
Abstract
Research into angiogenesis has contributed to progress in the fast-moving field of regenerative medicine. Designing coculture systems is deemed a helpful method to understand the dynamic interaction of various cells involved in the angiogenesis process. We investigated the juxtacrine and paracrine interaction between 3 different cells, namely rat marrow-derived mesenchymal stem cells (rMSCs), rat muscle-derived satellite cells (rSCs), and rat neonatal cardiomyocytes (rCMs), and endothelial cells (ECs) during angiogenesis process. In vitro Matrigel angiogenesis assay was performed whereby ECs were monocultured or cocultured with rMSCs, rSCs, and rCMs or their conditioned media (CM). In addition, in vivo Matrigel plug assay for angiogenesis was conducted to assess the angiogenic potential of the rCM-, rMSC-, and rSC-derived CM. Our results demonstrated that the rMSCs, rSCs, and rCMs elongated along the EC tubules, whereas the rMSCs formed tube-like structures with sprouting tip cells, leading to improved angiogenesis in the coculture system. Moreover, the rMSC- and rSC-derived CM significantly improved angiogenesis tube formation on Matrigel, accelerated EC chemotaxis, and increased the arteriolar density, vascularization index, and vascularization flow index in the Matrigel plug in vivo. Western blotting showed that rMSCs secreted a high level of vascular endothelial growth factor, basic fibroblast growth factor, and stromal-derived factor-1-alpha. Tie2 is also shed from rMSCs. This study demonstrated that stem cells interact with ECs in the juxtacrine and paracrine manner during angiogenesis, and marrow MSCs have superior angiogenic properties.
Introduction
T
Designing coculture systems is believed to be a helpful method to understand the dynamic interaction of various cells involved in the angiogenesis process [1]. The angiogenic potential of a number of progenitor cells have recently been investigated by some authors, who have proposed multiple hypotheses, including physical contact and paracrine mechanisms, to explain the positive, and sometimes negative, effects of these cells in angiogenesis. In vitro morphogenesis of endothelial cells (ECs), as well as in vivo neovascularization, is induced by mesenchymal stem cell (MSC) secretome [6]. Conditioned media (CM) derived from MSCs cocultured with mouse brain endothelial cells (MBECs) significantly increased MBEC expression of angiopoietin 1 (Ang1)/Tie2 and Flk1 compared with MBECs alone [7]. Crosstalk between MSCs and astrocytes accelerated vascular endothelial growth factor (VEGF) and Ang1/Tie2 expression by astrocytes in the coculture condition in comparison to astrocyte culture alone [7]. Other progenitors such as muscle-resident stem cells, known as satellite cells (SCs), are capable of producing a number of angiogenic growth factors. SC transplantation in experimentally injured mouse medial collateral ligament was reported to have accelerated angiogenesis and increased capillary density [8]. Moreover, in the coculture of rat SCs and microvascular fragments, the index of angiogenesis is greater than that of single culture alone [9]. Given the importance of vascular disease in the heart and muscle and dynamic interaction between ECs and the somatic and stem cell niche in the angiogenesis process, we aimed in this study to comparatively investigate the potential angiogenic effect of 3 different cells, namely rat MSCs (rMSCs), rat SCs (rSCs), and neonatal rat cardiomyocytes (rCMs). To study the physical and mechanical roles of these cells on EC tubulogenesis, we designed coculture systems of human umbilical vein endothelial cells (HUVECs) with rCMs, rMSCs, and rSCs on Matrigel. In addition, we assayed the angiogenic potential of rCM-, rMSC-, and rSC-derived CM in vitro and in vivo.
Materials and Methods
Animals
In this study, both green fluorescent protein (GFP)-positive (rat GFP+/+ ) ([Wistar-TgN (CAG-GFP) 184ys] strain) and GFP-negative adult male Wistar rats (rat GFP−/− ) were employed [10]. The rats, weighing 70–100g, were treated in accordance with the published guideline of The Care and Use of Laboratory Animals (NIH Publication No. 85-23, revised 1996). All implementation phases of this study were approved by the Animal Care Committee of the University of Tehran.
