
Abstract
Corneal endothelial dysfunction remains a major indication for corneal transplantation. Both corneal endothelial cells and stromal cells originate from the neural crest, but have distinct phenotypes and function in the adult cornea. We previously reported that stem cells isolated from the adult corneal stroma [cornea-derived precursors (COPs)] show characteristics of multipotent neural crest-derived stem cells. In this study, we report the induction of functional tissue-engineered corneal endothelium (TECE) from mouse and human COPs. TECE was engineered from Wnt1-Cre/Floxed EGFP mouse COPs in a medium containing retinoic acid and glycogen synthase kinase (GSK) 3β inhibitor (activator of Wnt/β-catenin signaling). The expression levels of major markers characterizing corneal endothelial function (Atp1a1, Slc4a4, Car2, Col4a2, Col8a2, and Cdh2) were significantly upregulated. Both retinoic acid and GSK 3β inhibitor upregulated the expression of Pitx2, a homeobox gene involved in the development of the anterior segment of the eye. GSK 3β inhibitor increased Atp1a1 expression and Na,K-ATPase pump activity of TECE, which was significantly higher than COPs or control 3T3 cells, and 2.6-fold higher than cultured mouse corneal endothelial cells. Mouse TECE transplanted into rabbit corneas maintained transparency and corneal thickness, whereas control corneas without TECE showed marked edema and increased corneal thickness. Furthermore, we successfully induced TECE from human COPs, and human TECE transplanted into rabbit corneas also maintained corneal transparency and thickness. This protocol enables efficient production of corneal endothelium from corneal stromal stem cells by direct induction, which may lead to a novel stem cell therapy for corneal endothelial dysfunction.
Introduction
The corneal endothelium consists of a single layer of hexagonal cells with a thick basement membrane (Descemet's membrane) covering the posterior surface of the cornea in a well-arranged mosaic pattern [8]. Corneal hydration is determined primarily by the balance between the movement of aqueous humor across the corneal endothelium into the stroma and the subsequent pumping of the fluid out from the stroma [8]. The accumulation of fluid in the stroma due to disturbance of this balance may lead to bullous keratopathy, which is characterized by an edematous cornea with a reduced transparency. The Na+- and K+-dependent ATPase (Na,K-ATPase) expressed in the basolateral membrane of corneal endothelial cells is primarily responsible for the pump function of the corneal endothelium [8]. Given that human corneal endothelial cells have a limited proliferative capacity, endothelial dystrophies, ocular trauma, corneal graft rejection, as well as insults associated with intraocular surgeries may result in corneal endothelial cell loss and permanent damage.
Corneal endothelium, as well as corneal stromal cells (keratocytes), originates from the cranial neural crest [9], and expression of the homeobox gene Pitx2 is required for ocular anterior segment development [10]. We previously reported the isolation of cornea-derived precursors (COPs) from the mouse corneal stroma, which have characteristics of multipotent neural crest-derived stem cells [11,12]. COPs initiate spheres by clonal expansion from a single cell, and express the neural crest markers Twist, Snail, Slug, and Sox9 [11]. Since both corneal endothelium and corneal stroma originate from the neural crest [9], we hypothesized that COPs can be made to differentiate into corneal endothelial cells by mimicking developmental process. Signaling crosstalk of retinoic acid and Wnt/β-catenin signaling was reported as a key regulators of development in the anterior segment of the eye [13,14]; therefore, retinoic acid and activation of Wnt/β-catenin signaling may be used for differentiation. Herein, we report how tissue-engineered corneal endothelium (TECE) derived from mouse COPs express markers associated with corneal endothelial cells, and show Na,K-ATPase pump function equivalent to cultured mouse corneal endothelial cells in vitro. We also show how TECE maintains corneal transparency and thickness in vivo using a rabbit corneal transplantation model. Finally, we show human TECE can be obtained from human corneal stroma by applying this protocol, which also functions to maintain corneal transparency and thickness in vivo.
Materials and Methods
Cell culture
For isolation of mouse COPs, cells were dissociated from adult Wnt1-Cre/Floxed EGFP mouse corneal stroma, and then cultured as described previously [11,12,15]. All animals were handled in full accordance with the Association for Research in Vision and Ophthalmology (ARVO) Statement for the Use of Animals in Ophthalmic and Vision Research. In brief, Descemet's membrane together with corneal endothelial cells was stripped off en bloc from the cornea by forceps. Next, the cornea was incubated in 2 mg/mL of dispase II (Roche Applied Science) for 60 min at 37°C, and corneal epithelial cells were totally stripped off from the stroma. After washing 3 times with phosphate-buffered saline (PBS) to avoid the contamination of endothelial or epithelial cells, corneal stromal discs were cut into small pieces and digested in 0.05% trypsin (Sigma-Aldrich) for 30 min at 37°C, followed by 78 U/mL collagenase (Sigma-Aldrich) and 38 U/mL hyaluronidase (Sigma-Aldrich) treatment for 30 min at 37°C. Stromal cells were mechanically dissociated into single cells and cultured in the Dulbecco's modified Eagle's medium (DMEM)/F-12 (1:1) supplemented with 20 ng/mL epidermal growth factor (EGF; Sigma-Aldrich), 10 ng/mL fibroblastic growth factor 2 (FGF2; Sigma-Aldrich), B27 supplement (Invitrogen), and 10 ng/mL leukemia inhibitory factor (Chemicon International) at a density of 1×105 cells per milliliter under a humidified atmosphere of 5% CO2 at 37°C. Initial culture was performed in 35-mm dishes and then subcultured to 25-cm2 culture flasks. The spheres were then further subcultured in 75-cm2 culture flasks after 14 to 21 days, which was repeated every 14 to 21 days. The medium was changed every 5 to 7 days. All dishes and flasks used for sphere culture were polystyrene, noncoated vessels obtained from Asahi Techno Glass. After the first passage, EGFP-positive sphere rates of COPs derived from Wnt1-Cre/Floxed EGFP mouse were counted in 4 independent view fields of 100 magnifications.
Human corneal stroma for isolation of human COPs were obtained from the U.S. eye bank corneas after the central buttons were used for corneal transplantation. After stripping endothelial and epithelial cells off by same method, corneal stromal tissue was cut into small pieces and digested in 400 U/mL type I collagenase (Sigma-Aldrich) for overnight at 37°C. Stromal cells were mechanically dissociated into single cells and washed by PBS 2 times, and cultured in DMEM/F-12 (1:1) supplemented with 20 ng/mL EGF, 20 ng/mL FGF2, B27 supplement, N2 supplement (R & D systems) 2 mM
In vitro differentiation
To examine endothelial cell differentiation, mouse COPs were dissociated into single cells and suspended at a cell density of 2×105 cells/cm2. Clonal COPs were plated and cultured on 0.1% gelatin-coated 35-mm dishes or 0.1% gelatin-coated polycarbonate Snapwell inserts (Corning) with a membrane pore size of 0.4 μm for Ussing Chamber study. For transplantation study, type I atelo collagen sheets (CM24; Koken) coated with 0.1% gelatin were used as carriers, and dissociated COPs were labeled with the fluorescent tracker, PKH26 (SIGMA), before plating on carrier sheets to observe cell localization after transplantation. The cells were cultured for 1 week under a humidified atmosphere of 5% CO2 at 37°C in the specific endothelium-inducing medium (SEIM), which was based on Eagle's minimum essential medium (MEM; Sigma-Aldrich) and supplemented with 1.0% fetal bovine serum (Sigma-Aldrich), all-trans retinoic acid (1 μM; Sigma-Aldrich), 6-bromoindirubin-3′-oxime [BIO; glycogen synthase kinase (GSK) 3β inhibitor] (1 μM; Calbiochem), transforming growth factor (TGF) β2 (5 ng/mL; Peprtotech), Y-27632 (10 μM; nacalai tesque), insulin (1 μM; Sigma-Aldrich), CaCl2 (1 mM; Sigma-Aldrich), sodium pyruvate (1 mM; Sigma-Aldrich), penicillin (100 U/mL; Sigma-Aldrich), streptomycin (100 μg/mL; Sigma-Aldrich), MEM amino acid (1×; Sigma-Aldrich), MEM essential vitamin mixture (1×; Bio-Whittaker). After the first week, cells were maintained with the MEM supplemented with CaCl2 (1 mM), sodium pyruvate (1 mM), penicillin (100 U/mL), streptomycin (100 μg/mL), MEM amino acid (1×), and MEM essential vitamin mixture (1×).
