
Abstract
Cerebral palsy (CP) is the most frequent neurological disorder associated with perinatal injury of the developing brain. Major brain lesions associated with CP are white matter damage (WMD) in preterm infants and cortico-subcortical lesions in term newborns. Cell therapy is considered promising for the repair of brain damage. Human umbilical cord blood mononuclear cells (hUCB-MNCs) are a rich source of various stem cells that could be of interest in repairing perinatal brain damage. Our goal was to investigate the potential of hUCB-MNCs to prevent or repair brain lesions in an animal model of excitotoxic brain injury. We induced neonatal brain lesions using intracranial injections of ibotenate, a glutamate agonist, in 5-day-old rat pups. hUCB-MNCs were injected either intraperitoneally (i.p.) or intravenously (i.v.) soon or 24 h after ibotenate injection, and their neurological effects were assessed using histology and immunohistochemistry. hUCB-MNCs injected i.p. did not reach the systemic circulation but high amounts induced a significant systemic inflammatory response and increased the WMD induced by the excitotoxic insult. This effect was associated with a significant 40% increase in microglial activation around the white matter lesion. hUCB-MNCs injected i.v. soon or 24 h after the excitotoxic insult did not affect lesion size, microglial activation, astroglial cell density, or cell proliferation within the developing white matter or cortical plate at any concentration used. We demonstrated that hUCB-MNCs could not integrate into the developing brain or promote subsequent repair in most conditions tested. We found that the intraperitoneal injection of high amounts of hUCB-MNCs aggravated WMD and was associated with systemic inflammation.
Introduction
It is hoped that stem cells will provide an inexhaustible source of therapeutic products that will enable neuroprotection and neuroregeneration in disorders affecting the brain and spinal cord. Various sources of stem cells have been described, but some, such as embryonic stem cells, are associated with ethical issues. Human umbilical cord blood is a rich source of stem and progenitor cells [4], notably mononuclear cells, which includes various stem cell populations that might be suitable for neuroprotection [5,6]. These human umbilical cord blood mononuclear cells (hUCB-MNCs) have many advantages.
• They are abundant in the cord blood and can be obtained easily with no harm to the infant or mother.
• They are less immunogenic than other stem cells because they are more immature [7].
• They provide a way to avoid the ethical issues associated with the use of embryonic stem cells.
hUCB-MNCs have been shown to have neuroprotective effects in adult animal models of traumatic or ischemic central nervous system injury [8,9]. However, not many groups are currently evaluating the interest of hUCB-MNCs in treating the perinatal brain, and those that do have revealed contradictory effects of transplanted hUCB-MNCs in animal models that mimic perinatal hypoxic-ischemic brain injury [10 –12]. For instance, some authors have demonstrated in a rat model of neonatal hypoxia-ischemia that hUCB-MNCs are associated with a reduction in the occurrence of spastic paresis [11] and significant improvements in sensorimotor performance and striatal neuron survival [12]. In contrast, another recent study has demonstrated no beneficial effects of hUCB-MNCs in the same model of neonatal brain damage [10]. Thus, the beneficial effects of hUCB-MNCs following perinatal brain damage are the subject of considerable controversy.
Here, we investigated the ability of hUCB-MNCs to prevent or repair white and gray matter lesions in a well-established animal model of excitotoxic brain injury that mimics the main features of brain damage in human preterm neonates [13,14].
Materials and Methods
Collection of hUCB-MNCs
To study the potential neuroprotection induced by hUCB-MNC infusion, human umbilical cord blood from both male and female neonates was obtained after normal full-term deliveries with maternal informed consent, according to approved institutional guidelines (Assistance Publique–Hôpitaux de Paris, Cord Blood Bank, Paris, France) and with the agreement of the Agence de la Biomédecine on behalf of the Assistance Publique-Hôpitaux de Paris.
The umbilical vein was punctured post partum, and blood was collected in umbilical cord blood collection bags containing citrate phosphate dextrose as an anticoagulant (Maco Pharma). Cord blood samples were processed within 24 h after collection.
Isolation of hUCB-MNCs
hUCB-MNCs were isolated by centrifugation on lymphocyte separation medium (Ficoll, d=1.077 g/mL; Eurobio), washed twice in phosphate-buffered saline (PBS)–ethylenediaminetetraacetic acid, and suspended in 0.9% sodium chloride solution or PBS before injection.
Monoclonal antibodies directed against CD34, CD45, CD3, CD4, CD8, CD14, CD19, CD133, or matched isotype controls and conjugated to either fluorescein isothiocyanate or phycoerythrin (all purchased from Becton Dickinson) were used for immunophenotyping, according to the manufacturer's protocol. Cell viability was assessed by using 7-AAD labeling. Data were acquired and analyzed using a 5-parameter flow cytometer (FACScalibur; Becton Dickinson) with CellQuestPro software (Becton Dickinson).
hUCB-MNC injection
hUCB-MNCs were injected within 12 h after their isolation. Before injection, cell viability was determined by the trypan blue dye exclusion method, and cell concentration was adjusted to obtain the desired concentration.
Animals were randomly assigned to 1 of 2 experimental groups: control, untreated animals (lesioned by ibotenate injection and injected with saline), and treated animals (lesioned by ibotenate injection and transplanted with hUCB-MNCs).
hUCB-MNCs were either injected intraperitoneally (i.p.) or intravenously (i.v.). The intraperitoneal injection of cells was either “early” (within 1 h following ibotenate injection) or “delayed” (24 h following ibotenate injection). Similarly, the intravenous injection of cells was either “early” (within 6 h following ibotenate injection) or “delayed” (24 h following ibotenate injection).
