
Abstract
Naïve and primed embryonic stem cells (ESCs) represent 2 pluripotent states of mouse embryonic stem cells (mESCs), corresponding to the pre- and postimplantation cells, respectively, in vivo. Primed ESCs are distinct from naïve cells in biological characteristics, genetic features, developing potentials, and antagonistic signal pathway dependences to support undifferentiated growth. In vitro, naïve mESCs are readily converted to primed cells upon transferring to primed pluripotency signaling. ESC-derived epiblast stem cells (ESD-EpiSCs) are stabilized primed cells derived from naïve mESCs in vitro, and cannot be maintained with leukemia inhibitory factor (LIF) signaling with or without mouse embryonic fibroblasts as the feeder layer. Here, we show that the undifferentiated growth of ESD-EpiSCs could be maintained with the basic fibroblast growth factor employing human amnion epithelial cells (hAECs) as the feeder layer. Upon exposure to LIF, ESD-EpiSCs could undergo a reprogramming process on hAECs and be converted to naïve-like cells converted ESCs (cESCs), in which naïve pluripotency markers were activated, and primed markers were suppressed. DNA methylation analysis also validated the epigenetic conversion from primed to naïve-like pluripotent status. The bone morphogenetic protein 4 (BMP4) is an important signaling factor in pluripotency controlling, germ cell development, and neural commitment. It showed that ESD-EpiSCs and cESCs exhibited different features toward BMP4. Our results prove that hAECs are ideal feeder cells for both naïve and primed ESCs. More importantly, the primed ESCs are allowed to be reprogrammed to naïve-like pluripotent cells on hAECs. These findings suggest that under suitable conditions primed ESCs have the potency of converting to naïve-like ESCs.
Introduction
M
Except separated directly from epiblast of developing embryos, primed pluripotent cells also could be achieved from differentiated naïve mESCs. ESC-derived epiblast stem cell (ESD-EpiSC) is an immortal cell line derived from naïve mESCs with the presence of activin A/bFGF [10]. Those cells have the same criteria as mEpiSCs derived from mouse embryos and are similar to in vivo egg cylinder-stage epiblast cells in gene expression, signaling dependence, and developing capacity. Previous study showed that ESD-EpiSCs could not be converted back into naïve mESCs when cultured with LIF instead of bFGF [10]. However, the in vitro reprogramming of mEpiSCs back to naïve pluripotency has been reported, and the requirements for this reprogramming were variously reported [11,13]. Different culturing systems applied by distinct groups might contribute to different conclusions.
In our previous work, we have successfully employed human amnion epithelial cells (hAECs) as feeder cells to maintain the undifferentiated growth of naïve mESCs and primed human embryonic stem cells (hESCs). Interestingly, the gene expression, epigenetic modification, and morphology of hESCs on hAECs differed from that on mouse embryonic fibroblasts (MEFs) [14 –16]. Amnion is a thin membrane-lined cavity, which is filled with fluid and serves to cushion the fetus during development and to prevent adhesion of the developing fetus to maternal structures. hAECs may secret specific proteins into the culturing medium, and consequently simulate the in vivo condition for stem cells to sustain pluripotency. Despite the same origin as mESCs (i.e., ICM), hESCs are distinct from mESCs. However, hESCs share the same gene expression profile and signaling dependence with mEpiSCs [17].
Based on this work, we believed that hAECs could also support the primed pluripotency of ESD-EpiSCs. More importantly, as the primed ESCs hold the converting potentials to be both naïve-like pluripotent and differentiated, investigations of ESD-EpiSCs on hAECs will offer opportunities to better understand the pluripotency determination mechanism.
Here, we sought to maintain and characterize the primed ESD-EpiSC employing hAECs as a feeder layer. Our results showed that both bFGF and LIF could stabilize the undifferentiated growth of ESD-EpiSCs, and cells exhibited distinct features under those 2 culturing conditions. Further studies proved that after being transferred from bFGF to LIF signaling, primed ESD-EpiSCs were converted to naïve-like cells. These findings provide evidences that the culturing system is critical for both naïve and primed pluripotency, which are capable of converting into each other under specific conditions without exogenous transcription factors.
