
Abstract
Endothelial cells (EC) generated in vitro from stem cells are desirable for their potential in a variety of in vitro models and cell-based therapeutic approaches; however, EC can take on a number of functionally and phenotypically distinct specializations. Here, we show the generation of functionally distinct EC subpopulations, including (1) the pro-angiogenic migrating tip-like and proliferative stalk-like EC, and (2) the less migratory cobblestone-shaped phalanx-like EC. Both embryonic stem cell (ESC)-derived EC subpopulations are generated from outgrowths of Flk-1+ vascular progenitor cells with high levels of vascular endothelial growth factor treatment, while the phalanx-like ESC-derived EC (ESC-EC) are subsequently isolated by selecting for cobblestone shape. Compared with the ESC-derived angiogenic endothelial cells (named ESC-AEC) that contain only 14% Flt-1+ and 25% Tie-1+ cells, the selected phalanx-like ESC-EC express higher numbers of cells expressing the phalanx markers Flt-1+ and Tie-1+, 89% and 90%, respectively. The ESC-AEC also contain 35% CXCR4+ tip cells, higher expression levels of stalk marker Notch-1, and lower expression levels of Tie-2 compared with the phalanx-type ESC-EC that do not contain discernible numbers of CXCR4+ tip cells. Perhaps most notably, the ESC-AEC display increased cell migration, proliferation, and 3 times more vessel-like structures after 48 h on Matrigel compared with the phalanx-like ESC-EC. This work analyzes, for the first time, the presence of distinct EC subtypes (tip/stalk, and phalanx) generated in vitro from ESC, and shows that phalanx-like EC can be purified and maintained in culture separate from the tip/stalk-like containing EC.
Introduction
T
The first specialized EC type within a sprouting vessel is called the tip EC. Tip EC are found at the leading edge of a sprouting vessel, and are distinct from other EC in their DLL4/Notch 1 signaling [20]. It is thought that the expression of Notch Delta-like ligand 4 (Dll4) in tip cells functions by suppressing tip cell fate of neighboring stalk cells via Notch signaling [21]. Other tip cell markers include CXCR4 [22] as well as receptors for axon guidance cues, such as the Netrin receptor UNC5B [23] and neuropilin-1 [24]. Tip cells also exhibit more organized stress fibers with numerous probing filopodia, and readily migrate toward an angiogenic stimulus [25], but do not form lumens and proliferate minimally [19,26,27]. Microarray analysis has shown that tip cells secrete a number of molecules thought to bind to associated receptors on the neighboring stalk cells, including angiopoietin-2 (Ang-2) [21]. The same study also identified the upregulation of various proteases in tip cells, especially uPAR, suggesting that production and release of uPAR by tip cells might also play a role in tip cell migration by facilitating matrix degradation at the leading edge of the vessel [21].
The stalk cells are known to trail behind the tip cells during angiogenic sprouting, forming the stalk of the sprout. These cells are predominantly defined by their morphological position behind the tip cell, however, and Notch signaling from the tip cells dampen the vascular endothelial growth factor (VEGF)-induced expression of Dll4 on stalk cells [20], enabling the tip cells to maintain their position at the leading edge of the sprouting vessel. Unlike tip cells, stalk cells readily proliferate, form lumens, and lay down extracellular matrix, but do not extend filopodia [27]. The Ang-2 receptor, Tie-2, is also found to be expressed by stalk cells, while not detectable in tip cells [21].
There is also a third population of less migratory EC, called a phalanx EC [28], which has been identified more recently. These cells are distinct in their cobblestone-shaped morphology, lower migratory and proliferate rates, and high levels of soluble and membrane-bound Flt-1[28]. Flt-1 is thought to mitigate the otherwise proangiogenic signals of VEGF [28], keeping the phalanx EC in a stable morphology. This is aided by the expression of VE-cadherin, which tightens the EC-to-EC adhesions, promoting a more quiescent phenotype. The phalanx cells also respond to VEGF in a different manner than the tip cells. Although phalanx-type EC are capable of responding to VEGF signaling, VEGF signaling in phalanx EC acts as an apoptosis rescue from serum-deprived conditions, rather than as the proliferative and mitogenic response seen in tip cells [28].
Based on these descriptions of distinct EC within the sprouting vessel, we set out to verify the populations of tip/stalk EC and phalanx EC, named angiogenic ESC-AEC and ESC-EC, respectively, within our cell populations derived, in vitro, from mouse ESC. We also purify the phalanx-like EC from the mixed EC cultures and show that these EC maintain their distinct subphenotype.
