
Abstract
Low back pain is common and degenerated discs (DDs) are believed to be a major cause. In non-degenerated intervertebral discs (IVDs) presence of stem/progenitor cells was recently reported in different mammals (rabbit, rat, pig). Understanding processes of disc degeneration and regenerative mechanisms within DDs is important. The aim of the study was to examine the presence of local stem/progenitor cells in human DDs and if these cell populations could respond to paracrine stimulation in vitro. Tissue biopsies from the IVD region (L3-S1) were collected from 15 patients, age 34-69 years, undergoing surgery (spinal fusion) and mesenchymal stem cells (MSCs) (iliac crest) from 2 donors. Non-DD cells were collected from 1 donor (scoliosis) and chordoma tissue was obtained from (positive control, stem cell markers) 2 donors. The IVD biopsies were investigated for gene and protein expression of: OCT3/4, CD105, CD90, STRO-1, and NOTCH1. DD cell cultures (pellet mass) were performed with conditioned media from MSCs and non-degenerated IVD cells. Pellets were investigated after 7, 14, 28 days for the same stem cell markers as above. Gene expression of OCT3/4 and STRO-1 was detected in 13/15 patient samples, CD105 in 14/15 samples, and CD90 and NOTCH1 were detected 15/15 samples. Immunohistochemistry analysis supported findings on the protein level, in cells sparsely distributed in DDs tissues. DDs cell cultures displayed more undifferentiated appearance with increased expression of CD105, CD90, STRO-1, OCT3/4, NOTCH1, and JAGGED1, which was observed when cultured in conditioned cell culture media from MSCs compared to cell cultures cultured with conditioned media from non-DD cells. Expression of OCT3/4 (multipotency marker) and NOTCH1 (regulator of cell fate), MSC-markers, CD105, CD90, and STRO-1, indicate that primitive cell populations are present within DDs. Furthermore, the possibility to influence cells from DDs by paracrine signaling /soluble factors from MSCs and from nondegenerated IVD cells was observed in vitro indicating that repair processes within human DDs may be stimulated.
Introduction
The degenerated disc (DD) is typically characterized by loss of disc height, fissure formation, cell clustering, and cell death, as well as a decrease in fluid binding ability. Hence, in DD disease, the flexibility properties of the IVD are drastically reduced. On the molecular level, a decrease in glycosaminoglycan production is seen as well as an increase in matrix degrading enzymes; for example, proteins of the metalloprotease family 1, 3, 9, and 13 [4 –6] and cytokines, for example, interleukin 1 beta [7,8].
It has been suggested that dysfunctional cells may be one plausible cause of DD. Cell-based therapy and/or stimulation of local immature cells might offer a more biological approach to regenerate the disc or slow down disc degeneration [9,10] as a complement to the treatment methods of today.
In nondegenerated IVDs, the presence of stem and progenitor cells has recently been reported in mammals in different localizations [11] and progenitor cells have been sorted out from the total IVD cell population [12 –14]. There is a gap of knowledge regarding these cell populations in human IVDs and their functions in DD. Therefore, to further understand the process of IVD degeneration, knowledge about regenerative mechanisms within DDs is of importance. Different types of cellular surface markers can be used to detect progenitor and stem cells within a tissue. OCT3/4 is a transcription factor and OCT3/4 expression is necessary for stem cells to sustain self-renewal and multipotent ability [15,16]. CD90 [thymocyte differentiation antigen 1 (THY-1)] is a glycoprotein expressed on mesenchymal stem cells (MSCs) [17,18]. Stromal cells antigen 1 (STRO-1) is a MSC marker associated with tissue growth and regeneration [19 –22]. CD105 (endoglin) is a transmembrane cell surface protein associated with, for example, human bone marrow MSCs, endothelia cells, and smooth muscle cells [17,23,24]. Furthermore, NOTCH signaling is a primitive interspecies universal signaling pathway that decides cell fate; for example, proliferation or differentiation in embryogenesis as well as in adult tissues [25,26]. JAGGED1 is a ligand to NOTCH1 that participate in NOTCH signaling [27,28] and is associated with cell communication present in stem cell niches [28,29].
The aim of the study was to examine the presence of local stem/progenitor cells in human DDs and their eventual response to stimulation by soluble factors in an in vitro model.
Materials and Methods
Human tissues and cells
Human DD tissue samples (2–3 on the Thompson scale, level LIV-SI) were collected from patients (n=15), (age range 33–69 years) undergoing spinal fusion surgery. All DD samples were divided for the following 2 analyses: (1) for RNA analysis, samples were placed in RNA later solution (Qiagen) and, thereafter, frozen in −80°C until analysis and (2) for immunohistochemical analysis, samples were placed in 4% formaldehyde (Histofix®), embedded in paraffin, and sectioned (5–7 μm). For cell cultures, DD cells were isolated from 4 of the donors; female age 32 (donor 1), male age 61 (donor 2), male age 41 (donor 3), male age 34 (donor 4), and non-DD cells were isolated from 1 patient with scolios undergoing anterior corrective surgery, age 19. MSCs were taken (iliac crest), from 2 donors; male age 50 [used for a conditioned medium in 1 of the pellet cultures (donor 1)] and from female, age 38 [used for a conditioned medium in 3 of the cell cultures (donors 2–4)]. Chordoma tissue (from the peripheral region of tumor) was taken from 2 donors; male age 69 and female age 70 during surgery. Different panels of cell surface markers, for example, the stem cell marker, C-KIT (CD117), are currently used (among laboratory methods) as part of diagnosis and classification of different cancer types [30,31]. Hence, chordoma tissues were hypothesized to serve as positive controls and as a comparison tissue type for the stem cell markers used in this study.
The study was approved by the local human ethics committee and the samples were collected with informed consent from all patients.
