
Abstract
Human embryonic stem cells (hESCs) are derived from the inner cell mass (ICM) of the blastocyst and can differentiate into any cell type in the human body. These cells hold a great potential for regenerative medicine, but to obtain enough cells needed for medical treatment, culture is required on a large scale. In the undifferentiated state, hESCs appear to possess an unlimited potential for proliferation, but optimal, defined, and safe culture conditions remain a challenge. The aim of the present study was to identify proteins in the natural environment of undifferentiated hESCs, namely, the blastocoel fluid, which is in contact with all the cells in the blastocyst, including hESCs. Fifty-three surplus human blastocysts were donated after informed consent, and blastocoel fluid was isolated by micromanipulation. Using highly sensitive nano-high-pressure liquid chromatography–tandem mass spectrometry, 286 proteins were identified in the blastocoel fluid and 1,307 proteins in the corresponding cells of the blastocyst. Forty-two were previously uncharacterized proteins—8 of these originated from the blastocoel fluid. Furthermore, several heat shock proteins (Hsp27, Hsp60, Hsc70, and Hsp90) were identified in blastocoel fluid together with zona pellucida proteins (ZP2–4), Vitamin D-binding protein, and Retinol-binding protein 4. Proteins that regulate ciliary assembly and function were also identified, including Bardet-Biedl syndrome protein 7. This study has identified numerous proteins that cells from the ICM of the human blastocyst are exposed to via the blastocoel fluid. These results can be an inspiration for the development of improved culture conditions for hESCs.
Introduction
Hitherto, the lack of highly sensitive methods for protein identifications, combined with the low amounts of proteins extractable from blastocoel fluid of 1–8 nL per blastocyst, has hampered an in-depth study of the human blastocyst proteome. However, recent developments in mass spectrometry-based proteomic techniques allow the identification and characterization of thousands of proteins from low microgram levels of protein extracted from tissues or cell cultures [7].
In the present study, we have collected the blastocoel fluid and the corresponding cells from 53 in vitro generated surplus human blastocysts, and we exploited the recent advances in proteomic technologies for mapping the proteome of this material. We identified 286 proteins in the blastocoel fluid and 1,307 proteins in the corresponding cells of the blastocyst.
Materials and Methods
If nothing else is mentioned, chemicals were purchased at Sigma-Aldrich (St. Louis, MA).
Generation and handling of human blastocysts
Surplus embryos were obtained from patients undergoing fertility treatment at Fertility Clinic IVF-SYD, Fredericia, Denmark, and Department of Women's and Children's Health, Uppsala University, Uppsala, Sweden, after informed consent and under the ethics approval numbers: H-KF-01-188/03 (Fredericia, Denmark) and Regional Ethics Review Board, Uppsala University 2008/046 (Uppsala, Sweden). The 2 clinics use slightly different culture methods for the human blastocysts, and therefore their culture methods are described separately. In both clinics, however, the embryos were generated from standard FSH-stimulated IVF cycles with conventional agonist/antagonist downregulation and human chorionic gonadotropin for ovulation induction.
Blastocyst generation in Fredericia, Denmark
Oocytes were fertilized in 0.5 mL medium (Universal IVF medium, ORIGIO, Maaloev, Denmark) in 4-well dishes (NUNC, Roskilde, Denmark) or using the standard intracytoplasmic sperm injection (ICSI) technique in cases of male factor infertility. After ICSI, the oocytes were transferred to a 30-μL microdroplet medium (ISM1-medium; ORIGIO) under liquid paraffin oil (ORIGIO), while conventional IVF-inseminated oocytes were transferred on day one to the ISM1 medium. All surplus embryos used for the study were thus cultured for a total of 5–6 days in ISM1.
Blastocyst generation in Uppsala, Sweden
Oocytes were fertilized in 30 μL microdroplets (G-IVF Vitrolife, Gothenburg, Sweden) under liquid paraffin oil (Ovoil 100; Vitrolife) or using the standard ICSI technique in cases of male factor infertility. Cultures of zygotes and embryos were maintained in droplets under oil. Cleavage-stage culturing was performed in a G-1 medium (Vitrolife), and blastocysts were cultured in the CCM medium (Vitrolife). Some embryos were cryopreserved on day 2–3 by a standard controlled rate cryopreservation using G-FreezeKit (Vitrolife), thawed, and cultured to blastocyst stage after reaching the maximum time limit for storage (5 years). At this point, the embryos where thawed using the Thawing Kit (Cook, Brisbane, Australia) and cultured to blastocyst stage in the CCM medium.
Isolation of the blastocoel fluid and sample collection
The blastocoel fluid was isolated from the blastocysts by micromanipulation using a Nikon Eclipse TE200 microscope (200×) with Narishige manipulators and pipettes from ORIGIO or The Pipette Company (Thebarton, Australia). A droplet of 5–10 μL culture medium (ISM1 or CCM) and 2 droplets of 3 μL 6 M urea were prepared in an ICSI dish and covered with paraffin oil. The blastocysts were transferred to the droplet with culture medium, and the dish was placed in the micromanipulator. The micropipette was coated with urea before sampling. In the medium droplet, the blastocyst was immobilized by the holding pipette with the ICM placed toward the holding pipette. The micropipette was then gently led in to the blastocyst cavity, and suction was applied to extract the fluid until the blastocyst collapsed (Fig. 1). The blastocoel was transferred to one urea droplet and the remaining cells to the other. After micromanipulation, the samples containing 6 M urea were transferred to low-bind PCR tubes using a urea-coated 10-μL pipette under a stereomicroscope and immediately frozen at −20°C or −80°C. To obtain adequate protein material for analysis, the samples were collected and analyzed in 2 pools. Initially, blastocoel fluid and blastocyst cells were isolated from 19 human blastocysts in Uppsala. Subsequently, the blastocoel and blastocyst cells were sampled from 34 blastocysts where 24 blastocysts were collected in Uppsala and 10 were collected in Fredericia. The MS data obtained from the total of 53 blastocysts are presented as 2 composed lists enclosing all proteins identified in the human blastocoel and remaining cells of the blastocyst, respectively. As a control of the background signal from the culture medium, a sample consisting of the ISM1 and CCM was analyzed as well and the proteins identified in the culture medium were excluded from the data presented in the Results section.