Materials and Methods
Cell culture
R1 murine ESCs were maintained on 0.5%-gelatin coated plates in a serum-free medium containing knockout Dulbecco's modified Eagle's medium (DMEM; Invitrogen), 15% knockout serum replacement (KSR; Invitrogen), 1×penicillin–streptomycin (Invitrogen), 1×nonessential amino acids (NEAA; Invitrogen), 2 mM
Cell proliferation
EC were harvested and resuspended in the LDSF medium and plated onto fibronectin-coated wells in a 6-well plate. Approximately 200,000 cells were seeded into each well. This process was repeated with MCEC onto gelatin-coated wells. Cells were allowed to expand for 72 h. They were subsequently harvested and counted using a hemacytometer. Using propidium iodide (PI), the cell cycle stage was also analyzed. Subconfluent cells were removed with dissociation buffer and fixed with −20°C absolute ethanol. After washing twice with phosphate-buffered saline (PBS), the cells were suspended in 500 μL of the PI solution: 50% RNase A (Qiagen), 50% PI (Sigma), and 1% Triton X-100 solution per million cells, for 40 min before FACS analysis.
In vitro angiogenesis
In a 24-well plate, 300 μL of growth factor-reduced Matrigel was added to each well and allowed to gel at 37°C. After 30 min, 50,000 cells were added to each well. The cells were observed at 12, 24, and 48 h and imaged using a phase-contrast microscope with a 25×objective (Fisher) using Micron software.
Cell migration
The EC were harvested from fibronectin-coated dishes, pelleted, and re-suspended in the EGM-2 medium (Lonza) without serum. Approximately 200,000 cells were added to a BD FluoroBlok transwell insert (BD) and placed in a well of a 24-well plate. In order to facilitate migration of the EC, 500 μL of EGM-2 with 10% FBS with and without 100 ng/mL of Ang-2 was added to the wells below each transwell. After 24 h, the inserts were fixed, permeablized, and fluorescently stained with Phalloidin 488 (Invitrogen) and 4′,6-diamidino-2-phenylindole (DAPI). The transwells were then imaged using a Leica DMI6000 scope. Migration was measured by counting the number of intact nuclei in the field of view. A Student's t-test was used to compare the migration of cells in both conditions.
Immunofluorescent analysis
Glass microscope slides were coated with fibronectin and seeded with either ESC-AEC or ESC-EC. For cytoskeletal analysis, the slides were then fixed, permeablized, and fluorescently stained with either intracellular phalloidin 488 (Invitrogen) and DAPI. For cell–cell adherens expression, the cells were first blocked with 5% donkey serum and then stained with extracellular (ie, the cells were not permeablized) goat anti-VE-cadherin (Santa Cruz) or goat anti-PECAM-1 (M-20; Santa Cruz) followed by a FITC-conjugated donkey anti-goat IgG (Abcam) and counterstained with DAPI. Coverslips were then mounted on the glass slides using mounting medium (Dako) and imaged with either a Leica DMI6000 scope or a confocal laser-scanning microscope.
Flow cytometry
The ESC-AEC and ESC-EC were harvested and stained for HSP27 phosphorylation at serine 82 [29], smooth muscle marker, calponin, fibroblast marker, DDR-2, or mesenchymal marker, vimentin. Briefly, the cells were isolated using cell dissociation buffer (Gibco), pelleted, fixed, and permeablized using ice-cold methanol. The cells were then incubated with pHSP27 (Santa Cruz), anti-calponin (Santa Cruz), goat anti-DDR2 (N-20; Santa Cruz), or rabbit anti-vimentin (Abcam) followed by anti-goat FITC secondary antibody (Abcam) or donkey anti-rabbit PE (Fitzgerald). Additional cell characterization for potential differences in tip, stalk, and/or phalanx surface markers (ie, no permeablization) included (1) FITC-conjugated mouse monoclonal IgG1 Notch-1 and FITC-conjugated IgG1 isotype control (Abcam), (2) PE-conjugated anti-mouse DLL4 with PE-conjugated armenian hamster IgG isotype control (BioLegend), (3) rabbit polyclonal CXCR4 IgG (Abcam), donkey anti-rabbit PE (Fitzgerald) and PE-conjugated IgG isotype control (Abcam), and (4) PE-conjugated rat anti-mouse Tie-2 with rat IgG1 isotype control (BioLegend). We also examined potential differences between extra- and intracellular Notch1 by permeablizing with 0.7% Triton X-100 for 10 min before staining. The cells were then analyzed using a BD LSRII. The results were analyzed using FlowJo software (TreeStar).