Histology
All tissues were treated with 4% formaldehyde before paraffin embedment. Paraffin sections (5–7 μm) were deparaffinized with xylene 2×10 min and rehydrated in 99%, 95%, and 70% ethanol for 5 min in each solution. Sections of DDs tissues, chordoma control tissues, and cell pellets were stained for morphology and proteoglycans with the Alcian Blue van Gieson method using standards protocols [32,33].
Isolation of RNA
The tissues were placed in RNA-later (Qiagen) and, thereafter, frozen in −80°C until analysis. Homogenization was performed in Trizol (Qiagen) using a Tissue lyser instrument (Qiagen). The homogenized tissue was centrifuged, the supernatant extracted with chloroform, RNA, and washed with ethanol. Thereafter, a clean-up step was done following the standard protocol for the RNeasy Mini Kit (Qiagen). cDNA was prepared from a minimum of 62.5 ng RNA using the High-Capacity cDNA Reverse Transcriptase kit with random hexamers. In 7/12 samples, preamplifications were performed using TaqMan PreAmp Master Mix (Applied Biosystems).
Quantitative real-time polymerase chain reaction analysis
All instruments, software and reagents were purchased from Applied Biosystems. Quantitative real-time polymerase chain reaction (PCR) analysis was performed using the instrument, 7900HT. The following TaqMan gene Expression Assays were used: OCT3/4 (Hs01895061_u1), CD105 (Hs00923996_m1), THY-1 (CD90) (Hs00174816_m1), BST1 (bone marrow stromal antigen 1, STRO1 (Hs00174709_m1), NOTCH1 (Hs00413187_m1), and Cyclophilin A (Hs99999904_m1) and CREBBP was used as reference genes. Real-time PCR was performed in 20 μL volume with 10 μL of 2×TaqMan® Gene Expression Master Mix and 1 μL of 20×gene expression assays, with standard temperature programs. Data were analyzed with the relative comparative method (the ΔΔCt method) and the gene expression results are presented in relative units according to User Bullentin 2 (Applied Biosystems).
Immunohistochemistry analysis
Sections were deparaffinized as described above. Sections thereafter treated with an antigen retrieval step which were used for all samples with a 10 mM citrate buffer, pH 6.0 in 90°C for 20 min. Sections stained for OCT3/4 (R&D systems) were blocked with 2% bovine serum albumin (BSA) (Sigma) and 0.5% Triton-X 100 diluted in the phosphate saline buffer and NOTCH1 samples with 3% BSA for 10 min in room temperature (RT). Secondary antibodies were applied to sections: goat anti-mouse horseradish peroxidase (HRP) (Jackson Laboratory) in dilution 1:150 or donkey anti-goat HRP (Santa Cruz Biotechnology). The sections were thereafter incubated in RT in a dark container for 3 h and visualized by using the TSA-Direct Cy-3 kit (Perkin Elmer) according to the manufacturer's instructions.
Sections stained for CD105, STRO1, NOTCH1, and CD90 were blocked with 3% BSA for 5 min, incubated over night in 4°C with primary antibodies: goat anti-CD105 (R&D Systems); dilution 1:100, mouse anti-STRO1 (R&D Systems); dilution 1:100, and mouse anti-CD90 (Abcam); dilution 1:500, NOTCH1 (Santa Cruz Biotechnology); dilution 1×100 in a 3% BSA blocking solution. Secondary antibodies; donkey anti-goat Alexa Fluor 546 and goat anti-mouse Alexa Fluor 568 (Invitrogen) dilution 1:250 were added and sections were incubated for 3 h in RT. Nuclear staining was performed using 4,6 diamino-2 phenylindole 0.05 μg/mL (Sigma-Aldrich) and sections were mounted with the Prolong Gold mounting medium (Invitrogen). Results were visualized by using NIKON eqlipse80i (NIKON, Tokyo, Japan) and NIS-elements software (NIKON, Tokyo, Japan). Negative controls were sections with isotype controls and/or sections with the primary antibody were omitted.
Harvesting of tissue/cells for cell cultures
Human MSC isolation and monolayer cell cultures
Human MSCs were aspirated from iliac crest, centrifugated in cell preparation tubes (Ficoll; Becton Dicksson) according to the manufacturer's protocol. The MSCs were seeded at a concentration 200×103 cells/cm2 and cultured in the Dulbecco's modified Eagle medium, with low-glucose, (DMEM-LG) medium (Invitrogen). The medium was supplemented with 2 mM L-glutamine, 100 U/mL/0.1 mg/mL penicillin/streptomycin (PAA Laboratories), 10 ng/mL β-fibroblast growth factor (Invitrogen), and 10% human serum. At 90% confluence, the MSCs were detached using the trypsin/ethylenediaminetetraacetic acid (EDTA) solution (Invitrogen), re-seeded in a concentration 15×103 cells/cm2 and thereafter cultured to passage 3 or 5.
Human IVD cell isolation and monolayer cell cultures
IVD cells were isolated from 4 patients with disc degeneration undergoing spinal fusion surgery and from 1 patient with scolios undergoing anterior corrective surgery. The IVD tissues were manually cut to small pieces with a scalpel. Tissue pieces were then treated with 0.8 mg/mL collagenase type II (Worthington) and incubated 24 h in an incubator (37°C, 7% CO2, 93% air). Single cells were seeded at the concentration 4.0×105 cells/cm2 in the DMEM/F12 (Invitrogen) supplemented with ascorbic acid 0.1 mg/mL (Apoteket), L-glutamine (2 mM; Gibco), penicillin/streptomycin (100 U/mL/0.1 mg/mL), 10% human serum in cell culture flasks. At 90% confluence, the NP cells were detached using the trypsin/EDTA solution (Gibco), harvested, and reseeded again at a concentration 7×103 cells/cm2 and cultured to passage 2.
