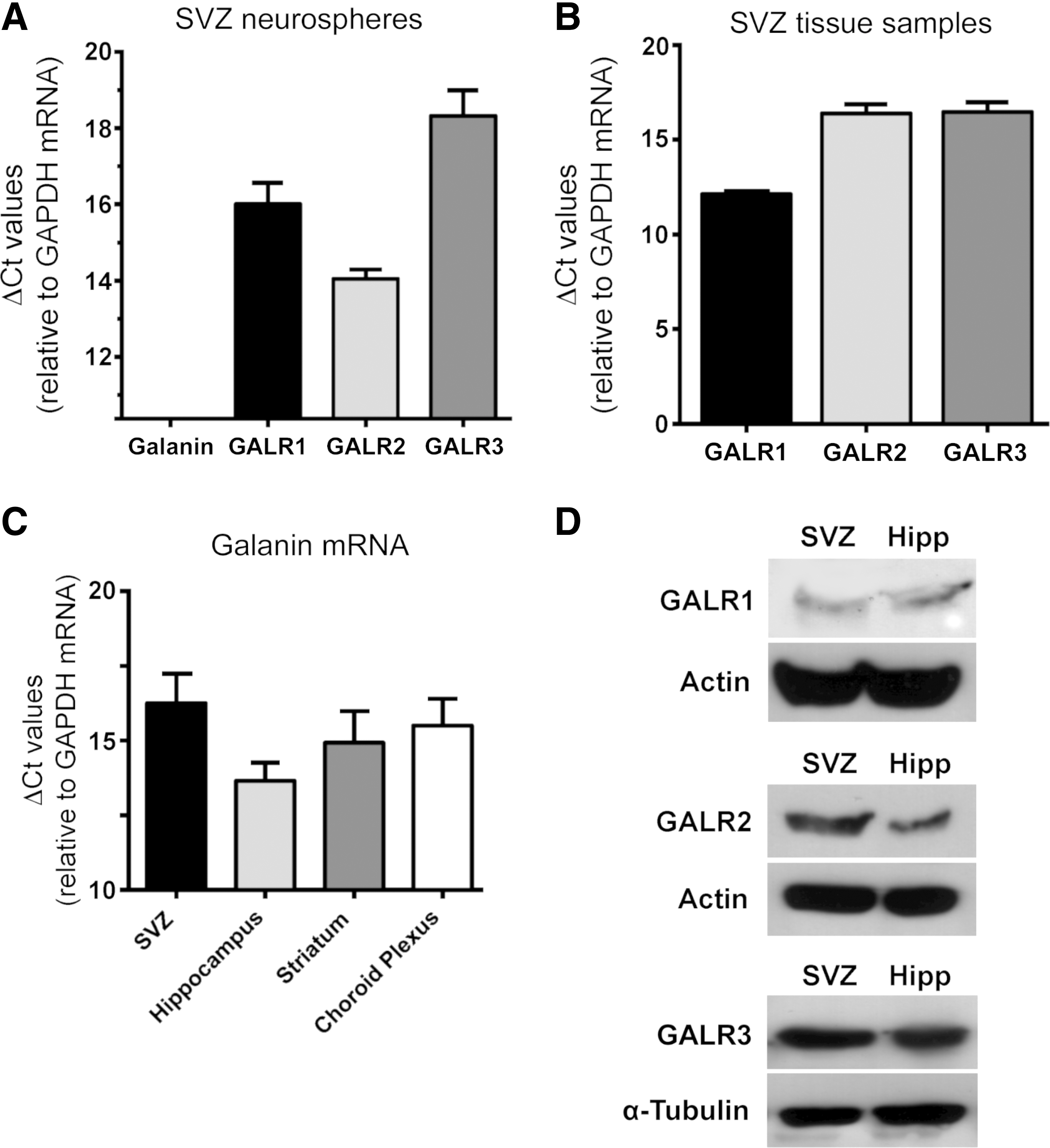
Detection of galanin and GALRs in subventricular zone (SVZ)-derived neurospheres, adult SVZ and brain tissues using quantitative polymerase chain reaction (qPCR) and western blotting.
Consistent with the expression of GALRs in the SVZ, specific [125I]-galanin binding was found corresponding to the SVZ of adult mouse brain slices (Fig. 2A–F). Moreover, low levels of galanin-stimulated [35S]GTPγS functional binding were also detected corresponding to the SVZ in the same animals, indicating that GALRs are functional in the SVZ (basal binding: 100.00%±2.52%; galanin-stimulated binding: 107.30%±0.49%; data not shown). To further confirm that GALRs are expressed in the SVZ in vivo, brain slices from adult mice encompassing the ependymal layer, SVZ, and striatum were stained for nestin, a marker of ependymal and stem/progenitor cells [37], and for GALR1, GALR2 and GALR3. Colocalization of GALRs was found with nestin-positive cells both in the ependymal layer and the SVZ (Fig. 2G–I). Faint labeling for the GALRs was also found in the striatum. These data correlate with the experiment shown in Fig. 2A–F. Similarly, expression of GALRs was found in nestin-expressing cells in SVZ neurospheres (Supplementary Fig. S2).
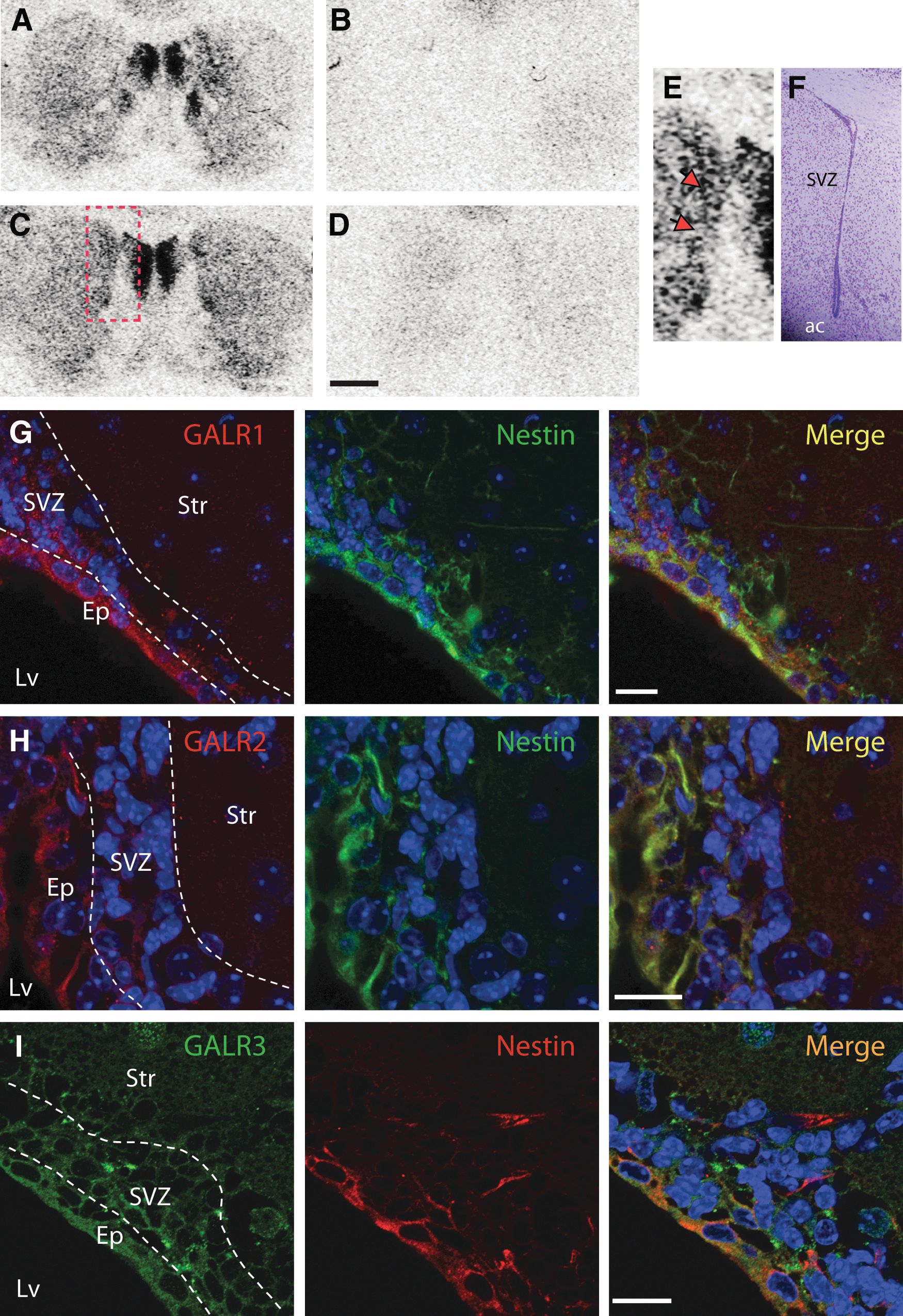
[125I]-galanin binding and immunodetection of GALR1, GALR2 and GALR3 in the murine SVZ. Total [125I]-galanin binding
Galanin does not affect self-renewal, proliferation or cell death
We determined whether galanin impacts on self-renewal capacity of SVZ cells by using the neurosphere assay as described previously [34,36,38]. The experiments were conducted in parallel with the hepatocyte growth factor (HGF, 50 ng/mL) known to induce self-renewal [38]. Exposure of freshly dissected SVZ cells to 1 μM galanin for 6 days did not affect the numbers of primary neurospheres generated as compared to nontreated cultures (control) (control: 100%±2.26%, 1 μM galanin: 92.03%±4.97%, HGF: 143.60%±13.17%, P<0.001, data not shown). Moreover, galanin had no impact on the self-renewal capacity of stem cells, since the numbers of secondary neurospheres were similar in control and treated conditions (control: 99.99%±2.65%, 1 μM galanin: 106.00%±4.90%, HGF: 140.20%±11.41%, P<0.05, data not shown). The effect of galanin on SVZ proliferation was tested in SVZ cultures using the BrdU incorporation assay. Cells were treated for 48 h with 10 nM, 100 nM, or 1 μM galanin, and with 100 nM of either the GALR1 (M617) or GALR2 (AR-M1896) agonists. BrdU was added to the cultures for the last 4 h of exposure to neuropeptides. As depicted in Table 1, none of the treatments affected proliferation, while incubation with EGF and FGF-2, used as a positive control, increased BrdU incorporation.