In vivo angiogenesis
Fertilized eggs were generously donated from a local hatchery (Waterford, CA) at day 2 postfertilization. On day 3 postfertilization, the eggs were windowed to facilitate visualization. First, a small hole was made at the large blunt end of the egg at the air sac. A 16-gauge needle with fitted syringe was inserted into the hole to remove 4–5 mL of albumenal fluid, thus creating a false air sack on the long side of the egg for cutting a small window. Clear tape was then placed over both the hole and window and the eggs were placed in a 37°C incubator with 1% CO2.
On day 6 postfertilization, a Matrigel scaffold was generated as the vehicle for cell transplantation. Briefly, 40 μL of Matrigel was mixed with 5,000 EC and allowed to gel for 30 min at 37°C on top of coverslips. The coverslips were then placed gel-side down onto the CAM. Controls also included a blank coverslip and cell-free Matrigel.
On day 10 postfertilization, the vascular pattern of the CAM was imaged using a Leica stereoscope using Axiovision software. These images were manually skeletonized using Adobe Photoshop and subsequently analyzed with Angioquant software. Tissue was also harvested from the CAM assays on day 10 postfertilization and fixed in 4% formaldehyde. The samples were then incubated in 0.3% H2O2 in methanol, rinsed for 3 min in PBS, and incubated in buffer (0.1% bovine serum albumin in PBS) for 20 min. The samples were then rinsed in PBS and incubated in the buffer with an anti-mouse horseradish peroxidase (Abcam)-conjugated antibody at room temperature for 30 min. The tissue was rinsed again with PBS and subsequently stained with 3,3-diaminobenzidine substrate kit (Abcam).
Statistical analysis
Student's t-tests were used to analyze statistically significant differences between experimental groups with statistical significance at P<0.05.
Results
EC markers
Although both ESC-AEC and ESC-EC contain high levels Flk-1+ and VE-cadherin+ cells, the ESC-AEC contained reduced numbers of Flt-1+ (14%) and Tie-1+ (25%) compared with purified ESC-EC (Fig. 1).
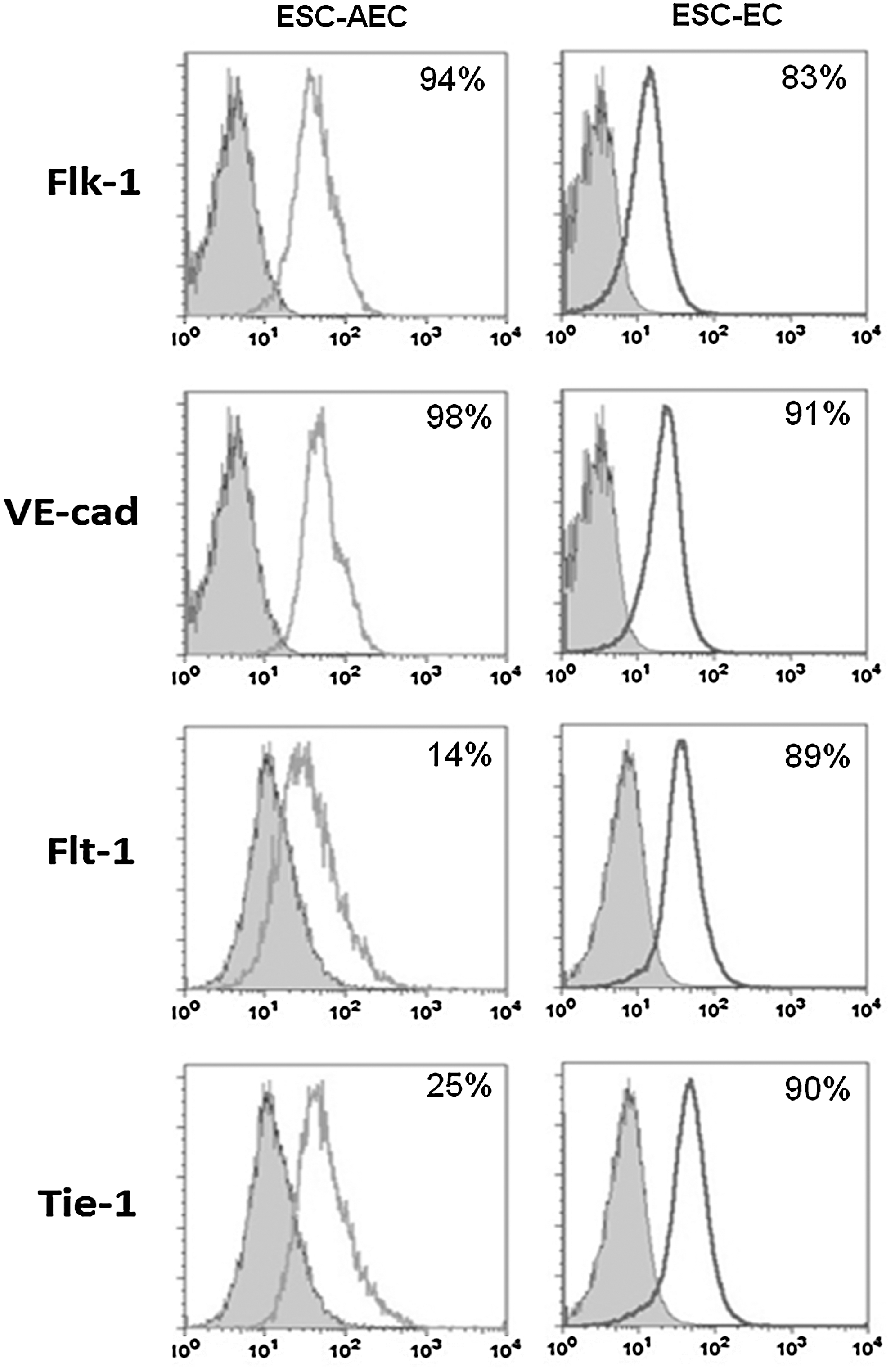
EC derived in chemically defined conditions express appropriate endothelial makers. We examined the expression of endothelial markers Flk-1, VE-cadherin, Flt-1, and Tie-1. The histograms include the ESC-AEC (left) and the purified ESC-EC (right) derived using R1 ESC under chemically defined conditions. EC, endothelial cell; ESC, embryonic stem cell; AEC, angiogenic endothelial cells.