Flow cytometry analysis
The MSCs (2 donors) were characterized for a set of accepted mesenchymal cell surface markers [34], such as CD105, CD166, CD34, and CD45 by flow cytometry (passage 0 and 3–5). Fluoroscecin-, phycoerythrin-, or peridinin-conjugated antibodies (Ancell) were used in flow cytometry analysis using a FACSAria flowcytometer instrument (Becton Dickson).
Preparation of conditioned medium
For studies of differentiation and proliferation of DD cells in a three-dimensional (3D) culture (pellet mass system (DD pellets) potentially influenced by soluble factors, the conditioned medium was prepared from MSCs (a conditioned MSC medium) and from non-DD cells (a conditioned NDDC medium). The cells were radiated at 25 Gy, and then seeded at 6×103/cm2 in cell culture flasks coated with 0.1% gelatin. The MSCs were cultured in the DMEM/F12-LG and the non-DD cells were cultured in the DMEM/F12 for 5 days. The medium was changed twice and collected and pooled during this period. The 2 types of conditioned medium were sterile filtered and stored at −20°C until usage.
3D DD cell cultures (pellet mass)
After expansion, DD cells (passage 2–3) from 4 donors were placed in a 3D cell culture system (pellet mass). About 200,000 cells were applied in polypropylene conical tubes and 0.5 mL of the conditioned medium/tube was added. The collected conditioned medium was supplemented with 5.0 μg/mL linoleic acid (Sigma-Aldrich), insulin, transferrine, and selenium (concentrate no. 41400-045; Life Technologies), 1.0 mg/mL human serum albumin (Equitech-Bio), 10 ng/mL transforming growth factor -β1 (R&D Systems), 10−7 M dexamethasone (Sigma-Aldrich), 14 μg/mL ascorbic acid (Sigma-Aldrich), and 1% penicillin–streptomycin (PAA Laboratories). These are additives commonly used in the chondrogenic medium [35,36]. The cells were thereafter centrifuged at 500 g for 5 min. 0.5 mL of the conditioned medium from MSC or non-DD cells (scolios) was added to the cultures. The medium was changed twice a week and the pellets harvested after 7, 14, and 28 days (except in one case were pellets were cultured only to day 7 and 14 due to shortage of cells).
Results
Histology of DD tissues
All the human IVD samples displayed morphological signs characteristic of degeneration, such as disorganized irregular extracellular matrix appearance and cell clusters (cell foci) (Fig. 1).
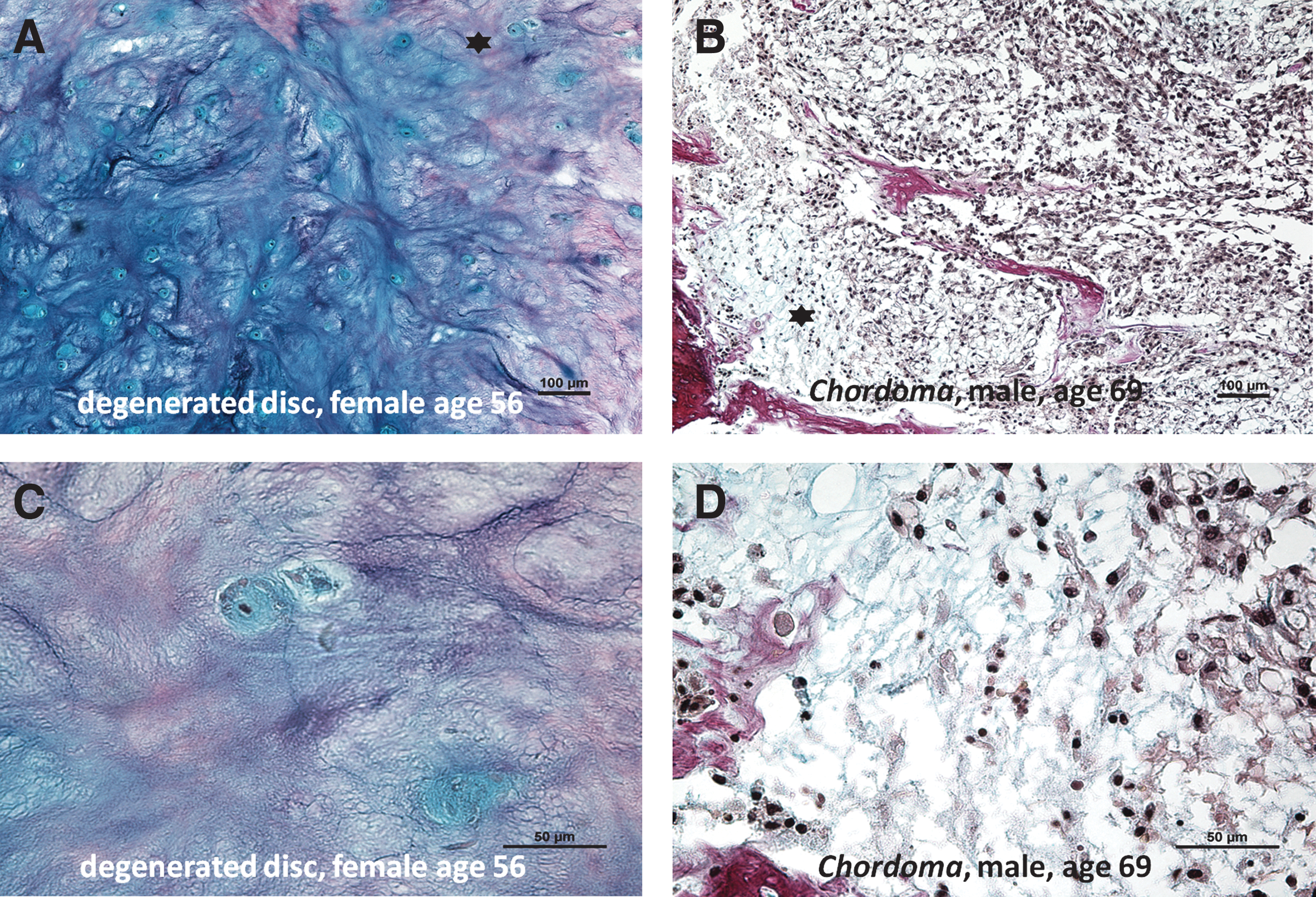
Alcian blue Van Gieson histological stainings (proteoglycans appears blue and collagens red) of
Stem cell/progenitor cells—gene expression in human DD tissue
Gene expression of OCT3/4 and STRO-1 was detected in 13/15 patient samples, CD105 in 14/15 samples, and CD90 and NOTCH1 was detected 15/15 samples (Fig. 2).
