
Abstract
In a recent study, we showed that the adult pituitary gland is capable of regenerating transgenically ablated growth hormone-producing (GH+) somatotropes. Here, we investigated whether the gland's regenerative capacity is more general and also applies to the other major hormonal cell type, the prolactin-producing (PRL+) lactotropes. We set up the transgenic PRLCre/inducible diphtheria toxin receptor (iDTR) mouse model, in which the PRL promoter drives expression of Cre that induces DTR in lactotropes. Injection of female mice with DT for different periods causes a gradual ablation of PRL+ cells, reaching a maximum of 70% after 10-day DT treatment. During the following weeks, lactotropes progressively reappear achieving a 60% restoration after 6 weeks. The Sox2+ stem/progenitor cell compartment displays a prompt reaction to the DT-triggered cell ablation injury, including expansion of the marginal-zone niche and coexpression of PRL, the latter only very rarely observed in control pituitary. Throughout the regeneration period (2–6 weeks), Sox2+ as well as double Sox2+/PRL+ cells continue to be more abundant than in control pituitary. In addition to this stem cell reaction, surviving or newborn lactotropes increase their proliferative activity, and bihormonal PRL+/GH+ cells become detectable suggesting somatotrope-to-lactotrope transdifferentiation. In conclusion, the adult pituitary gland is capable of restoring lactotrope cells after destruction, further confirming its regenerative competence. Repair of lactotropes appears to be driven by a combination of mechanisms, including recruitment from stem cells, proliferation of lactotropes, and transdifferentiation of somatotropes.
Introduction
A
In many adult organs, stem cells participate in the repair process after damage [8 –11]. In a recent study, we demonstrated that the pituitary stem cells promptly react to injury inflicted at adult age through transgenic ablation of somatotropes, and that the destroyed GH+ cells are substantially replaced, most plausibly from the activated stem cells [12]. Thus, the mature pituitary is capable of regeneration, an issue unsettled and controversial until this study [11,13,14]. Here, we investigated whether the pituitary's regenerative capacity is not only limited to somatotropes, and used a comparable transgenic model in which the other main hormonal cell type, the PRL-producing lactotrope, is destroyed using the “diphtheria toxin (DT)/inducible DT-receptor” (iDTR) system [12,15,16]. We found that also lactotropes are regenerated, but that in contrast to the somatotrope restoration, not only stem cells, but also other mechanisms appear to be involved.
Materials and Methods
Transgenic mice
PRLCre mice express Cre recombinase under control of the rat 3.2-kb PRL promoter, and were kindly provided by Dr. LeTissier (Division of Molecular Neuroendocrinology, National Institute for Medical Research, The Ridgeway, Mill Hill, London, UK) [17]. For genotyping, genomic DNA was subjected to PCR with 5′-TGACGGAAATAGATGATTGG-3′ as forward primer, and 5′-ACGCGTTGAAAGATGGCCTTTC-3′ (rat) and 5′-TTCTGGTCTTGAGTAGTCAC-3′ (mouse) as backward primers, yielding a transgene-specific amplicon of 1,480 bp and an endogenous (mouse) amplicon of 1,600 bp. iDTR mice harbor the DTR gene preceded by a STOP cassette, which is flanked by LoxP sites. Cre recombinase binds to these sites and excises the STOP sequence, thereby rendering the Cre-, thus the DTR-expressing cells susceptible to ablation by DT through apoptosis [12,15,16]. Homozygous iDTR/iDTR mice were crossed to hemizygous PRLCre/- animals, yielding PRLCre/iDTR mice (hemizygous for both transgenes) and −/iDTR littermates (controls).
Mice were bred on the original C57Bl6/CBA genetic background and kept in the Animal Housing Facility of the University (KU Leuven, Belgium), providing constant temperature, humidity, day/night cycle, and water and food ad libitum. Animal experiments were approved by the KU Leuven Ethics Committee.
Treatment with DT
Adult (8- to 12-week-old) PRLCre/iDTR and −/iDTR mice were intraperitoneally (i.p.) injected with DT (4 ng/g body weight, twice a day; Sigma-Aldrich) or vehicle (PBS; Invitrogen) for the time periods mentioned (see Figs. 1A and 2A). Toxicity was not observed as assessed by overall health, including body weight, hair, posture, walk, and explorative behavior [12]. For independent experiments, 3–6 mice were grouped per condition. Since highest ablation was obtained in female mice (see Results), all experiments (except one mentioned at the beginning of the Results) were performed with this gender (random cycle). Control experiments were done as described before [12]; neither DTR nor Cre expression per se, nor DT treatment in the absence of DTR expression affected the pituitary architecture and the proportions of the different hormonal cell populations (data not shown).