Cell morphology and proliferation
Looking more closely at these 2 EC subpopulations, it is immediately apparent that they exhibit distinct morphologies in tissue culture (Fig. 2A–D). The ESC-AEC form lines and loops that do not collapse at confluence (Fig. 2C), while the purified ESC-EC display the distinct cobblestone shape (Fig. 2D) described for phalanx EC, and continued to exhibit this phalanx-like morphology after several passages. The ESC-AEC are also more proliferative in culture compared with ESC-EC. This is evident in Fig. 2A, where over half the cells in the culture dish are rounded up as single and double cells as if they are undergoing or have just completed mitosis, striking comparison to the ESC-EC (Fig. 2B), where only about 10% of the cells appear to be actively dividing. When quantified for differences in the proliferation rates of these 2 cell populations, the average proliferation rates for ESC-AEC were much greater (with an approximate doubling time of 18 h) compared with the ESC-EC (over 25 h) (Fig. 2E). From the cell cycle analysis, the majority of the ESC-AEC were confirmed to be in S-phase (Fig. 2F), while most of the ESC-EC were in G0 (Fig. 2G).

Cell morphology and proliferation of ESC-AEC and ESC-EC. Differences in cell morphology were very apparent between the ESC-AEC
In vitro angiogenesis
Although the ability of these ESC-derived EC to form vascular networks exhibiting loops and lumens in 3-dimensional collagen gels has been previously reported [30], the angiogenic cells should exhibit increased migration (tip) as well as the increased proliferation (stalk) compared with purified phalanx-like EC. Therefore, the ability for these cells to undergo angiogenesis and vasculogenesis in vitro was directly compared. After seeding onto Matrigel, both EC populations migrate into cord-like structures with some additional sprouting within 12 h (Fig. 3A, B). However, after 48 h (Fig. 3C, D), the ESC-EC are regressing into cell clumps and are undergoing apoptosis (a normal part of vessel regression, verified previously [31]), while the ESC-AEC continue to proliferate and sprout robustly for over 48 h, generating 3 times more vessel-like structures (Fig. 3E), and remained stable for up to 5 days (not shown).

Increased migration and sprouting activity in ESC-AEC compared with ESC-EC.