Cells isolation and expansion
rMSCs, rSCs, and rCMs were isolated as previously described. A complete description of the cell isolation techniques is provided in the Supplementary Methods section (Supplementary Data are available online at
CM preparation
The impact of CMs on EC tubulogenesis and migration was tested by collecting rMSC, rSC, and rCM CMs. The third passage of rMSCs, rSCs, and rCMs (with 70%–80% confluence) were cultured in Dulbecco's modified Eagle's medium-low glucose (DMEM/LG) for 5 days, and then the medium was subsequently discarded and washed 3 times with phosphate-buffered saline (PBS). Thereafter, 10 mL of serum-free DMEM/LG was added and incubated for further 24 h [6,11,12]. The CMs were then harvested, centrifuged at 1,000 g for 10 min at 4°C (Hettich universal 320R), and filtered through a 0.20-μm-pore syringe filter (GVS Filter Technology, Inc.). The harvest was stored at −80°C until use [6] (Supplementary Fig. S1).
Western blotting
The concentrated CMs were subject to protein expression analysis through western blotting. Detailed information is provided in the Supplementary Methods section.
EC migration assay
For migration assay, 24-well polycarbonate membrane cell culture plate inserts with 8-μm-pore size were obtained (Catalog No. 35224; SPL Life Science). For this propose, the ECs from the third subconfluent passage were utilized. In brief, ECs were serum-starved overnight [13] and were thereafter trypsinized. Next, 5×104 cells were suspended in 100 μL serum-free DMEM/LG and seeded on the top chamber of the transwell Inserts (triplicate per one sample). The bottom chambers contained 750 μL of serum-free CMs, each from the rMSCs, rCMs, and rSCs, or a combined serum-free CM (a cocktail containing 250 μL CM for each cell type). The serum-free DMEM was used as control [14]. After incubation for 24 h in a CO2 incubator, 6 random fields per well were counted. All experiments were performed in triplicate.
EC labeling with CM-Dil
The ECs were labeled with CellTracker™ CM-Dil (Catalog No. C-7000; Molecular Probes) according to the manufacturer's instructions. The cells were incubated with 2 μM cell tracker for 20 min at 37°C, washed with PBS, trypsinized, washed 3 times with PBS, and then resuspended in DMEM/LG [15].
In vitro tube formation assay (Matrigel assay)
In vitro tube formation was assessed via growth factor-depleted Matrigel (Catalog No. 356231; BD Biosciences). For Matrigel assay, a 3D culture system was designed in 96 well plates (Catalog No. 167008; Nunclon), in which the wells were coated with 50 μL of overnight pre-cooled Matrigel followed by incubating at 37°C for 30 min to be solidified [16]. Three different protocols were used to perform the in vitro angiogenesis assays: Protocol I: single-cell culture (ECs, rCMs, rSCs, and rMSCs); protocol II: coculture system (EC-rCMs, EC-rSCs, and EC-rMSCs) and combined cell culture (ECs–rMSCs–rSCs–rCMs); and protocol III: CM-dependent culture (ECs—200 μL rCM CM, ECs—200 μL rSC CM, ECs—200 μL rMSC CM, and EC-cocktail CM containing ∼66.7 μL CM for each cell) (Supplementary Fig. S1). Three independent experiments with duplicate wells were performed: a total of 2×104 cells were seeded in each well. For coculture experiments, a 4:1 ratio of cells was used (16,000 ECs:4,000 non-ECs). The wells were checked daily by an inverted microscope (Nikon; Eclipse TS100), which was equipped with a digital imaging device (Sight DS-L2; Nikon). In our preliminary studies on in vitro Matrigel assay, we investigated different time points for tubulogenesis and found that the best time for tubulogenesis assay was at day 7. At day 7, each well was photographed and analyzed [15]. In vitro angiogenesis was assessed quantitatively through examining 25 colonies in each well, and an assigning score from 0 to 4 to each colony: 0, aggregate with no budding; 1, colony budding without arborization; 2, with arborization; 3, with anastomosis; and 4, development of a complex network (Supplementary Fig. S2). The total score per well was calculated by adding all the 25 scores, with a maximum possible score of 100. The average tubular length of 50 tubes in 10 serial microscopic fields and the average of tube area per total field area in 10 serial microscopic fields were also measured. Tubular length was measured in micrometer using an image-analyzing software (Sight DS-L2; Nikon), while the tube area was calculated as pixel2 using AxioVision Version Rel 4.8 Software (Carl Zeiss Microimaging, Inc.). Fluorescent images were captured with an inverted microscope (Olympus IX71), and merged using analysis LS Starter software.