Table depicts the values of absorbance measured at 450 nm after revelation of the peroxidase activity associated with an anti-BrdU antibody. SVZ monolayers were treated for 48 h with galanin and GALR agonists. As a positive control, SVZ cultures were incubated with EGF and FGF-2 and, as expected, proliferation was significantly increased in these cultures. Data represent means±SEM of four independent cultures. a P<0.01, using one-way analysis of variance with Dunnett's post hoc test for comparison with the control condition.
EGF, epidermal growth factor; FGF-2, fibroblast growth factor-2; SEM, standard error of the mean; SVZ, subventricular zone.
To rule out the possibility that galanin may exert an effect on proliferation of stimulated cells, the assay was conducted in the presence growth factors (EGF and FGF-2). Again, no effect of galanin was obtained, while EGF and FGF-2 were highly efficacious (control: 100.00%±4.06%; 1 μM galanin: 99.42%±9.01%; EGF+FGF-2: 163.10%±22.87%, **P<0.001, five cultures tested, data not shown). The effects of galanin and GALR agonists on cell death were evaluated by TUNEL staining to detect apoptotic nuclei. Neither galanin nor the GALR agonists affected SVZ cell death as the numbers of apoptotic nuclei did not differ from control levels (control: 19.09%±1.50%, galanin: 17.68%±1.98%, 100 nM M617: 17.89%±1.32%, AR-M1896: 14.31%±1.65%, data not shown). Taken together, these results show that galanin neither affects cell proliferation nor cell death in the SVZ.
Galanin promotes neuronal differentiation through GALR1 and GALR2 activation
The effects of galanin on neuronal differentiation were first tested in neurospheres plated on poly-D-lysine-coated coverslips and incubated for 7 days in the absence (control) or the presence of 1 μM galanin, and then stained to reveal βIII tubulin-positive neurons and GFAP-expressing astrocytes. As depicted in Fig. 3A, the neuronal network appeared denser in galanin-treated cultures as compared to the untreated cultures. To quantify neuronal differentiation, cells were immunostained for the neuronal nuclear marker NeuN, NeuN-positive (NeuN+) nuclei were counted (Fig. 3B, C). The result shows that galanin promotes neuronal differentiation as more NeuN+ cells were counted in the presence of galanin as compared to control (control: 11.79%±1.61%, galanin: 28.49%±4.83%, P<0.01; Fig. 3C).