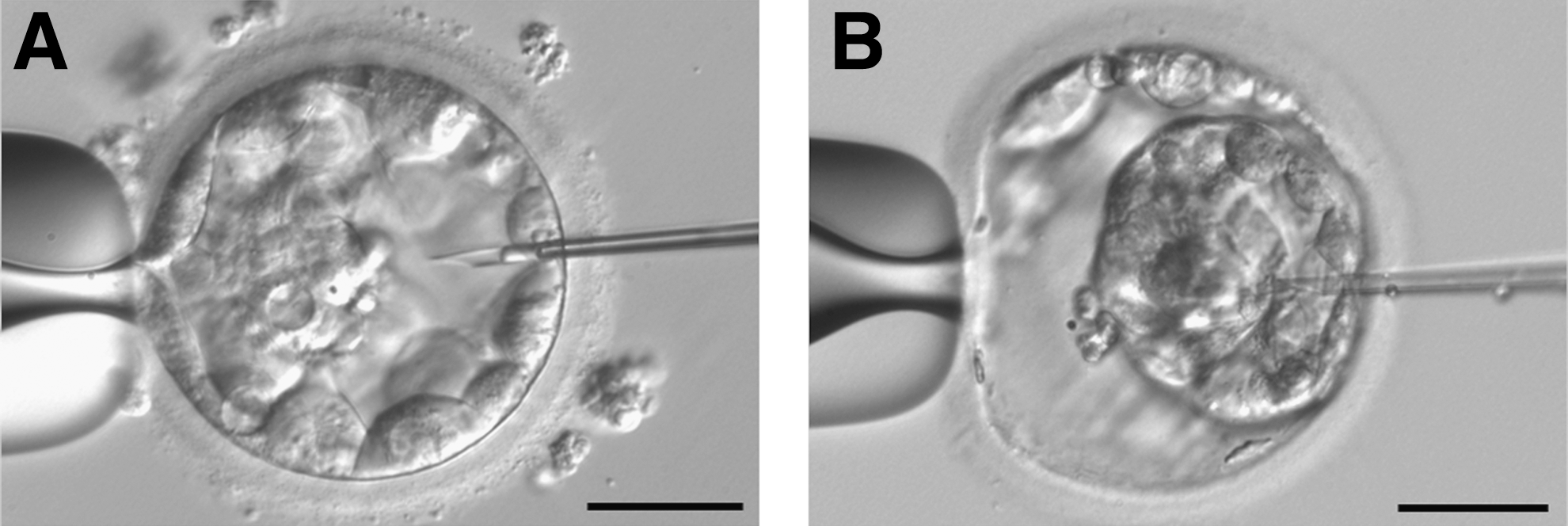
Photographs from the micromanipulation procedure. The human blastocyst is immobilized on a holding pipette
Proteomic analysis
The proteins of the blastocoel fluid or the lysed cells taken up in 6 M urea were digested by the addition of 10 ng lysC (Wako Chemicals, Richmond, VA), followed by a reduction of the disulfide bonds using 5 mM dithiothrethiol and blocking of free cysteine with 15 mM iodoacetamide. The samples were diluted to a urea concentration of 0.75 M and further proteolytically cleaved by adding 25 ng sequencing-grade trypsin (Promega, Madison, WI). The resulting peptide mixtures were purified using reversed-phase microcolumns and analyzed in duplicates by nano-LC MSMS analysis using a Dionex UltiMate 3000 nano-high-pressure liquid chromatography (HPLC; Thermo Scientific, Sunnyvale, CA) coupled to a Thermo Scientific LTQ Orbitrap Velos mass spectrometer (Thermo Scientific, Bremen, Germany). The samples were separated using nano-HPLC as previously described [8]; briefly, the samples (5 μL) were loaded onto a custom-made fused capillary column (20-cm length, 360-μm outer diameter, 75-μm inner diameter) packed with ReproSil Pur C18 3-μm resin (Dr. Maish, Ammerbuch-Entringen, Germany) with a flow of 600 nL/min and sequential eluted using a linear gradient from 98% solution A (0.1% formic acid) to 35% B (90% Acetonitrile in 0.1% formic acid) over 86 min, followed by 10 min at 90% B and 14 min at 98% A at a flow rate at 300 nL/min. Mass spectra were acquired in positive ion mode applying automatic data-dependent switch between one Orbitrap survey MS scan in the mass range of 300 to 1,500 m/z followed by HCD or CID fragmentation and Orbitrap detection of the 10 most intense ions observed in the MS scan. Target value in the Orbitrap for MS scan was 1,000,000 ions at a resolution of 30,000 at m/z 400. Fragmentation in the HCD cell was performed at normalized collision energy of 40 and CID at normalized collision energy of 35. Ion selection threshold was set to 10,000 counts. Selected sequenced ions were dynamically excluded for 60 s. A combined MASCOT-SEQUEST was performed where peak lists (.mgf files) were processed using the Proteome Discoverer 1.3 (version 2.2.3). The search parameters were set to MS accuracy 15 ppm, MSMS accuracy 0.1 Da for HCD data and 0.4 for CID, trypsin digestion with 2 missed cleavages allowed, fixed modification of cysteine blocked with carbamidomethyl, and variable modifications: methionine oxidation, deamidated asparagines, lysine acetylation, and lysine and N-terminal carbamylation. Tandem mass spectra were searched against the SwissProt database (downloaded January 30th 2012) restricted to humans. False protein identifications (1% or 5%) were estimated by searching MSMS spectra against the corresponding reversed-sequence (decoy) database. Proteins inferred on a basis of at least one unique peptide with a MASCOT score higher than 30 or a SEQUEST probability score (Xcorr) higher than 2.8 were included in the protein list. Trypsins originating from the protein digestion step were removed from the data. To estimate protein abundance in the blastocoel fluid and in the remaining cells of the blastocyst, the msms data were used, and the relative protein amount was estimated using the high-three approach in Proteome discover software [9].