Matrigel implantation in rats
In this phase, 500 μL of CM, each from rCM, rMSCs, and rSCs, or from the control DMEM (DMEM/LG that was incubated in 37°C in a CO2 incubator), was added to 500 μL of overnight precooled growth factor-depleted Matrigel and mixed by gentle vortexing for homogenization. The final volume for injection was 1,000 μL. The mixture was loaded in a syringe and maintained in 4°C until injection. The rats were separated into 4 experimental groups (n=4 each); in each group, the mixture of CM from each cell (or control DMEM) and Matrigel was injected subcutaneously. Before injection, rats were anesthetized with xylazine (5 mg/kg) and ketamine (80 mg/kg), and the left flank was shaved and disinfected by a surgical disinfectant solution.
Ultrasonography imaging
For ultrasonography imaging, all animals underwent an ultrasonographic analysis at days 0, 3, and 7 postinjection using a Voluson 730 ProSystem (GE Medical Systems) fitted with a 5–12-MHz linear 3D–4D transducer. The anesthetized rats were placed on the right side before the injection site was impregnated with acoustic gel for the positioning of the probe. Raw-power Doppler 3D data were selected and processed using 4D view version: 5.0 software (GE Medical Systems Kretztechnik GmbH & Co. OHG Company). The 3D power Doppler of the injected site (hypoechoic compared to the surrounding tissue) was measured and demonstrated by Vocal® analysis in the software, which was calculated as vascularization index (VI), flow index (FI), and vascularization-flow index (VFI) of the cut volume (in terms of cm3) by the following formulas. All calculations were done by the software (Fig. 1). Orthogonal and reconstructed 3D ultrasonography imaging of Matrigel mass that was injected subcutaneously. The swollen form of Matrigel beneath the skin during injection changed and flattened by day 3 or 7 (upper right part in this figure). After ultrasonography imaging of the injection site, the hypoechoic mass was selected, and 3D power Doppler information was measured by Vocal analysis. In the bottom right, the 3D reconstructed image of the gel is shown. This image is related to Matrigel impregnated with control Dulbecco's modified Eagle's medium (DMEM) at day 7. Red points that are seen in the gel are indicative of blood vessels. On the left side, the upper column is indicative of gray scale g (0–100), and the lower column represents color value c (0–100). Color images available online at 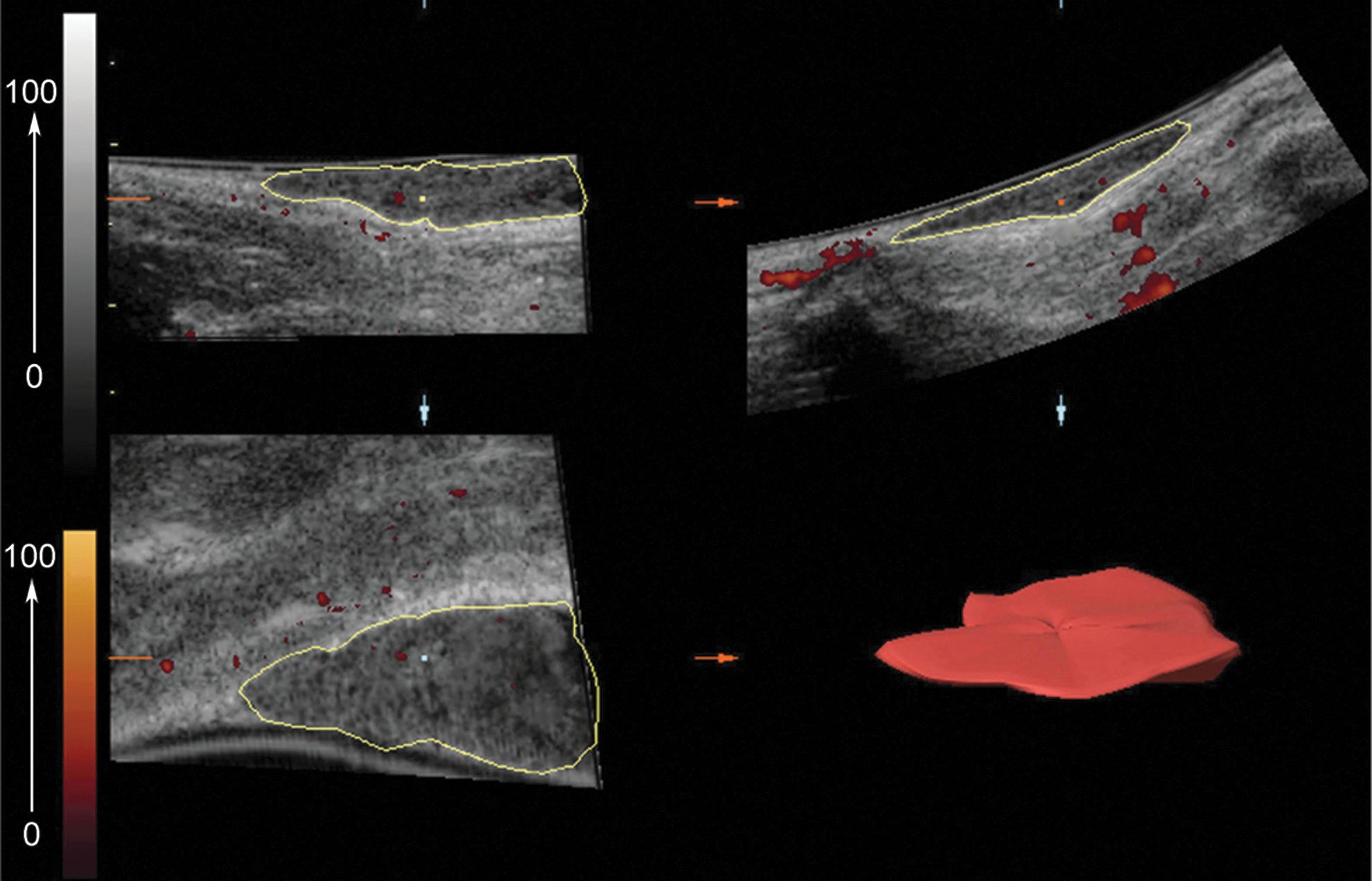