Cell migration
The EC subpopulations were then examined for migration in response to a (1) general nutrient or (2) specific chemotactic chemical signal. For these studies, FBS was used as a general nutrient attractant, and Ang-2 was used as the specific chemotactic migration signal. As expected, all cells migrated through a Boyden chamber transwell (Fig. 3F) toward both the FBS nutrient and Ang-2, exhibiting a stronger response to Ang-2 compared to the FBS (Fig. 3G). Although the addition of angiopoeitin-2 increased the migration activity by ∼1.7-fold in the ESC-EC, the ESC-AEC migrated only sparingly toward the FBS attractant while migrating more strongly toward the specific angiopoeitin-2 chemokine, resulting in a much larger, 5-fold, increase in migration compared with FBS alone.
Because endothelial and smooth muscle cells (SMC) are known to be derived from a common Flk-1+ progenitor cell [15] and studies have implicated a role for SMC or fibroblast coculture in increased angiogenesis [32,33], the cells were also examined for the presence of calponin+ vascular SMC, DDR2+ fibroblasts, or vimentin+ mesenchymal cells. The flow cytometry histograms indicate that neither ESC-EC nor ESC-AEC subpopulations contained significant numbers of any of these potentially contaminating cell types (Supplementary Fig. S1; Supplementary Data are available online at
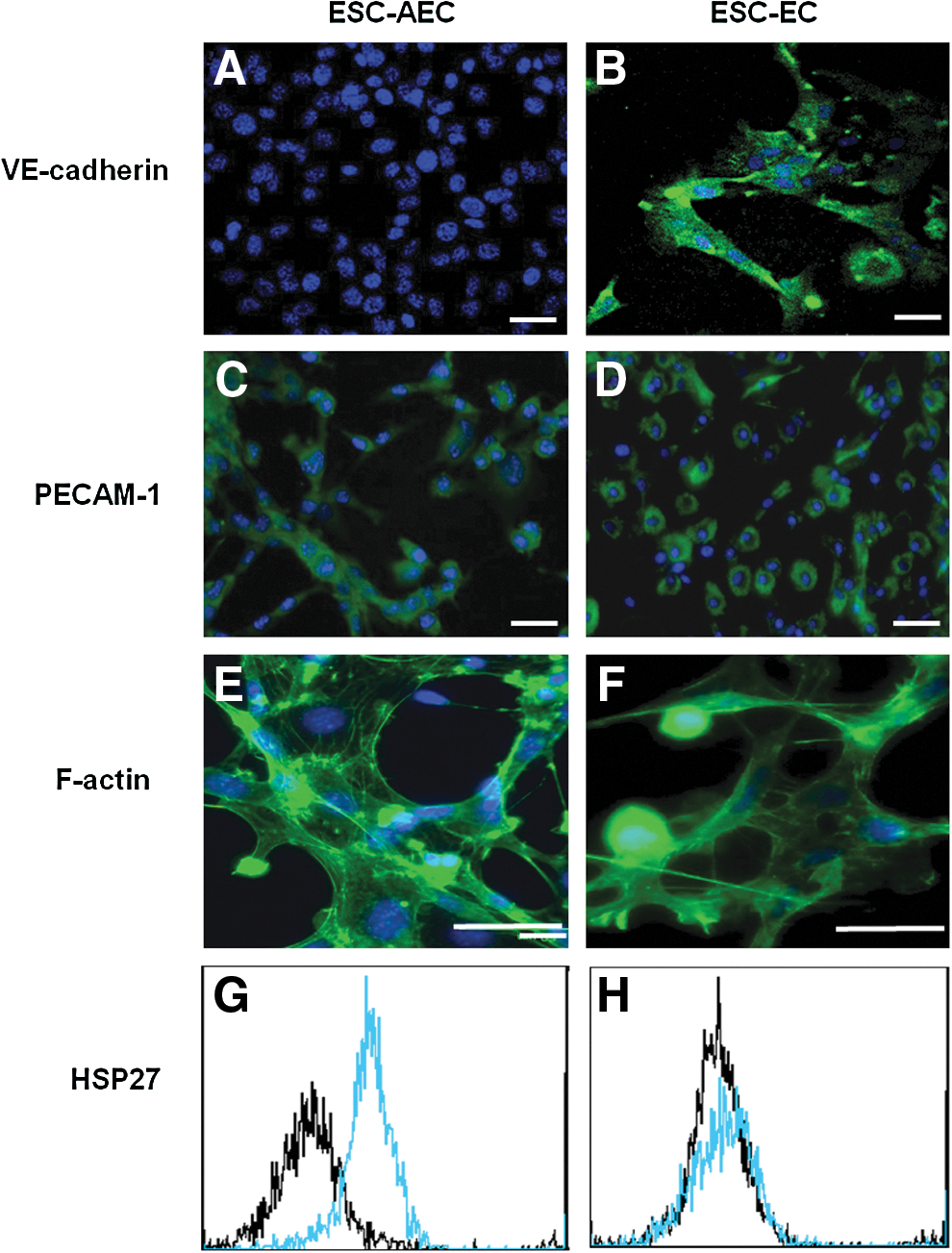
ESC-EC express VE-cadherin and ESC-AEC express increased levels of actin stress fibers and HSP27 phosphorylation. The presence of extracellular VE-cadherin adhesion molecule is not observed to be expressed in many of the
Tip/stalk and phalanx EC markers
Based on the hypothesis that the ESC-AEC contain tip and stalk EC, while the purified ESC-EC are composed of primarily phalanx EC, we examined some additional putative markers for tip, stalk, and phalanx EC. Additionally, both extracellular and total (intra- and extracellular) Notch1, tip cell marker CXCR4, and Tie-2 were examined (Fig. 5A–H). Both ESC-AEC and ESC-EC populations express Notch1, but