For endothelial cell differentiation from human COPs, human COPs were dissociated into single cells, labeled with PKH26, and suspended at a cell density of 2×105 cells/cm2. Clonal COPs were plated and cultured on 0.1% gelatin and 1.0 μg/mL laminin-coated 35-mm dishes or similarly coated type I atelo collagen sheets (Koken). The cells were cultured for 1 week under a humidified atmosphere of 5% CO2 at 37°C in the SEIM.
Immunocytochemistry
Corneal endothelial cells and TECE cultured on the 3.5-mm culture dish were fixed at room temperature for 10 min in 4% formaldehyde in PBS. After 3-time PBS washes, the specimens were incubated for 30 min in Morphosave (Ventana Medical Systems). After 2-time PBS washes, the specimens were incubated for 30 min in 10% normal donkey serum to block nonspecific binding. This was followed by overnight incubation at 4°C with 1:200 diluted rat anti-ZO-1 antibody (Santa Cruz Biotechnology; final concentration 1 μg/mL) or rabbit anti-Na,K-ATPase α-subunit antibody (Epitomics), and 3-time washes in PBS. The cells were then incubated for 1 h in a 1:200 dilution of Cy3-conjugated donkey anti-rat IgG antibody (Jackson ImmunoResearch; final concentration 30 μg/mL) or anti-rabbit IgG antibody (Jackson ImmunoResearch) and again washed 3 times in the dark. The specimens were mounted on dishes with an anti-fading mounting medium containing 4′,6-diamino-2-phenylindole (DAPI), and the slides were inspected with the microscope (Axio Imager; Carl Zeiss, Inc.).
RT-PCR analysis and quantitative RT-PCR analysis
Mouse corneal epithelium, stroma, endothelium, COPs, and TECE derived from COPs were used for reverse transcription–polymerase chain reaction (RT-PCR) analysis. Epithelium was mechanically removed from the eye by forceps and collected before enucleating the eyeball. Next, eyeballs were enucleated and washed with PBS to avoid corneal epithelial cell contamination, and corneas were excised. Descemets' membrane together with corneal endothelial cells was stripped off en bloc from the cornea by forceps, and residual corneal stroma was washed with PBS to avoid corneal endothelial cell contamination. Total RNA was isolated from mouse corneal epithelium, endothelium, COPs, and TECE with the use of an RNeasy kit (Qiagen). Corneal stroma was homogenized in 1.2 mL of Trizol Reagent (Molecular Research Center). About 0.24 mL chloroform was added in the reagent and centrifuged 15,000 g for 15 min at 4°C for phase separation. The aqueous phase was mixed with 0.6 mL isopropyl alcohol and centrifuged 15,000 g for 15 min at 4°C for RNA precipitation. After washing by 75% ethanol, total RNA from corneal stroma was dissolved in RNase-free water. The RNA from corneal epithelium, stroma, endothelium, COPs, and TECE was subjected to RT-PCR analysis with a ReverTra Ace α kit (Toyobo). The PCR protocol was designed to maintain amplification in the exponential phase. The sequences of the PCR primers are in Supplementary Tables S1 and S2 (Supplementary Data are available online at
For quantitative RT-PCR analysis, RNA was extracted from TECE, COPs before derivation, COPs cultured with MEM supplemented with 1.0% fetal bovine serum, and either 1 μM retinoic acid or 1 μM BIO. Then, cDNA was synthesized by using an RNeasy kit. Quantitative RT-PCR analysis was performed by SYBR Green enzyme mixture (Thunderbird SYBR qPCR mix; Toyobo) and a thermal cycler (Step One Plus; Applied Biosystems). Primer pairs are listed in Supplementary Table S3.
Western blot analysis
Mouse COPs and TECE were washed with PBS twice, and dissolved in a lysis buffer (M-PER; Thermo Fisher Scientific) with a protein inhibitor cocktail (Thermo Fisher). Western blot analysis was performed by using a standard protocol with primary antibodies for β-actin (rabbit; Abcam), Na,K-ATPase α-subunit (Epitomics), and Pitx2 (Aviva Systems Biology). Chemiluminescence image was analyzed by using a CCD camera system (ImageQuant LAS 4000; GE Healthcare).
Measurement of Na,K ATPase pump function
The pump function of confluent monolayers of various type of cells was measured with the use of an Ussing chamber basically as described previously [16 –18]. TECE were cultured under the condition mentioned in the In Vitro Differentiation section, and other cells (mouse corneal endothelial cells, 3T3 cells, and COPs before derivation) were cultured under a humidified atmosphere of 5% CO2 at 37°C in Eagle's minimum essential medium (Sigma-Aldrich) supplemented with 10% fetal bovine serum (Sigma-Aldrich), CaCl2 (1 mM; Sigma-Aldrich), sodium pyruvate (1 mM; Sigma-Aldrich), penicillin (100 U/mL; Sigma-Aldrich), streptomycin (100 μg/mL; Sigma-Aldrich), and MEM amino acid (1×) (Sigma-Aldrich), MEM essential vitamin mixture (1×) (Bio-Whittaker) on 0.1% gelatin-coated Snapwell inserts until the cells became confluent. For positive control, the human corneal endothelial cell line B4G12 cells were purchased from DSMZ and cultured in Human-Endothelial-SFM (Invitrogen) supplemented with 1.0% fetal bovine serum (Sigma-Aldrich) and 10 ng/mL FGF2 on 0.1% gelatin-coated polycarbonate Snapwell inserts. To standardize the measurement condition, all media were changed to the MEM supplemented with 1.0% fetal bovine serum, CaCl2 (1 mM), sodium pyruvate (1 mM), penicillin (100 U/mL), streptomycin (100 μg/mL), MEM amino acid (1×), and MEM essential vitamin mixture (1×) after each cell became confluent, and each cell was cultured additional 48 h. The cells cultured on Snapwell inserts were placed in an Ussing chamber EM-CSYS-2 (Physiologic Instruments). The endothelial cell surface side was in contact with one chamber, and the Snapwell membrane side was in contact with another chamber. The chambers were carefully filled with Krebs-Ringer bicarbonate (120.7 mM NaCl, 24 mM NaHCO3, 4.6 mM KCl, 0.5 mM MgCl2, 0.7 mM Na2HPO4, 1.5 mM NaH2PO4, and 10 mM glucose bubbled with a mixture of 5% CO2, 7% O2, and 88% N2 to pH 7.4). The chambers were maintained at 37°C by an attached heater. The short-circuit current (SCC) was sensed by narrow polyethylene tubes positioned close to either side of the Snapwell, filled with 3 M KCl and 4% agar gel, and connected to silver electrodes. These electrodes were connected to a computer through an Ussing system EVC4000 (Physiologic Instruments) and an iWorx IX/408 data acquisition system (iWorx Systems), and the SCC was recorded by Labscribe 2 Software for Research (iWorx Systems). After the SCC had reached a steady state, the specific Na,K-ATPase inhibitor ouabain (final concentration, 10 mM; Sigma-Aldrich) was added to the chamber, and the SCC was measured again. The SCC attributable to Na,K-ATPase activity was calculated as the difference in SCC measured before and after the addition of ouabain.