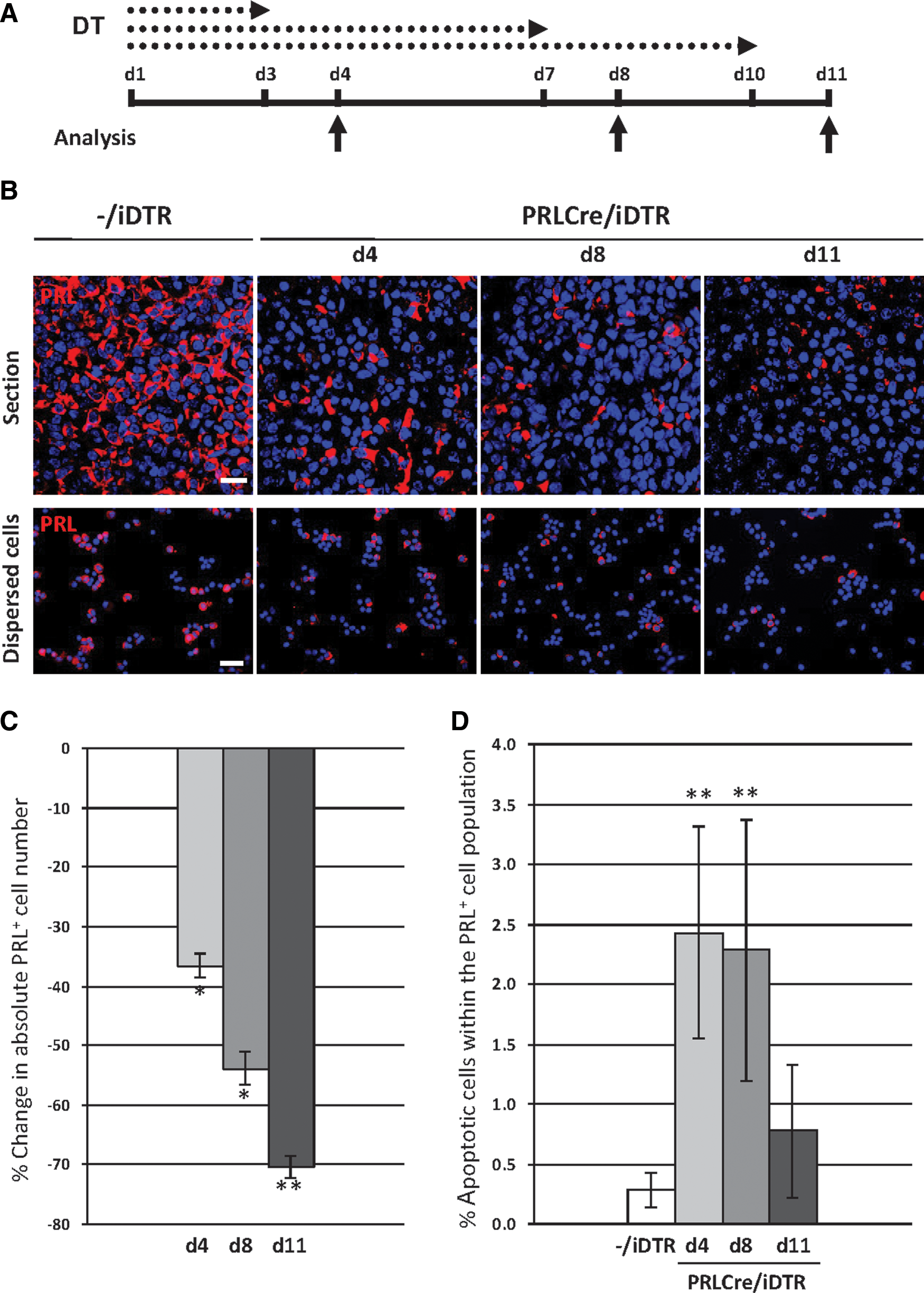
Gradual ablation of lactotropes in DT-treated PRLCre/iDTR mice.

Regeneration of lactotropes after DT-mediated ablation.