the ESC-AEC contain greater numbers of Notch1+ cells (Fig. 5A, C) compared with ESC-EC (Fig. 5B, D). This difference is most pronounced in the extracellular Notch1 expression found on stalk cells, 92% of ESC-AEC (Fig. 5A) compared with only 56% in ESC-EC (Fig. 5B). The tip cell-specific molecule, CXCR4 [22], was expressed in 35% of the ESC-AEC (Fig. 5E), but not found in the ESC-EC (Fig. 5F). Both cell populations contained high numbers of Tie-2+ cells, but the ESC-EC displayed greater expression levels (based on higher levels of fluorescence intensities, or right shift, in the cell population; Fig. 5G) compared with the ESC-AEC (Fig. 5H).

ESC-AEC expressed increased numbers of stalk and tip cell markers compared with ESC-EC. The 2 cell populations were stained for both extracellular (stalk cells) and total (intra- and extracellular) Notch1, tip cell marker CXCR4 and Tie-2. Compared with isotype controls (left histogram),
In vivo angiogenesis
Lastly, the ability for these ESC-derived EC subpopulations to integrate with host vasculature was confirmed in an in vivo CAM assay. The results indicate that all EC populations, as well as Matrigel alone, were able to promote vascular development in vivo (Fig. 6B–E) and all cells integrated into host vasculature, indicated by brown staining of graft cells in Fig. 6G–J, compared with the vehicle control (Fig. 6F). The undifferentiated R1 ESC also exhibited integration into the chick vasculature (Fig. 6G), possibly by first differentiating into vascular cells in the proangiognenic environment of the CAM. Additionally, the quantified vascular activity, measured in average length of vessels, of all of the EC treatments (MCEC, ESC-AEC, and ESC-EC) increased significantly compared with controls (Fig. 6K). However, in the in vivo CAM assays, the EC populations performed similarly in their contributions to the native vasculature. Since this was an in vivo assay, we expect that the sophisticated microenvironment in the CAM compensates for the subtle differences between the graft cell populations.