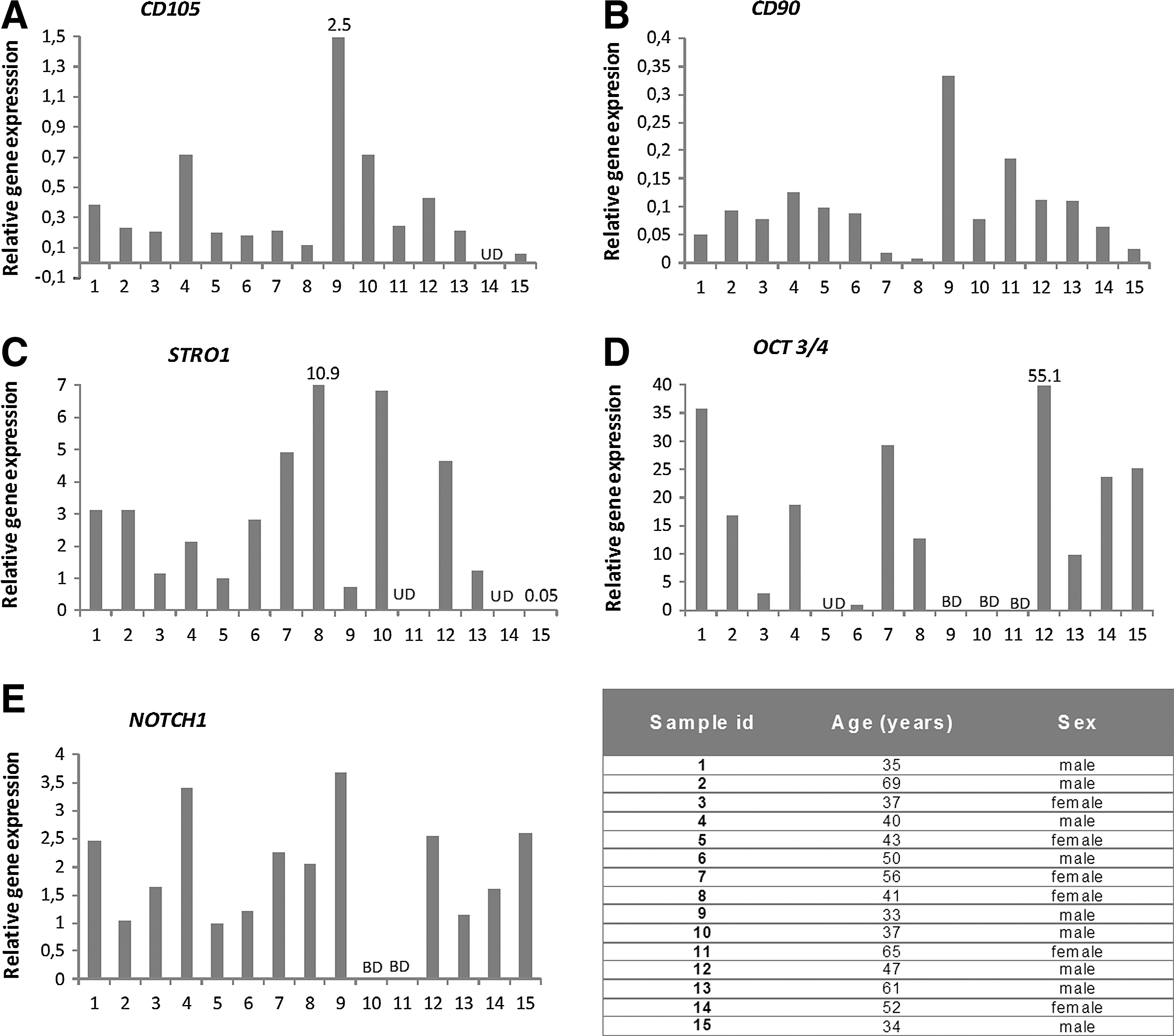
Tables are illustrating gene expression of stem/progenitor markers and NOTCH1 (marker involved in cell fate proliferation/differentiation) in human degenerated discs:
Stem cell/progenitor cells—protein expression in human DD tissue
CD105+, STRO-1+, OCT3/4+, and NOTCH1+ cells were found in 12/15 samples
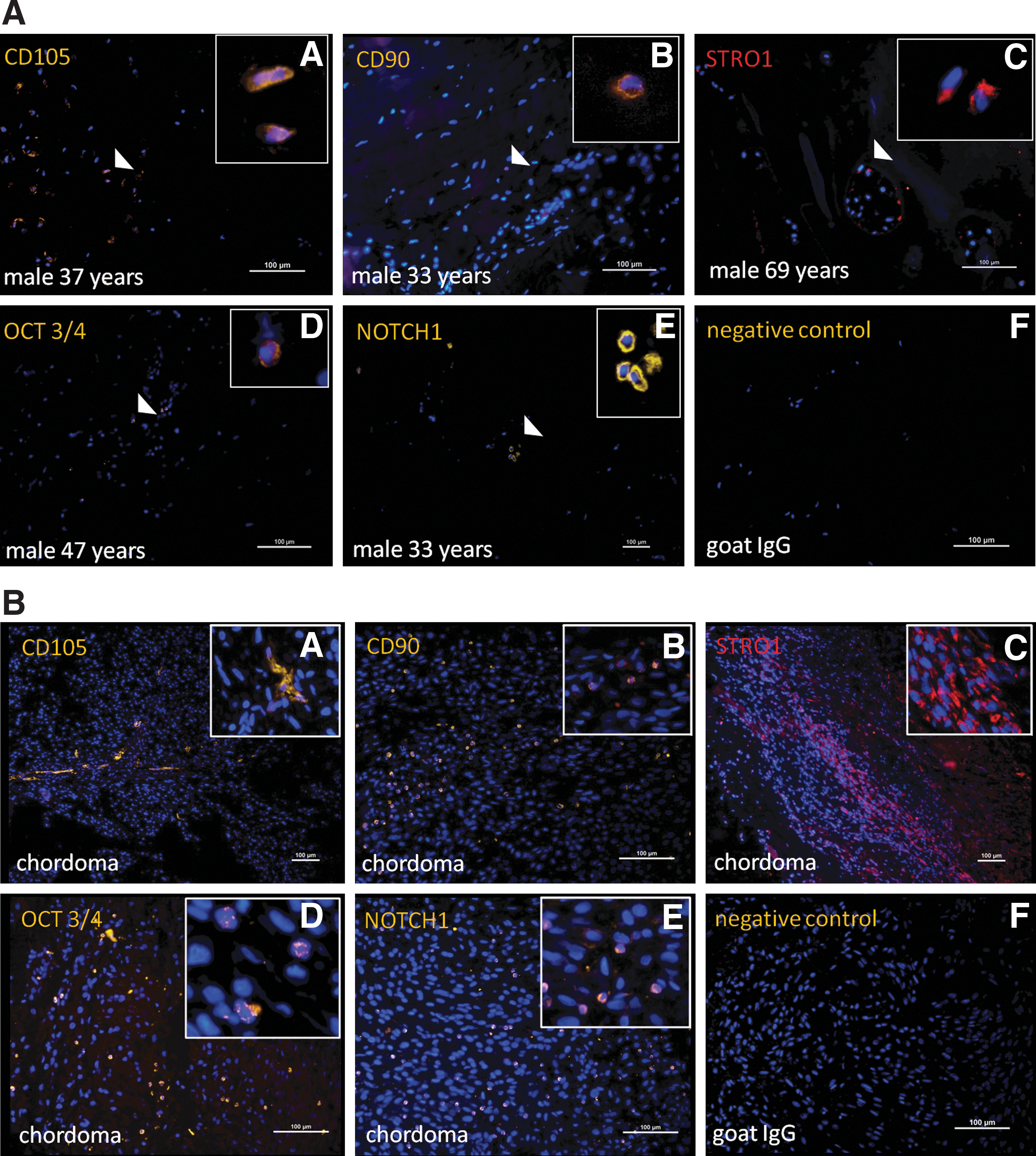
Score for tissue biopsies: 0=no detectable expression of marker, 1=< 2% positive cells/section, 2=2%–5% positive cells/section, 3=5%–10% positive cells/section, and 4=> 10% positive cells/section.
The chordoma tissue was used as a positive control for the stem/progenitor markers used in the analyzes.
Score for tissue biopsies: 2=2%–5% positive cells/section, 3=5%–10% positive cells/section, and 4=> 10% positive cells/section.
Flowcytometry results—human MSCs
Cells isolated from human iliac crest were characterized by staining for a set of accepted markers for MSCs; CD105 and CD166 (MSC markers), CD34 (expressed on hematopoietic stem cells), and CD45 (expressed on leukocytes) [34]. The flowcytometry analysis showed that MSCs markers increased during culturing time and with passages. Cell surface markers of MSCs were as follows (both donors passage 3 and 5): >92% of the cells were positive for CD105, >90% were positive for CD166, and <5% were positive for the markers, CD45 and CD34 (data not shown).
Stem cell/progenitor cells—protein expression in DD cell pellets stimulated with 2 types of conditioned medium