Immunofluorescence, AMCA labeling, and TUNEL analysis of dissociated AP cells
Mice were euthanized with CO2, and the anterior pituitary (AP) lobe carefully isolated and dissociated into single cells using trypsin as described before [2,3,12]. Cells were resuspended in a serum-free pituitary-optimized cell culture medium (Invitrogen) [2,3,12] and spun down on cytospin slides (Superfrost Plus; Thermo Scientific). Cells were fixed with paraformaldehyde (4% in PBS), permeabilized with saponin (Sigma-Aldrich; 0.5% in PBS), and incubated overnight with the following antibodies: rabbit anti-mouse GH (1/10,000), guinea pig anti-rat GH (1/5,000), rabbit anti-rat PRL (1/10,000), guinea pig anti-rat PRL (1/2,500), rabbit anti-rat ACTH (1/5,000), guinea pig anti-rat αGSU (1/1,000) (all obtained from Dr. A. F. Parlow, NHPP, Harbor-UCLA Medical Center, Torrance, CA), goat anti-human Sox2 (1/250–1/750; Immune Systems), and rabbit anti-Ki67 (1/50; Thermo Scientific). Subsequently, cells were incubated during 1 h with Alexa Fluor 488-, 555- (Invitrogen), and/or FITC- (Jackson Immunoresearch Europe) labeled secondary antibodies (at 1/1,000), and then 4′, 6-diamidino-2-phenylindole (DAPI; 0.5 μg/mL; Sigma-Aldrich). Images were captured using an Olympus IX81 microscope equipped with Cell^M software (Olympus). Cells were counted using ImageJ (
Because of the reduction of the total cells within the AP when lactotropes are destroyed, unaffected hormonal cell types logically increase in proportion. Therefore, proportions do not correctly reflect the real abundance of the cell types, and absolute cell numbers must be determined for appropriate interpretation and comparison. The absolute cell number of each hormonal cell population was calculated by multiplying its proportion by the total number of AP cells (as obtained after AP dispersion).
Figures shown were prepared with ImageJ and assembled with Microsoft PowerPoint (2007). Negative controls for immunostaining (without primary antibodies) did not show any signal [2,3,12; and data not shown].
Folliculostellate (FS) cells were identified by the uptake of the fluorescent dipeptide β-Ala-Lys-Nɛ-7-amino-4-methylcoumarin-3-acetyl (further referred to as AMCA; Biotrend) as described before [12]. In the AP, uptake of AMCA is a specific property of FS cells [21]. Briefly, dispersed AP cells were incubated with AMCA (40 μM) for 2 h at 37°C, and then fixed and immunostained for Sox2 and Ki67 as described above.
Apoptosis was assessed by terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) analysis in cytospin preparations of fixed AP cells using the FragEL kit (Calbiochem) according to the manufacturer's protocol [12].
Immunofluorescence of pituitary vibratome sections
Coronal pituitary vibratome sections were obtained and immunostained as described in detail before [2,3,12,22]. Briefly, pituitaries were isolated, fixed in 4% paraformaldehyde, embedded in 2% agarose (MP Biomedicals), and coronally sectioned to 45-μm slices using a vibratome (Microm HM 650V; Prosan). After permeabilisation with 0.4% Triton X100, sections were incubated with antibodies as described above for cells, followed by nuclear labeling with ToPro-3 (0.01 mM; Invitrogen) and scanning using a confocal laser scanning microscope (Zeiss LSM 510; Zeiss). Images were prepared with Zeiss LSM Image Brower and Microsoft PowerPoint (2007).
Statistical analysis
One-way analysis of variance (ANOVA) was applied (SPSS, Version 13.0; IBM). For comparison of 3 groups, ANOVA was followed by Least Significant Difference Post Hoc Multiple Comparison. Statistical significance was defined as P<0.05.
Results
Gradual ablation of lactotropes in DT-treated PRLCre/iDTR mice
Adult PRLCre/iDTR and control (−/iDTR) mice were injected with DT for different periods (3, 7, or 10 days), and pituitaries were analyzed 1 day later (d4, d8, or d11) (Fig. 1A). Lactotrope ablation was not very efficient in male mice (35% reduction of absolute PRL+ cell number after 10-day DT treatment; n=5). Because we aimed at the maximal cell obliteration to study the regenerative capacity of the gland, all further experiments were performed with female mice.