For the intraperitoneal injection, cells were diluted in 200 μL of saline solution and cell number was adjusted to test 3 different doses (106, 3.106, and 107 cells). The injection was carried out using a 26G needle (BD Biosciences). For the intravenous injection, cells were injected into the right jugular vein. Briefly, animals were anesthetized with inhaled isoflurane (1.5%), a skin incision was made to expose the jugular vein, and a 30G needle (BD Biosciences) was used to inject cells or control solution. This 30G needle was directly connected to a length of rigid polyethylene tubing (Phymep, I.D. 0.28 mm, O.D. 0.61 mm) that was connected at its other end to a 26G needle (BD Biosciences) together with a 1 mL syringe (Terumo) containing the solution to be injected. First, 100 μL of 0.9% NaCl was injected to check for the correct insertion of the needle into the jugular vein. Cells (106 or 107 per animal) diluted in 200 μL of 0.9% NaCl solution were then injected into treated animals over a 60-s period. Untreated animals received the same volume of 0.9% NaCl. Finally, 100 μL of 0.9% NaCl was again injected into all animals to avoid hUCB-MNC leakage during needle retraction. After needle retraction, mild pressure was immediately applied to the vein to prevent bleeding and animals were rapidly sutured.
No immunosuppressants were used in any experimental group.
Rat model of excitotoxic brain lesions
All experiments were carried out in compliance with the ethical rules of the INSERM. The study and animal protocols were approved by the research council review board of the Robert Debré hospital animal housing facility. Animals (Sprague-Dawley, Janvier S.A.S, Le Genest-St-Isle, France) were housed in a temperature-controlled room (19°C–23°C) with a 12 h/12 h light cycle and food and water ad libitum.
Because excitotoxicity is known to play a key role in developing brain vulnerability, we used in this study a well-characterized rat model of excitotoxic-induced brain injury [15 –17].
Briefly, ibotenate (10 μg; Tocris) was injected intracerebrally (i.c.) into postnatal day 5 (P5) rat pups, as previously described [13]. Ibotenate is a glutamate analogue that activates both N-methyl-D-aspartate and metabotropic receptors, but does not activate alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionate or kainate receptors. From each treatment group, 7–15 animals from both genders were sacrificed by decapitation 5 days after the injection (i.e., on P10), and brains were processed as previously described [18]. In all the experiments described below, lesion size in each pup was determined by 2 investigators blind to the treatment group.
Determination of lesion size
The brains of rat pups sacrificed as described previously were immediately fixed in 4% formalin and kept in this solution for 5–7 days before paraffin embedding. Coronal sections of 16-μm thick were cut and every third section was stained with cresyl violet. The size of neocortical and white matter lesions can be defined by their extent along 3 orthogonal axes: the lateral-medial axis (in the coronal plane), the radial axis (also in the coronal plane, from the pial surface to the lateral ventricle), and the fronto-occipital axis (in the sagittal plane). In previous studies [13,18], we found an excellent correlation among measurements along the 3 axes. Based on these findings, we cut serial sections of the entire brain in the coronal plane for this study. This permitted an accurate and reproducible determination of the fronto-occipital extent of the lesion in the sagittal plane (equal to the number of sections in which the lesion was present multiplied by 16 μm). We used this measure as an index of lesion volume.
Immunohistochemistry
In each experimental group, we studied 6 to 10 pups sacrificed on P10 in 3 separate experiments. Immunolabeling with primary antibodies [Tomatolectin (Vector; B-1175), Ki-67 (Abcam; ab27619), and Glial Fibrillary Acidic Protein (GFAP; Sigma; G3893)] was visualized using the streptavidin-biotin-peroxidase method as previously described [19]. Alternatively, a streptavidin-Cy3 conjugate or streptavidin-Alexa Fluor 488 conjugate was used to obtain fluorescent labeling. Quantitative analysis was performed in the same region for all brain sections (+2.16 to −0.36 mm from the bregma). Labeled cells were counted at 40× magnification within a 0.065 mm2 area on 4 sections per animal. For the quantification of Tomatolectin-positive cells, labeled cells were counted along the cystic lesion perimeter. For each section, the lesion perimeter was measured using a computerized image analysis system (Image J 1.41o, NIH) and the number of labeled cells per unit of perimeter length was calculated.
Luminex analysis of serum cytokine concentrations
Serum concentrations of 9 cytokines were measured using the Luminex xMAP (multianalytic profiling) technique according to the manufacturer's guidelines (Bio-Rad). The detection and quantification of cytokine levels was performed using a Bio-Plex 200 system (Bio-Rad). Analysis of data was performed using Bio-Plex Manager 5.0 software (Bio-Rad).
Polymerase chain reaction
To determine whether human cells were recruited and persisted in different rat organs, we checked for the presence of human DNA in different organs and at different time points after cell injection. Animals (n=3 per group) were sacrificed by decapitation at different time points after cell or saline injection (5 min, 30 min, 2 h, 24 h, and 5 days). Organs (liver, spleen, lungs, and brain cortices including the white matter) were rapidly harvested and immediately snap-frozen in liquid nitrogen and stored at −80°C. Genomic DNA extraction was performed using the QIAamp DNA mini kit (Qiagen; Cat. No. 51304) following the manufacturer's instructions.
DNA samples (100 ng per reaction) were analyzed using polymerase chain reaction (PCR) to specifically detect a fragment of the human CEP68 gene [20]. PCR was performed with a Bio-Rad MyiQ Thermal Cycler using the Taq DNA Polymerase kit (Qiagen; Cat. No. 201205), with hybridization at 62.4°C for 35 cycles. The following primers were used: CEP68-Forward: 5′-gcaccttgatatgggctctc-3′ CEP68-Reverse: 5′-aagtgcctggtcaacatgaa-3′. PCR products were analyzed by 2% agarose gel electrophoresis.
Statistical analysis
Values are expressed as means±SEM. Statistical comparisons between control and experimental groups were performed using one-way analysis of variance followed by Dunnett's multiple comparison test, and a nonparametric Mann-Whitney test in other cases. Statistical tests were run on GraphPad Prism 5.0 for Mac OS (GraphPad Software).