CD105+, CD90+, STRO+, and OCT3/4+ cells were found in DD cell pellets cultured in the conditioned medium from MSC as well as from non-DDs at the time point 7 days (except for donor 1, where no CD90+ cells were observed at time point 7 days in either medium). Thereafter, the DD pellets cultured in the conditioned MSC medium maintained the expression of stem cell markers CD105, CD90 and OCT 3/4 more pronounced and for a longer time period when compared to the pellets cultured in the conditioned medium from non-DD cells. STRO1+ cells displayed a maintained expression during the full culture time in both culture systems; however, the expression was more pronounced in pellets cultured in the conditioned medium from MSCs.
At the time point 7 days, NOTCH1+ and JAGGED1+ cells were present in the DD pellets cultivated in the conditioned MSC medium in 4/4 donors and in 3/4 donors when cultured in the conditioned NDDC medium. At the time point 14 days, NOTCH1+ cells were found in DD pellets from 3/4 donors cultured in either type of medium and JAGGED1+ cells were detected in DD pellets from 3/4 donors cultured in the conditioned MSC medium and in 2/4 donors in the conditioned NDDC medium.
At the time point 28 days, NOTCH1+ cells were found in DD pellets cultured in 2/3 donors for the conditioned MSC medium and in 1/3 donors for the conditioned NDDC medium. JAGGED1+ expression was detected in 3/3 donors in the conditioned MSC medium, while no expression was found in the pellets cultivated in the conditioned NDDC medium at the time point 28 days.
The expression of NOTCH and JAGGED was overall more pronounced in the DD pellets cultured in the conditioned MSC medium compared to the conditioned NDDC medium.