Materials and Methods
Cell culture
hAECs were prepared as reported before [15,18]. Human amnion membranes, which were negative for HIV-I, hepatitis B, and hepatitis C were obtained with written and informed consent from pregnant woman in the International Peace Maternity and Child Health Hospital. The appropriate use of human amnion was approved by the institutional ethics committee. hAECs were separated from amnion membranes by 0.25% trypsin/ethylenediaminetetraacetic acid digestion for 45 min at 37°C. Freshly prepared hAECs were directly used as feeder layers when cells grown to a density of 80% confluence. ESD-EpiSCs [10] were maintained on fresh prepared hAECs in the Dulbecco's modified Eagle medium: Nutrient Mixture F-12 (DMEM:F12) medium (Invitrogen) supplemented with 2 mM
Polymerase chain reaction analysis
RNA was extracted with Trizol (Invitrogen) according to the instructions, and then treated with DNase I (Fermentas) to exclude DNA contamination. One microgram of RNAs were reverse-transcribed into cDNA with random primers by a ReverTra Aca-α- Kit (Toyobo) in a final volume of 20 μL. Reactions were performed at 37°C for 15 min, followed by 98°C for 5 min. Real-time polymerase chain reaction (PCR) was performed with SYBR Green Real-time PCR Master Mix-Plus (Toyobo) using Mastercycler ep realplex (Germany). Reaction mixtures contained a total volume of 20 μL consisting of 10 μL Master mix (2×), 2 μL plus solution, 2 μL diluted cDNA (1:10), 1.6 μL of forward and reverse primed (10 μM), and water. The PCR conditions were as follows: 95°C for 60 s, 45 cycles of 95°C for 15 s, and 60°C for 60 s. A melting curve was also performed to exclude the unspecific amplification. Primers used in this study are list in Table 1. The comparative concentration was calculated from cycle thresholds, using GAPDH as an internal standard control. Three independent experiments were performed.
Immunofluorescence staining
The immunofluorescence staining was performed as described before [18]. Oct4 was assessed with rabbit monoclonal antibodies (1:400; Santa Cruz); Nanog was detected with rabbit polyclonal antibodies (1:200; Chemicon); Klf4 was examined with rabbit polyclonal antibodies (1:100; Cell Signaling Technology); E-cadherin was detected with rabbit monoclonal antibodies (1:100; Cell Signaling Technology); Foxa2 was assessed with rabbit monoclonal antibodies (1:400; Cell Signaling Technology) and then visualized with fluorescein isothiocyanate (FITC)-conjugated goat anti-rabbit immunoglobulin Gs (IgGs) (Santa Cruz). Goat polyclonal antibodies against DAZL (1:200; Santa Cruz), Stella (1:200; Santa Cruz), and T (1:50; R&D) were used as primary antibodies, and then labeled with FITC-conjugated donkey anti-goat IgGs (Santa Cruz). Nuclei were stained with 4′6-diamidino-2-phenylindole (DAPI). Activity of alkaline phosphatase (AKP) was detected by the BCIP/NBT Alkaline Phosphatase Color Development Kit (Beyotime Institute of Biotechnology) according to the instruction.
In vitro differentiation assay
Embryoid bodies (EBs) were achieved by growing cell clumps for 8 days with ultra-low attachment dishes (Corning) in the differentiating medium: DMEM:F12 (Invitrogen) with 2 mM
Single-cell cloning efficiency assay
Colonies were picked and dissociated into single cells with 0.25% trypsin (Invitrogen) at 37°C. 1×104 cells were seeded onto fresh prepared hAECs feeder layers in one well of a 6-well plate (Corning). Each cell line was analyzed in triplicate. The medium was changed every day. Colonies were counted 5 days later. Average and standard deviation were calculated.
DNA methylation analysis
The CpGs methylation of promoter regions of Rex1 and Stella was analyzed by bisulfite sequencing analysis. Genomic DNA was extracted with the Genomic DNA Extraction Kit (Sangon Biotec) according to the manual, and then treated with bisulfite as reported before [20]. After DNA purification, promoter regions were amplified by PCR with primer pairs as described before [20]. DNA fragments were then cloned into the pUC18-T vector (Sangon Biotec). Ten white clones were randomly picked from IPTG/X-gal plat and sequenced with the M13 primer.
Karyotype analysis
Cells were arrested at metaphase by incubation with 40 μg/mL colcemid (Sigma) for 1 h at 37°C. Cells were collected by trypsinization, and then resuspended in a hypotonic solution (0.075 M KCl) for 30 min at 37°C. After centrifugation, the pellet was fixed with cold methanol/acetic acid (3:1) 3 times before dropping onto cold microscope slides. After being incubated in 0.005% trypsin for 5 s at 37°C, slides were stained with 5% Giemsa for 10 min. More than 20 metaphase spreads were analyzed.