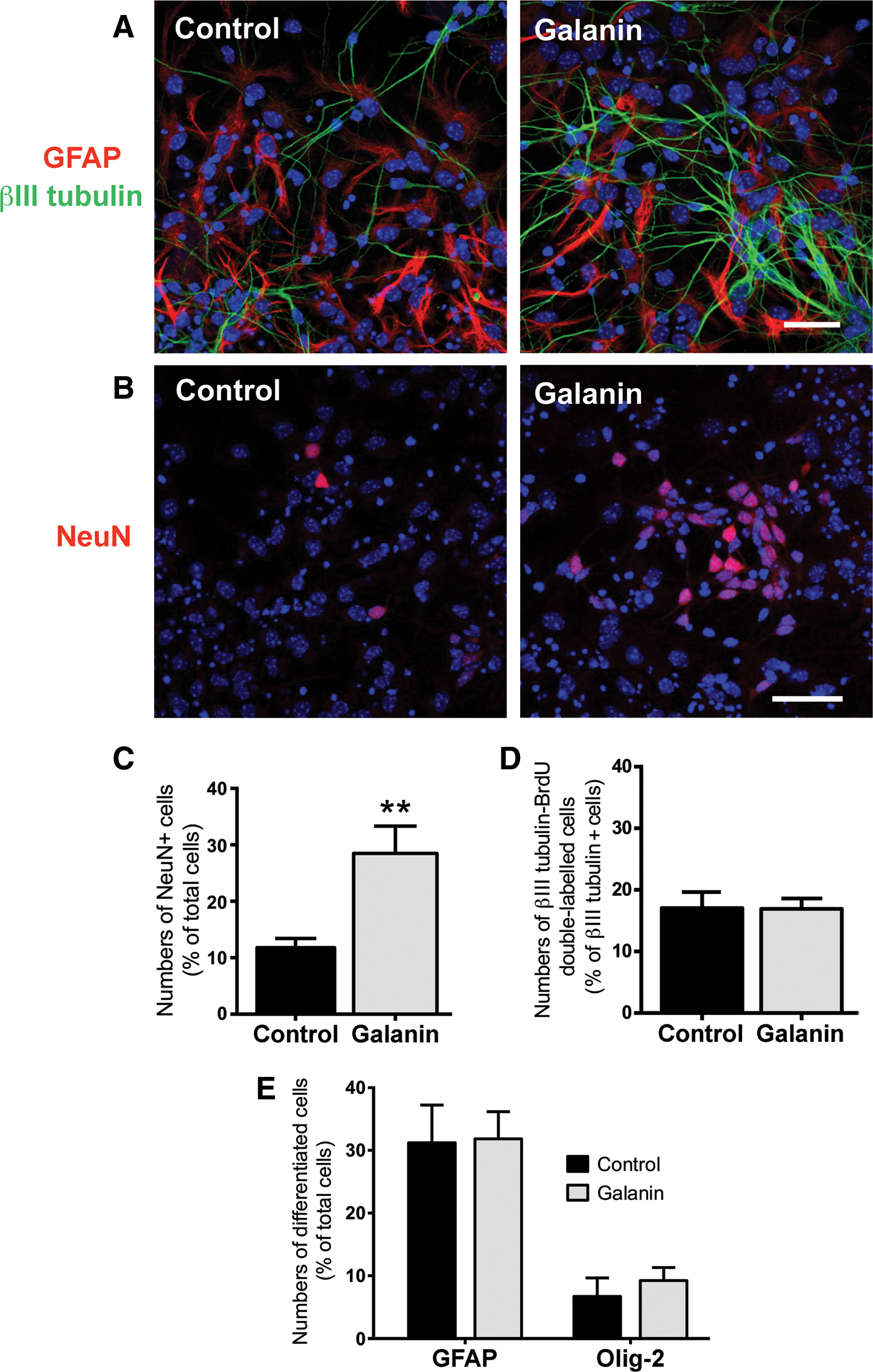
Galanin stimulates neuronal differentiation. Representative digital images of SVZ cells treated or untreated (control) for 7 days with 1 μM galanin and immunostained for glial fibrillary acidic protein (GFAP, red), βIII tubulin (green)
As an increase in the number of neurons can result from an increase in the proliferation rate of neuronal progenitors (or neuroblasts), we incubated the cells with galanin for 48 h and marked the DNA of proliferating cells with BrdU for the last 4 h of the cultures. Proliferating neuroblasts were labeled for BrdU and βIII tubulin. We counted the numbers of double-labeled cells and found that galanin does not induce the proliferation of neuroblasts (control: 17.06%±2.53%, galanin: 16.91%±1.72%; Fig. 3D). To discard the possibility that galanin may enhance the proliferation of more immature progenitors, we performed the same experiments and labeled the cells for the C cell type marker Dlx2 and counted the number of BrdU-Dlx2 double-labeled cells (expressed as a percentage of proliferating cells). No significant difference was obtained between the conditions (control: 33.85%±1.57%, galanin: 37.73%±3.26%; three independent cultures, data not shown), indicating that galanin also does not affect proliferation of immature progenitors.
The effects of galanin on glial cell differentiation were assessed by counting the numbers of GFAP+ and Olig-2+ cells in SVZ cultures incubated for 7 days in the absence (control) or the presence of 1 μM galanin. No differences in the number of astrocytes and oligodendrocytes were observed between the conditions (GFAP, control: 31.21%±6.01%, galanin: 31.84%±4.34%; Olig-2, control: 6.71%±2.97%, galanin: 9.24%±2.08%; Fig. 3E). Moreover, quantification of GFAP expression was also performed by western blotting and we found no effect of galanin treatment (control set at 100%, 1 μM galanin: 101.60%±8.00%, three independent cultures, data not shown), indicating that galanin does not affect glial differentiation.
Functional neuronal differentiation was evaluated by measuring [Ca2+]i variations in single cells after KCl and histamine stimulation. KCl depolarization leads to the massive entry of Ca2+ into neurons through voltage-dependent calcium channels, while stimulation with histamine increases [Ca2+]i in SVZ immature cells but not in neurons or in glial cells. We previously showed that SVZ-derived neuronal-like mature cells display histamine/KCl ratios below 0.8 [29,35,39]. Figure 4A–D represents characteristic profiles of fluorescence records displayed by 20 cells of control (A), 100 nM galanin-treated cells (B), 1 μM galanin, and 100 nM M871-treated cells (C) and 1 μM galanin and 1 μM galantide-treated cells (D). In control cultures, most of the cells responded to histamine but not to KCl. After 7 days of treatment with 100 nM or 1 μM galanin, the percent of neuronal-like cells increased, suggesting a proneurogenic effect of galanin (control: 11.80%±1.65%; 10 nM galanin: 25.72%±2.90%; 100 nM galanin: 45.67%±7.45%, P<0.01; 1 μM galanin: 44.98%±5.98%, P<0.01; Fig. 4E). This proneurogenic effect of galanin was abolished by cotreatment with the nonselective GALR antagonist galantide, while galantide by itself did not affect neuronal differentiation (1 μM galanin+100 nM galantide: 10.74%±5.52%, data not shown; 1 μM galanin+1 μM galantide: 15.50%±5.51%; 1 μM galantide: 6.36%±2.15%; Fig. 4D, E).