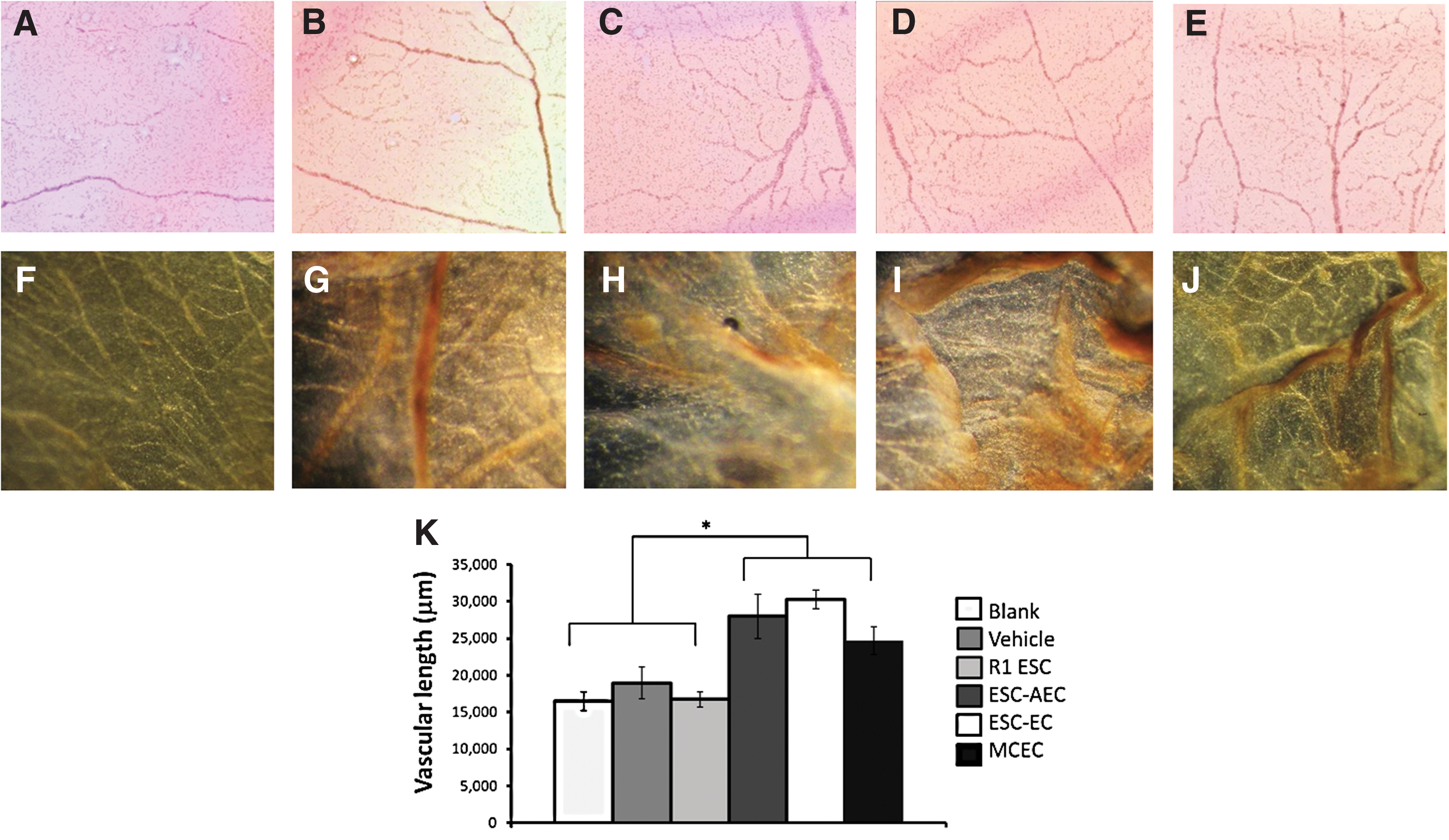
ESC-AEC and ESC-EC behave similarly in CAM assays. Images of CAM vasculature resulting from the
Discussion
The ability to generate EC subphenotypes in vitro from stem cells has not been well explored, especially identification and/or purification of tip/stalk EC compared with the more quiescent phalanx EC found in stable vessels. This work, for the first time, confirms the presence of these tip/stalk, and phalanx EC subpopulations derived in vitro from ESCs by generating 2 distinct EC populations with notable marker and functional differences. A summary highlighting these differences is provided in Fig. 7A. Note that the cobblestone-shaped ESC-EC contain high numbers of EC expressing Flt-1, Tie-1, Tie-2, and extracellular VE-cadherin, whereas the ESC-AEC contain CXCR4+ tip-like and increased Notch-1+ stalk-like cells exhibiting increased cytoskeletal organization and F-actin stress fibers [36 –38], and increased phosphorylation of HSP27, correlating with increased motility toward Ang-2. These cell populations also maintained distinct morphological properties in vitro. The ESC-EC continued to exhibit a cobblestone shape, while the ESC-AEC were slightly more elongated and generate loops on the culture dish that did not collapse at confluence. Lastly, the most robust functional difference between these distinct EC populations was seen in the increased sprouting activity and maintenance of vessel-like networks by the ESC-AEC compared with ESC-EC. These data indicate that the angiogenic ESC-AEC contain the all endothelial subphenotypes (including tip-, stalk-, and phalanx-like EC) typically represented in the developing vessel (Fig. 7B), whereas the purified ESC-EC contain >90% phalanx-like EC.

Summary of ESC-AEC and ESC-EC characterization.