PRL-immunocreactive (PRL+) cells gradually decline in PRLCre/iDTR pituitaries, the ablation becoming more pronounced with a longer DT treatment period (Fig. 1B). Some of the surviving lactotropes become hypertrophic, and some are connected and organized in clusters (data not shown). To determine the ablation grade, PRL-immunoreactive (PRL+) cells were counted after AP cell dispersion, and absolute cell numbers were calculated (see Materials and Methods) to take into account the reduced total AP cell number as observed from d8 (∼30% reduction, corresponding to a decline in pituitary weight; see Supplementary Fig. S1B, C). PRL+ cell ablation reaches ∼70% after 10-day DT treatment (Fig. 1C), which does not further increase (data not shown). This ablation grade is concordant with the reported activity of the PRLCre transgene in 70%–80% of the lactotropes in female mice [17]. In analogy, apoptotic PRL+ cells are highest between d4–d8 and return to basal (control) levels at d11 (Fig. 1D), indicating that all susceptible lactotropes are killed by that time. We further examined whether the other hormonal cell lineages are influenced. ACTH-immunopositive (ACTH+) corticotropes and α-glycoprotein subunit-immunoreactive (αGSU+) thyrotropes, gonadotropes, and αGSU-only cells are not affected (Supplementary Fig. S1D–F). GH+ somatotropes, however, slightly, but significantly decline in absolute number (∼20% at d11; Supplementary Fig. S1D–F). Possible explanations for this decay are given in the Discussion. There were no signs of inflammation (data not shown), neither infiltration of mononuclear immune cells (lymphocytes, monocytes, macrophages), nor of polymorphonuclear immune cells (neutrophils, basophils, eosinophils). Absence of inflammatory infiltrate is in accordance with our previous study [12] and other studies using DT-mediated cell ablation through apoptosis [15, 23 –25].
Taken together, we set up a transgenic mouse model to conditionally obliterate lactotropes, which allowed us to study whether and how the pituitary gland repairs this injury and restores the destroyed cells.
Gradual regeneration of lactotropes after DT-induced ablation in PRLCre/iDTR mice
Female mice were injected with DT for 10 days to obtain maximal lactotrope ablation, and pituitaries were analyzed at d11 as well as 2, 4, and 6 weeks later (Fig. 2A). From in situ examination of pituitary sections, it is clear that PRL+ cells gradually reappear (Fig. 2B, upper row), which is confirmed by cell counting; the ∼70% decline in absolute PRL+ cell number in PRLCre/iDTR mice (as compared to control mice) progressively decreases to ∼30% after 6 weeks (Fig. 2C). Further regeneration was not observed (as analyzed after 12–16 weeks; data not shown). Of note, the absolute GH+ cell number is restored to control levels after 4 weeks (Supplementary Fig. S2A–C), while ACTH+ and αSU+ cells do not change (Supplementary Fig. S2B, C and data not shown).
Taken together, the pituitary gland is capable of regenerating lactotropes to a considerable extent within weeks, as well as recovering the slightly declined somatotropes.
Multiple mechanisms of lactotrope regeneration
Lactotropes may be regenerated after destruction through several mechanisms, including recruitment from stem cells, proliferation of surviving or newborn PRL+ cells, and/or transdifferentiation from somatotropes. In our previous study, we advanced development from stem cells as a major mechanism in the restoration of GH+ cells after somatotrope ablation [12].
Involvement of stem cells in lactotrope regeneration
The Sox2+ pituitary stem cells promptly react to the DT-provoked lesion in the PRLCre/iDTR mice. At some locations, in particular, the marginal-zone niche and the wedge region (where the anterior and intermediate lobes meet; see [12,22]) Sox2+ cells are clearly expanded at d8–d11 (Fig. 3A, B). At some sites, a multilayer Sox2+ marginal zone is observed, whereas a unicellular layer is generally seen in control mice (Fig. 3A). The absolute number of Sox2+ cells within the total AP cells is increased by ∼2-fold (Fig. 3C). As observed before [3,12,22], the Sox2+ cell compartment does not only include cells with Sox2 in the nucleus, but also some cells with Sox2-immunoreactive signal in the cytoplasm, proposed to represent cells in which the nuclear action of Sox2 is stopped, thereby allowing differentiation [3,12,22]. Exclusion of Sox2 from the nucleus by, for instance, nucleocytoplasmic shuttling, has also been shown to permit differentiation in other stem cells [26]. These Sox2-cytoplasmic cells are more frequently encountered—mostly grouped—, in the anterior lobe's parenchyma of PRLCre/iDTR mice, particularly at the end of the peak period of lactotrope ablation (d8; Fig. 3A), which is also confirmed by counting (Fig. 3C). The expansion of the Sox2+ stem/progenitor cells is, at least partially, explained by increased proliferative activity at d4 and d8 (as assessed by Ki67 immunostaining), returning to the basal control level of practically not observable dividing Sox2+ cells at d11 (Fig. 3D, E). Also the FS cells, previously proposed as pituitary stem/progenitor cells and significantly overlapping with the Sox2+ stem/progenitor cell compartment [3 –6,11,12], increase in number (as visualized by AMCA uptake), although to a lower extent than the Sox2+ cells (Fig. 3D and Supplementary Fig. S3A). In analogy, proliferative activity of the AMCA+ cells increases, but not as prominently as in the Sox2+ cell compartment (and not statistically significant) (Fig. 3D and Supplementary Fig. S3B). Of note, the proportion of double Sox2+/AMCA+ cells within the AMCA+ cell population shows a tendency (P>0.05) of increase from ∼80% (control) to ∼90% at d4 and d8, and a subsequent decrease at d11 (Supplementary Fig. S3C). Together, the AMCA+ cells may also contribute to the enlargement of the Sox2+ cell compartment, by first expanding (either as AMCA+/Sox2+, or as AMCA+ cells that then start to express Sox2), and subsequently loosing the AMCA uptake ability.