When summarizing the scores for all donors for one time point, the DD pellets cultivated in the conditioned MSC medium displayed a higher expression of all markers compared to DD pellets cultivated in the conditioned NDDC medium (Fig. 4; Table 3).
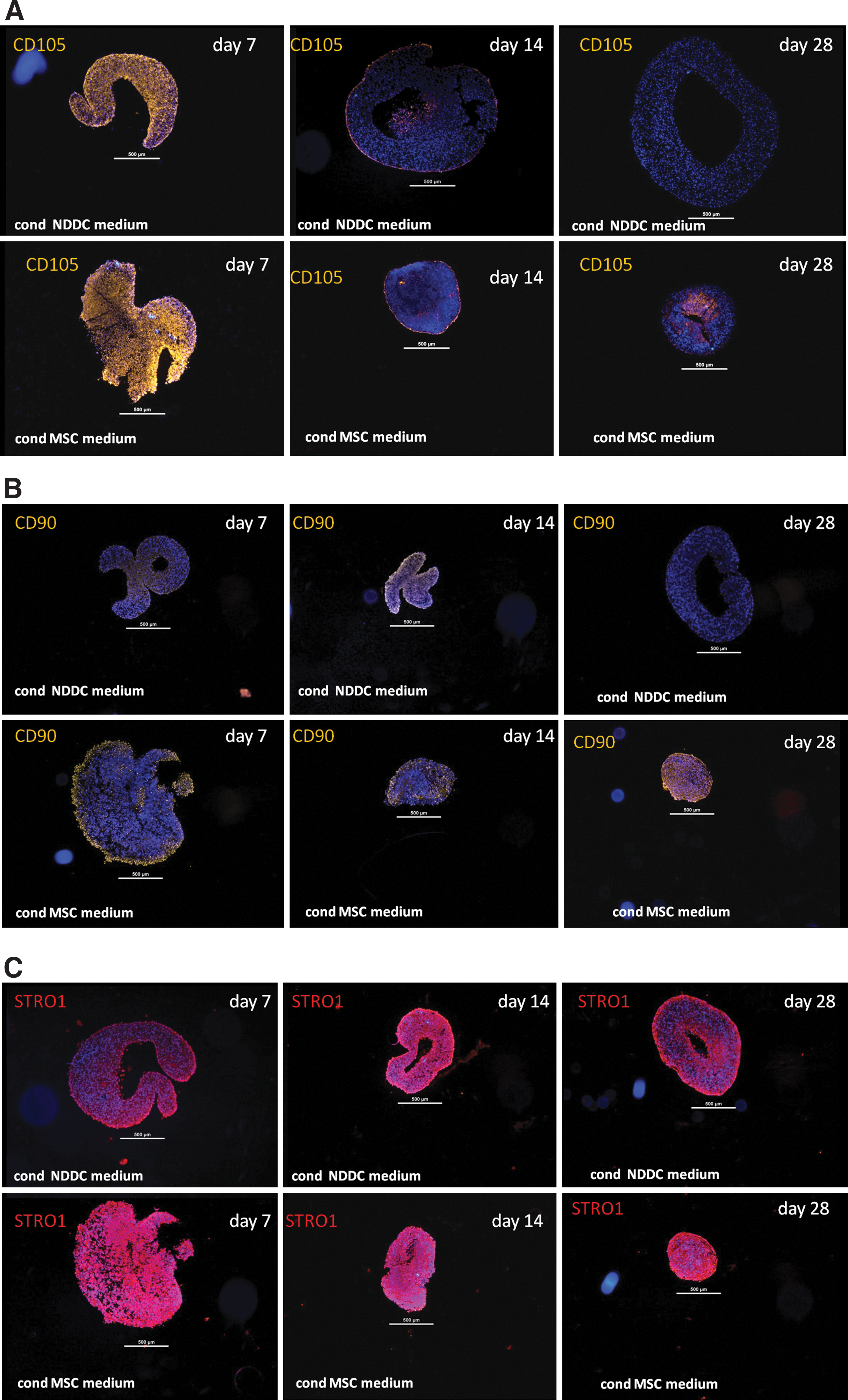
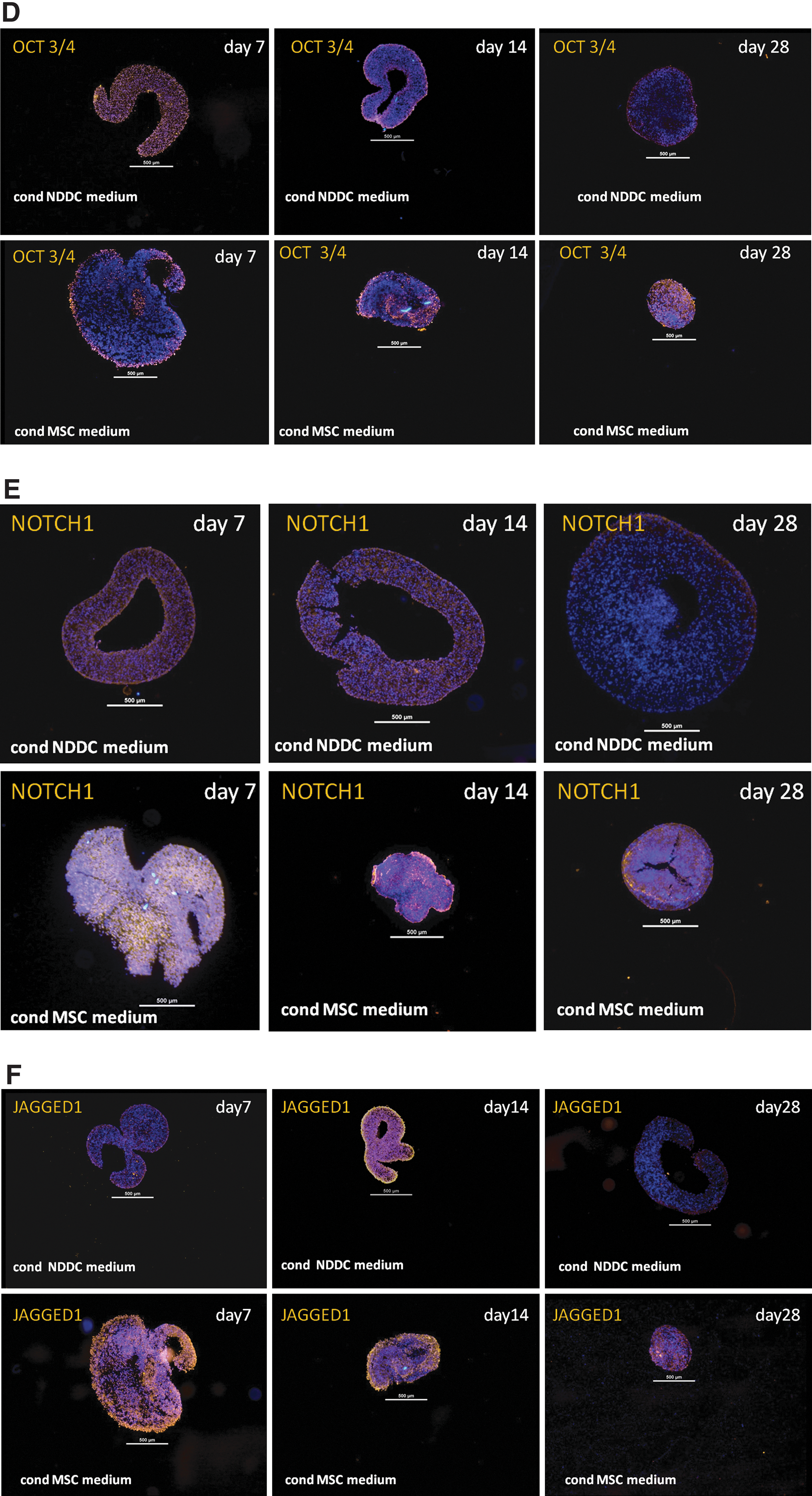
Score for pellets: 0=no detectable expression of marker, 1=< 10% positive cells/section, score 2=10%–50% positive cells/section, and score 3=> 50% positive cells/section.
The DD pellets cultivated in conditioned medium from mesenchymal stem cells (cond MSC medium) displayed a higher total sum of scores (all time points counted and summed) compared to degenerated disc cells pellets cultivated in conditioned medium from non-degenerated disc cells (conditioned NDDC medium).
Histology of DD cell pellets stimulated with conditioned medium
The DD cells formed cell pellets with various rounded shapes in the 3D cell cultures with the conditioned NDDC medium. Matrix accumulation was seen in pellets at the time points day 14 and 28. In the cultures with the conditioned MSC medium, the DD cells formed cell pellets with rounded or irregular shapes in the 3D cell cultures. No distinct matrix accumulation was observed in DD pellets cultured in the conditioned MSC medium at any time point (Fig. 5).
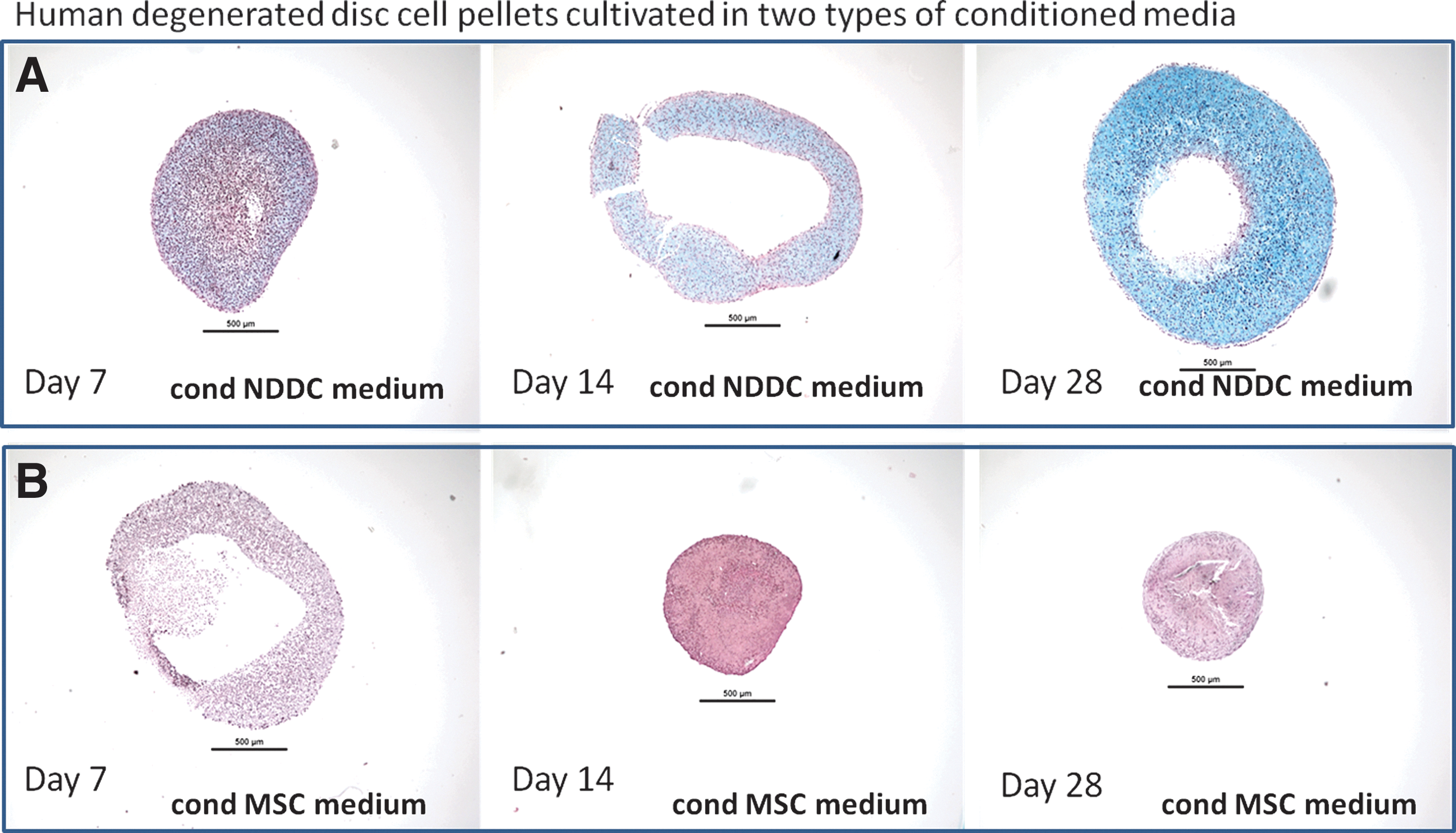
Alcian blue Van Gieson histological stainings (proteoglycans appears blue and collagens red) of a set of representative human (male age 40) degenerated discs cell pellets cultured in:
Discussion
The results from the present study shows the presence of cells with expression on both gene and protein level of OCT3/4, marker of multipotency and NOTCH1, regulator of cell fate, together with the classical MSC markers CD105, CD90, and STRO-1 are present in human degenerated IVDs. However, a relatively low expression of these markers was found in the IVD tissue which is to be expected. This indicates that primitive minor MSC populations are present within DDs. The results from this study is in alignment with a few other studies that has identified the presence of skeletal progenitor cells and stem/progenitor cells within human DDs [13,14], as well as in normal IVDs of other mammal species [11,12].