FI describes the average of color intensity, showing the blood flow in 3D volume indirectly, where c stands for the color value in the cut volume at the range interval of 0–100. VI calculates the number of blood vessels in the cut volume. In this formula, g is the abbreviation of gray scale value in the cut volume at the range of 0–100, and c stands for color value in the cut volume at the range interval of 0–100 again. Additionally, hg(g) presents the frequency of gray scale value in 3D volumetric image, and hc(c) scores the frequency of color value in 3D volumetric image. VFI is the multiplication of the blood vessel number by the blood flow.
Histopathology of Matrigel plug
Seven days postinjection, the rats were sacrificed, and the Matrigel plug was obtained. A fixative solution containing 2% paraformaldehyde and 1% glutaraldehyde (Catalog No. 1.04239.0250; Merck) was used for better fixation and preventing of Matrigel depolymerization. Paraffin-embedded sections were cut into 5-μm sections for immunohistochemistry and hematoxylin and eosin staining. To assess the effect of the CM in angiogenesis induction in the Matrigel plug, an anti-alpha-smooth muscle actin (α-SMA) antibody (Catalog No. M0851; Dako) was used to stain smooth muscle cells and myofibroblasts [17]. Capillary ECs were stained with an antibody against von Willebrand factor (vWF; Catalog No. M0616; Dako). After deparaffinization process, the slides were incubated in 3% H2O2 solution for 30 min to inhibit endogenous peroxidase. For antigen retrieval, the slides were submitted to autoclaving in a citrate buffer, pH 6.0, for 30 min. After cooling in room temperature and rinsing with PBS, the sections were incubated with the antibody for 30 min, washed twice with PBS, and colored with EnVision+ Dual Link System HRP kit (Dako) according to the manufacturer's instructions. 3,3′-Diaminobenzidine and Mayer's hematoxylin solution were utilized as the chromogen and counterstain, respectively. All parts of the slides were examined serially in a high-power field view.
Statistical analysis
Data on the angiogenesis scoring and on the Matrigel tube formation are expressed as mean±standard error; other data are expressed as mean±standard deviation and were analyzed using the SPSS package version 16 (SPSS, Inc.). In our study, the data analyses between groups were performed via the one-way analysis of variance (ANOVA) or Kruskal–Wallis test with a significant level of 95%. The results in any given group at different times were compared using the repeated-measure ANOVA test. Mean difference was significant at P<0.05. In histograms, statistical difference between the groups is shown by brackets with *P<0.05 and **P<0.005.