Results
hAECs could support the undifferentiated growth of ESD-EpiSCs
Naïve and primed ESCs require antagonistic signal pathways to maintain their pluripotency. While naïve ESCs need LIF and bone morphogenetic protein 4 (BMP4) signaling to maintain the pluripotency, primed ESCs rely on bFGF and Activin A/Nodal signaling [4,5,21]. It has been proved that ESD-EpiSCs have the same characteristics as mEpiSCs derived from mouse embryos, and their pluripotency can be stabilized in a chemically defined medium containing Activin A and bFGF (CDM/AF) [10].
First, we asked whether hAECs could be used as feeder layers to maintain the pluripotency of ESD-EpiSCs. In some case, primed mEpiSCs could survive and be reprogrammed to a naïve-like state with LIF signaling [11]; however, the undifferentiated growth of ESD-EpiSCs could not be maintained when exposed to LIF signaling no matter with or without MEF as the feeder layer [10]. When we maintained ESD-EpiSCs on hAECs with bFGF, we also tried to culture them with LIF on hAECs. Figure 1A shows that on hAECs, homogenous colonies of ESD-EpiSCs were obtained with bFGF, and those domed colonies were slightly different from the flat monolayer colonies on MEF as reported [10]. Compact colonies were picked, dissociated, and then seeded onto hAECs with LIF. Cells were split every 3–4 days. Interestingly, clean bounded colonies could be formed with LIF signaling, and inside cells showed high nuclear–cytoplasmic ratios (Fig. 1B). Colonies in LIF were smaller than original ESD-EpiSC colonies in bFGF, and the aggregation were better rounded. After being extensively propagated for more than 70 generations, colonies under both conditions do not change obviously. At the same time, naïve mESCs (R1) were also maintained on hAECs with LIF signaling (Fig. 1B).

The undifferentiated growth of EDS-EpiSCs could be stabilized on hAECs with either bFGF or LIF.
Nanog and Oct4 are 2 crucial transcription factors, which mark pluripotent cells and constitute the central loop for pluripotency controlling [22,23]. As shown in Fig. 1C, no matter with bFGF or LIF, Nanog and Oct4 could be clearly detected by immunostaining in ESD-EpiSCs on hAECs. R1 cells were employed as a positive control. The expression of those pluripotency markers suggested that the undifferentiated growth of ESD-EpiSCs could be supported on hAECs. To further characterize ESD-EpiSCs on hAECs, their developing capability was assessed in vitro.
It has been shown that ESD-EpiSCs were able to differentiate into derivatives of all 3 primary germ layers under the standard culturing condition [10]. To determine if the ESD-EpiSCs on hAECs retained a developing potential after extensively propagated with bFGF or LIF, we performed the EB differentiation assay. EBs were obtained by growing small cell clumps in suspension as aggregates for 8 days. Spheroidic EBs were formed from these 2 kinds of cells in absence of any growth factors (Fig. 2A). EBs were then transferred to gelatin-coated culture dishes, allowed to attach for 1 day, and differentiate for another 7 days. Under a microscope, somatic cells with distinct morphology could be observed (Fig. 2B). Cells were harvested for RNA extraction, and germ layer marker genes were detected by reverse transcription-PCR (Fig. 2C). It showed that markers of all 3 germ layers could be detected, including Sox1, Ck18, and CK19 (ectodermal), Flk1, Sox17 (mesodermal), Gata4 (endodermal), and Eomes (trophectodermal). Results demonstrated that under both maintaining conditions (with bFGF or LIF) ESD-EpiSCs retained a wide developing potency. Again, it indicated that on hAECs, both bFGF and LIF could support the undifferentiated growth of ESD-EpiSCs.

ESD-EpiSCs were able to differentiate into 3 primary germ layers in vitro.
ESD-EpiSCs exhibited distinct features after being transferred to LIF signaling
Under the traditional condition, undifferentiated ESD-EpiSCs could not be supported with LIF [10]. However, ESD-EpiSCs could be maintained with LIF on hAECs. To better define the molecular properties of ESD-EpiSCs on hAECs, we analyzed their expression profile of pluripotent marker genes. For the ESD-EpiSCs cultured with LIF signaling, we collected cells at 7, 14, 28, and 35 days, respectively, after prolonged culture with LIF, and performed the real-time PCR compared with cells maintained with bFGF (Fig. 3A).