However, the presence of OCT3/4+ cells in the human IVD detected in this study is a novel finding that has not been described before. Further, the chordoma tissues displayed the presence of all the markers and illustrate that MSCs are present in this tumor type, including that chordoma tissue can function as a positive control on the protein level for the investigated markers. Chordomas are rare malignant bone tumors that in most cases develop at the sacrococcygeal region or the skull base, but in rare cases also in the mobile spine. They are believed to derive from notochordal cell remnants in the IVD and/or vertebrae and usually have a locally invasive growth pattern and metastasize late [37 –40].
Furthermore, in recent years, cancer also has come to be believed as a disease which at least in some cases, originates from abnormal hyperactive stem cells [14,41 –43] and it has been observed that chordomas can develop from the IVD region [39].
In addition, in vitro 3D cultures supported the existence of cells expressing stem/progenitor markers and indicated that DD cells can be influenced by paracrine stimulation with soluble factors from MSC as seen in the pellet cultures. This was demonstrated by the higher cellular expression of the markers OCT3/4, STRO1, CD90 and CD105, NOTCH and JAGGED1 (see the summed score for all markers in Table 3) in the DD cell pellets from all donors when cultured with the conditioned medium from MSCs compared to the medium from non-DDs cells.
During embryonic development of cartilage, mesenchymal progenitor cells undergo extensive proliferation followed by condensation. The pellet mass culture is a 3D cell culture system that mimics embryonic chondrogenesis and has been demonstrated to be a good model for studies involving processes, such as tissue induction and cellular differentiation and results in a differentiated, matrix-producing cell type when cultured in the chondrogenic medium [35,44,45]. Recently, an in vitro (pellet mass culture) study has shown that cell proliferation and chondrogenesis are tightly coupled processes when MSCs differentiate into chondrocytes [46,47]. In the same study, label retaining cells (syn. slow cycling cells e.g., stem cells) [48] were observed in MSC pellets by labeling of DNA methods. This type of interactions between proliferation and differentiation has been observed during embryonic cartilage development [49,50].