Results
Dynamics of angiogenesis in the coculture system of ECs with rMSCs, rSCs, and rCMs
Matrigel tube formation assay was employed to evaluate the effects of coculture on tube formation. First, the tube formation capability of each cell was assessed using monoculture systems, in which rMSCs GFP+/+ , rSCs GFP+/+ , and rCMs GFP+/+ were individually seeded onto Matrigel-coated wells: the rCMs, rMSCs, and rSCs formed cellular aggregates with no evidence of budding or elongation in Matrigel monoculture systems (Fig. 2). Aggregates were larger and less frequent for rCMs than those for the rMSCs or the rSCs (Fig. 2). For cocultures, the ECs were labeled with CellTracker™ CM Dil, and mixed with the rMSCs GFP+/+ , rSCs GFP+/+ , rCMs GFP+/+ , or their combination. Within 48 h, cells started to mix and elongate. After a week, the tubular-like structures and networks were sufficiently large, possessing lumens lined with ECs. Interestingly, co-alignment of the rMSCs, rSCs, and rCMs with the ECs was seen in the coculture systems (Fig. 3A–D). The rMSCs, rSCs, and rCMs elongated along the tubules, although aggregated cells were occasionally present. Moreover, compared to the EC monoculture, the formation of vascular-like networks was enhanced in the coculture conditions. The rMSC-EC coculture exhibited a significantly improved angiogenesis scoring (P=0.026), and increased tube length and tube area (P=0.001) compared to the EC monoculture (Fig. 4A–C). Specifically, in the rMSC-EC coculture, GFP-positive tip cells, sometimes with anastomosis, were discernible, indicating that the rMSCs contributed to sprouting angiogenesis (Fig. 3C). In the combined cell culture, dual-fluorescent cells, which appeared yellow, were detected (Fig. 3D); this could be due to vesicle trafficking between the adjacent ECs and GFP+ cells [1].