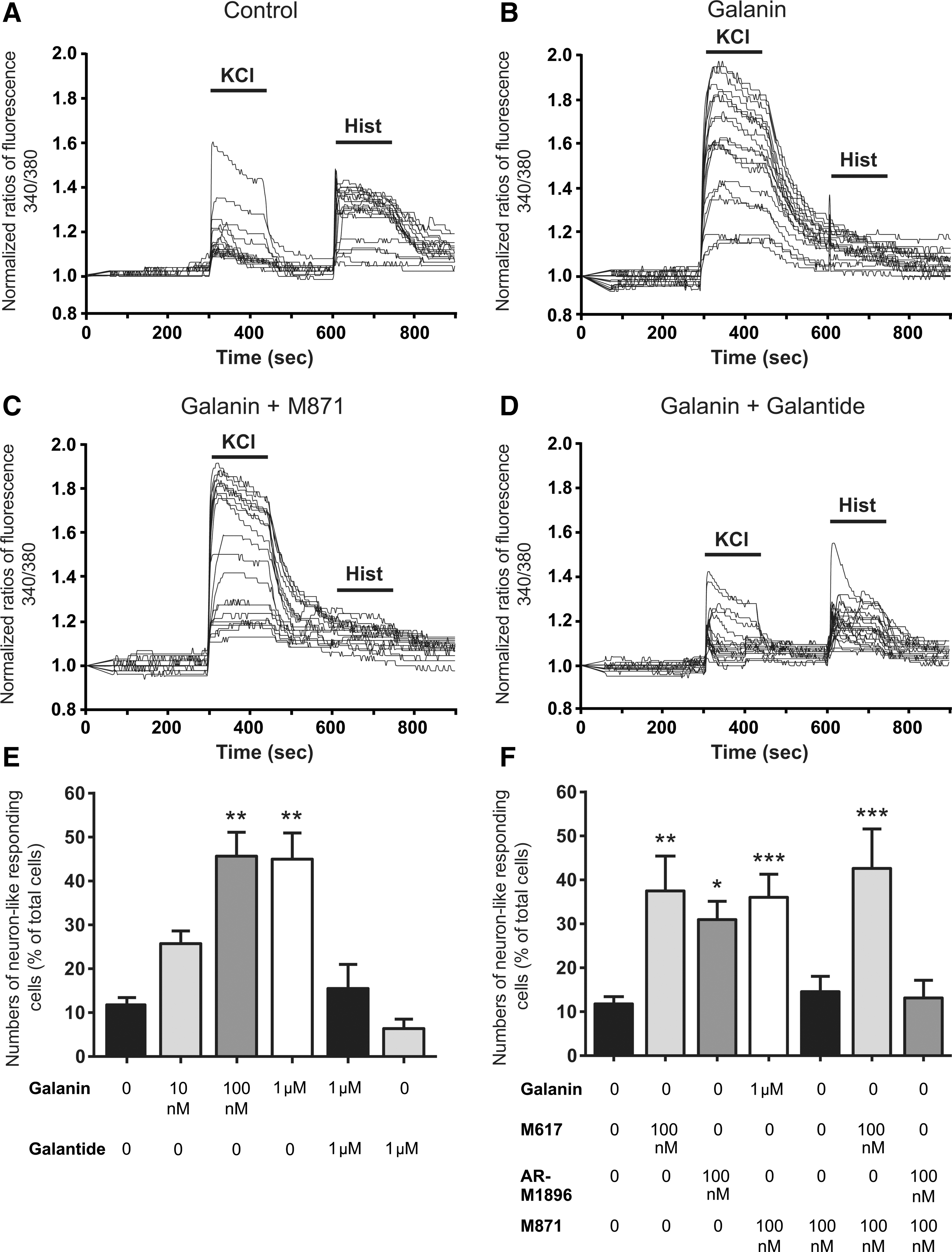
Galanin induces neuronal differentiation in SVZ cultures through GALR1 and GALR2.
To determine which receptor mediates the proneurogenic effect of galanin, SVZ cells were treated for 7 days with 10 and 100 nM of either the GALR1 agonist M617 or the GARLR2 agonist AR-M1896. Single-cell calcium experiments showed that the proneurogenic effect of galanin is reproduced after incubation with 10 and 100 nM of the GALR1 agonist, suggesting that galanin effects are mediated by GALR1. However, treatment with 100 nM but not 10 nM GALR2 agonist also increased neuronal differentiation (10 nM M617: 32.47%±8.39%, P<0.05, data not shown; 100 nM M617: 37.49%±7.95%, P<0.01; 10 nM AR-M1896: 15.31%±6.85%, data not shown; 100 nM AR-M1896: 30.95%±4.20%, P<0.05; Fig. 4F). As the IC50 for the GALR2 agonist AR-M1896 on GALR1s has been reported to be 879 nM [40] and as AR-M1896, in our experiments, triggered neuronal differentiation when added at 100 nM, the proneurogenic effects obtained after GALR2 stimulation seems to be specific.
To confirm that the proneurogenic effect obtained with the GALR1 agonist M617 was exclusively due to GALR1 stimulation and that AR-M1896 was selective for GALR2, SVZ cells were treated with the GALR2 antagonist M871. M871 alone at 100 nM had no effect on neuronal differentiation (14.58%±3.49%). Antagonizing the GALR2 receptor with M871 did not prevent the proneurogenic effects exerted by M617 and galanin (100 nM M871+100 nM M617: 42.61%±8.98%, P<0.01; 100 nM M871+1 μM galanin: 36.04%±5.26%, P<0.01; Fig. 4C, F). In contrast, the antagonist M871 was able to prevent the proneurogenic effect exerted by AR-M1896 (100 nM M871+100 nM AR-M1896: 13.14%±4.02%, Fig. 4F), indicating that GALR2 is also involved in the neurogenic effects of galanin. These results indicate that both GALR1 and GALR2 mediate the proneurogenic effect of galanin. However, GALR1 and GALR2 agonists had no synergistic effects (data not shown).
Galanin interferes with early stages of neuronal differentiation
To determine whether galanin affects early stages of neuronal differentiation, proliferating SVZ cells were labeled with BrdU for the first 24 h of treatment followed by a chase of 6 days in the absence (control) or the presence of galanin (Fig. 5A). Galanin treatment caused a significant increase in the proportion of NeuN- and BrdU-positive cells among the NeuN-positive cells (control: 4.64%±0.85%, 1 μM galanin: 9.05%±1.57%, P<0.05, Fig. 5A). To determine whether galanin impacts on proneuronal gene expression, levels of mRNA for the proneuronal transcription factors Mash1, Dlx2, and Ngn1 were measured in cultures treated for 2, 3, and 4 days with galanin. Galanin treatment significantly increased Ngn1 mRNA levels at 3 days (control set to 1, Mash1: 1.00±0.28, Dlx2: 0.72±0.38; Ngn1: 6.48±1.83, P<0.001, Fig. 5B) but not at 2 and 4 days (data not shown), while no effect was found on Mash1 and Dlx2 mRNA levels. Similarly, expression of the proglial transcription factor Olig-2 was not affected (3 days, Olig-2: 1.27±0.61).