Data analysis
Keratins listed in the cRAP database [10] were excluded from the data as contaminants from the sample procedure or protein analysis. The remaining keratins were searched in the Uniprot database [11], and proteins with function or localization merely to hair or skin were excluded as well. The proteins were grouped after their associated functions in specific biological processes using the gene ontology terms (GO terms) provided by the bioinformatics tool ProteinCenter (ThermoScientific) [12].
Culture of hESCs
LRB010, a hESC line derived in Laboratory of Reproductive Biology under the ethics approval number H-KF-01-188/03, was cultured as previously described in [13]. Briefly, the cells were cultured for 7 days on Matrigel (BD, Franklin Lakes, NJ) and in an hESC medium with knockout serum replacement (Life Technologies, Carlsbad, CA) and bFGF (R&D systems, Minneapolis, MN).
Immunohistochemical analysis
Human blastocysts were fixed in 4% paraformaldehyde and embedded into 4% agar. The agar was cut around the blastocysts, and the piece of agar was embedded in paraffin and cut into serial sections (3-μm thick), which were mounted on glass slides. Sections were subjected to immunohistochemical (IHC) analysis as previously described [14]. Briefly, sections were dewaxed, rehydrated, washed in PBS, and blocked for 30 min with 0.1 mg/mL BSA, 2% goat serum, and 0.01% Tween-20 in PBS followed by incubation with primary antibodies in a blocking buffer overnight at 4°C and with secondary antibodies for 45 min at room temperature. Primary antibodies used were as follows: rabbit anti-thioredoxin (TRX, 1:200) (ab26320; Abcam, Cambridge, MA), rabbit anti-ZP3 (1:1,000) (sc25802; Santa Cruz Biotechnology, Santa Cruz, CA), Aurora A kinase (AURA, 1:50) (4718; Cell Signaling, Danvers, MA), and rabbit anti-β-Catenin (β-CAT, 1:100) (9582; Cell Signaling). Secondary antibody used was Alexa-fluor-488-conjugated goat anti-rabbit (1:500; Life Technologies). The nuclei were stained with 1.43 mM DAPI (Life Technologies) followed by mounting of the slides with the Prolong Gold mounting medium (Life Technologies). Differential interference-contrast microscopy and fluorescence microscopy analysis were carried out with an Olympus BX63 microscope. Images were acquired using an Olympus DP72 color CCD camera, and images were processed for publication in Adobe Photoshop version 6.0.
SDS PAGE and western blotting analysis
Twenty-nine human blastocysts were lysed in a lysis buffer containing 0.5% SDS with a complete protease inhibitor cocktail tablet (Santa Cruz Biotechnologies) and sonicated in a water bath for 3×1 min, followed by centrifugation at 18,000 g for 5 min. Cell lysates for SDS-PAGE, electrophoretic transfer of proteins to nitrocellulose, and immunoblotting were prepared as previously described [15]. Membranes were blocked in Tris-buffered saline with Tween-20 and 5% nonfat dry milk for 2 h at room temperature before incubation with primary antibodies overnight at 4°C, followed by incubation with secondary antibodies for 45 min at room temperature. Primary antibodies used were as follows: rabbit anti-TRX (1:50) and rabbit anti-AURA (1:50). Secondary antibody used was horse-radish peroxidase (HRP)-conjugated goat anti-rabbit IgG (DAKO, Glostrup, Denmark). The blot was developed with Luminata Forte Western HRP substrate
Results
The aim of the study was to examine the protein composition of human blastocoel fluid and the corresponding cells of the human blastocyst. We isolated blastocoel fluid by micromanipulation, and proteins in the fluid and cell material were identified by proteomic analysis. A total of 286 proteins were identified in the blastocoel fluid, while 1,307 proteins were identified in the corresponding cells of the blastocyst. Forty-four proteins were identified in the background medium, where 21 of these proteins were present in the blastocoel fluid and 25 proteins in the remaining cells (see Venn diagram in Fig. 2). Subtracting the proteins identified in the background medium, 265 proteins were identified in the blastocoel fluid and 1,282 proteins in the remaining cell material. Of the blastocoel proteins, 149 proteins were identified with a false-positive discovery rate of 1%, while the remaining 137 proteins were identified with a false-positive discovery rate of 5%. For the cell material, 729 proteins were identified with a false-positive discovery rate of 1% and 578 proteins with 5%. Nineteen proteins were identified specifically in the background medium without presence in the blastocoel or blastocyst cells.