Monoculture systems. Phase-contrast and fluorescent micrographs of rSCs
GFP+/+
, rCMs
GFP+/+
, and rMSCs
GFP+/+
, which were separately cultured on Matrigel. As is seen, these cells formed cellular aggregates, without capability to form tube-like vascular structures alone. Scale bars: 100 μm. rSCs, rat muscle-derived satellite cells; rCMs, rat neonatal cardiomyocytes; rMSCs, rat marrow-derived mesenchymal stem cells. Color images available online at
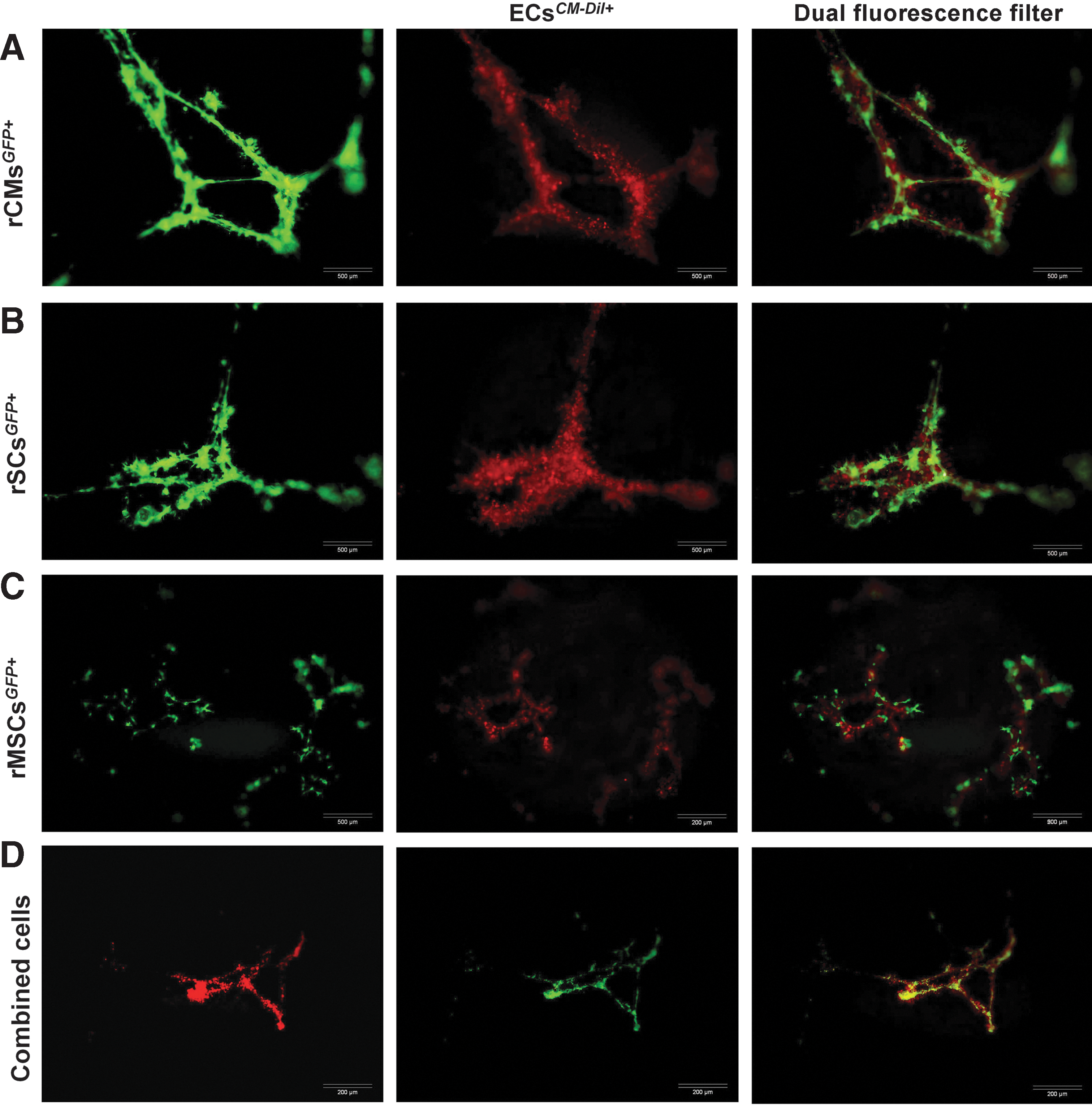
Representatives of tube formation by ECs
CM-Dil+
cocultured with rCMs
GFP+/+
, rSCs
GFP+/+
, rMSCs
GFP+/+
, or in combined cell culture
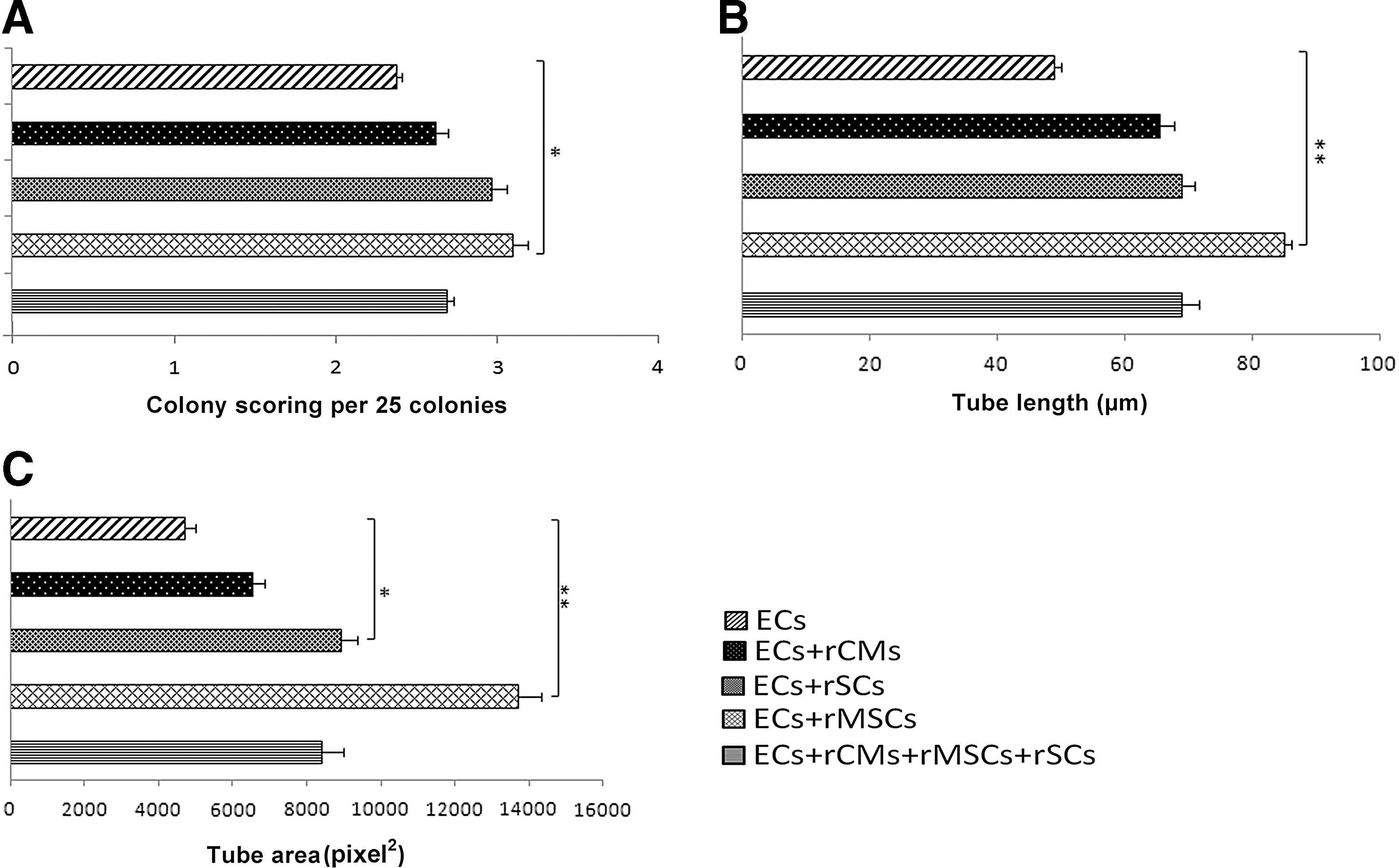
Improvement in Matrigel angiogenesis in cocultures compared to EC monoculture. Coculture of rMSCs with ECs significantly increased the angiogenesis scoring
Promotion of angiogenesis by cell-derived CM
The paracrine interaction between the rMSCs, rSCs and rCMs with ECs was evaluated by assessing endothelial tube formation on Matrigel, where the ECs were incubated with the rMSCs CM, rSCs CM, rCMs CM, and cocktail CM for 7 days. Statistical analysis revealed no significant effect of the rCM-derived CM on the ECs angiogenesis scoring, whereas the rMSCs, rSCs and the cocktail CM significantly increased angiogenesis scoring, tube length and tube area compared to the ECs incubated with DMEM alone (Fig. 5A–C). The rMSC-derived CM showed a higher colony scoring index than the rSC-derived CM (P=0.001). In addition, tube length increased significantly by