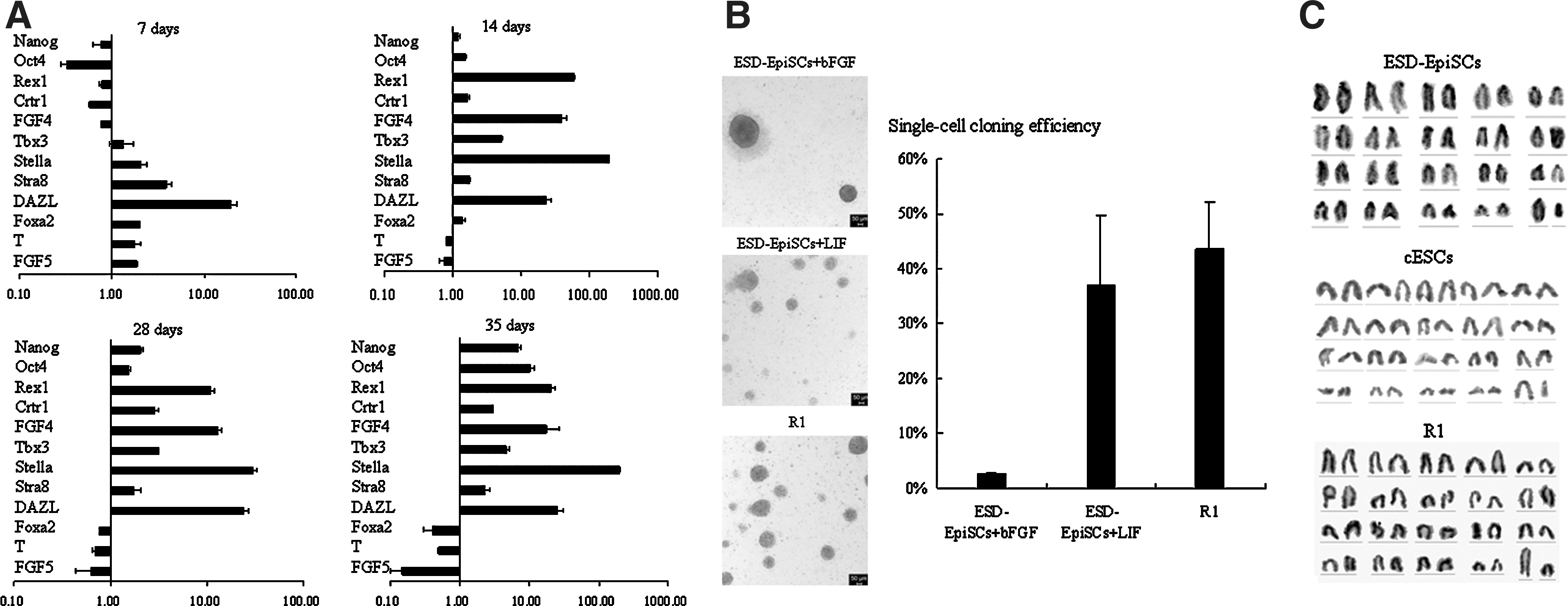
Genetic and biological features of ESD-EpiSCs were changed when switched from bFGF to LIF signaling on hAECs.
It has been demonstrated that Oct4 and Nanog were expressed at equivalent levels in both naïve and primed ESCs [4,5,12]. Consistently, ESD-EpiSCs express the core pluripotent genes (Oct4 and Nanog) no matter with bFGF or LIF signaling. However, during the first week after switching to the LIF medium, the expression of Oct4 and Nanog was repressed. On the other hand, Foxa2, T (also known as brachyury), and FGF5, which are markers of primed pluripotency and early germ layers were induced after the first week. The expression of pluripotent markers was reduced, which maybe because most cells were differentiated when switched to the LIF medium and only a small amount of cells were still grown as compact colonies. Following continuing passage, pluripotent cells selectively survived, and the expression of Nanog and Oct4 was recovered (14 days), and then even higher (28 days) than the original cells cultured with bFGF.
Rex1, Crtr1, FGF4, and Tbx3 are specifically expressed in naïve ESCs and ICM, while Foxa2, T, and FGF5 are associated with primed ESCs and the postimplantation embryo [4,5,10,11]. It has been demonstrated that in CDM/AF, ESD-EpiSCs strongly expressed the primed markers, but not the naïve markers [10].