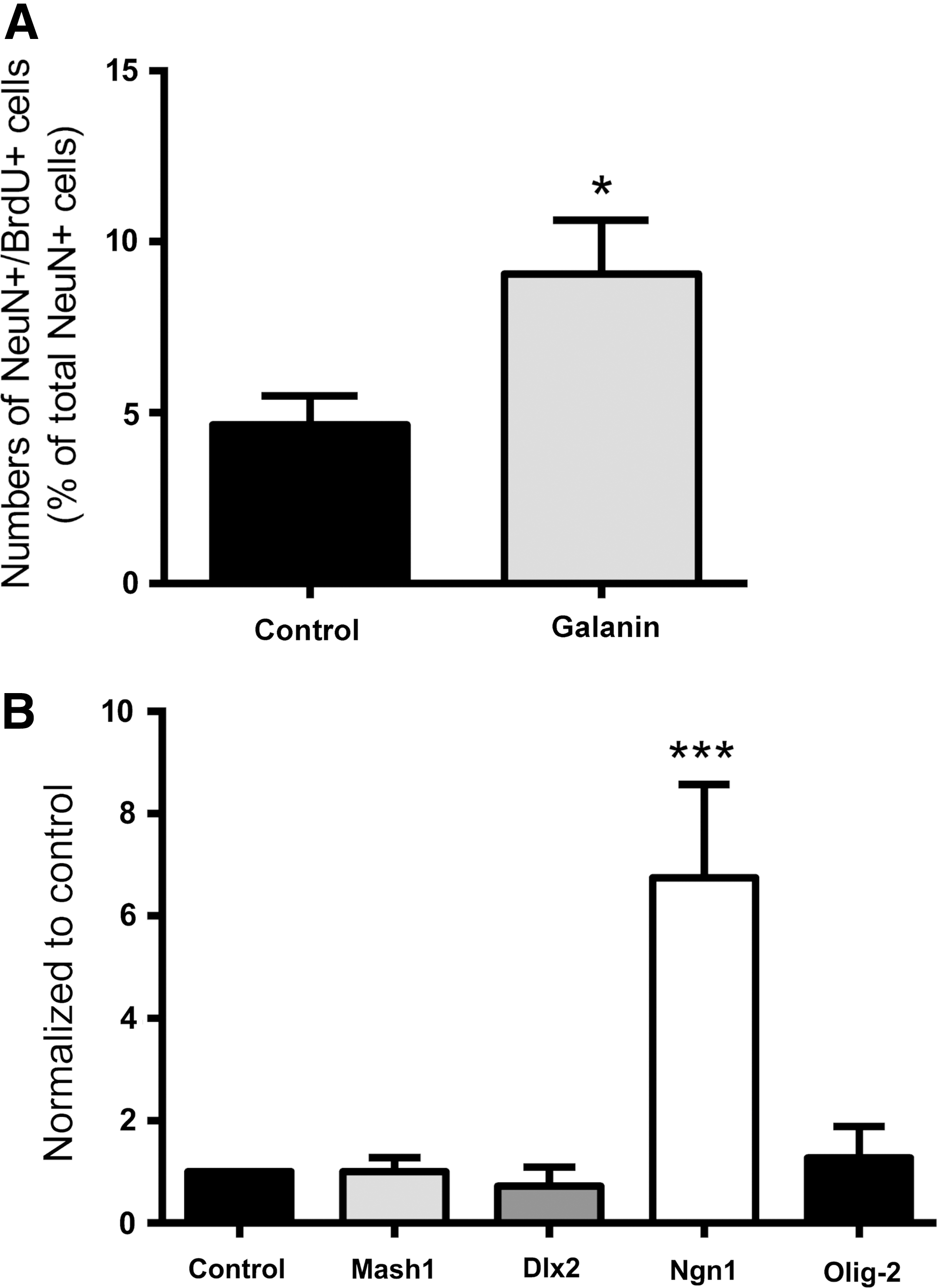
Galanin induces neuronal commitment of SVZ cells.
Moreover, galanin elicited neuronal differentiation via chromatin modification of the Ngn1 promoter region. qChIP analysis which determines the interaction between specific proteins and DNA (eg, promoter regions) was used to study if galanin treatment promoted the availability of the selected proneurogenic transcription factor Ngn1 to transcription complexes. In particular, we used qChIP against the H3K36m3, a protranscriptional histone modification known to induce gene expression. Six and 24 h treatment with galanin increased H3K36m3 in the promoter region of Ngn1 as assessed (control set to 1, 6 h: 3.75±2.63, 24 h: 2.10±0.82, three to six independent cultures, data not shown). Together these results demonstrate that galanin triggers neuronal commitment.
Galanin promotes both GABAergic and dopaminergic differentiation
SVZ-derived neurons have been shown to develop into GABAergic interneurons with a minority of cells differentiating into dopaminergic interneurons in the olfactory bulb [41]. As galanin triggered neuronal differentiation in SVZ cells, we sought to determine whether galanin would increase these former neuronal populations. SVZ cells were treated for 7 days in the absence (control) or the presence of 1 μM galanin, and immunocytochemistry was performed to detect GABAergic neurons (expressing VGAT) and dopaminergic neurons (expressing the enzyme TH). Since glial expression of TH has been reported [42], double immunolabelling for TH and βIII tubulin was performed and TH positive neurons counted. Galanin increased both the numbers of GABAergic neurons (VGAT, control: 10.55%±1.06%, 1 μM galanin: 16.90%±2.38%, P<0.05, Fig. 6A, B) and dopaminergic neurons (TH+neurons, control: 8.03%±1.13%, 1 μM galanin: 13.58%±1.92%, P<0.05; Fig. 6C, D).
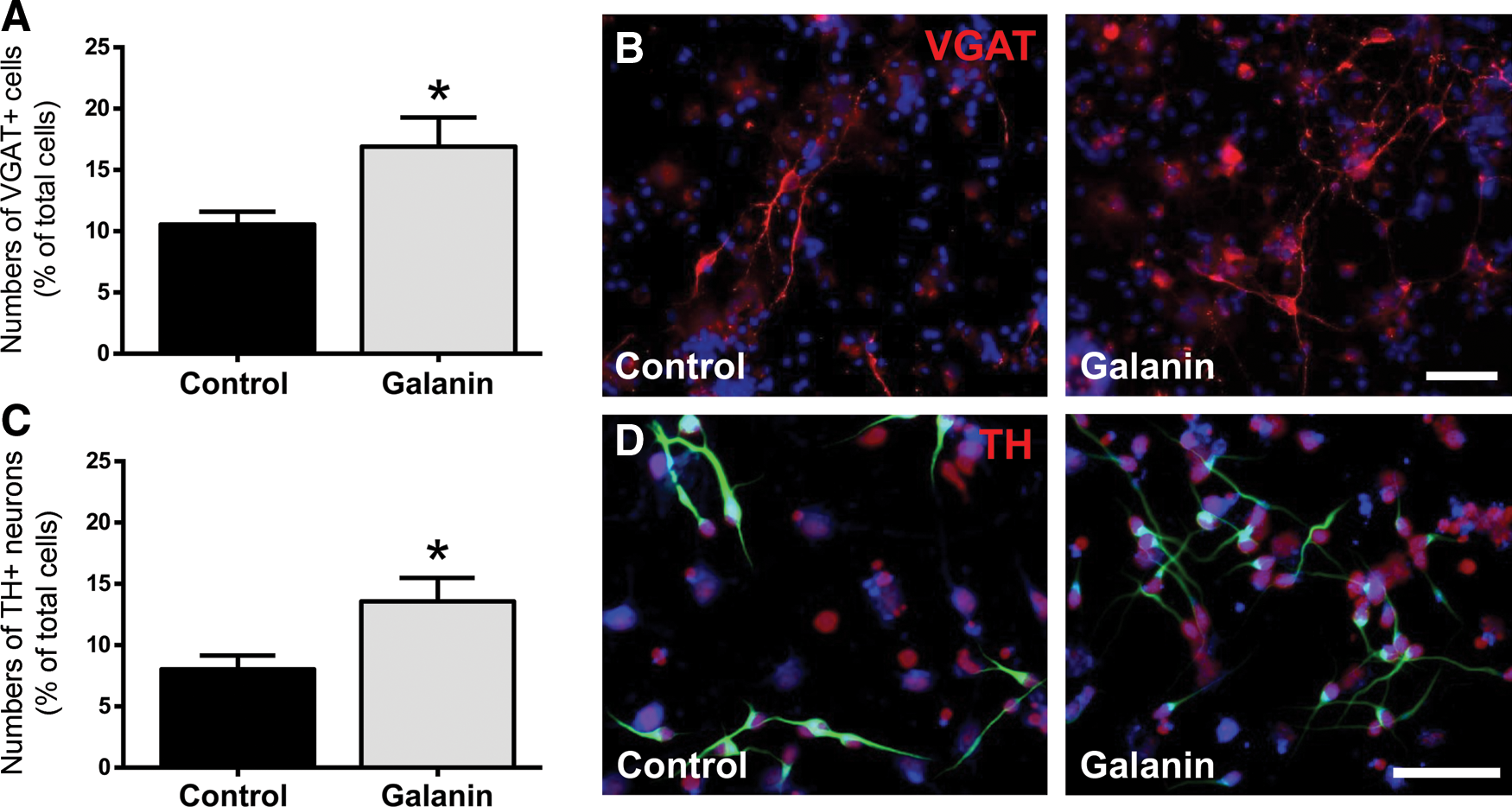
Galanin promotes GABAergic and dopaminergic neuronal differentiation. Bar graph depicts the numbers of either
Galanin promotes neuronal maturation
To examine whether galanin affects axonal formation, activation of the SAPK/JNK MAPK pathway, known to interfere with axonal formation, was investigated in branches derived from plated SVZ neurospheres incubated for 6 h with 1 μM galanin. The numbers and total length of branches which were immunoreactive for the phospho(p)JNK in SVZ cultures were measured. We ascertained that pJNK+ (green) branches colocalized with the axon-specific protein Tau (red) (Fig. 7A; see [43]). Exposure of SVZ cells to galanin significantly increased the number as well as the total length of pJNK+ branches per neurosphere as compared with control cultures (total length of ramification per neurosphere: control: 134.80±26.19 μm; 1 μM galanin: 280.80±38.27 μm, P<0.01, Fig. 7B; numbers of branches per neurosphere, control: 2.00±0.38; 1 μM galanin: 3.32±0.37 P<0.05; Fig. 7C). The same experiments were performed in the presence of 20 μM SP600125, an inhibitor of JNK activity. Length and number of pJNK/Tau+branches per neurosphere decreased in cultures treated with SP600125 alone or together with 1 μM galanin (total length of ramification per neurosphere, control+SP600125: 53.57±10.69 μm; 1 μM galanin+SP00125: 33.23±9.98 μm, P<0.001 as compared to galanin, Fig. 7B; numbers of branches per neurosphere, control+SP600125: 0.62±0.16; 1 μM galanin+SP00125, 0.44±0.11; P<0.001 as compared to galanin, Fig. 7C), attesting the crucial role of SAPK/JNK in mediating axonal elongation. Altogether, these data show that galanin promoted axonogenesis via activation of the SAPK/JNK pathway in SVZ cultures.