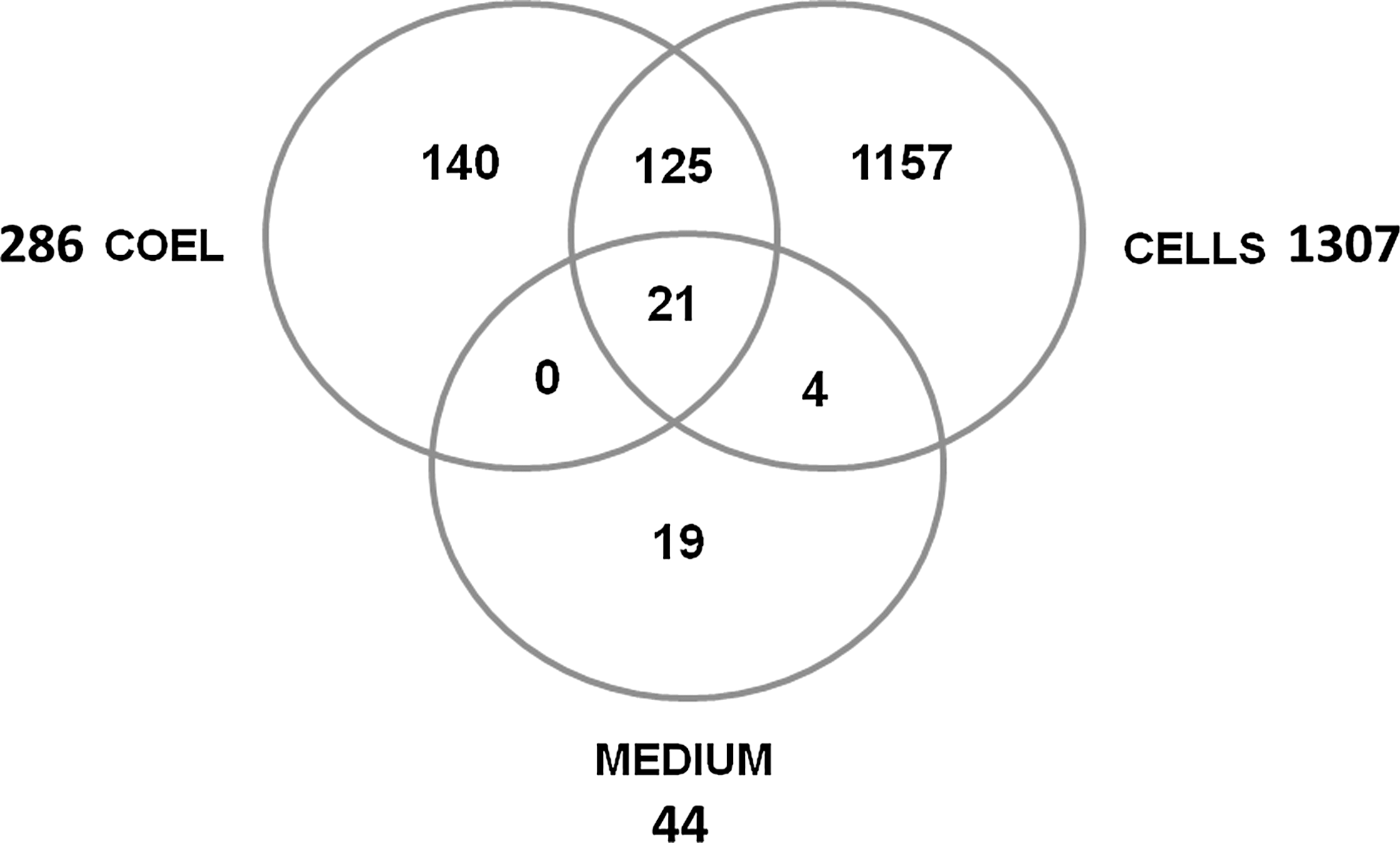
Venn diagram. The number of proteins identified in the blastocoel fluid (COEL), in the remaining cells of the blastocyst (CELLS), and in the background medium (MEDIUM). Figure not to scale.
In the blastocoel fluid, the protein amount spans over 4 orders of magnitude. Serum albumin is by far the most abundant protein; most of the other identified proteins are present in 3-to-4 order-of-magnitude lesser amount. In the blastocyst cell preparation, the dynamics are also 4 orders of magnitude, and serum albumin is also the most abundant protein. However, most of the identified proteins are only 2 orders of magnitude less than serum albumin (see Supplementary Fig. S1; Supplementary Data are available online at
The identified proteins were grouped according to their function in specific biological processes using GO terms provided by the bioinformatics tool ProteinCenter. The distribution of the proteins is presented in pie charts in Figs. 3 and 4. Proteins of potential interest in human reproduction and developmental biology or in culture conditions for hESCs or embryos after IVF were retrieved by using the GO terms cell proliferation, development, and reproduction. Selected proteins from these groups are presented in Tables 1 and 2. Microarray data on human blastocysts and hESCs from the HumEr database from Vassena et al. [16] are included in the tables whenever there is overlap in the identification of mRNA and protein.