Results
Effect of intraperitoneal injection of hUCB-MNCs on excitotoxic brain lesions in the neonatal rat
Rat pups injected on P5 with ibotenate i.c. developed cortical lesions and periventricular white matter cysts in all cases. The cortical lesion was characterized by neuronal loss in all neocortical layers and the almost complete disappearance of neuronal cell bodies along the axis of excitotoxin injection. WMD was characterized by a periventricular cystic lesion surrounded by glial activation. The effect of an intraperitoneal injection of hUCB-MNCs on these excitotoxic brain lesions was investigated using various cell doses (106, 3.106, or 107 per animal). The lowest dose of hUCB-MNCs used at an “early” time point (see the Materials and Methods section) following ibotenate injection was associated with a small but significant decrease in the cortical brain lesion (P<0.05). In contrast, in the white matter, the injection of hUCB-MNCs early after the excitotoxic insult resulted in a mild dose-dependent increase of lesion size (Fig. 1A). This effect was found to be statistically significant only with the highest dose of intraperitoneal hUCB-MNCs used (107 per animal). Similar deleterious results in the white matter were observed when a high dose of hUCB-MNCs was injected at a “delayed” time point (see the Materials and Methods section) following ibotenate injection, that is, 24 h after the insult (data not shown). Because the intracerebral injection of excitotoxins is associated with the infiltration of monocytes/macrophages to surround white matter lesions, we measured microglial cell density using Tomatolectin, a marker for macrophages. We found that an intraperitoneal injection of 107 hUCB-MNCs was associated with a significant 40% increase in the number of Tomatolectin-positive cells per unit length of the lesion perimeter (Fig. 1B–D). These findings were all independent of gender. To analyze cell types in the glial scars in the injured white matter, the density of mature reactive astrocytes was assessed by counting GFAP-positive cells in P10 rat pups (Fig. 1F). This density was found to be comparable in control pups and pups exposed to intraperitoneal hUCB-MNCs. Similarly, cell proliferation as assessed using an antibody to Ki-67 was not affected by intraperitoneal hUCB-MNCs (Fig. 1E).
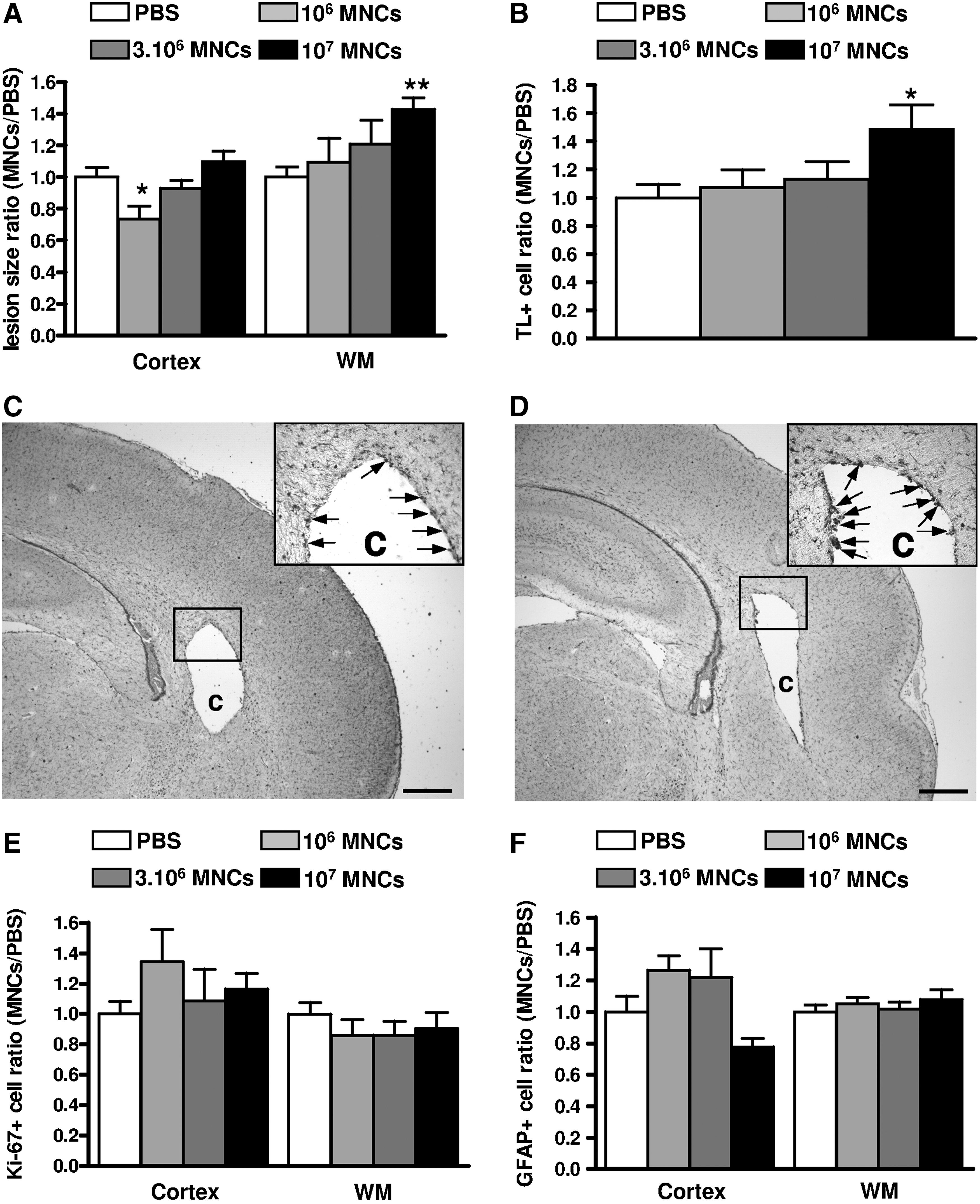
Intraperitoneal injection of human umbilical cord blood mononuclear cells (hUCB-MNCs) aggravates excitotoxic white matter (WM) lesions in neonatal rats.