Although Notch 1 was observed in both EC populations, the higher expression of total and extracellular Notch 1 in the ESC-AEC is consistent with the active Notch1 signaling observed in tip/stalk cells. This is consistent with the in vivo scenario in which the VEGF-induced expression of Notch1 ligand, Dll4, is upregulated in cells at the tip of the growing vascular sprout exposed to the highest levels of VEGF [21]. The Dll4 then binds to extracellular Notch 1 on neighboring stalk EC, resulting in the downregulation of VEGFR2 signaling and dampening the VEGF-induced expression of Dll4 in the stalk cells, and preventing these cells from leaving their position in the stalk [19].
Tip cells are also known to express lower levels of Flt-1 [39] compared with phalanx EC [28]. The low number of Flt-1+ cells in the ESC-AEC population is consistent with the tip-containing subpopulation, whereas the ESC-EC expressed higher levels of Flt-1, consistent with a phalanx-like phenotype [28]. This is congruous with the proposed function for Flt-1, serving as a VEGF repository (with greater binding affinity to VEGF) and mitigating EC migration in response to VEGF signaling [40], and allowing the generation of a more stable endothelium through upregulation of extracellular VE-cadherin to the cell–cell junctions [19].
Although the ESC-EC expressed greater numbers of Tie-1+ cells and expression levels of the Tie-2 molecule, the roles of Tie-1 and Tie-2 are distinct. While Tie-2 expression is usually constitutively expressed by EC, it seems that Tie-1 expression is more strongly regulated. It has been shown that Tie-1 mRNA is upregulated at sites of turbulent flow [41], and leads to the upregulation of the pro-inflammatory cell adhesion molecules (CAM): VCAM-1, E-selectin, and ICAM-1 [42]. Likewise, Tie-2 activation and upregulation via Ang-1-Tie-2 signaling contributes to the maintenance of the stable quiescent EC phenotype. In confluent (quiescent) EC, Ang-1 induces Tie-2 translocation to the EC-to-EC contacts to facilitate the formation of homotypic Tie-2 complexes [43], leading to inhibition of paracellular permeability [44,45]. While the mechanisms for Tie-1 and Tie-2 expression and activation are not equivalent, both would be expressed on stable phalanx EC that line vessel lumens for the regulation of pro-inflammatory adhesion molecules that facilitate leukocyte adhesion and transmigration [46,47], consistent with the ESC-EC subpopulation.
It is important to note that the cell culture medium used for generation of these subpopulations has not yet been explored. The presence of tip/stalk-like cells in the ESC-AEC is not surprising, as it is well known that high levels of VEGF signaling induce and help guide angiogenic tip cells in [27]. However, we expect that because FGF signaling, known to preserve the integrity of the endothelial adherens junction VE-cadherin/p120-catenin complex [48], may play a role in the specification and/or maintenance of the phalanx EC. A second candidate for phalanx specification is BMP-9, which is shown to inhibit angiogenesis and induce quiescence in vascular EC in vivo [49]. Ongoing studies will examine the signaling mechanisms for directing the fate of these EC subpopulations.
Conclusions
The ability to generate phenotypically distinct EC subpopulations in vitro from stem cells would enable the specialization of various EC-specific studies and therapeutic treatments. For example, tip cells are thought to play a dominant role in tumor angiogenesis, and therefore, these cells could provide an ideal in vitro model for screening anti-angiogenic molecules for cancer therapies. The tip/stalk cells would also be a more ideal cell type for use as cell therapies for ischemic vascular disease [50]. Conversely, because the phalanx EC are found in stable arteries [44,45] and probably sites of atherosclerosis [42], this EC subpopulation may be ideal for studying anti-atherosclerotic treatments. The phalanx EC would also be ideal for lining the lumens of stents or small-diameter vascular grafts [51,52].
This work confirms, for the first time, that tip/stalk- and phalanx-like EC populations can be found in cultures generated from ESCs, and that a growing blood vessel structure is not required for these cells to take on tip/stalk-like subphenotypes. Moreover, the phalanx-like EC can be isolated and cultured separately, and maintain their distinct phenotype after several passages despite being cultured in identical cell culture medium as the ESC-AEC. The generation/identification of these EC subpopulations authenticates corroborating studies examining the mechanisms leading to commitment of maintenance of these distinct EC populations.
Footnotes
Acknowledgments
This work was funded by a National Institutes of Health–National Service Award (NIH-NRSA) from the National Heart Lung and Blood Institute (NHLBI) no. F31HL087716 and a New Faculty Award from the California Institute of Regenerative Medicine (RN2-00921-1).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.