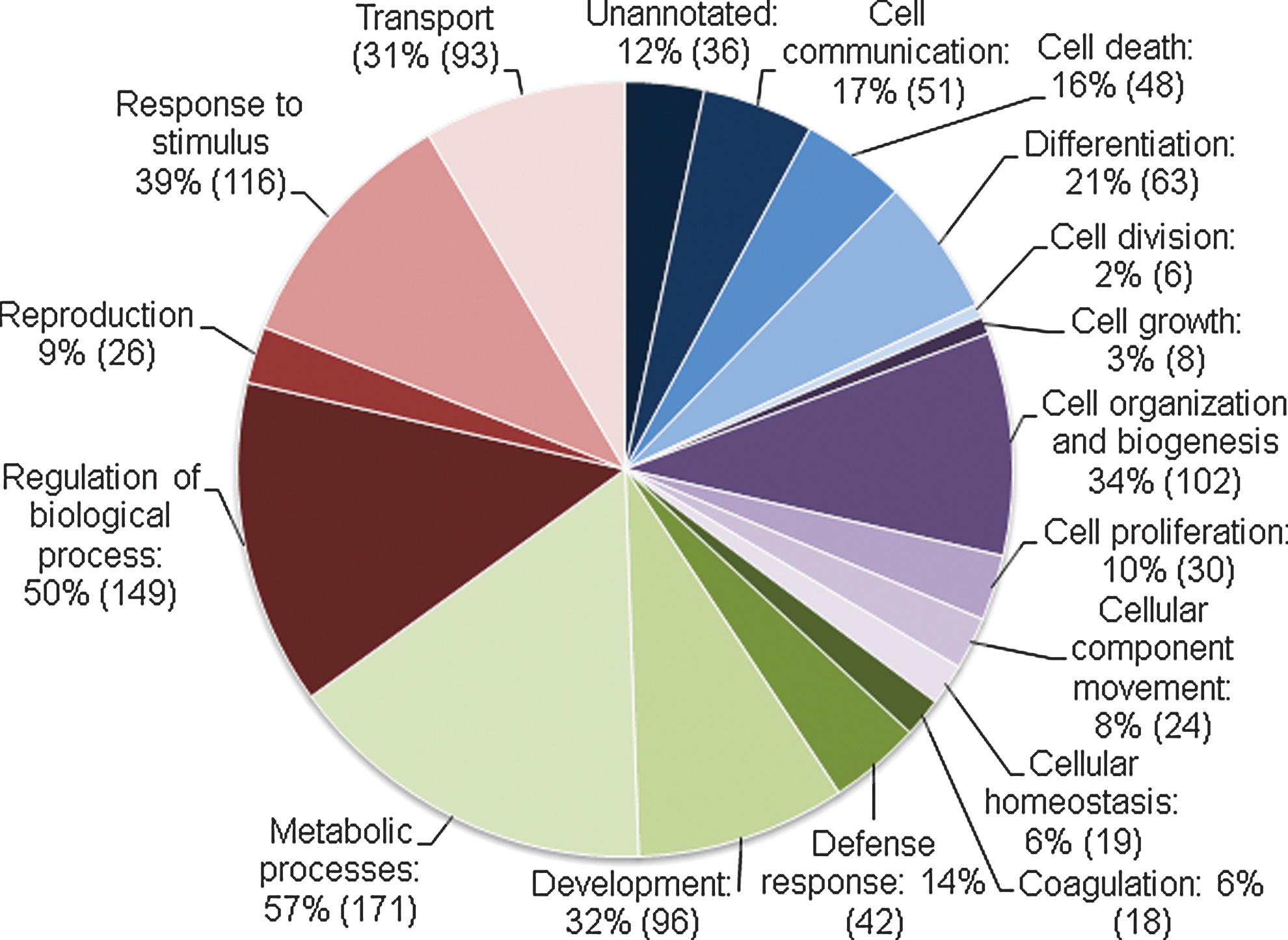
Biological processes in the blastocoel. Proteins identified in the blastocoel grouped according to the gene ontology terms of Biological Processes. Numbers in parentheses are the number of proteins in each group. The same protein can appear in more than one group.
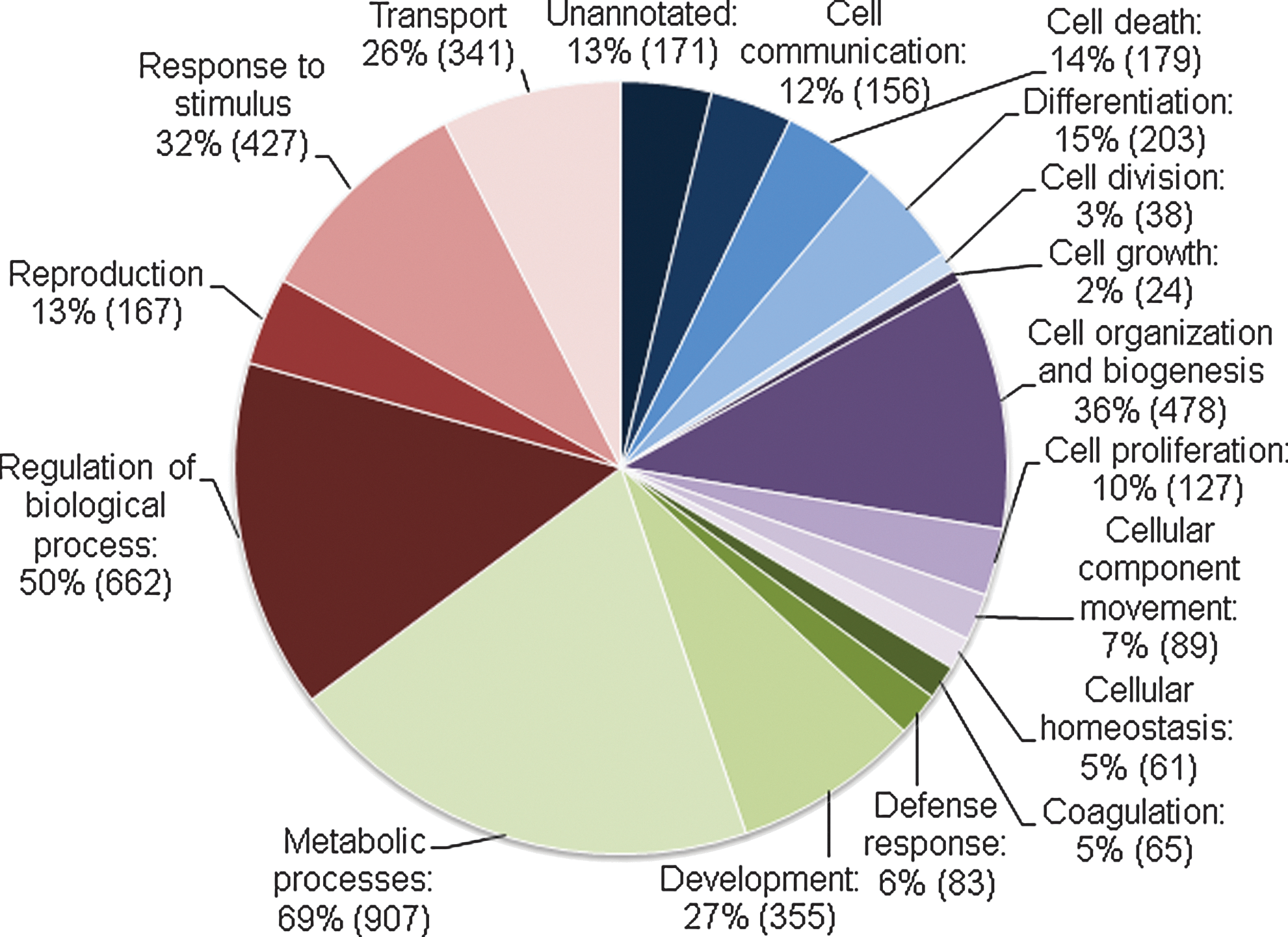
Biological processes in the remaining cells. Proteins identified in the remaining cells of the blastocyst grouped according to the gene ontology terms of Biological Processes. Numbers in parentheses are the number of proteins in each group. The same protein can appear in more than one group.