Systemic inflammatory response to the intraperitoneal injection of hUCB-MNCs
Because microglial activation in the developing brain could be related to systemic inflammation [21], to which the developing white matter is known to be highly vulnerable, we next explored the impact of intraperitoneal hUCB-MNCs on cytokine production in serum of injected rat pups. The concentrations of several cytokines, including the main pro-inflammatory cytokines interleukin (IL)-1α, IL-1β, IL-6, and interferon-γ, were found to be increased 24 h after the intraperitoneal injection of hUCB-MNCs when compared with concentrations in PBS-injected controls (Fig. 2). This systemic inflammatory response was not related to the presence of detectable numbers of injected cells, as assessed using PCR for a human-specific gene fragment of CEP68 [20], in either the blood or the lung at any experimental time point from 5 min to 5 days (Fig. 3A, B).
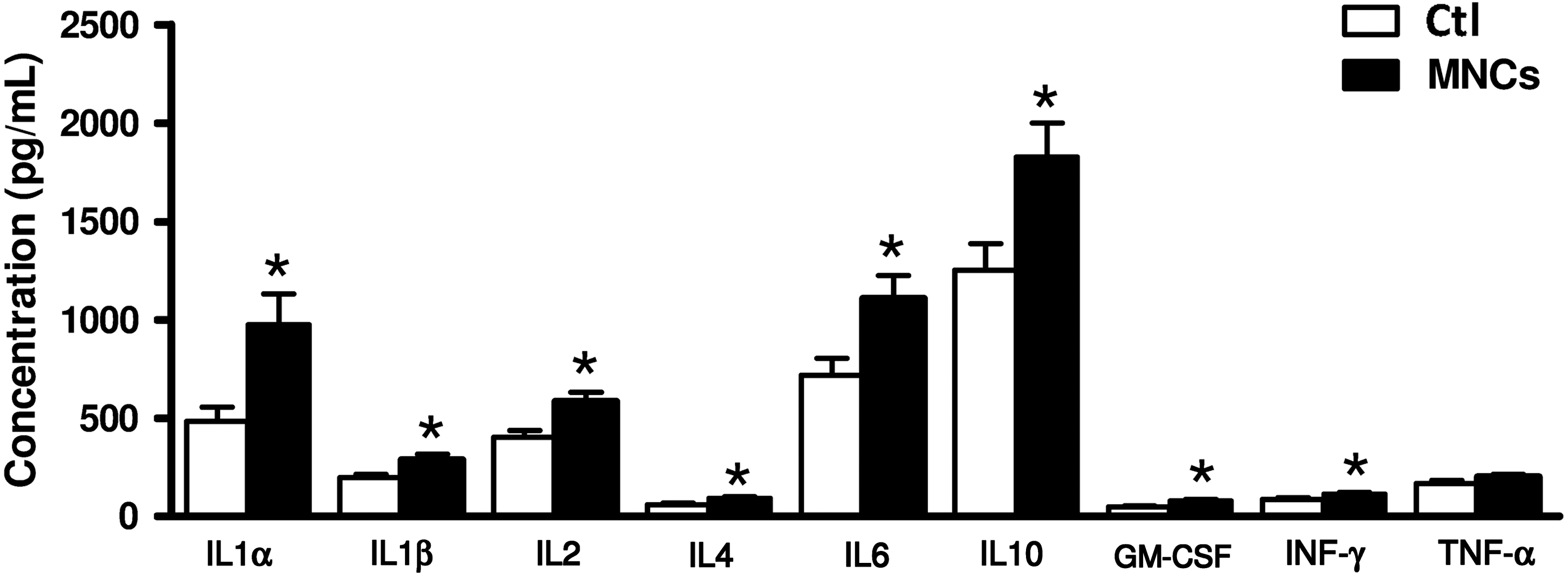
Systemic inflammatory response following intraperitoneal injection of hUCB-MNCs. LUMINEX multiplex assay for serum cytokines in P6 rat pups administered of 3.106 hUCB-MNCs i.p. and in PBS-treated controls (Ctl) (n=6 for each group). Cytokine concentrations were measured 24 h after treatment. *P<0.05 for MNC-treated animals compared with PBS-treated controls, using one-way ANOVA with the Dunnett's correction.
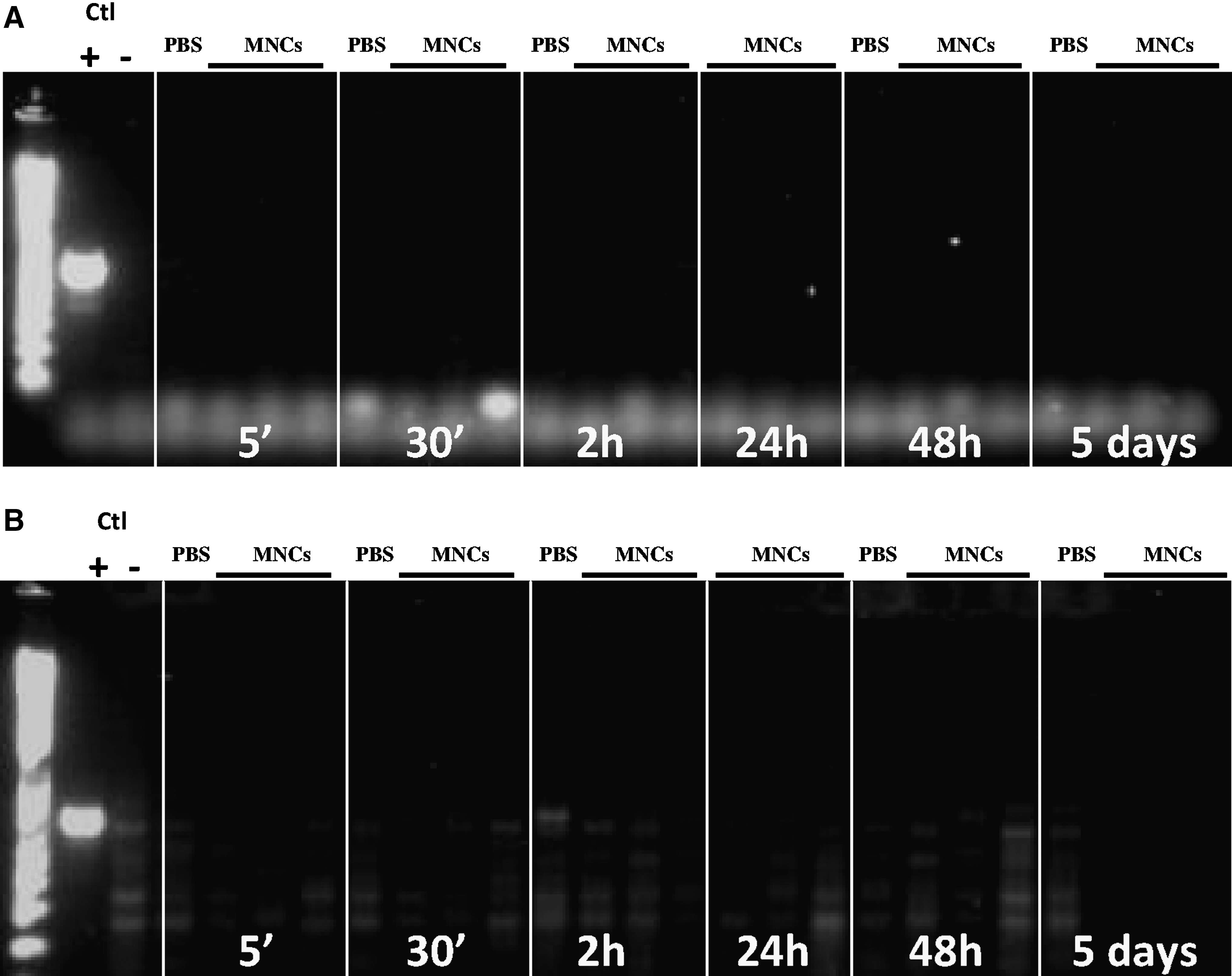
Detection of hUCB-MNCs in the blood and lung after intraperitoneal injection. No hUCB-MNCs were detected using polymerase chain reaction (PCR) for a human-specific gene fragment of CEP68 in either the blood