After propagating for 14 days, the transcripts of Rex1, Crtr1, FGF4, and Tbx3 were strongly detected in cells cultured with LIF, but not in original cells maintained with bFGF signaling (Fig. 3A). At the same time, the expression of Foxa2, T, and FGF5 were downregulated, and then further suppressed after culturing with LIF for 28 days. However, the gene expression pattern of ESD-EpiSCs stabilized with bFGF did not change obviously. DAZL, Stella, and Stra8, which are associated with germ line cells, are expressed in naïve mESCs, and could be barely detected in primed mEpiSCs [5]. As analyzed by real-time PCR, the expression of DAZL, Stella, and Stra8 increased significantly after being cultured on hAECs with LIF for 1 week (Fig. 3A).
While naïve mESCs could survive as single cells, primed mEpiSCs are sensitive toward trypsin treating; and the growth rate of mEpiSCs is lower than mESCs [4,5]. ESD-EpiSCs could not survive as single cells either. Consequently, cells were dissociated with collagenase IV or by mechanical separation [10]. During propagating, we observed that the generation time of ESD-EpiSCs was shorter with LIF than with bFGF. Normally, ESD-EpiSCs were passaged every 3–4 days with bFGF, whereas cells had to be propagated every 2 days when supplied with LIF. To further characterize ESD-EpiSCs maintained under LIF signaling, the single-cell cloning efficiency was determined after trypsin treatment.
After being treated with trypsin, cells were seeded at the same density onto fresh prepared hAECs feeder layer with bFGF or LIF. The medium was changed every day. Colonies were counted after 5 days. As shown in Fig. 3B, under bFGF signaling, a few flat monolayer colonies (∼3%) were formed after trypsin treating. However, with LIF, the single-cell cloning efficiency was much higher (∼37%) (Fig. 3B) and the colonies were still kept as compact domed aggregations (Fig. 3B).The tolerance to trypsin further indicated that after being transferred to LIF signaling, ESD-EpiSCs share similar biological features with naïve mESCs other than original ESD-EpiSCs.
To distinguish those 2 kinds of cells, cells stabilized with LIF were named converted ESCs (cESCs). The chromosomal stability of ESD-EpiSCs and cESCs has been confirmed by karyotype detection. ESD-EpiSCs (passage ∼30) and cESCs (passage ∼25) were harvested for karyotype assess by G-banding examination. Twenty metaphase spreads were examined for each cell line, and over 80% spreads showed a normal karyotype (Fig. 3C). Up to now, cESCs have been extensively passaged for over 70 generations. After freezing and thawing in liquid nitrogen, their morphology and gene profile did not change obviously, suggesting that the cESCs have been stabilized as independent cell lines.
ESD-EpiSCs and cESCs were independent cell lines
To validate that cESC was an individual cell line, we compared the expression of naïve and primed marker genes at the protein level. As reported before, Klf4 and E-cadherin are specifically expressed in naïve cells, while Foxa2 and T are restricted in primed cells [4,5,11,13,24]. Germ cell markers, such as DZAL and Stella, are expressed in naive mESCs, other than in primed EpiSCs [4,5].
We examined the expression of those markers by immunostaining (Fig. 4A). Klf4 and E-cadherin were expressed in cESCs and R1 cells, but could not be detected in ESD-EpiSCs. On the other hand, Foxa2 and T were limitedly expressed in ESD-EpiSCs. As shown in Fig. 4A, DAZL and Stella were clearly detected in cESCs and R1 cells, whereas they were not expressed in ESD-EpiSCs. These findings indicated that naïve marker genes were activated and primed marker genes were depressed in cESCs. Those results indicated that the gene expression pattern of cESCs was distinct from ESD-EpiSCs, but alike to naïve R1 cells.