P value is the probability of false-positive identifications, estimated by searching MSMS spectra against the corresponding reversed-sequence (decoy) database.
Microarray data are from the database HumER [16] and the number states mRNA level in percent-rank expression. A percent-rank is defined as the rank of a value in a dataset as a percentage of the dataset. The probe with the highest intensity will get the rank 100%, whereas the probe with the lowest intensity will get the rank 0%.
hESC, human embryonic stem cells.
P value is the probability of false-positive identifications, estimated by searching MSMS spectra against the corresponding reversed-sequence (decoy) database.
Microarray data are from the database HumER [16], and the number states mRNA level in percent-rank expression. A percent rank is defined as the rank of a value in a dataset as a percentage of the dataset. The probe with the highest intensity will get the rank 100%, whereas the probe with the lowest intensity will get the rank 0%.
Several proteins known to be involved in assembly, maintenance, and function of mammalian cilia were also identified and primarily in the blastocyst cells. These included intraflagellar transport proteins (IFT74, IFT88, and IFT122) [17], kinesin-like proteins (KIF17 and KIF27) [17,18], axonemal dynein components [19], AURA [20], and Ellis-van Creveld syndrome protein [21 –23]. Bardet-Biedl syndrome protein 7 (BBS7), which critically regulates ciliary assembly through the BBsome [17], was also identified in the blastocoel fluid.
In addition, eight previously uncharacterized proteins were identified in the blastocoel fluid, and 34 uncharacterized proteins were identified in the remaining cells of the blastocyst. Full lists of proteins identified in the blastocoel fluid and blastocyst cells along with the proteins identified in the culture medium are to be found as Supplementary Data (Supplementary Tables S1–S3).
To validate the proteomic findings in our study, we used IHC and western blot (WB) analyses to confirm the expression of four of the identified proteins: AURA, TRX, ZP3, and β-CAT (Fig. 5). In IHC, AURA and β-CAT were visualized in the cytoplasm and in the nucleus, whereas TRX primarily was expressed in the cytoplasm. ZP3 clearly localized to the zona pellucida surrounding the blastocyst (Fig. 5A). In WB, we confirmed the expression of AURA and TRX in lysates of 29 human blastocysts and equivalent amounts of hESCs (LRB010) (Fig. 5B).
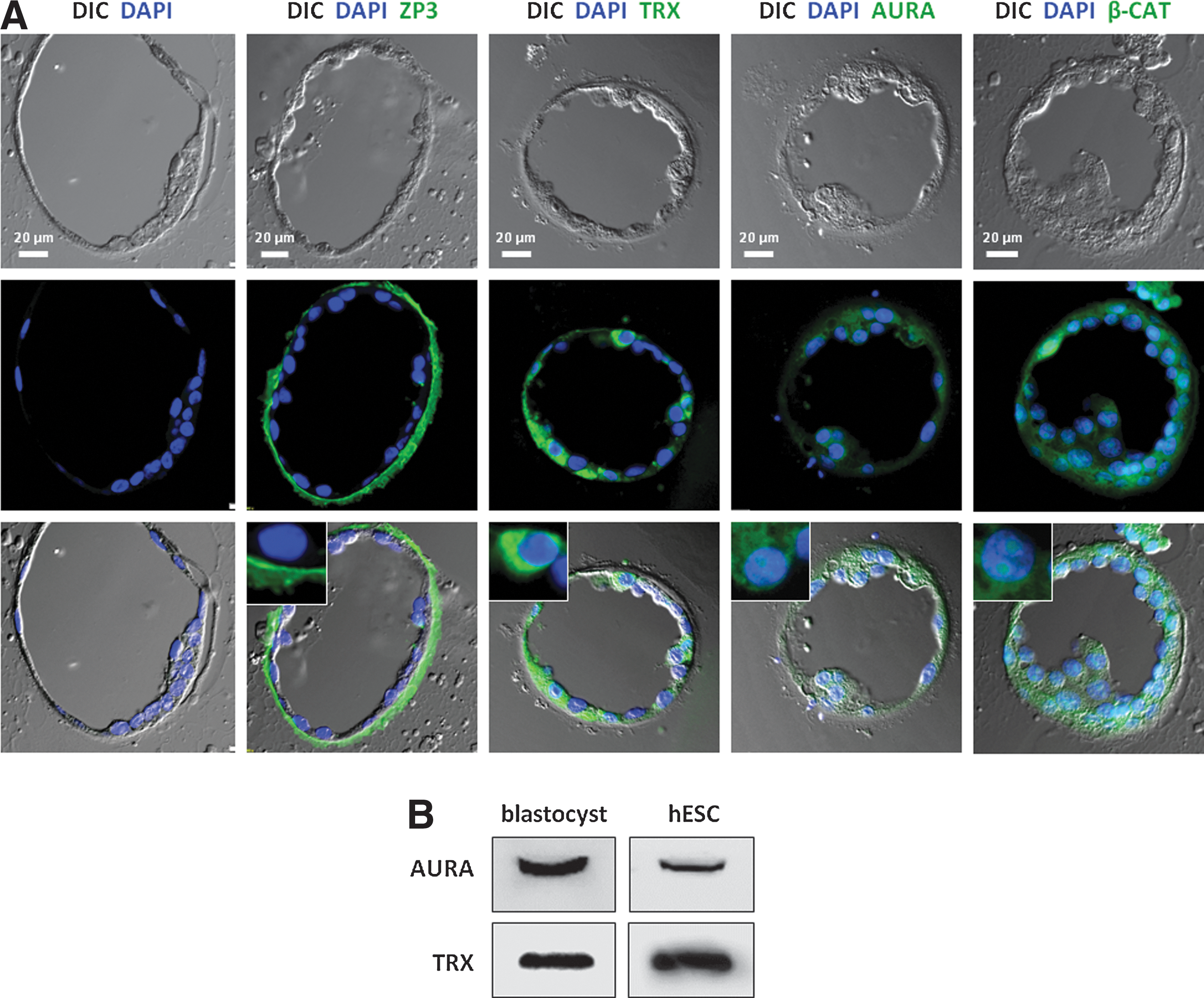
Validation of mass spec findings.