These data strongly support the hypothesis that hUCB-MNCs (107 cells) injected i.p. did not enter the systemic circulation but were able to induce a systemic inflammatory response and subsequent deleterious effects on the developing white matter challenged by the excitotoxic insult.
Effect of intravenous injection of hUCB-MNCs on excitotoxic brain lesions in the neonatal rat
Since hUCB-MNCs failed to translocate from the peritoneum to the blood circulation, and since intraperitoneal injections induced significant inflammation, we next investigated the effect of hUCB-MNCs injected directly i.v. in P5 rat pups. First, we measured serum cytokine concentrations 24 h after the intravenous injection of hUCB-MNCs and compared them with those in control pups. We found no difference in any of cytokines measured between these 2 experimental groups (Fig. 4). We then asked for how long hUCB-MNCs would remain detectable by PCR in the blood circulation as well as in several other organs, including the lung, liver, spleen, kidney, and brain. As expected, hUCB-MNCs were detectable at 5 min and up to 2 h after intravenous injection mainly in the blood, lung, and liver (Fig. 5A–C). In contrast, only small amounts of the PCR product were detected in the brain, indicating that the number of hUCB-MNCs entering it was low (Fig. 5D). hUCB-MNCs were no longer detectable in any tissue 24 h after intravenous injection.
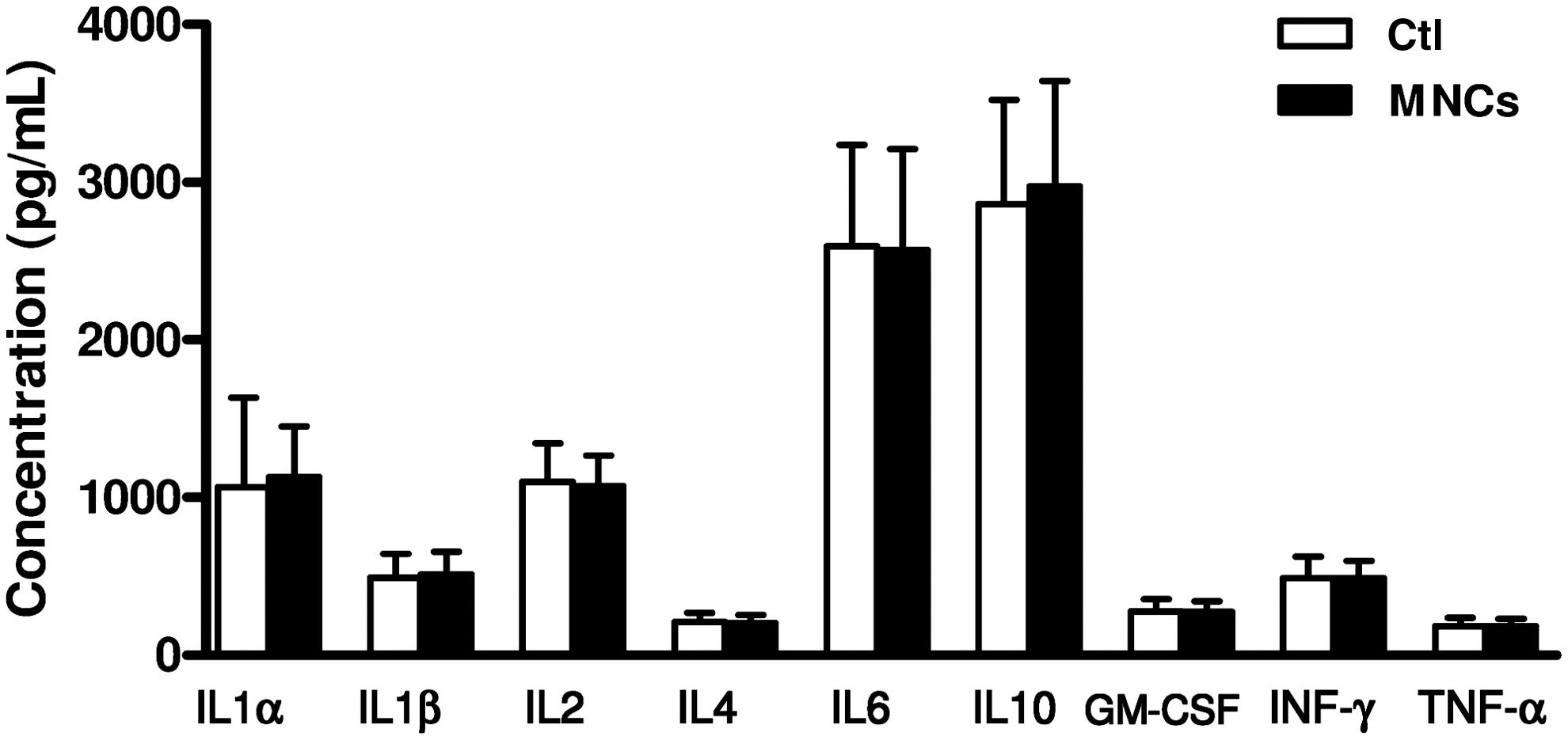
Systemic inflammatory response following intravenous injection of hUCB-MNCs. LUMINEX multiplex assay for serum cytokines in P6 rat pups administered 3.106 hUCB-MNCs intravenously (i.v.) and in NaCl-treated controls (Ctl) (n=6 for each group). Cytokine concentrations were measured 24 h after treatment. One-way ANOVA with the Dunnett's correction was used to compare MNC-treated animals with PBS-treated controls.