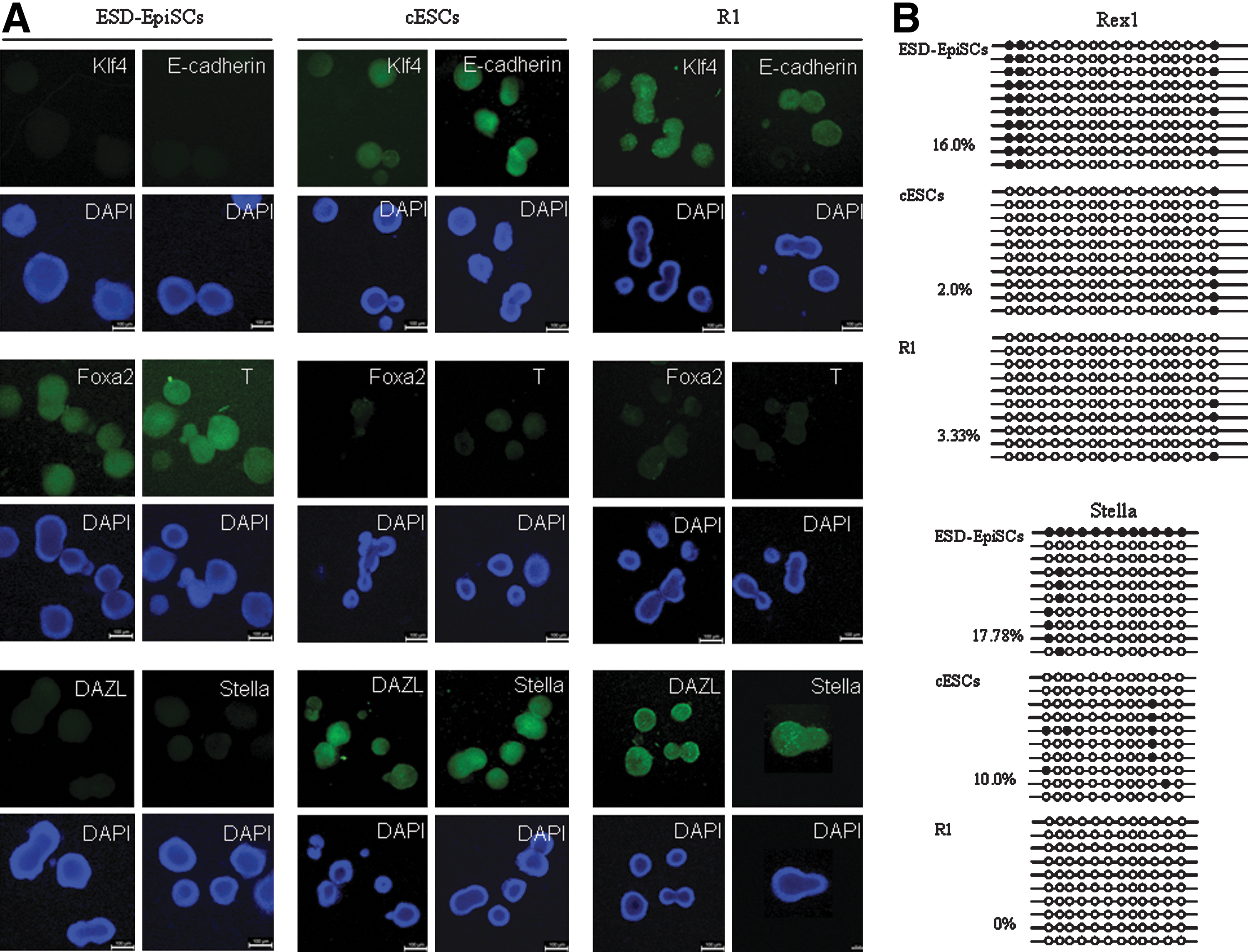
cESCs resembled naïve R1 cells in genetic and epigenetic characterizations. EDS-EpiSCs were cultured with bFGF; cESCs and R1 were maintained with LIF on hAECs (passage 25–30). Scale bars, 100 μm.
We then wondered whether the differences in gene expression resulted from distinct epigenetic modifications. DNA methylation on promoter regions of Rex1 and Stella was examined by bisulfite sequencing PCR. Consistent with reported before [11,12], promoters of Rex1 and Stella were hypo-methylated in naïve R1 cells (Fig. 4B). Compared to original ESD-EpiSCs, cESCs also showed reduced methylation, revealing that the promoters of Stella and Rex1 were de-methylated after transferred to LIF signaling. Those findings confirmed that ESD-EpiSCs and cESCs were 2 indistinguishable cell lines, and that cESCs represented naïve-like pluripotency.
ESD-EpiSCs and cESCs showed distinct features upon exposing to BMP4
BMP4 is a critical signal in pluripotency maintaining [25 –27], neural commitment [10,22,28,29], and primordial germ cell (PGC) differentiation [30]. BMP4 exhibits antagonistic functions in naïve and primed ESCs [4,27]. BMP4 plays a positive role in self-renewal [22,27,28] and a negative role on neural commitment of naïve mESCs [25,28,29], whereas BMP4 promotes ESD-EpiSCs to differentiate into non-neural lines and inhibits the neural commitment [4,5,10].
To determine the different characteristics of ESD-EpiSCs and cESCs in detail, we analyzed the effection of BMP4 on those cells. Cells were induced with BMP4 (20 ng/mL) for 3 days on hAECs. Consistent with reported, after being induced with BMP4 for 3 days, ESD-EpiSCs started to differentiate: clones became flattened and differentiated cells expanded outside the periphery. The activity of AKP that marks the pluripotent cell was absent in those cells (Fig. 5A).