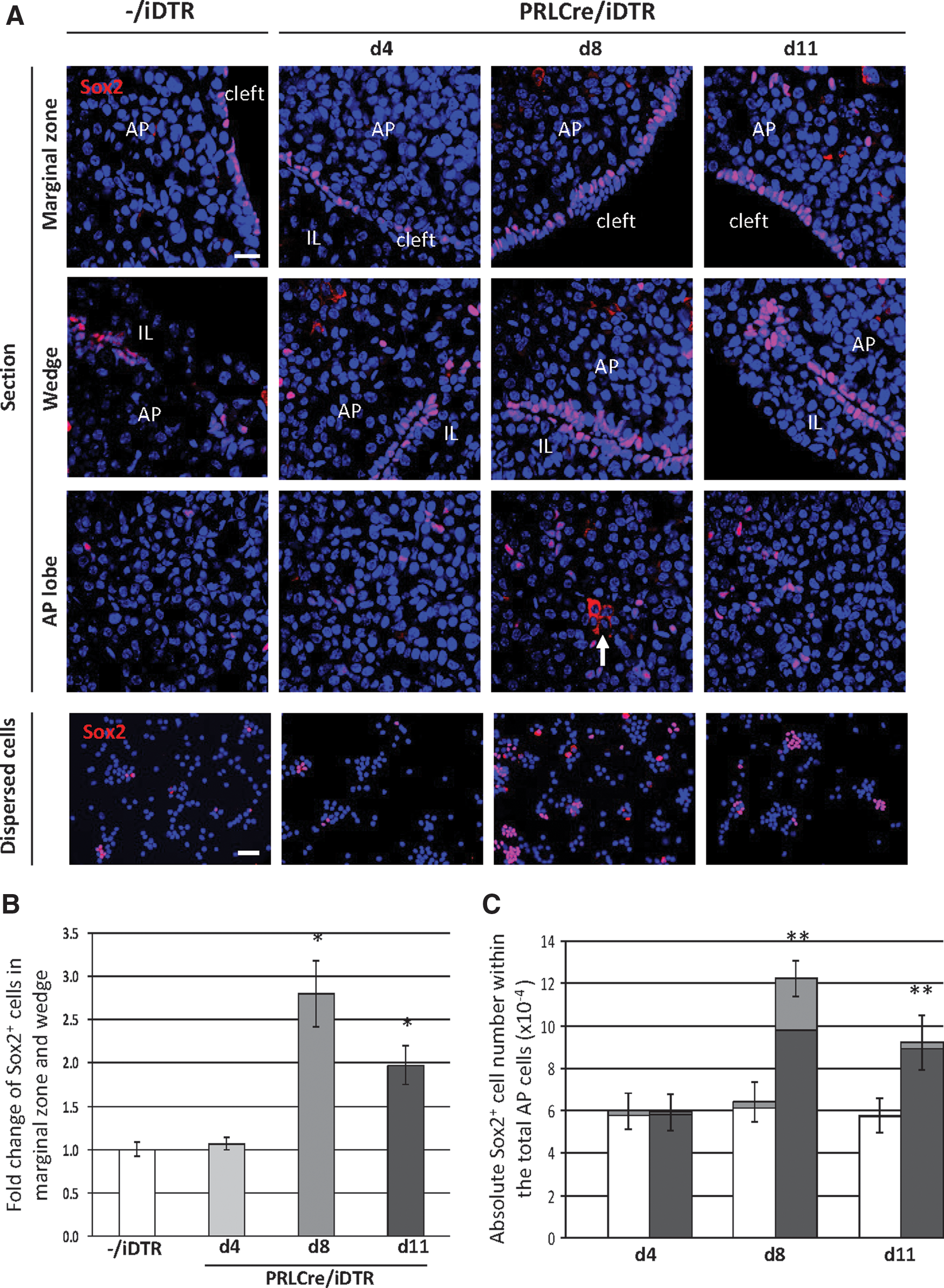
Acute response of the Sox2+ stem/progenitor cell compartment to DT-induced lactotrope ablation.