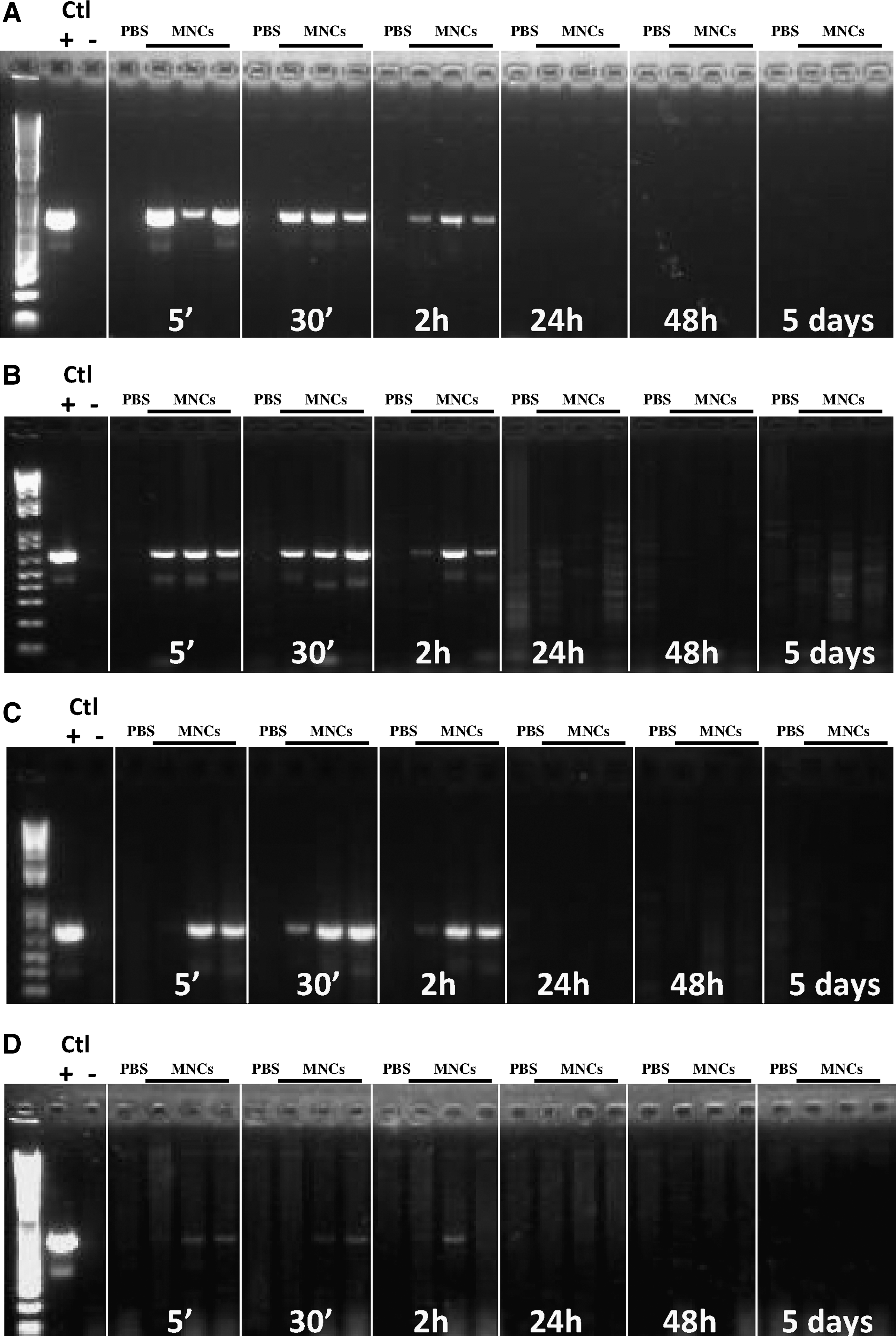
Detection of hUCB-MNCs after intravenous injection in the blood, lung, liver, and brain. PCR for a human-specific gene fragment of CEP68 was used to detect hUCB-MNCs in the blood
We explored the effect of intravenous hUCB-MNC injection after early (within 6 h) or delayed (24 h) injection following the excitotoxic insult. hUCB-MNCs used at 2 concentrations (106 or 107 per animal) did not affect lesion size in either the developing white matter or the cortical plate (Fig. 6A, B). Moreover, intravenous hUCB-MNC injection failed to induce any change in microglial activation around the cystic lesion (Fig. 6C, D). Both in the cortex and the white matter, the density of proliferating Ki-67+ cells and GFAP+ astrocytes was found to be similar in hUCB-MNC-injected animals and in controls (Fig. 6E–H).