ESD-EpiSCs and cESCs represent different properties upon exposing to BMP4 signaling. Scale bars, 100 μm.
On the other hand, cESCs and R1 clones were still compact and exhibited high AKP activity after induced by BMP4 for 3 days (Fig. 5A). It indicated that BMP4 was positive for the undifferentiated growth of cESCs.
To further assess the effect of BMP4 on ESD-EpiSCs and cESCs, gene expression profile was analyzed by real-time PCR (Fig. 5B). As discussed above, pluripotency markers, such as Nanog and Oct4, expressed at a high level in undifferentiated ESD-EpiSCs and cESCs (Fig. 1B). The addition of BMP4 did not modify the expression of pluripotency marker genes in cESCs and R1. The expression of FGF4, which was limitedly expressed in cESCs and R1 cells, was even lightly increased after BMP4 treatment (Fig. 5B). In contrast, the expression of those pluripotency markers (Nanog, Oct4, and Sox2) was depressed by BMP4 in ESD-EpiSCs. FGF5, the specific marker for primed pluripotency, was downregulated by BMP4 in ESD-EpiSCs (Fig. 5B). At the same time, differentiating markers (CK18, CK19, Flk1, Sox17, Gata4, Gata6, and Eomes) were activated in ESD-EpiSCs, but not in cESCs or R1 cells (Fig. 5B). It indicated that BMP4 addition induced ESD-EpiSCs to differentiate.
Upon exposure to BMP4, primed EpiSCs are primed for differentiating into PGC-like cells, and germ cell markers (such as DAZL and Stella) are readily activated [5,31]. As shown in Figs. 3A and 4A, those PGC markers were absent in ESD-EpiSCs, nevertheless, they could be detected in cESCs and R1. To determine the effect of BMP4 on ESD-EpiSCs and cESCs, the expression of marker genes was examined by real-time PCR (Fig. 5B). Addition of BMP4 led to an enhanced expression of DAZL and Stella in ESD-EpiSCs, while BMP4 did not upregulate those markers in cESCs or R1. It suggested that ESD-EpiSCs had the potential of differentiating to PGCs upon BMP4 treatment, while cESCs and R1 were resistant to this induction.
As demonstrated before, BMP4 promotes non-neural lineage differentiation and suppresses the neural commitment of ESD-EpiSC [10]. Although the addition of BMP4 induced ESD-EpiSCs to differentiate, the expression of neural markers (Hesx1, Sox1) were inhibited (Fig. 5B), suggesting that BMP4 supressed neural commitment. Similar to naïve R1, the expression of neural markers did not change obviously in cESCs upon BMP4 supplement.
Together, results indicated that toward BMP4, cESCs shared a similar response with naïve mESCs, being distinct from primed ESD-EpiSCs. It suggested that a reprogramming process occurred in ESD-EpiSCs after transferred from bFGF to LIF signaling, and the converted cells (cESCs) gained the naïve-like phenotypes.
Discussion
Immortalized naïve and primed ESCs correspond to distinct in vivo development stages, and are maintained in vitro with LIF or bFGF signaling, respectively [4,5]. ESD-EpiSC is a primed cell line derived from naïve mESCs through programming mESCs with bFGF/Activin A, and could be maintained in the same condition [10]. When exposed to LIF signaling, ESD-EpiSCs differentiated no matter with or without MEF as feeder cells [10]. hAECs have been successfully employed as feeder cells to support the undifferentiated growing of naïve mESCs and primed hESCs [14,18]. The biological and molecular features of hESCs on hAECs were distinct from traditional hESCs maintained on MEF [14]. The culture system is one essential issue for stem cell studying. Both ESD-EpiSCs and hAECs provide valuable molds to study how maintaining conditions affect the pluripotent status of ESCs in vitro.
In this work, we tried to maintain ESD-EpiSCs on hAECs with bFGF or LIF signaling. Both bFGF and LIF signaling could maintain the undifferentiated growth of ESD-EpiSCs on hAECs. Differs from the flat monolayer colonies of primed ESCs as reported [4,5,10], ESD-EpiSCs could form compact domed colonies on hAECs. Analysis of gene expression showed that the pluripotency markers, such as Nanog and Oct4, are expressed in ESD-EpiSCs maintained on hAECs. EBs of ESD-EpiSCs could be obtained and were capable of differentiating into all 3 germ layers in vitro. It indicated that hAECs could maintain the undifferentiated state and development potentials of ESD-EpiSCs with bFGF or LIF.