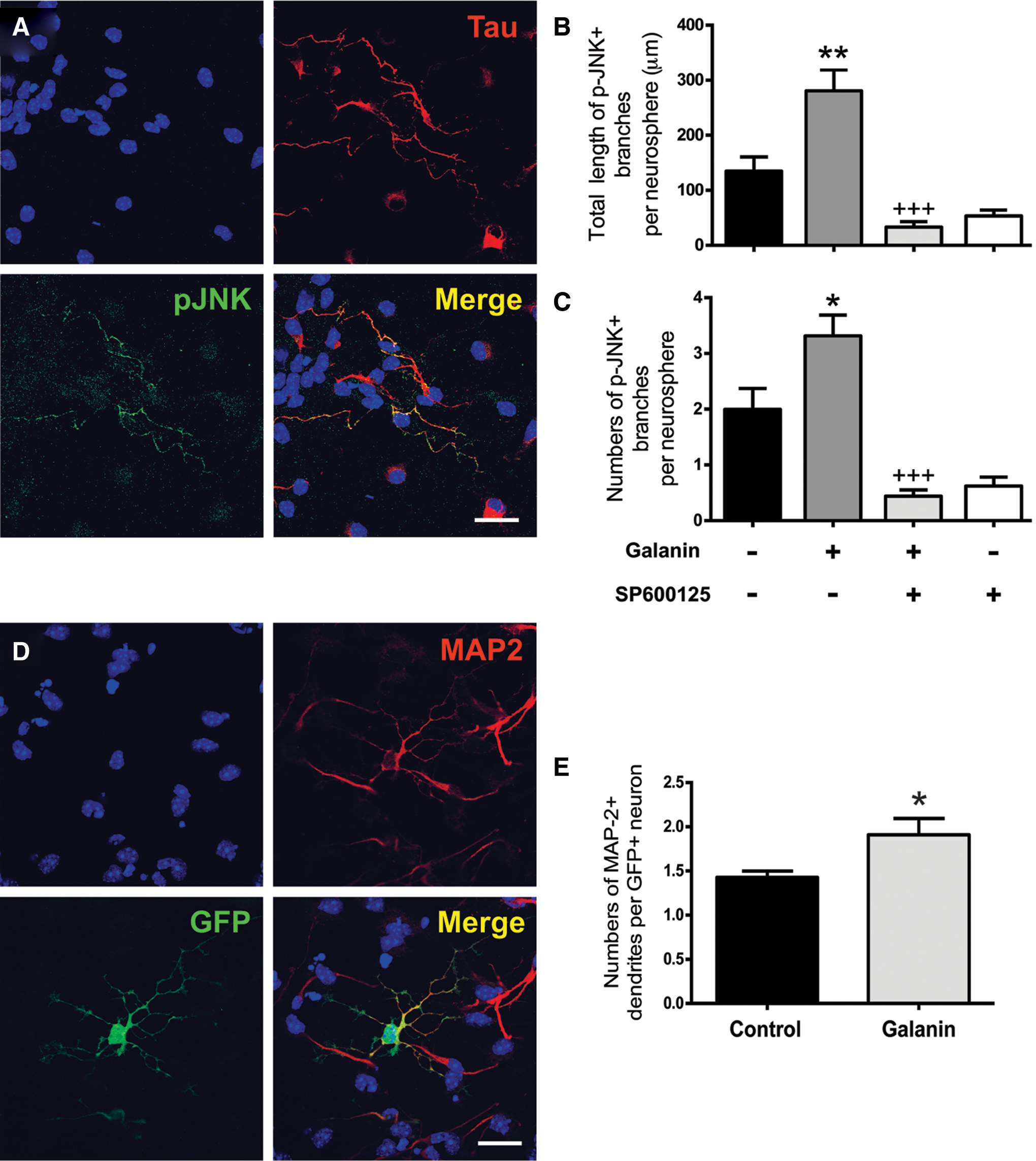
Galanin promotes neuronal maturation.
The effects of galanin on neuritogenesis were also evaluated in WT and GFP mixed SVZ cell cultures. Neurites were immunostained for MAP-2 and numbers of GFP/MAP-2+ neurites per neuron, total GFP/MAP-2 length of neurites and mean length per GFP/MAP-2+ neurite were evaluated. Figure 7D provides a representative aspect of GFP+ neurons labeled for MAP-2. Galanin increased the numbers of dendrites per neuron (control: 1.43±0.07; 1 μM galanin: 1.91±0.18; P<0.05; Fig. 7E), while not affecting the other parameters significantly (total length: control: 659.00±64.75 μm; 1 μM galanin: 808.60±92.11 μm; mean length per neurite: control: 32.83±2.01 μm; 1 μM galanin: 35.81±1.50 μm, data not shown).
Galanin reduces SVZ cell migration
As galanin increased neuronal differentiation, we tested whether the neuropeptide would also exert an effect on the migration pattern of SVZ-derived neuroblasts using Boyden chambers. The cells were plated in the upper compartment of a transwell filter and allowed to migrate for 6 h through a microporous membrane separating the upper and lower compartments. Galanin was added in the lower compartment and found to decrease the number of migrating cells (control: 100.02%±2.68%; galanin: 76.75%±3.87%; P<0.05; Fig. 8). NPY was chosen as a positive control since its promigratory effects had been previously demonstrated [34].