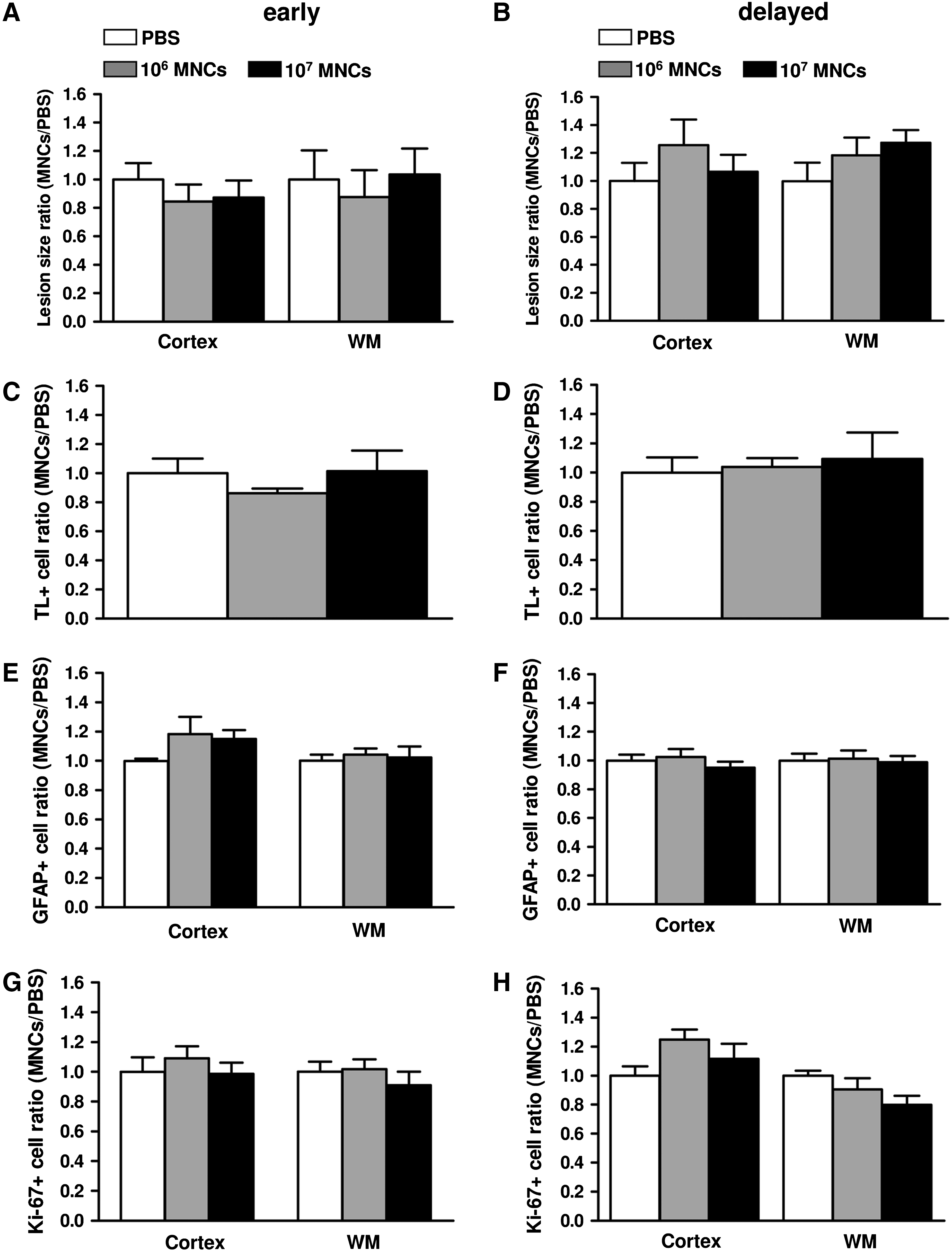
Intravenous injection of hUCB-MNCs has no effect on excitotoxic brain lesions in neonatal rats.
Discussion
This study shows that hUCB-MNC infusion is unable to reduce excitotoxin-induced neonatal brain damage in most conditions tested. Further, the intraperitoneal injection of high doses of hUCB-MNCs is associated with an inflammatory response in blood circulation and a mild increase in lesion size together with central nervous system inflammation.
A few groups have already reported the failure of hUCB-MNCs to induce neuroprotection either in the developing or the mature central nervous system. De Paula et al. demonstrated that P7 rats subjected to hypoxic-ischemic injury and injected with intravenous hUCB-MNCs showed no improvement in functional outcome (including performance in the Morris Water Maze) or lesion volume [10]. In another study conducted in spontaneously hypertensive adult rats subjected to stroke, the delayed intravenous infusion of hUCB-MNCs failed to reduce infarct volume or cell death [22]. According to Mäkinen et al., the systemic administration of hUCB-MNCs did not improve functional recovery or histological outcome in rats subjected to middle cerebral artery occlusion because of the limited migration of cells into the ischemic brain [23]. In all these cases, hUCB-MNCs displayed no or limited homing and engraftment into the ischemic brain. These reports are consistent with the findings of Coenen et al. showing that hUCB-MNCs transplanted into the embryonic ventricular zone remain largely confined to subventricular clusters, with little evidence for intraparenchymal integration [24]. In our study, a very sensitive PCR-based technique to detect hUCB-MNCs did not reveal their presence in the brain 24 h following intravenous injection or at any time point following intraperitoneal injection. We could speculate that low levels of PCR product seen in the brain at earlier time points following intravenous injection could be related to small amounts of residual blood harvested together with the brain parenchyma. We did not try other systemic routes, for example, intracarotid artery infusion, with which high number of cells would be docking directly into the ipsilateral hemisphere with lesion site. This route may be rather unrealistic in the case of neonatal rats and also not feasible in a clinical setting.