Discussion
This study is the first to identify specific proteins in the blastocoel fluid and corresponding cells of the human blastocyst; 286 proteins were identified in the fluid, and 1,307 proteins in the remaining cells of the blastocyst. Human blastocysts are scarce material, and the blastocoel fluid of 1–8 nL per blastocyst is difficult to retrieve, while the small volume and protein concentration are challenging to analyze. Our protein analysis of the blastocoel fluid and remaining cells from 53 human blastocysts, using the state-of-the-art-sensitive nano-LC tandem mass spectrometry, is therefore unique. We acknowledge that the protein lists are not an exhaustive description of the human blastocyst proteome, and the results are limited by the volume of blastocoel fluid available. The study could obviously be improved by collecting even more blastocoel fluid. However, this study represents a first attempt to uncover the protein composition of this very scarce human fluid and cell material.
The protein expression of four individual proteins could be validated by IHC and WB analyses. In addition, most of the proteins presented in Tables 1 and 2 have been identified at the gene expression level in human blastocysts in the microarray study on human preimplantation development and hESCs performed by Vassena et al. [16]. Most often, the mRNA was highly expressed (over 60% percent-rank expression), and all the proteins from our study were also identified at the gene expression level in hESCs [16]. This overlap in mRNA/protein identification is further supportive of our data.
Heat shock proteins
Several heat-shock proteins (HSPs) were expressed in both the blastocyst cells (heat-shock 70-kDa protein 1A/1B, heat-shock 70-kDa protein 4, heat-shock 70-kDa protein 4L, and HSP 75 kDa) and in both the blastocoel fluid and in the blastocyst cells (heat-shock cognate 71-kDa protein, heat-shock protein beta-1 (HSP27), 60-kDa HSP, heat-shock protein HSP 90-alpha and HSP 90-beta). HSPs are highly conserved proteins that act as molecular chaperones, assisting correct protein folding in addition to a variety of functions in cellular housekeeping. Some HSPs function especially during cellular stress from the environment, such as reactive oxygen species, high temperature, ethanol, UV light, and heavy metals [24]. In mouse models, it appears that HSPs are essential for successful preimplantation development, and it is believed that the specific expression pattern of HSPs may play both an essential role for differentiation and a protective role against apoptosis in the developing embryo [25]. HSPs have not to our knowledge been described in the human embryo before, but all of the mentioned HSPs, except for HSP 75 kDa, are also expressed in hESCs, induced pluripotent stem cells [26], and in porcine zygotes [27].
Zona pellucida proteins
Zona pellucida (ZP) proteins were detected in both the blastocoel fluid (ZP2–4) and in the remaining cells of the blastocyst (ZP1–4), and ZP3 surrounding the blastocyst was visualized in IHC (Fig. 5A). The ZP consists of 4 glycoproteins, ZP1–4, which are important for the survival of growing oocytes, successful and species-specific fertilization, and the passage of the early embryo through the fallopian tube [28]. Knockout of the genes encoding ZP glycoproteins in mice adversely affects the developmental competence of the eggs [29]. Throughout the preimplantation period of development, ZP is gradually degraded before hatching and just before implantation of the blastocyst in the endometrium. It has been speculated that ZP glycoproteins are enzymatically digested by peptidases released from cortical granules after egg activation. These degenerated products are thought to be released from the developing embryos in the fallopian tube with a function in the transmission of information about the developing embryo to the immune system in the female reproductive tract [28].
It is expected to find ZP proteins in the blastocyst cell sample, since the blastocysts were sampled before hatching from the ZP. However, it is surprising to identify 3 out of 4 ZP proteins in the blastocoel fluid. It is possible that the ZP proteins are brought in to the blastocoel with the micropipette during the micromanipulation procedure and thus is an artifact of contamination. Conversely, it cannot be excluded that the ZP proteins are present in the blastocoel fluid carrying out yet unknown functions. The identified peptides from the MS analysis could also be degenerated products of ZP digestion as described above. In addition, microarray data from Vassena et al. [16] show expression of ZP1-4 mRNA in hESCs, which underlines that the ZP proteins can carry out functions other than those previously described.
Vitamins and vitamin-binding proteins
Retinoic acid (RA) has a known function as a morphogen in the early mammalian embryonic development, where both deficiency and excess doses can cause developmental abnormalities [30]. We identified Retinol-binding protein 4 (RBP4) in both the human blastocoel fluid and the remaining cells of the blastocyst, and it may be speculated that this protein secures low levels of RA to prevent premature differentiation of the blastocyst cells. RBP4 has also been identified in the yolk syncytial layer of zebra fish early embryos, where knockdown of the protein resulted in shortened yolk extension and impaired liver development [31]. In addition, RA-induced protein 1 was identified in the remaining cells of the blastocyst. The function of this protein is largely unknown besides the fact that deletion of the gene causes Smith-Magenis syndrome, a neurobehavioral disorder with a variety of symptoms, including obesity, and craniofacial and skeletal abnormalities [32].