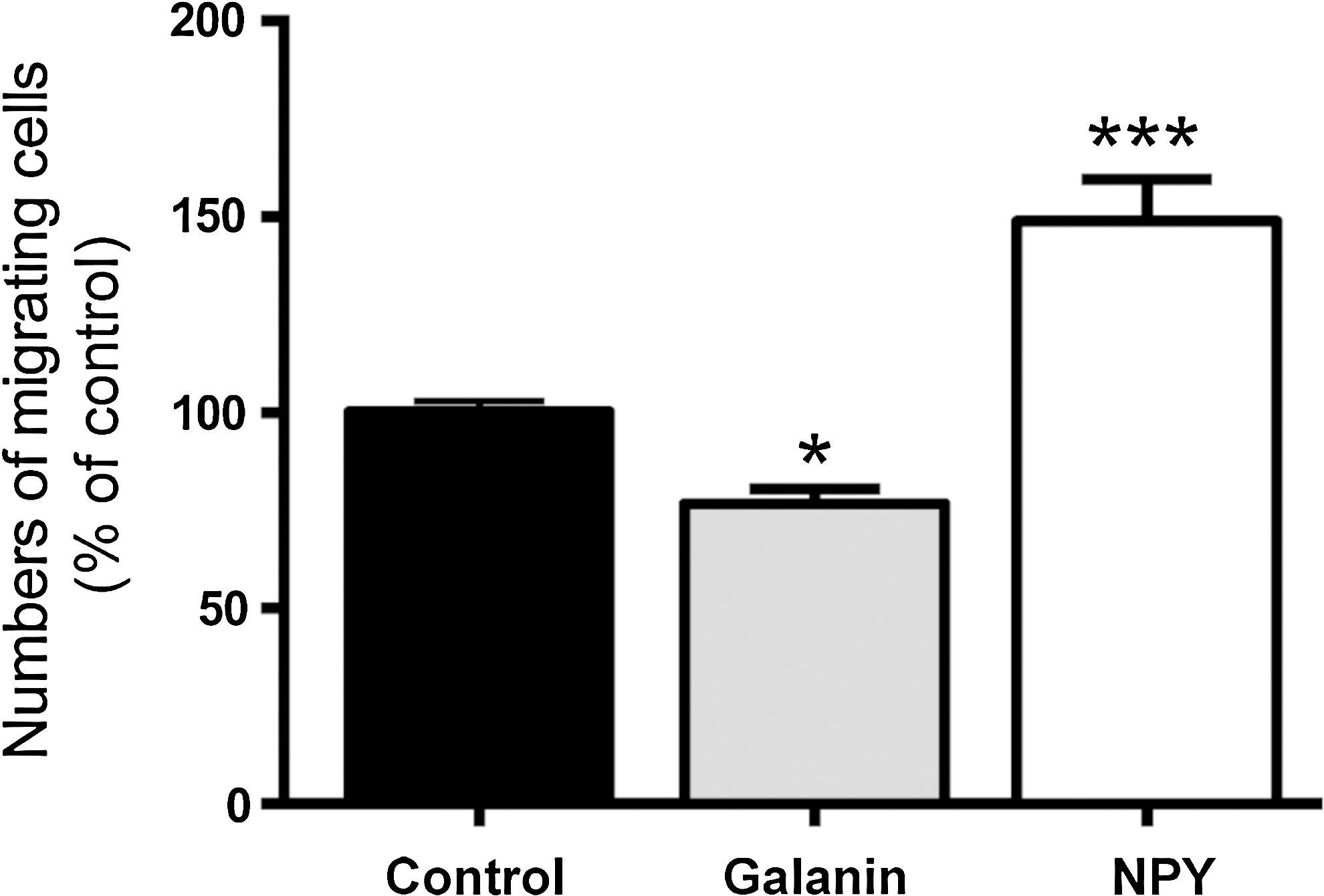
Galanin reduces SVZ cell migration. Bar graph depicts numbers of migrating cells determined after 6 h incubation with 2 μM galanin, or 2 μM neuropeptide Y (NPY) as a positive control, and expressed as a ratio to that obtained in serum-free medium (control). Data represent means±SEM of three independent experiments. *P<0.05, ***P<0.001, using one-way ANOVA with Dunnett's post hoc test for comparison with the control condition (set to 100%).
Discussion
This study tested the effects of galanin on stem cell properties and neurogenesis in SVZ stem/progenitor cell cultures. We first showed by qPCR that galanin is not expressed in SVZ neurospheres, while the mRNA for all galanin receptors are present. However, galanin mRNA was found in situ in dissected SVZ tissue samples in accordance with a previous study [44], suggesting that SVZ cells may secrete galanin within their microenvironment. Nevertheless, we cannot conclude whether the mRNA is present in SVZ stem cells in vivo or rather in the differentiated cells of the neurogenic niche. Indeed, galanin is expressed by a variety of cell types, including non-neuronal cells, such as oligodendrocyte precursors and astrocytes [44 –46] and astrocytes are cellular components of the SVZ [2,37]. Expression of galanin by mature cells of the SVZ in vivo rather than by stem/progenitor cells would explain why no galanin mRNA was found in neurospheres since the culture conditions applied sustain exclusively the expansion of stem/progenitor cells. In addition and consistent with a previous mouse study [47], galanin binding sites were found to be present in adult mice corresponding to the SVZ and, more importantly, galanin-stimulated [35S]GTPγS functional binding was also present, further establishing that galanin may exert biological effects on SVZ cells. We also identified galanin mRNA in the choroid plexus, suggesting that galanin is secreted into the cerebrospinal fluid and thereby may exert its action on SVZ cells. Another potential source of galanin could be the adjacent caudate-putamen which was shown in the present study to contain galanin mRNA and has been shown to contain galanin-immunoreactive fibres in mice [48]. Recent studies showed that galanin is expressed by pluripotent embryonic stem cells and by bone marrow mesenchymal stem cells [25 –28] suggesting that galanin expression may be used as a functional marker for stemness. However, we found no expression of galanin in SVZ cells, at least in vitro, and no effect of galanin on self-renewal, indicating that galanin expression is not a hallmark for stemness. Similarly, galanin is not found in nestin positive stem cells from the adult pancreas islets [49].
The principal finding of the present work is that galanin triggers neuronal differentiation of SVZ-derived cells. The increase in neurons obtained with galanin is independent of an induced proliferation of neuroblasts and/or progenitors and does not affect glial differentiation. Our results suggest a role for galanin as an enhancer of the process of neuronal differentiation, firstly, by increasing the commitment of progenitors towards the neuronal fate and, secondly, by “accelerating” neuronal differentiation and maturation as evidenced by the increase in the proneurogenic transcription factor Ngn1 transcripts and activation of the Ngn1 gene (qChIP experiment). Moreover, we showed that galanin enhances neuronal maturation, promoting the acquisition of neurotransmitter phenotypes and extension of neurites. Therefore, galanin seems to act as an accelerator of the neuronal differentiation process. Using immunocytochemistry to detect NeuN+ neurons and functional evaluation of neuronal capacity to increase calcium after depolarization, we showed that neuronal differentiation was achieved through the activation of GALR1 and/or GALR2 receptors. It seems that the signaling induced by each of the two GALRs is necessary and sufficient for inducing the proneuronal differentiation effect of galanin since cotreatment of the SVZ cells with both GALR agonists had no additive effects. This suggests that a common signaling pathway is activated by the receptors. GALR1 and GALR2 couple to Gi and GALR2 also signals through G0 and Gq/11. GALR1 and GALR2 both activate the MAPK pathway via Gi and G0, respectively [9,50], and MAPK pathways are known to mediate neuronal differentiation [51]. Indeed, the MAPK proteins MEK1/2 and ERK1/2 mediate sensory neural development and neurite outgrowth induced by retinoic acid [52]. Moreover, galanin induces neurite outgrowth in hippocampal neural progenitor-derived neurospheres via GALR2/G0 and ERK activation [53], and increases neuronal differentiation in rat hippocampal precursor cell cultures [24].