Transplantation of TECE sheets into a rabbit model
All animals were treated in accordance with experimental procedures approved by the Committee for Animal Research of Keio University School of Medicine and Tokyo Dental College Ichikawa General Hospital. Before transplantation, cell densities of TECE sheets were manually counted by a microscope. Japanese white rabbits (female, 2.5 kg body weight; Shiraishi Experimental Animal Breeding Farm) were anesthetized intravenously with a mixture of diazepam (0.5 mg/kg; Takeda pharmaceutical) and pentobarbital sodium (30 mg/kg; Kyoritsu Seiyaku). Transplantation was performed on the left eye of each animal only.
Corneal buttons were prepared by 8.0-mm Barron donor cornea punch (Barron Precision Instruments) from the center of donor corneas purchased from Funakoshi Co., Ltd., and Descemet's membrane along with the entire endothelium was stripped from the corneal buttons. PKH26-labeled TECE cell sheets with carrier collagen sheets were harvested by same size punch and immediately placed on the stromal bed. Within 5 min, the sheets produced stable attachment to the stroma. The recipient central cornea was incised and opened by Hassburg-Barron Vacuum Trephine (Barron Precision Instruments), and the corneal buttons with TECE cell sheets or the control corneal buttons with neither TECE cell sheets nor endothelium were then placed on the graft bed of the originally surgically operated eye and sutured with 16 interrupted sutures (10–0 nylon). Antibiotics (0.3% ofloxacin) and steroids (0.1% betamethasone) were applied topically 3 times a day. After transplantation, eyes were carefully observed by slit-lamp microscopy, and serial photographs were obtained. Central corneal thickness was measured with an ultrasound pachymeter (Tomey), and intraocular pressure (IOP) was measured by Accupen (White Medical) on 1, 2, 5, and 8 days after transplantation. Central corneal thickness measurement and IOP measurement were performed by 2 different investigators who were masked to the details of transplantation in each group. Six eyes from 6 individual rabbits were used for each experimental group. Finally, 8 days after surgery, rabbits were sacrificed, and the operated eyes were enucleated, and the corneas of each operated eye were cut and paraffin-embedded, and mounted on dishes or glass slides with an antifading mounting medium containing DAPI. Localization of transplanted cells labeled by PKH26 was observed by the Axio Imager.
Statistical analysis
Data are presented as means±SD and were compared by Student's t-test or multiple t-test with Bonferroni correction after ANOVA with the use of Excel 2007 software (Microsoft). A P value of<0.05 was considered statistically significant.
Results
Morphology of TECE
In our original report, COPs were maintained in sphere cultures (Fig. 1A) [11,12]. COPs derived from Wnt1-Cre/Floxed EGFP mouse expressed EGFP fluorescence, indicating the neural crest origin of these cells (Fig. 1B) [15]. After the first passage, EGFP-positive spheres/total number of COP spheres in 4 independent fields of view were 45/45, 34/34, 28/28, and 20/20, yielding a mean EGFP-positive sphere rate of 100%. When COPs had been cultured in a DMEM/F12 medium without any other supplements, cells differentiated into keratocytes with dendritic morphology forming a meshwork type of cellular network (Fig. 1C). However, when COPs were cultured in a specific endothelium-derived medium for a week (see the Methods section), the cells formed a monolayer with a mosaic pattern (Fig. 1D). TECE maintained a similar monolayer mosaic pattern on collagen sheets as well (Fig. 1E). These tissue-engineered corneal endothelial cells (TECE) were similar to cultured mouse corneal endothelial cells in the cell size, cell shape, nuclear size, and nuclear cytoplasm ratio (Fig. 1F). Immunocytochemistry of ZO-1 also revealed that both TECE derived from Wnt1-Cre/Floxed EGFP mouse COPs and cultured Wnt1-Cre/Floxed EGFP mouse corneal endothelial cells expressed the tight junction maker, ZO-1, and formed a similar mosaic pattern (Fig. 1G, H). Na,K-ATPase α−subunit, a component of the corneal endothelial pump, was also expressed on the lateral side of all cells (Fig. 1I). Both TECE and corneal endothelial cells also expressed EGFP fluorescence, suggesting that these cells were of neural crest origin. After that, the monolayer mosaic pattern of the cells with ZO-1 expression was maintained for at least additional 2 weeks (Fig. 1J).