It is not yet clear whether vitamin D exerts an important role in embryonic development. While an early study showed that the differentiation of cancer cells is induced by vitamin D [33], later studies conclude that vitamin D deficiency does not prohibit development, nor does vitamin D receptor knockout, so the current understanding is that vitamin D is not a major developmental hormone [34]. We identified vitamin-D-binding protein in the blastocoel fluid of the human blastocyst and vitamin D 25-hydroxylase (activates vitamin D by hydroxylation) in the remaining cells of the blastocyst. Thus, the function of these proteins in the developing embryo remains to be discovered.
Ciliary proteins
Cilia protrude from the cell membrane into the extracellular environment in most eukaryotic cells [35]. In particular, primary cilia are formed as solitary organelles from most quiescent mammalian cell types and coordinate a variety of signaling pathways, including Hedgehog (Hh), Wnt, and Receptor Tyrosine Kinase signaling, which control cellular processes during embryonic development [36 –38]. Consequently, defects in the assembly or function of these cilia leads to a plethora of developmental disorders and diseases, now commonly referred as to ciliopathies [39,40]. Previously, we have used electron microscopy and immunofluorescence microscopy to show that hESCs form primary cilia that coordinate Hh signaling [41]. It is therefore not surprising to find ciliary proteins in the cells of the human blastocyst, such as Ellis-van Creveld syndrome protein, which controls Hh signaling via the primary cilium [21 –23], and when mutated causes numerous developmental disorders, including congenital heart defects and skeletal dysplasia [42]. However, this is the first time that ciliary proteins have been identified in the blastocyst, and our data confirm that the human embryo at this early stage indeed possess primary cilia, which may play a critical role during early development. BBS proteins form a protein complex required for ciliary assembly, and when mutated causes a series of developmental disorders [40]. BBS7 mRNA is expressed in hESCs [16], and we here show that BBS7 is present in the blastocoel fluid. Although speculative at this point, these data are indicative of a role of BBS7 in the blastocoel fluid in maintenance of primary cilia of the blastocyst.
Uncharacterized proteins
Several previously uncharacterized proteins were identified both in the blastocoel fluid and in the remaining cells of the blastocyst, and only five out of 42 uncharacterized proteins were not supported by microarray data from Vassena et al. [16]. The uncharacterized proteins are proteins that have been identified in other protein studies, for example, general human proteome analyses, but have not yet been assigned a function.
Technical considerations
The human blastocysts were generated by IVF using slightly different protocols at two different laboratories. Since the proteins of the human blastocyst have not been examined before, it is difficult to say how much the culture conditions contribute to the specific protein composition of the blastocyst.
In proteomics studies with unlimited cell material, it is possible to identify more than 4,000 proteins in single experiments [43]. Therefore, it is clear that the limitation of sample material in our case consequently leads to a limitation of the number of proteins we were able to identify.
The analysis of the culture medium for the blastocysts identified 44 proteins in the culture medium. These proteins were extracted from the presented datasets of the blastocoel and remaining blastocyst cells. However, it cannot be excluded that some proteins present in the culture medium also could be present in the blastocyst in vivo. The only protein source in the culture medium is human serum albumin and insulin as stated by the companies, so the remaining proteins are most probably impurities copurified with the serum albumin.
One hundred forty-six proteins were identified in both the blastocoel fluid and the remaining cells. This overlap can be due to contamination of the blastocoel sample with the cell material under the micromanipulation procedure, but it is also reasonable to believe that some proteins are to be found in both blastocoel fluid and blastocyst cells. Nineteen proteins were specifically identified in the background sample, but were not present in the blastocoel or remaining cells, indicating that the contamination from the background medium was minor.
A number of keratins were identified both in the blastocoel and remaining cell material. It is well known that keratins can be the source of contamination during proteomic sample preparation. Therefore, we have excluded from our data all keratins listed in the cRAP database (common Repository of Adventitious Proteins) [10] together with proteins with expected expression merely in hair or skin as judged by data from the Uniprot database [11]. Remaining keratins are kept and presented in the dataset (Supplementary Tables S1–S3), because we find it likely that some of the keratins are present in our blastocoel and cell material. In the humER database from the study of Vassena et al. [16], mRNA from 51 specific keratins were detected in human preimplantation embryos, and several other studies can confirm the presence of keratins and cytokeratins in the early mammalian embryonic development [44 –47].
Today, many fertility clinics routinely remove the blastocoel fluid of human blastocysts in an attempt to enhance survival after cryopreservation [48 –50]. Usually, the fluid is discarded, but can instead be a source of future knowledge about the earliest developmental conditions in the blastocyst and also an inspiration for the further development of culture conditions for both human embryos and hESCs.
Concluding Remarks
In this study, we identified 286 proteins in the human blastocoel fluid and 1,307 proteins in the remaining cells of the human blastocyst. Of these proteins, 42 were previously uncharacterized proteins. Several ciliary proteins were identified as well, supporting previous findings that the human blastocysts possess primary cilia. These results contribute new and important knowledge about the early human development and can be an inspiration for the development of safe and efficient culture conditions for human embryos and hESCs. Nonetheless, we encourage more studies on the blastocoel fluid, which is routinely derived in the fertility clinics to obtain more detailed knowledge of these earliest stages of human development.
Footnotes
Acknowledgments
We thank Dr. Poul M. Hyttel and M.D. Tonny S. Petersen for technical assistance with the blastocyst sections and IHC procedure. Iben R. Veland, MSc, is thanked for technical assistance with the WB procedure, and Dr. Henriette Pilegaard is thanked for kindly providing access to her CareStream Image Station. This work was funded by the Lundbeck Foundation (R54-A5642; STC), the Novo Nordic Foundation (STC), and the Danish Agency for Science, Technology and Innovation, Danish Ministry of Science, Innovation, and Higher Education (PLJ).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.