the rMSC-derived CM when compared with the rCMs derived CM, cocktail CM or ECs incubated with DMEM alone (P=0.003, P=0.018, and P=0.000 respectively). The area of tube formation by the ECs incubated with the rMSC- and rSC-derived CM was also larger than that by the ECs incubated with the rCM-derived CM and ECs incubated with the DMEM alone. Taken together, the paracrine stimulation of the rMSCs derived-CM on the ECs tube formation was more potent than that of the other cell-derived CM used in this study. The rCM-derived CM had no better effect than the ECs incubated with the DMEM alone. In the mixed cocktail CM, the angiogenesis scoring and tube formation were less than those in the rSC- and rMSC-derived CM probably due to the dilution of soluble angiogenic factors after mixing the CM from the 3 different cells.
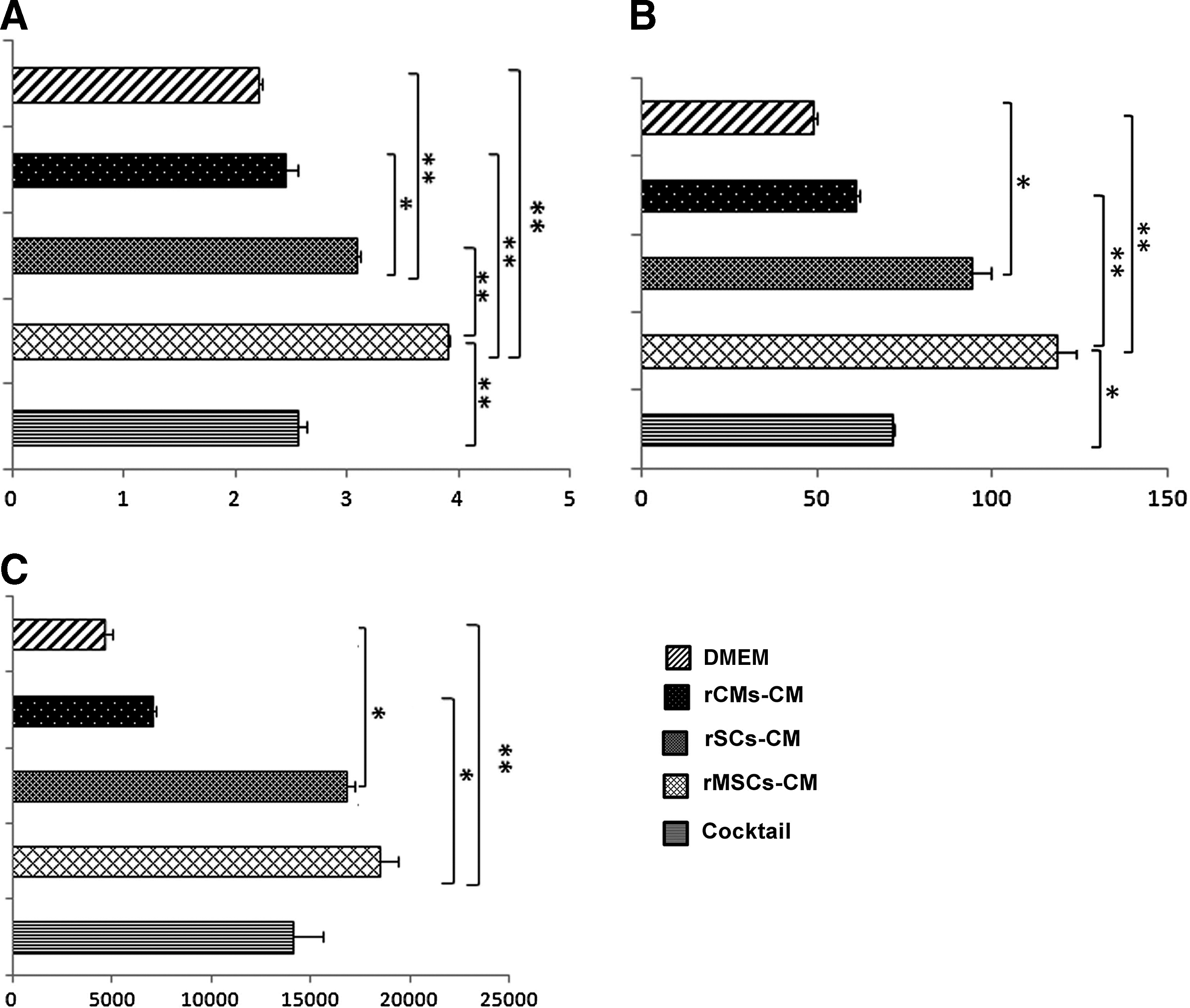
Angiogenesis scoring
Chemotactic potency of CM toward ECs
Next, the effect of cell-derived CM on ECs cell migration was examined. The results showed that the rMSC-derived CM profoundly stimulated the ECs chemotaxis compared to the rCM-derived CM and DMEM, indicating that the rMSC-derived CM has the highest level of chemotactic soluble factors, which accelerate the ECs migration and invasion through the membrane (Fig. 6). Also, the rSC-derived CM as well as the cocktail CM promoted the ECs migration, which was significantly higher than that of control DMEM (P=0.013 and P=0.035 respectively). In contrast, the rCM-derived CM had no chemotactic effect on the ECs (Fig. 6).

Effect of cell-derived CM on EC migration. The rMSC-derived CM significantly enhanced EC migration. Data are mean±standard deviation of the number of migrated cells per high-power field (HPF).*P<0.05, **P<0.005.