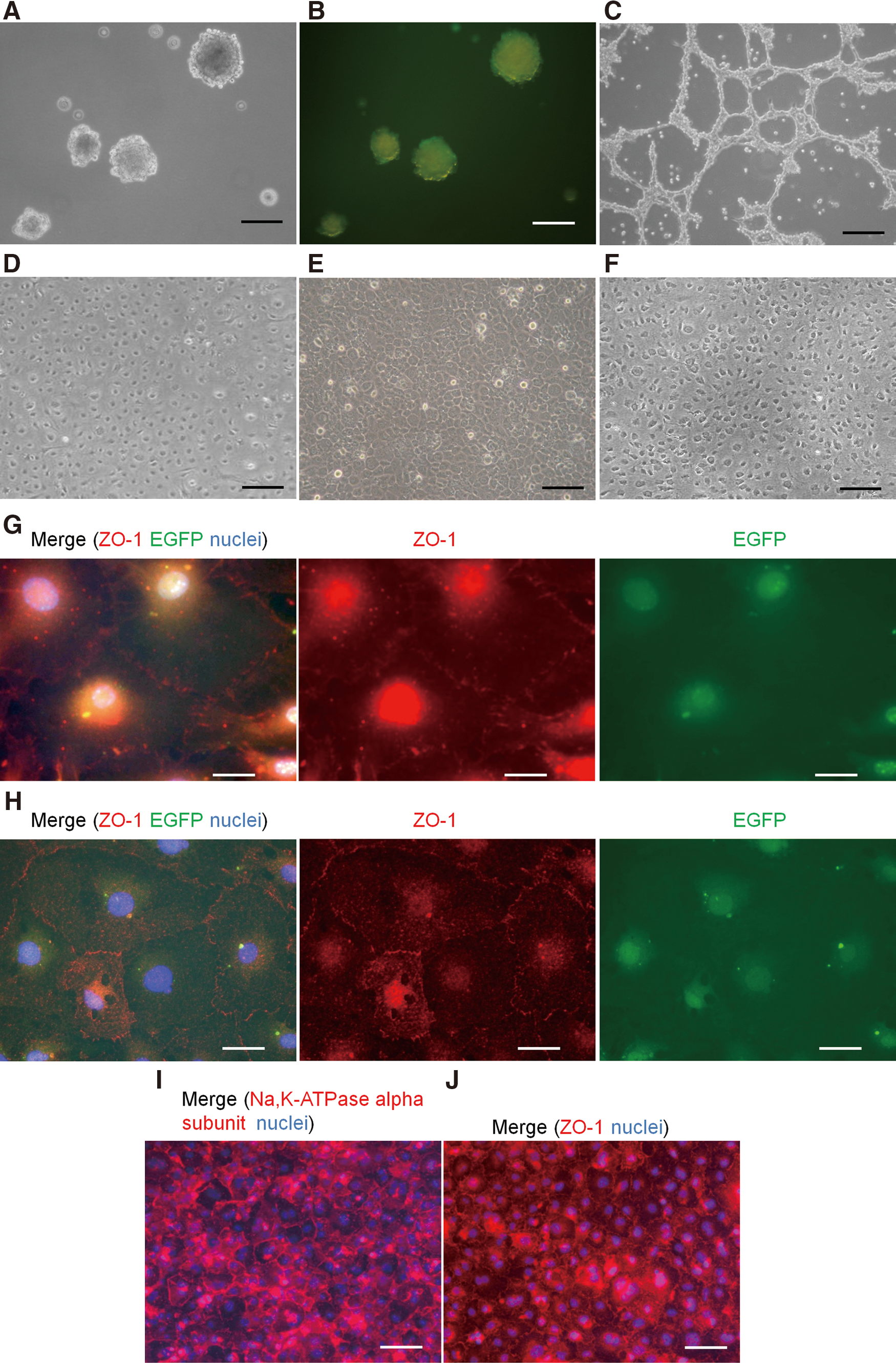
Morphology of tissue-engineered corneal endothelium (TECE). COPs derived from Wnt1-Cre/Floxed EGFP mouse were maintained in sphere culture
Changes in expression of corneal endothelium markers, Pitx2 and Abcg2
We selected Atp1a1 (Na,K-ATPase α-subunit), Slc4a4 (Na,HCO3 co-transporter), Car2 (carbonic anhydrase type 2), Col4a2 (collagen type IV), Col8a2 (collagen type VIII), and Cdh2 (N-cadherin) as corneal endothelium markers. Figure 2A shows the expression of these markers in corneal epithelium, stroma, and endothelium by RT-PCR. Some, but not all, markers were also expressed in epithelium or stroma, but only the endothelium expressed all markers. Among these, Col4a2 and Col8a2 were expressed specifically in the endothelium.
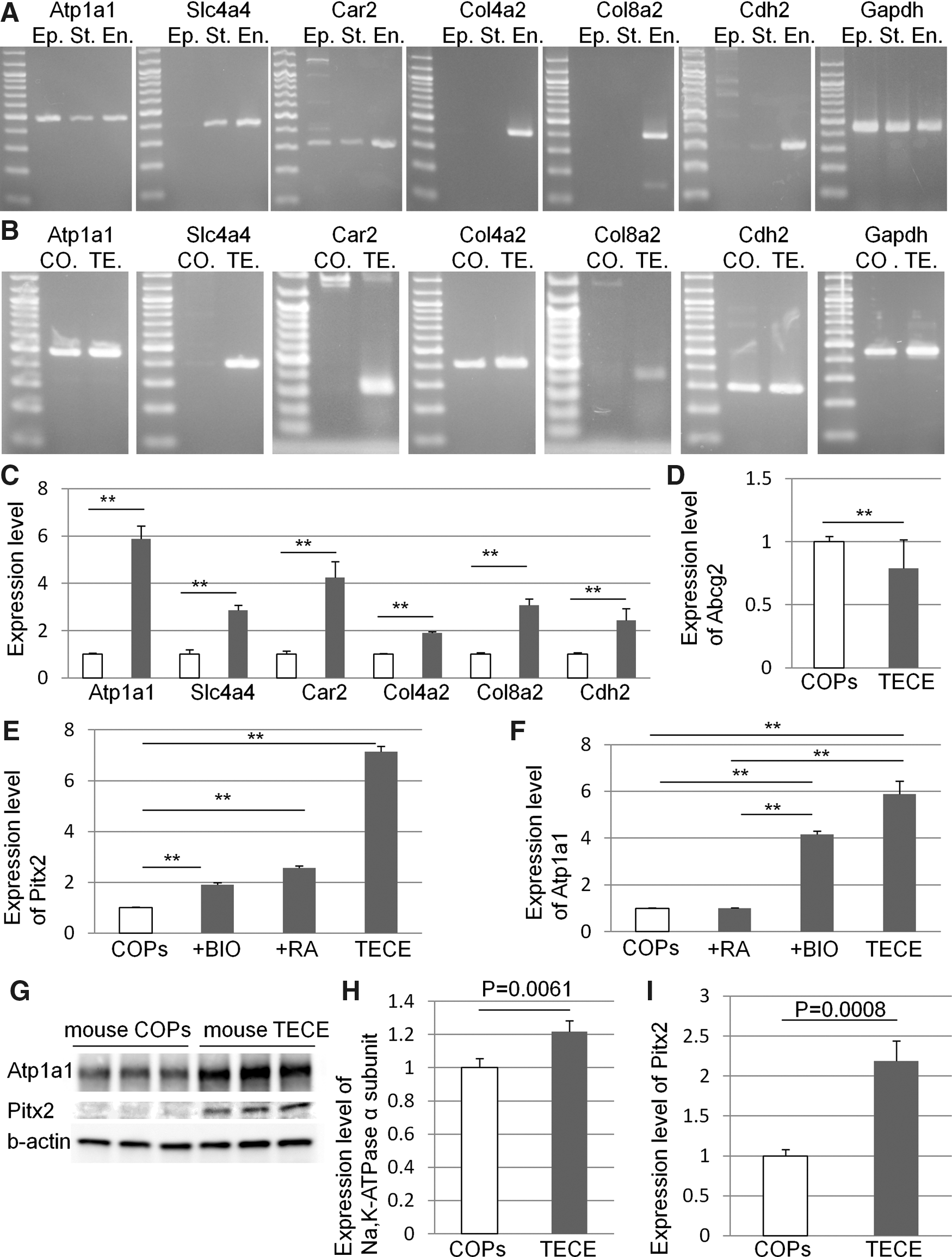
The expression of corneal endothelium markers by RT-PCR.
Figure 2B shows the expression of corneal endothelial markers before and after the induction of TECE from COPs. Before derivation, COPs expressed only limited markers (Atp1a1, Col4a2, and Cdh2); however, after the derivation, TECE expressed all 7 markers. Figure 2C shows the change in the expression level of corneal endothelial markers before and after deriving TECE from COPs. TECE express significantly higher levels of all corneal endothelial markers compared to original COPs. We previously reported that COPs expressed the stem cell marker Abcg2 when examined by RT-PCR and immunocytochemistry [11]. Figure 2D shows that the expression level of Abcg2 was significantly decreased in TECE compared to COPs.