Furthermore, positive effects on matrix formation by pellet cocultures of DD cells and MSCs have been reported [36,51].
Observations from this study, where cell populations displayed ability to be stimulated in vitro to differentiate or to maintain a more undifferentiated state due to soluble factors secreted by different cell types suggest that DD cells may be possible to stimulate in vivo.
The limitations of the present study are that a relatively small sample size was used for the in vitro analyses. However, the soluble factors involved and their exact mechanisms in stimulation of DD cells remain to be identified and investigated.
In conclusion, this study showed that minor cell populations expressing stem cells and progenitor markers, including the marker of multipotency OCT3/4 are present in the adult human degenerated IVD. It was also observed that the chordoma biopsies displayed a higher expression of the stem cell/progenitor markers OCT3/4, NOTCH1, CD90 and CD105 compared to the DDs biopsies.
Possibly, these local immature cell populations present in the human degenerated IVD take part in repair attempts and may also be possible to stimulate with soluble factors as observed in the in vitro experiments performed in this study.
Further, this study shows that DDs cells can respond differently to stimuli through intercellular paracrine signaling; for example, soluble factors secreted from variant cell types in vitro and that the 3D-model used is a good model for such analyses. An increased knowledge about dynamic cell signaling processes can be a useful tool when developing future biological treatment options for patients with degenerated IVD disease.
Footnotes
Acknowledgments
This study was supported with grants from the Swedish Research Council, Dr. Felix Neubergh Foundation, A.L.F. Västra Götaland, Gothenburg Medical Association, the Swedish Society of Medicine, Vilhelmoch Martina Lundgrens Science Foundation 1, the Memorial Foundation of Sigurd and Elsa Golje, The Hjalmar Svensson Foundation and Sahlgrenska University Hospital Foundations.
Author Disclosure Statement
No competing financial interests exist.