In this study, galanin triggered an increase in the mRNA levels of the proneural gene Ngn1, while not affecting Mash-1 and Dlx2 mRNA levels. Moreover, galanin treatment increased chromatin modification of Ngn1 promoter region consistent with active transcription of the gene. These results suggest that proneurogenic effects of galanin are mediated via Ngn1 expression. Expression of the basic helix-loop-helix transcription factor Ngn1 correlates with neuronal commitment and differentiation. Several studies have demonstrated that the induced expression of Ngn1 is sufficient per se to commit P19 cell-line [54], embryonic stem cells [55], mesenchymal stem cells [56] into neurons, even in the absence of other neurogenic stimuli. Also, transgenic mice that lack Ngn1/2 genes exhibit a loss of neuronal progenitor cell population [57]. Importantly, Wang and colleagues have reported that a short-interfering RNA targeting Ngn1 abolished human recombinant erythropoietin-induced neuronal differentiation [58]. Recent studies also showed that Ngn1 mediates the proneurogenic effects of histamine on NSCs from the SVZ and the embryonic cortex [36,59].
Numerous pathways are activated after binding of galanin to the GALR1 and GALR2 in particular the MAPK pathways. It has been demonstrated that MAPK activation can alter gene expression by directly binding to promoter regions of target genes or by inducing the recruitment of histone modification and chromatin regulation complexes (for review see [60]). For instance, novel object recognition is favored in young female mice due to the activity of the hormone 17β-estradiol in the dorsal hippocampus. Estrogen activates ERK1/2 which leads to histone 3 acetylation and DNA methylation and mediates memory consolidation [61]. Also, GALR activation regulates the cAMP Responsive Element Binding Protein (CREB) pathway: in the hippocampus, galanin impairs LTP, and this is correlated with a decrease in CREB phosphorylation [62]. The CREB pathway is involved in the regulation of proneural gene, such as the basic helix/loop/helix NeuroD [63]. It could be speculated that galanin induction of Ngn1 expression in our cultures may be modulated by these pathways. Further studies are needed to confirm this issue.
We also showed that galanin does not specifically direct differentiation towards a specific neuronal phenotype as both populations of GABAergic and dopaminergic neurons are increased upon galanin treatment. In addition, we showed that galanin promotes neuronal maturation as it increases axonal elongation and dendrite branching in SVZ-derived neurons. Positive effects of galanin on neurite outgrowth and branching have already been established in cultured sensory neurons of the dorsal root ganglia [64 –67]. Moreover, galanin also induces neurite outgrowth in cultures of neurospheres from the hippocampus [53]. In vivo, galanin induces axonal sprouting of cranial motor neurons [67]. These effects have been attributed to activation of the GALR2 and subsequent PKC activation [64,65].
No effect on proliferation of SVZ cells was shown upon incubation with galanin, GALR1 or GALR2 agonists. Previous studies testing the effects of galanin and GALR agonists on stem/progenitor cell cultures have reported contradictory results. Indeed, the GALR2 agonist AR-M1896 has been reported to induce proliferation in rat hippocampal precursor cells and Prox1+ granule cell neurons [24], while galanin reduced proliferation in stem cells from E14 embryonic rat brain [68]. The fact that galanin did not affect SVZ cell proliferation in our study, while it promoted and inhibited proliferation in hippocampal precursor cells and in E14 brain stem cells, respectively, further demonstrates the specificity of different types of tissue-derived stem cells in terms of regulation and intrinsic properties. Interestingly, in other cell types, there is relative consensus that galanin inhibits proliferation by acting either via GALR1 or GALR2. Galanin reduced cell proliferation of glia olfactory ensheathing cells [46], head and neck cancer cells [69], and of GALR2 transfected pheochromocytoma (PC12) cells [70]. In oral squamous cell carcinoma, inhibition of proliferation of malignant cells is mediated by GALR1 activation [71,72]. Nevertheless, presence of galanin and GALRs is not always correlated to modulation of proliferation: in glioma, for instance, GALR1 and galanin are highly expressed, while no correlation between proliferation and GALR1 activation was demonstrated [73]. We found no effect of galanin or GALR1/GALR2 agonists on cell survival in contrast to other studies showing that galanin is a trophic factor, promoting the survival of basal forebrain cholinergic neurons and dorsal root ganglia cells [74,75]. Moreover, Abbosh and collaborators [24] showed that galanin exerts trophic effects on hippocampal precursor cells. Even further, galanin expression is increased upon injury both in the central and peripheral nervous systems, and galanin upregulation was correlated to neuroprotection [76]. Hippocampal neuronal cell death is exacerbated in both GALR1 and GALR2 knockout mice subjected to seizures, indicating that galanin is neuroprotective [21,22]. Galanin is upregulated in the brains of Alzheimer's disease patients and this increase has been suggested to promote the survival of cholinergic basal forebrain neurons, thereby supporting their cognitive functions [77]. Neuroprotective effects of galanin have been demonstrated in cultures of mouse septal/neuroblastoma (SN56) cells [78] and in hippocampal neuronal cells subjected to amyloid β toxicity [18].
Few studies have been conducted on the effect of galanin on cell migration. Galanin has been shown to stimulate migration of rat microglial cells [79] and bone marrow mesenchymal stem cells [28]. In contrast, our results clearly point to an inhibitory effect of galanin on SVZ-derived cell migration. The functional implications of this finding remains to be established; however, an inhibitory effect of cell migration could contribute to the processes of differentiation and maturation. In conclusion, we here report that galanin is an efficacious inducer of neuronal differentiation and maturation in SVZ stem/progenitor cells. As SVZ is the major reservoir of stem cells in the adult brain, galanin deserves further attention as a potential inducer of neuronal differentiation in future stem cell based therapies.
Footnotes
Acknowledgments
The authors acknowledge the Fundação para a Ciência e a Tecnologia–Portugal (POCTI/SAU-NEU/68465/2006; PTDC/SAU-NEU/104415/2008; PTDC/SAU-NEU/101783/2008), Fundação Calouste Gulbenkian (96542), Lundbeck Foundation, Beckett Foundation, Aase and Ejnar Danielsen Foundation, Family Hede Nielsen Foundation, Foundation of 2 July 1984 for the Fight Against Parkinsonism, La Ligue Contre le Cancer Grand Ouest Comité de la Vienne. The authors thank Dr. Florence Hofman for their helpful comments on the manuscript. Part of this work has been presented as a poster at the 2009 Society for Neuroscience annual meeting (Chicago, IL, USA).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.