To clarify the role of Wnt/β-catenin signaling and retinoic acid in Pitx2 and Atp1a1 expression and Pitx2 and Atp1a1 expression levels of COPs, COPs cultured with 1.0% fetal bovine serum and 1 μM BIO, COPs cultured with 1.0% fetal bovine serum and 1 μM retinoic acid, and TECE were compared. Figure 2E shows the change in the Pitx2 expression level by these cells. Both BIO and retinoic acid significantly increased Pitx2 expression levels; however, expression was highest in TECE, suggesting that our differentiation protocol mimics embryonic development of corneal endothelial cells. Figure 2F shows the change in Atp1a1 expression levels by these cells. TECE and BIO-treated cells significantly increased Atp1a1 expression levels, whereas retinoic acid did not affect Atp1a1 expression. Western blot analysis also showed that TECE express significantly higher protein levels of Na,K-ATPase α-subunit and Pitx2 compared to COPs (Fig. 2G, H).
Na,K-ATPase pump function of TECE
Figure 3 shows Na,K-ATPase pump function measured by the Ussing chamber in cultured mouse corneal endothelial cells, 3T3 cells (control), COPs (before derivation), COPs cultured with 1.0% fetal bovine serum and 1μM BIO, and TECE derived from COPs. As expected, cultured mouse corneal endothelial cells have a higher SCC (116.3±34.4 μA/cm2) than 3T3 cells (40.8±9.6 μA/cm2) and COPs (15.2±4.6 μA/cm2). COPs cultured with BIO have a significantly higher SCC (127.4±28.8 μA/cm2) than 3T3 cells and COPs, and it was equal to that of cultured mouse corneal endothelial cells. The pump function of TECE (298.8±53.3 μA/cm2) was also significantly higher than 3T3 cells and COPs. Surprisingly, the TECE pump function was 2.6-fold significantly higher than that of cultured mouse corneal endothelial cells.
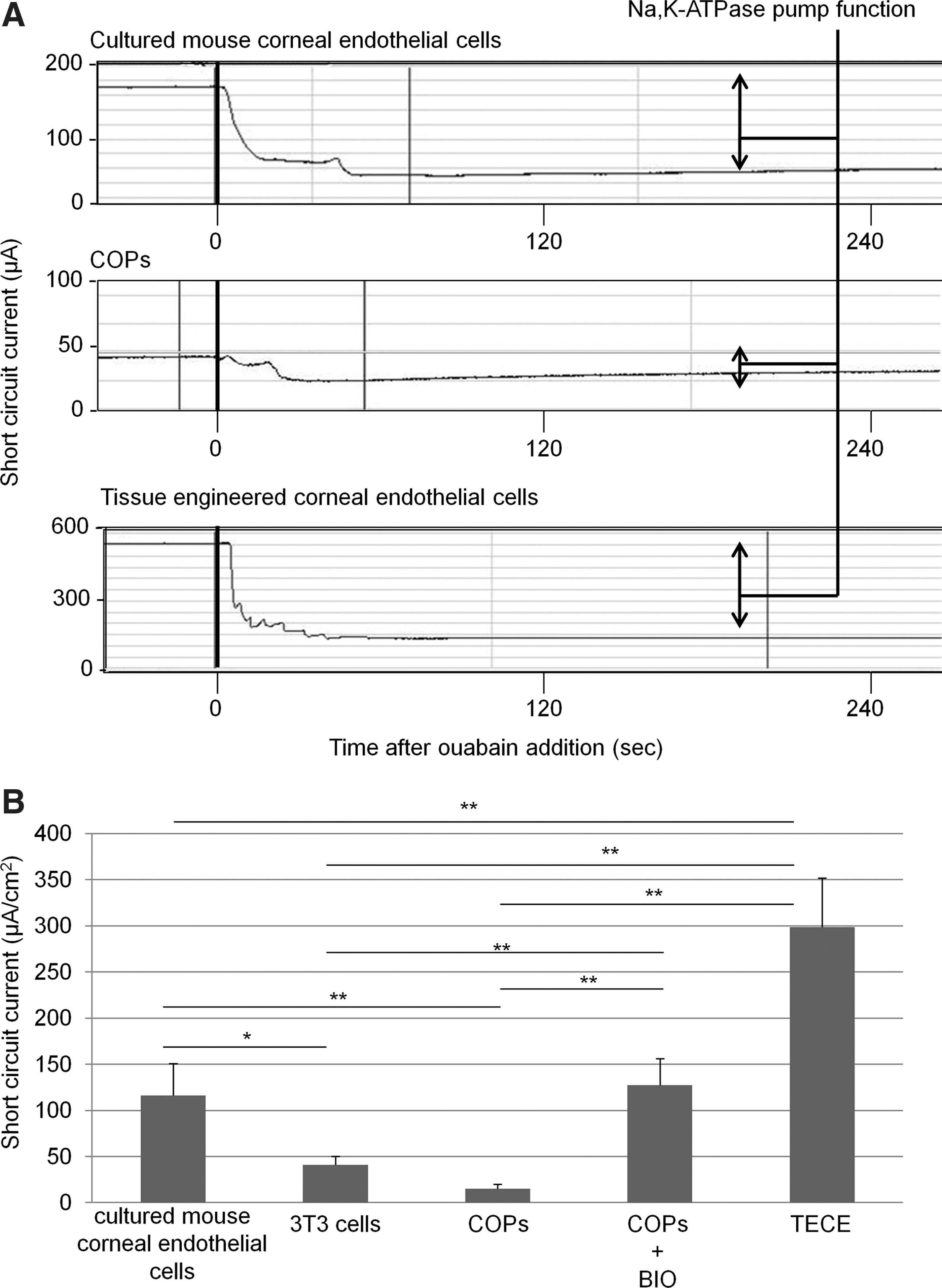
Measurement of pump function by the Ussing chamber system.
In vivo transplantation of TECE sheets
COPs were plated and cultured on 0.1% gelatin-coated type I atelo collagen sheets to induce TECE (see the Methods section), and the collagen sheets with TECE kept solid state during in vitro differentiation for at least 1 week, until transplantation was performed. Six TECE sheets with a cell density of 2470.8±654.9 cells/mm2 and 6 control grafts were transplanted to rabbit corneas by penetrating keratoplasty. Original corneal endothelium and Descemet's membrane were stripped from corneal buttons. TECE on carrier collagen sheets were attached to the posterior surface of the donor buttons and transplanted with 16 interrupted sutures, whereas corneal buttons with neither corneal endothelium nor TECE sheets were transplanted in control eyes. Figure 4A and B show anterior segment photographs of rabbit eyes 8 days after transplantation. Eyes transplanted with TECE were less edematous and maintained transparency (Fig. 4A), whereas corneas of control eyes suffered severe corneal edema and thickened stroma (Fig. 4B). Carrier collagen sheets without TECE were also attached to the grafts and transplanted, but these corneas also suffered severe corneal edema and thickened stroma 8 days after the transplantation (data not shown). Figure 4C and D show harvested corneas 8 days after transplantation. PKH26 signal was detected in harvested corneas transplanted with TECE, suggesting that transplanted TECE remained on the cornea (Fig. 4C), whereas no signals were found on control corneas (Fig. 4D). Host rabbit corneal endothelial cells did not invade from the periphery into the central 8.0-mm area in both TECE sheet-transplanted corneas (Fig. 4E) and control corneas (Fig. 4F). The vertical section of harvested corneal graft with TECE (Fig. 4G) also shows monolayer TECE with both PKH26 signals and EGFP fluorescence on carrier collagen sheets attached to stroma of the corneal graft, which also showed autofluorescence.