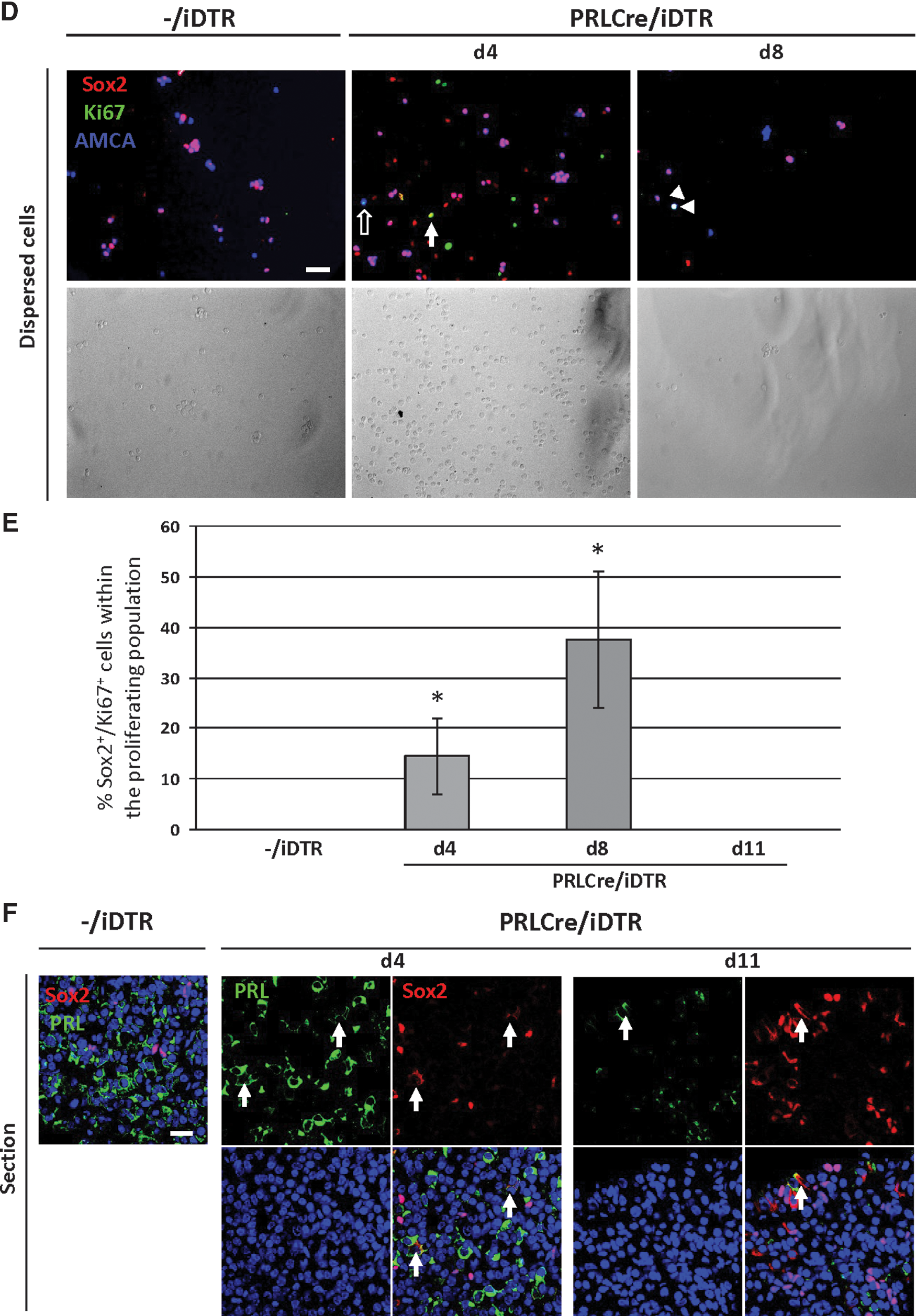
To investigate whether the activated Sox2+ stem/progenitor cell compartment is involved in lactotrope regeneration, double Sox2/PRL immunostaining was performed. During the acute reaction (d4–d11), PRL is found in Sox2-cytoplasmic cells in the PRLCre/iDTR mice (Fig. 3F). Although at low frequency, these cells are more readily encountered (1.6%±0.6% of the Sox2+ cells; mean±SEM of n=4) than in the control mice (<0.1%). GH was virtually not found together with Sox2 (Supplementary Fig. S4A). In agreement with our previous studies, some sporadic Sox2-cytoplasmic cells contain ACTH or αGSU, however, not different from control pituitary (Supplementary Fig. S4A), considered to represent normal turnover events in the slowly renewing adult gland [1,3,12,22].