On the other hand, there have been several promising reports of the use of hUCB-MNCs to repair brain injury both in adulthood and during development. For instance, hUCB-MNCs injected either i.v. or i.c. induced functional recovery after ischemic injury in adult rats [25]. hUCB-MNCs also lessened inflammation and the overall consequences of brain damage resulting from adult stroke or traumatic insults [26]. In addition, they suppressed the recruitment of proinflammatory isolectin-binding cells during the period of infarct expansion after middle cerebral artery occlusion in adult rats [27]. In the developing brain, a few studies have revealed beneficial effects of transplanted hUCB-MNCs in animal models that mimic perinatal hypoxic-ischemic brain injury. A recent report has demonstrated a reduction in the occurrence of spastic paresis in a rat model of neonatal brain lesion (the Levine model) after the intraperitoneal administration of hUCB-MNCs [11], accompanied by a high incorporation rate at the lesion site with no signs of transdifferentiation. The authors have proposed a possible candidate for the chemotactic homing of transplanted hUCB-MNCs to the hypoxic-ischemic lesion [28]. Pimentel-Coelho et al. have also demonstrated improvements in sensorimotor performance and striatal neuron survival after intraperitoneal hUCB-MNC transplantation, although with very limited engraftment, in the developing brain [12].
As conventional cell replacement mechanisms become less probable, alternative pathways for graft-mediated brain repair, involving neurotrophic effects resulting from release of various growth factors that promote cell survival and angiogenesis and reduce inflammation, have recently been proposed [25]. Yasuhara et al. have reported that mannitol, by inducing blood–brain barrier permeabilization, facilitates neurotrophic factor expression and behavioral recovery in ischemic neonatal rats treated with hUCB-MNCs [29]. One limitation of many of these studies, however, is the lack of a clear identification of the mechanisms associated with neuroprotection and neuroregeneration.
A recent study has reported that delayed intravenous cord blood infusion reduces the inflammatory response to brain ischemia [26]. In contrast, in our study, the intraperitoneal injection of high doses of hUCB-MNCs was found to be associated with a systemic inflammatory response. Activated microglia/macrophages appear to play a central role in the pathophysiology of perinatal brain damage [30]. Indeed, microglial activation is more rapid after brain injury in neonates than in adults following transient focal ischemia with reperfusion [31], hypoxia-ischemia [32,33], or excitotoxic injury [34]. This could contribute to variable impact of cell therapy according to the developmental stage of the injured brain. The transplantation-induced neuroinflammation described here could be an additional indicator of the sensitivity but also potential serious risk of hUCB-MNC intraperitoneal transplantation (with a transition of M1 to M2 macrophage/microglia response).
Our study also has some limitations. First of all, the lack of a neuroprotective effect in most conditions tested in our study could be related to the absence of immunosuppressive treatment. Some studies have shown the survival of various types of transplanted cells for periods as long as 4 months, even though no immunosuppression was used [35 –37]. Conversely, in similar animal models, few transplanted cells could be identified a few weeks after transplantation in immunosuppressed animals [38]. Further studies are thus required to clarify whether immunosuppression is a help or a hindrance to obtaining the most beneficial outcome in xenotransplantation experiments.
A technical challenge faced by us and others was the detection of injected hUCB-MNCs at the lesion site. Indeed, despite similar routes of injection, previous studies have revealed highly variable cell homing rates after cerebral hypoxic-ischemic damage in neonatal rats [11,12]. In our experiments, we used several techniques, including fluorescent markers (Vybrant DiI, Invitrogen), human leucocyte antigen system (HLA) markers, and PCR for a human-specific DNA fragment (CEP68). Unlike the PCR product, we found that HLA markers and DiI were not specific to human cells. For example, we detected HLA markers in both injected and control animals. DiI labeling was found to be colocalized with a rodent microglial marker (ED1), suggesting that a transfer of this cytoplasmic marker to endogenous microglia is possible. Indeed, the in vivo transfer of several intracellular labels (DiI, BrdU, Bisbenzimide, and GFP) from locally implanted cells to resident macrophages and even neurons and astrocytes has already been demonstrated [39 –41]. Therefore, the molecular detection of human DNA as in our current study could be the most reliable way to assess the presence of transplanted human cells in rodents.
Finally, no behavioral assessment was carried out in this study. Some authors have shown functional improvements using hUCB-MNCs in the absence of an effect on lesion size [11,12,29]. Nevertheless, in our model and with most experimental conditions used we observed either a pro-inflammatory effect of a high dose of hUCB-MNCs when using the intraperitoneal route or no effect on microglial activation or the astrocytic response around the lesion with other injection protocols. Thus, although we cannot rule out the possibility, any behavioral improvement under these conditions would be highly unexpected.
In summary, we demonstrated that hUCB-MNCs administered through the intraperitoneal or intravenous routes could not integrate within the developing brain or promote its subsequent repair in most conditions used. On the contrary, we found that the intraperitoneal injection of a high dose of these cells aggravated the size of the brain lesion and was associated with systemic inflammation. While further preclinical studies are required to determine the protocol for obtaining the most effective neuroprotection, in this study we explored several critical experimental parameters, including the time window for transplantation after injury, the dosage of cells to be transplanted, and the route of administration, in a well-established rat model of neonatal brain injury (one condition 106 cells inject early intraperitoneally, was associated with interesting results on the lesion size, currently under study). The next step could be to use a similar experimental protocol to investigate the neuroprotective potential of other umbilical-cord-derived cells. While a number of promising neuroprotective strategies have been formulated using bone-marrow-derived mesenchymal stem cells following brain damage, both bone marrow puncture and the preparation of these cells through culture are severe limitations to their use in very preterm infants. The use of umbilical-cord-derived mesenchymal stem cells, easily obtained from the umbilical cord stroma at birth, could therefore provide a viable alternative to both hUCB-MNCs and adult stem cells.
Footnotes
Acknowledgments
This work was supported by Institut National de la Santé et de la Recherche Médicale, “Fondation de l'Avenir pour la Recherche Médicale Appliquée” project ETO-599, and “PremUP” Foundation. The authors are grateful to Sophie Lebon, Guillaume David, and Nadege Brunelle for helpful comments and discussion and technical assistance. This study was supported by grants from APHP (contrat d'interface to Dr. Pierre Gressens), Fondation Roger de Spoelberch, Fondation Grâce de Monaco.
Author Disclosure Statement
No competing financial interests exist.