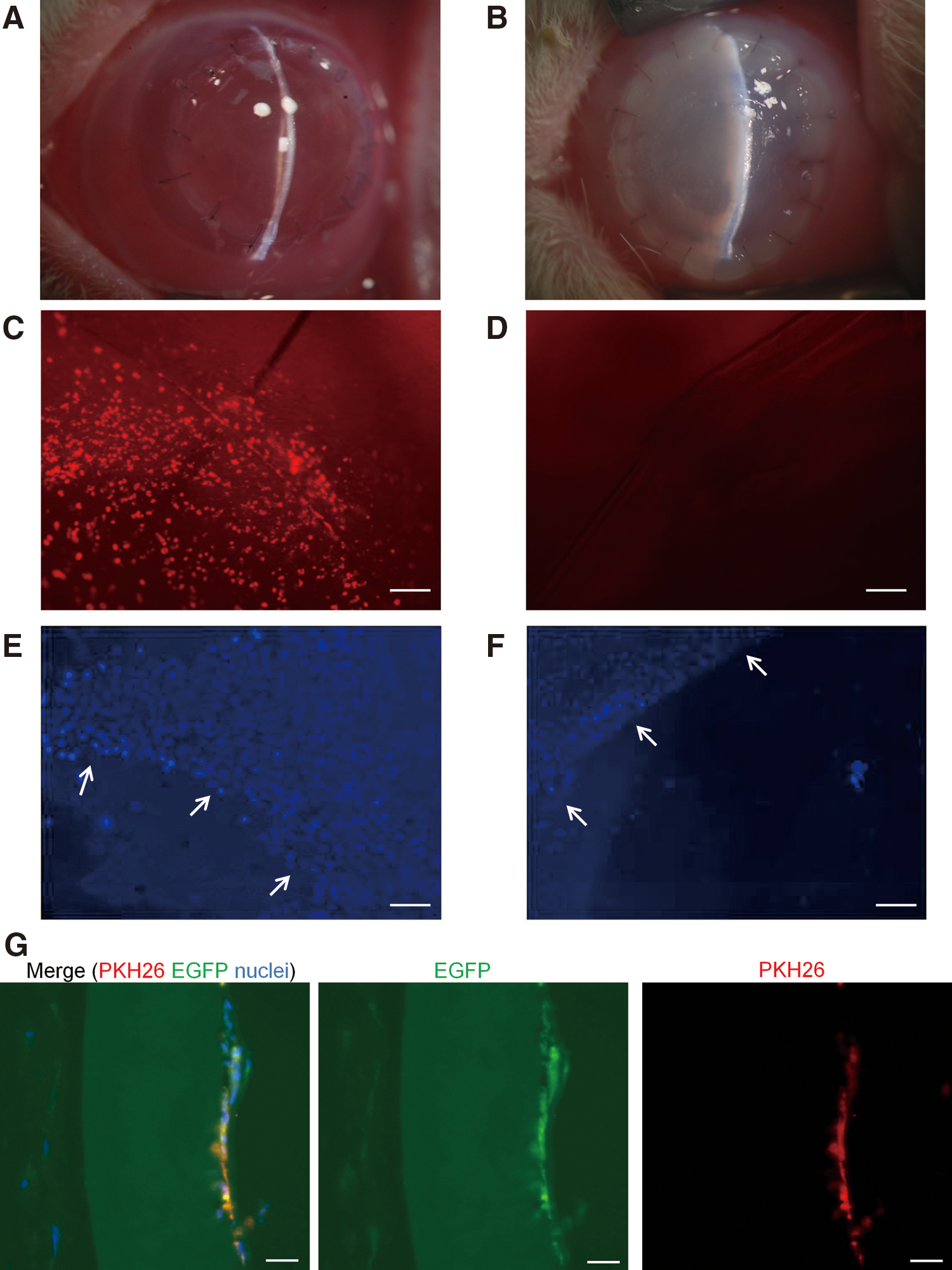
In vivo transplantation of mouse TECE to rabbit cornea.
Figure 5A shows changes in the corneal thickness in the normal control group without any operation, control cornea group transplanted with corneal buttons without Descemet's membrane and corneal endothelium, and TECE sheet-transplanted group. After transplantation, corneal thickness of control eyes increased, whereas TECE-transplanted eyes maintained significant lower corneal thickness than control eyes throughout the postoperative period. Eight days after the operation, the corneal thickness of control eyes was 1105.8±165.9 μm, while the corneal thickness of TECE sheet-transplanted eyes was 497.5±104.9 μm, and the corneal thickness of normal eyes was 398.3±14.3 μm. Figure 5B shows IOP changes after transplantation. IOP increased in both TECE sheet-transplanted eyes and control eyes for several days after transplantation. However, IOP subsequently decreased gradually and was maintained within the 10–20-mmHg range on 8 days after transplantation (control eyes; 20.0±2.2 mmHg, TECE sheet-transplanted eyes; 18.7±7.4 mmHg, normal eyes; 17.1±4.6 mmHg). There was no significant difference in IOP between TECE sheet-transplanted eyes and control eyes. The results indicate that the difference in the corneal thickness was due to TECE function, but not IOP.