During the regeneration period (2 to 6 weeks after the 10-day DT injection), Sox2+ cells remain more abundant in the PRLCre/iDTR than the control pituitary (Fig. 4A–C). Sox2-cytoplasmic cells are most numerous after 2 weeks, and then return to control levels (Fig. 4C). This picture may mirror the restorative attempt at d4–d8, but in contrast, this response can now develop because DT has been cleared [12,15,23 –25] and nascent lactotropes are no longer killed (in contrast to d4–d8). During the regeneration period, PRL remains to be found together with Sox2 more easily in the PRLCre/iDTR than the control mice, although their occurrence remains low (Fig. 4D; 2.1%±0.9% of the Sox2+ cells at 2 weeks after injection versus <0.1% in control mice; n=4). GH remains virtually absent from Sox2+ cells, whereas ACTH and αGSU are present in some sporadic Sox2-cytoplasmic cells, not different from control mice (Supplementary Fig. S4B). Both in control and PRLCre/iDTR mice, ACTH (not the other hormones) is sometimes found in cells that have Sox2 in the nucleus (Supplementary Fig. S4B, arrowheads), as also reported before [22]. At present, we do not know the meaning of this observation.

Phenotype of the Sox2+ stem/progenitor cell compartment during the regeneration process.
Taken together, these data indicate that stem/progenitor cells promptly react to the lactotrope-ablation injury by expanding, and suggest that they are involved in the regeneration process, at least partially, by initiating PRL expression.
Involvement of lactotrope proliferation in the regeneration process
We further examined the proliferative activity of surviving (or newborn) PRL+ cells as a possible mechanism of lactotrope regeneration. During the acute period (d4–d11), proliferating (Ki67+) lactotropes increase 4- to 7-fold when compared to control mice (Fig. 5A, B). During the further regeneration period (2–6 weeks), even a higher proportion (2%–4%) of the PRL+ cells is dividing in the PRLCre/iDTR mice (Fig. 5A, B), which is likely explained by the fact that proliferating PRL+ cells (surviving or newborn) are not killed anymore after clearance of DT from the system.

Lactotrope proliferation in DT-treated PRLCre/iDTR mice.
Thus, the pituitary acutely attempts to compensate the loss of lactotropes, at least, in part, through proliferation of surviving PRL+ cells. During the further regeneration period, lactotrope restoration also appears partly driven by proliferation of (surviving or newborn) PRL+ cells.
Involvement of transdifferentiation in lactotrope regeneration
Transdifferentiation between somatotropes and lactotropes, reported to take place under certain circumstances (see eg, [27,28]), may also be involved in the regeneration process. Therefore, we investigated the presence of transitory bihormonal lactosomatotropes. At all time points analyzed (d4 till 6 weeks), double GH+/PRL+ cells are more abundant in the pituitary of DT-treated PRLCre/iDTR mice when compared to control mice (Fig. 6). Thus, transdifferentiation may also, at least, in part, be responsible for the lactotrope restoration process. Proliferating GH+ cells, not detected in control mice, but in PRLCre/iDTR mice at 2, 4, and 6 weeks after DT (although rare and estimated 0.01%–0.05% of the total AP cells; data not shown), may provide the source of the transdifferentiating cells while keeping the number of somatotropes stable.

Analysis of double GH+/PRL+ cells (lactosomatotropes) in DT-treated PRLCre/iDTR mice.
In conclusion, our data show that lactotropes are regenerated after transgenic ablation and support the involvement of multiple processes, including recruitment from stem cells, from proliferating lactotropes, and from transdifferentiating somatotropes. Lineage-tracing studies are required for further proof, but the necessary transgenic models are at present not available (see Discussion).
Discussion
The adult pituitary gland can restore somatotropes after transgenic ablation [12]. Here, we show that also lactotropes can be substantially renewed after destruction, thereby supporting a more general regenerative capacity of the mature gland. In both pituitary injury models, stem cells promptly react by expanding (most obvious at the marginal-zone and wedge niches) and by coexpressing the ablated hormone, thus supporting their differentiation toward the affected cell type. However, there are also noticeable differences between the somatotrope (see [12]) and lactotrope regeneration process (as described here). Whereas surviving somatotropes do not proliferate in response to the cell destruction within their population, lactotropes clearly react by increasing their proliferative activity. Along the same line, lactosomatotropes, generally considered as transitory stages during transdifferentiation, are virtually absent in the somatotrope-ablated pituitary, but clearly present in response to lactotrope obliteration. Thus, mechanisms of repair appear to depend on the cell type that is affected. Indeed, whereas scarce for somatotropes, there are several indications that lactotropes represent a plastic, dynamic cell population in the pituitary [28 –30]. The dissimilar plasticity of both cell types, and the inherent link to either a single or multiple mechanisms involved, may further explain the different pace of regeneration, being weeks for lactotropes and months for somatotropes, as well as the final restoration grade, being more efficient for lactotropes. On the other hand, the final ablation of somatotropes was higher (90%; see [12]) compared with lactotropes here (70%), which may also explain the superior restoration of the lactotropes. Moreover, the different degrees of ablation may also explain the different regeneration strategies employed. In pancreas, it has been shown that only extensive ablation activates the resident stem cells for regeneration, whereas lower degrees of injury can be repaired by self-replication of β-cells [25,31].