Our results showed that the gene expression patterns of ESD-EpiSCs were distinct under different culturing conditions. Consistent with previous reports, the primed markers were expressed and the ICM markers were absent in ESD-EpiSCs when cultured with bFGF on hAECs. This further validated that hAECs could maintain the primed pluripotency of ESD-EpiSCs supplemented with bFGF. Interestingly, when cells were stabilized and passaged in the LIF medium on hAECs, the naïve pluripotent markers were upregulated, while the primed markers were downregulated. The distinct genetic properties suggested that ESD-EpiSCs were reprogrammed into a new cell line (named cESC) upon transferring to LIF signaling.
Epigenetic studies showed that the Stella and Rex1 promoters of cESCs were de-methylated, resulting in similar modification as naïve R1 cells. Further examinations demonstrated that cESCs shared similar features with R1 upon exposure to BMP4 signaling. It was apparent that cESC was a distinguishable cell line from original ESD-EpiSCs, and cESCs represent the naïve-like pluripotency.
Those findings demonstrated that on hAECs primed ESD-EpiSCs could convert to naïve-like pluripotency just upon switching the culture medium. It suggests that under suitable conditions both primed and naïve stem cells are capable of converting to each other, and this program is reversible. However, in other works, the primed mESCs are stable cell lines that could not be maintained in the traditional naïve mESC medium without genetic manipulations [4,5,13]. Nevertheless, it also reported that primed mESCs were able to convert to a ground pluripotency state through switching to LIF signaling without the exogenous transcription factor [11]. Different genetic background of individual cell lines may contribute to this contract. However, even the ESD-EpiSCs could not be maintained with LIF in CDM [10]. It indicates that hAECs offer a proper environment to ESD-EpiSCs for reprogramming to the naïve-like pluripotency.
HAECs develop from the epiblast by 8 days after fertilization before gastrulation, and retain the premature properties. Cultured hAECs themselves retain pluripotent characteristics [32]. Since hAECs do not express telomerase and can only be expanded for limited passages, hAECs feeder layers were not inactivated by mitomycin C or with gamma irradiation. Fresh hAECs may secrete some proteins into the culture medium, or they may provide some membrane proteins for undifferentiated growth of stem cells. In the human amniotic membrane, certain growth factors were detected: epidermal growth factors, keratinocyte growth factor, hepatocyte growth factor, bFGF, TGF-α, TFG-β1, TFG-β2, and TFG-β3 [33]. Cultured hAECs have been proved to release acetylcholine, catecholamines, and neurotrophins [34]. In our previous studies, we have proved that hAECs could secret LIF highly into the medium [14,18]. Released factors might participate the pluripotency controlling pathway, and consequently, promote ESD-EpiSCs to be ready to reprogram upon specific inductions. However, identifying the critical factor(s) and uncovering its (their) mechanism in pluripotency controlling need further studies.
Despite the common in vivo origin as mESCs, hESCs resemble many properties as primed mESCs and exhibit primed pluripotency under the traditional artificial system [6,24,35]. It has been reported that the XaXa state, which marks the ground pluripotency, could be established through being cultured under physiological oxygen [36]. This observation suggests that hESCs also possess the potential of naïve pluripotency. Defining an ideal in vitro system for hESCs to obtain the naïve hESCs is an essential issue for the clinical therapeutic application. Employing human origin hAECs could avoid the risks of zoonosis, and this is required for obtaining the clinical-grade hESCs. Additionally, as human placental amnion is abundantly available as routinely discarded tissues, there are no ethical and practical limitations to prepare hAECs as the feeder layer. Here, our data provide evidences that primed stem cells could be reprogrammed to naïve-like cells on hAECs. Taken together, hAECs feeder layer is an ideal in vitro maintaining system for hESCs in cell transplantation and regenerative medicine.
Footnotes
Acknowledgments
The work was surppoted by grants from the National Natural Science Foundation of China (No. 81070533), and Laboratory of Molecular Cell Biology, Chinese Academy of Science, Shanghai; the Shanghai Jiao Tong University School of Medicine (No. 11xj21061), and the International Peace Maternity and Child Health Hospital. We are especially grateful to Professor Naihe Jing for providing the ESD-EpiSC cell line. We are also grateful to Dr. Lingyu Li for helpful discussions.
Author Disclosure Statement
No competing financial interests exist.