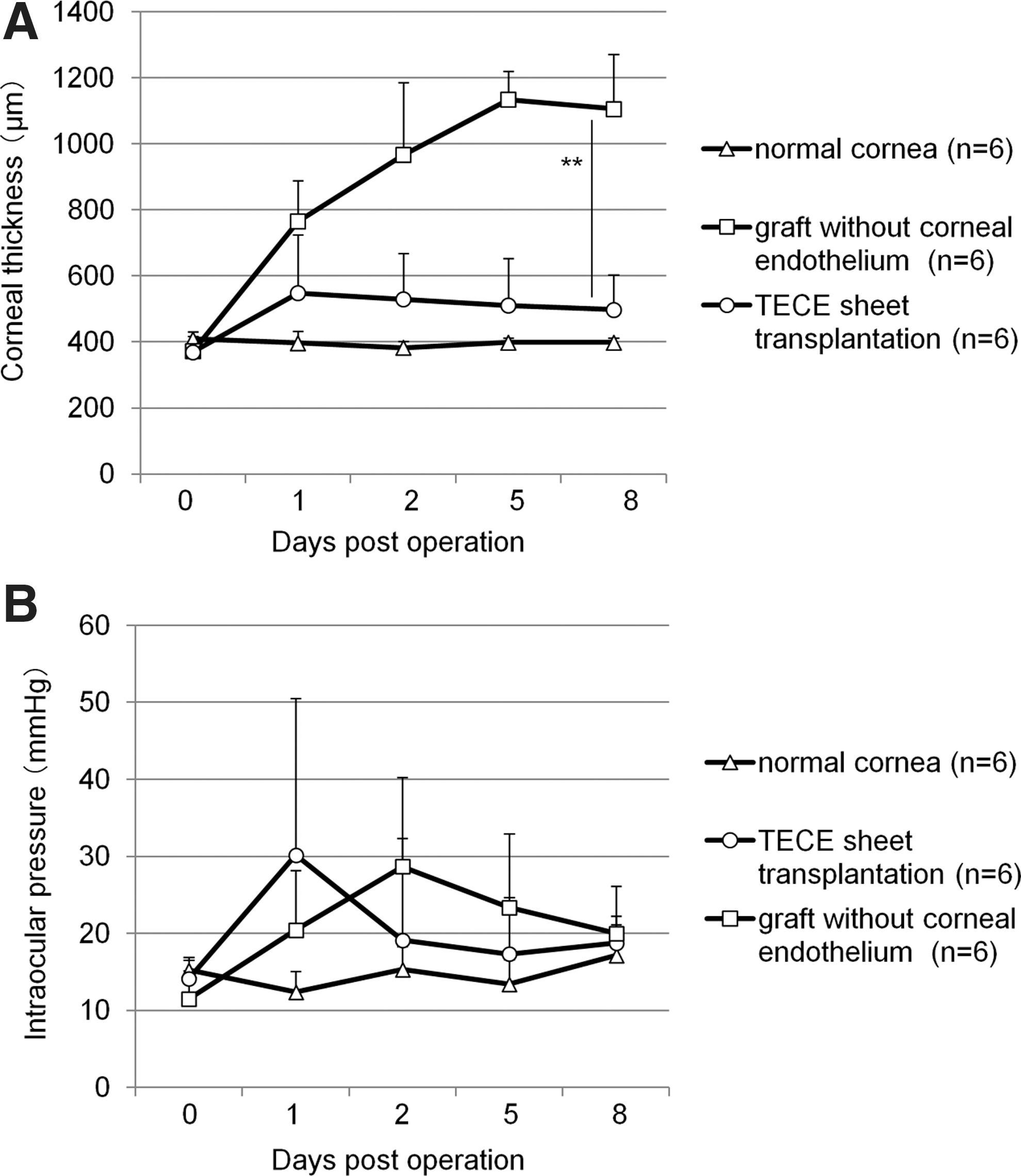
Changes in corneal thickness and intraocular pressure (IOP) of in vivo transplanted rabbit corneas.
Functional TECE induced from human corneal stroma
Human COPs were isolated from human corneal stroma by applying the method in the mouse study with slight modifications (Fig. 6A). Human TECE sheets were also successfully induced, which formed a monolayer with a mosaic pattern (Fig. 6B). Human TECE derived from human COPs expressed all corneal endothelium markers (Fig. 6C). The pump function of human TECE (250.5±42.3 μA/cm2) was significantly higher than 3T3 cells (40.8±9.6 μA/cm2) and the human corneal endothelial cell line B4G12 (117.0±19.1 μA/cm2) (Fig. 6D, E). For in vivo transplantation, 6 human TECE sheets labeled with PKH26 were obtained from the remnants of 3 donor corneas, which were used for keratoplasty. The ages of the donors were 66-, 67-, and 69-years old, and they were all men. In vivo transplantation of human TECE was performed in rabbit eyes by penetrating keratoplasty in the same way as the mouse study. The cell density before transplantation was 2545.8±313.6 cells/cm2. Eyes transplanted with human TECE were less edematous and maintained transparency (Fig. 6F), whereas corneas of control eyes suffered severe corneal edema and thickened stroma (Fig. 6G). The vertical section of the harvested corneal graft with TECE also shows TECE with PKH26 signals (Fig. 6H). TECE-transplanted eyes also showed significantly less corneal thickness than control eyes throughout the postoperative period (Fig. 6I). Eight days after the operation, the corneal thickness of human TECE sheet-transplanted eyes was 589.5±39.6 μm. There was no significant difference in IOP between TECE sheet-transplanted eyes (19.7±11.4 mmHg) and control eyes (20.0±2.2 mmHg) at 8 days after transplantation (Fig. 6J).
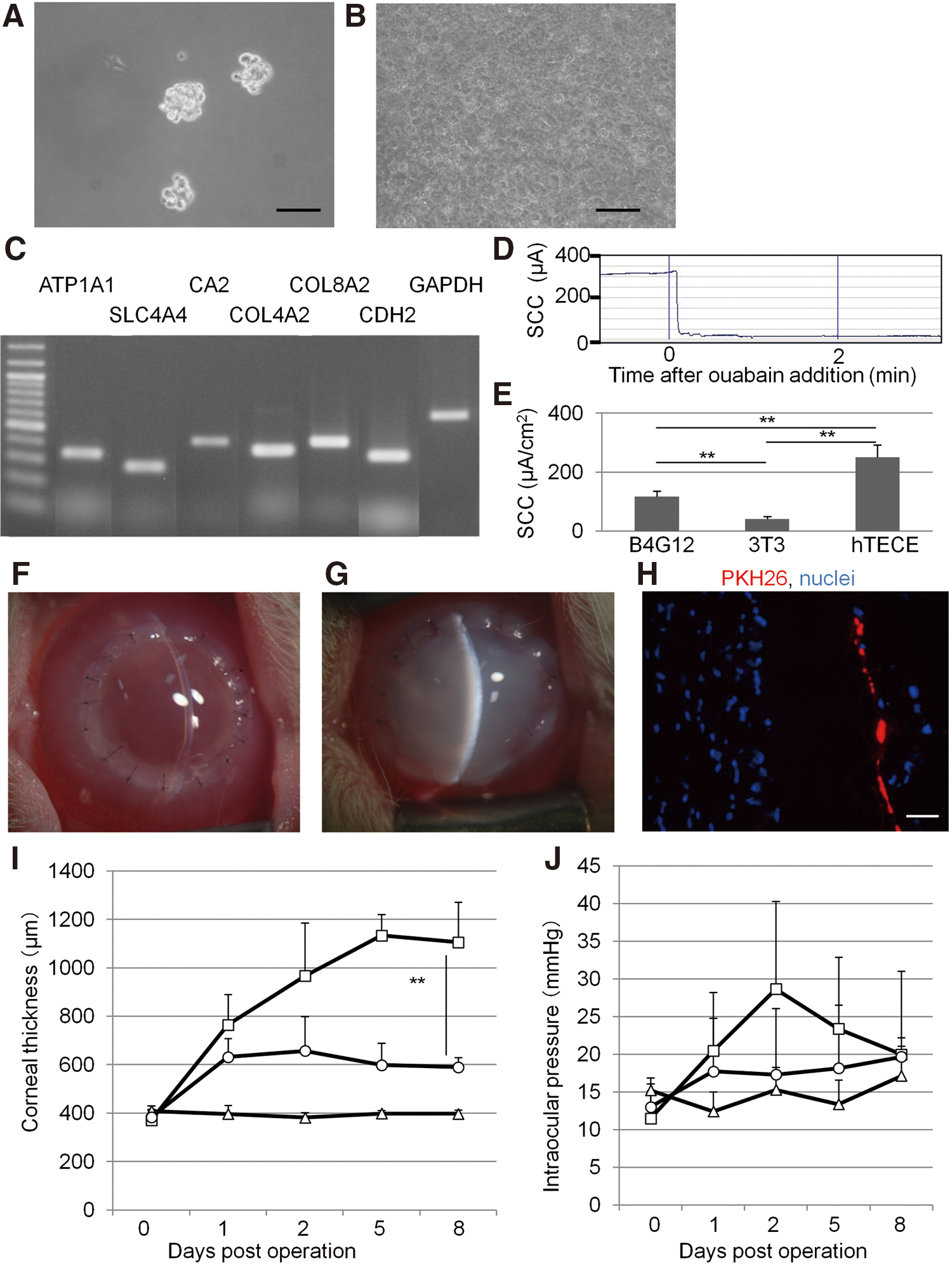
Functional human TECE induced from human COPs.