Also, within the stem cell response, differences are noted between both models. The expansive reaction is faster and depicts as more prominent in the somatotrope ablation model (already after 1–2 days of DT injection; see [12]), which may be accounted for by the swifter and more vigorous ablation process (injury) than in the PRLCre/iDTR model described here. In addition, peculiar small Sox2-cytoplasmic cells immediately start to coexpress GH after somatotrope ablation (from d3; [12]), whereas in the PRLCre/iDTR model here, such small cells are not observed and the coexpression (in larger Sox2-cytoplamic cells) starts later (d4). The meaning of these observations requires further investigation.
Further support and direct demonstration of development of new lactotropes through the different mechanisms proposed requires multiple lineage-tracing studies, in which, for instance, Sox2+ stem cells or GH+ somatotropes are marked with a label that remains present in the progeny. However, the Cre-driven ablation in our model precludes the use of a Cre-based approach for lineage tracing (such as Cre-induced reporter activation regulated by tamoxifen). At present, no appropriate (pituitary) models using other tactics (such as Flp/Frt) are available. In addition, we can so far not exclude that (partial) dedifferentiation of surviving and/or DT-affected lactotropes or of somatotropes plays a role; these cells may regain a stem cell phenotype (with or without reinitiation of Sox2 expression), and subsequently restart lactotrope differentiation. Finally, proliferating PRL+ cells may represent surviving lactotropes, but also newborn PRL+ cells that just recently developed from Sox2+ stem cells. Again, complex and compound transgenic models are needed, but not available (e.g., strictly time-controlled and inducible PRL, GH, and/or Sox2 lineage-tracing systems, not using Cre/LoxP).
Full recovery was not attained, neither after somatotrope [12], nor after lactotrope ablation, even following longer lag periods. Possibly, no full restoration is needed to attain satisfactory hormone functions, or the mature organ, in contrast to the embryonic tissue, is not competent anymore for full restoration, as also found in other organs (see eg, [32 –34]). In analogy, also other mature organs (e.g., thyroid gland, heart) only rarely regenerate to a full extent [8,35,36].
In addition to the transgenically targeted lactotropes, somatotropes also decline in the PRLCre/iDTR model. Because less than 1% of GH+ cells show transgenic promoter activity in the PRLCre mouse [17]—also encompassing the GH+ cells that have expressed PRL at some time point during their development—, it is unlikely that the 20% somatotrope reduction is due to a direct impact of DT through DTR expressed in these cells. The acute decrease in somatotropes may be explained by their transdifferentiation to complement the loss in the affected lactotrope population, which, however, does not succeed because the newborn lactotropes are killed as long as DT is present [12,15,23 –25]. Alternatively, the dying lactotropes may liberate factors inducing death of neighboring cells, most of which are somatotropes. Finally, we cannot exclude that loss of trophic signals from the disappearing lactotropes may play a role [37]. However, this paracrine impact on the somatotropes would then occur at a very fast pace. Later, during the regeneration period (4–6 weeks), somatotropes are restored and further kept stable despite the putative transdifferentiation to lactotropes. Self-duplication may be one underlying mechanism, since we observed proliferating somatotropes in the PRLCre/iDTR, but not the control pituitary. Transdifferentiation from the surviving and/or newly formed lactotropes may represent another possibility. Thus, somatotropes recur here in a different manner than after their more vigorous ablation in the GHCre/iDTR model, where increased proliferation and transdifferentiation are not observed [12].
In conclusion, the mature pituitary can restore more than one cell type after injury, and restorative mechanisms seem to depend on the cell type that is affected. Further studies deciphering molecular pathways (including hypothalamic influence) underlying pituitary cell regeneration, may guide to potential clinical applications. As suggested by this and our previous study [12], different therapeutic approaches may be needed for different cell types.
Footnotes
Acknowledgments
This work was supported by the Fund for Scientific Research-Flanders (Belgium) (FWO-Vlaanderen) and the Research Fund (Onderzoeksfonds) of the KU Leuven. Q.F. received a Selective Bilateral Agreement (SBA) Scholarship (KU Leuven).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.