Discussion
Previously, we reported that COPs isolated from corneal stroma have characteristics of neural crest cells [11,12], which may explain the ability for these cells to differentiate into corneal endothelial cells, which are also of neural crest origin [9]. Recently, multipotent stem cells were also isolated from human corneal stroma as ATP-binding cassette transporter G-family member 2 (ABCG2)-expressing side population cells by cell sorting [19]. In this study, we demonstrated that COPs derived from mouse and human cornea stroma could be a source for TECE. We also showed that COPs derived from Wnt1-Cre/Floxed EGFP mouse corneal stroma expressed EGFP fluorescence, confirming that COPs are neural crest origin.
Previous studies reported that key factors involved in the development of corneal endothelium from neural crest include signaling crosstalk of retinoic acid and Wnt/β-catenin signaling, which are essential for activation and maintenance of Pitx2 expression, a key regulator of development in the anterior segment of the eye [10,14,20]. Inhibition of GSK 3β by BIO leads to increased β-catenin levels and activation of Wnt-dependent transcription [21]. The results in this study show upregulation of Pitx2 by both retinoic acid and GSK 3β inhibitor, a Wnt/β-catenin-signaling activator, supporting the fact that these are key factors for inducing corneal endothelium.
Although Wnt-signaling may lead to another pathway via Rho kinase, the planar cell polarity pathway [22], the specific ROCK inhibitor Y-27632 prevents this pathway. The inhibition of Rho/ROCK signaling by Y-27632 was also reported to promote the adhesion of corneal endothelial cells and inhibit apoptosis [23]. TGF β2 is also essential for anterior chamber development [24], and is responsible for growth arrest and maintenance of mature endothelium during endothelial development [25]. When mouse COPs were cultured with 5 ng/mL TGF β2, expression levels of Col8a2 (collagen type VIII; component of Descemet's membrane) increased, suggesting that TGF β2 affects basement membrane production (Supplementary Fig. S1.). By including retinoic acid, GSK 3β inhibitor, and these supplementary factors in culture medium, we successfully mimicked developmental process from the neural crest to corneal endothelium in vitro. This is shown by how TECE derived from COPs show similar morphology to cultured corneal endothelial cells, express higher levels of all selected corneal endothelial cell markers and Pitx2. Further studies are needed, however, for revealing how Pitx2 causes morphological and functional changes from COPs to corneal endothelium.
Since specific markers of corneal endothelial cells have not yet been established, we selected markers that characterize corneal endothelial function. It is well known that corneal endothelial cells are rich in Na, K-ATPase and carbonic anhydrase [8], and Na,HCO3 co-transporter is necessary for transporting sodium and HCO3 from stroma to endothelium [26,27]. Descemet's membrane consists of collagen type IV and VIII, which are produced by corneal endothelial cells [28]. We therefore selected Atp1a1 (Na,K-ATPase α1-subunit), Slc4a4 (Na,HCO3 co-transporter), Car2 (carbonic anhydrase type 2), Col4a2 (collagen type IV), Col8a2 (collagen type VIII), and Cdh2 (N-cadherin) [9] as corneal endothelium markers. Increased expression of these markers in TECE suggested functional change from COPs to corneal endothelium.
The ability to transport ions from the corneal stroma, causing water to simultaneously flow out of the cornea, is the main function of corneal endothelial cells. Na,K-ATPase expressed in the basolateral membrane of corneal endothelial cells is primarily responsible for this function. COPs cultured with GSK 3β inhibitor and TECE express significantly higher levels of Atp1a1 compared to COPs, whereas supplementation of retinoic acid alone did not change the expression level. Comparison of Na,K-ATPase pump function between COPs and TECE by the Ussing chamber system suggests that our culture protocol successfully derived COPs to TECE with sufficient Na,K-ATPase pump function in vitro, and Wnt/β-catenin signaling had an important role in increasing pump function.
Xenograft transplantation of mouse TECE to rabbit cornea revealed that TECE was able to control corneal hydration in vivo. Compared to control corneas, the eyes that received TECE sheets demonstrated significantly reduced corneal swelling, as well increased corneal transparency. Corneal thickness also depends on IOP [8]; however, there was no significant difference in IOP between TECE sheet-transplanted eyes and control eyes after transplantation, indicating that the final difference in corneal thickness was due to TECE function, but not IOP. Since rabbit corneal endothelial cells are known to proliferate in vivo, TECE were labeled with the cell tracer PKH26 before transplantation to verify that functional TECE after transplantation were responsible for the observed improvement in endothelial function. PKH26-positive TECE was retained over the entire corneal button, and moreover peripheral rabbit corneal endothelial cells were not observed invading into the central 8.0-mm area in both experimental groups. These findings suggest that the retained corneal thickness and transparency were due to endothelial function of transplanted TECE sheets and not due to proliferation of host rabbit corneal endothelial cells.
By applying the above protocol in mice with slight modifications, we showed that functional human TECE sheets can also be engineered from human COPs. Human TECE-transplanted eyes maintained corneal transparency, and also showed significantly lower corneal thickness than control eyes throughout the postoperative period.
Induction of corneal endothelial cells from human COPs may lead to a novel stem cell therapy for corneal endothelial dysfunction. Cultivated corneal endothelial cell transplantation in animal models has been reported; however, clinical application is still not possible, since adult human donor corneal endothelial cells have limited proliferative capacity [29]. Fetal donor corneal endothelial cells may have higher proliferative capacity; however, ethics issues need to be resolved before clinical use. Recently, isolation of endothelial progenitor cells from bone marrow was reported [30]. However, co-culture of bone marrow-derived cells with corneal endothelial cells was required for derivation [30]. On the other hand, the protocol shown in this study does not require co-culture with endothelial cells or feeder cells. Therefore, derivation of endothelial cells from expanded COPs may be a much easier approach for the efficient production of endothelial grafts from corneal stroma. Furthermore, if this protocol can be applied to neural crest cells derived from autologous sources other than the cornea, TECE from somatic cells may be obtained that are free from immunological rejection.
In conclusion, we successfully induced functional corneal endothelial cells from stromal stem cells using a defined medium, including retinoic acid and a GSK 3β inhibitor. Our protocol has the potential to address 2 major issues concerning corneal transplantation, including the shortage of donors worldwide by mass-producing endothelial cell sheets and immunological rejection by using autologous sources. Further refinements to efficiently culture neural crest stem cells are underway.
Footnotes
Acknowledgment
This study was supported by the grants from the Ministry of Education, Culture, Sports, Science and Technology, Japan, especially, the Project for Realization of Regenerative Medicine, with support for the core institutes for iPS cell research (to H.O. and S.S.). This work was also supported by a grant-in-aid from the Global COE Program of MEXT to Keio University.
Author Disclosure Statement
The authors declare no conflict of interest. The sponsor or funding organization had no role in the design or conduct of this research.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.