In vivo angiogenesis induced by CM in Matrigel plug
To further confirm the angiogenic potency of the cell-derived CM used in this study, in vivo angiogenesis assay was performed by mixing an equal amount of Matrigel with each CM and injecting the mixture subcutaneously into rats (Supplementary Fig. S1). On the third post-injecting day, some evidence of blood flow was observed at the peripheral aspect of the Matrigel plug (Fig. 7A–D; Supplementary Movies M1–M5 available in Supplementary Methods). Cell-derived CM had a positive effect on the VI and VFI indices over time. The repeated measures ANOVA showed no significant difference between the groups at days 0 and 3. Data analysis revealed that VFI and VI were not statistically different from day 0 to 7 in the control group (P=0.06), whereas the rMSC-derived CM substantially increased the VI index (P=0.006) as well as the VFI index (P=0.013) compared to the control group. In the rMSC-derived CM group, both VFI and VI indices were significantly different between day 0 and 7 (P=0.013 for VFI0–7 , and P=0.01 for VI0–7 ). Also, in the rats that were injected with Matrigel impregnated with the rSC-derived CM, both indices significantly increased from day 0 to 7 [VFI0–7 (P=0.001), VI0–7 (P=0.000)]. In addition, in this group, the indices were substantially higher than those of the control group at day 7 (P=0.001 for VI, and P=0.004 for VFI). The rCMs secretome also improved the indices by day 7 [VFI0 –7 (P=0.012), VI0–7 (P=0.011)], although the indices were not statistically different when compared with those of the control group (Fig. 7E, F).
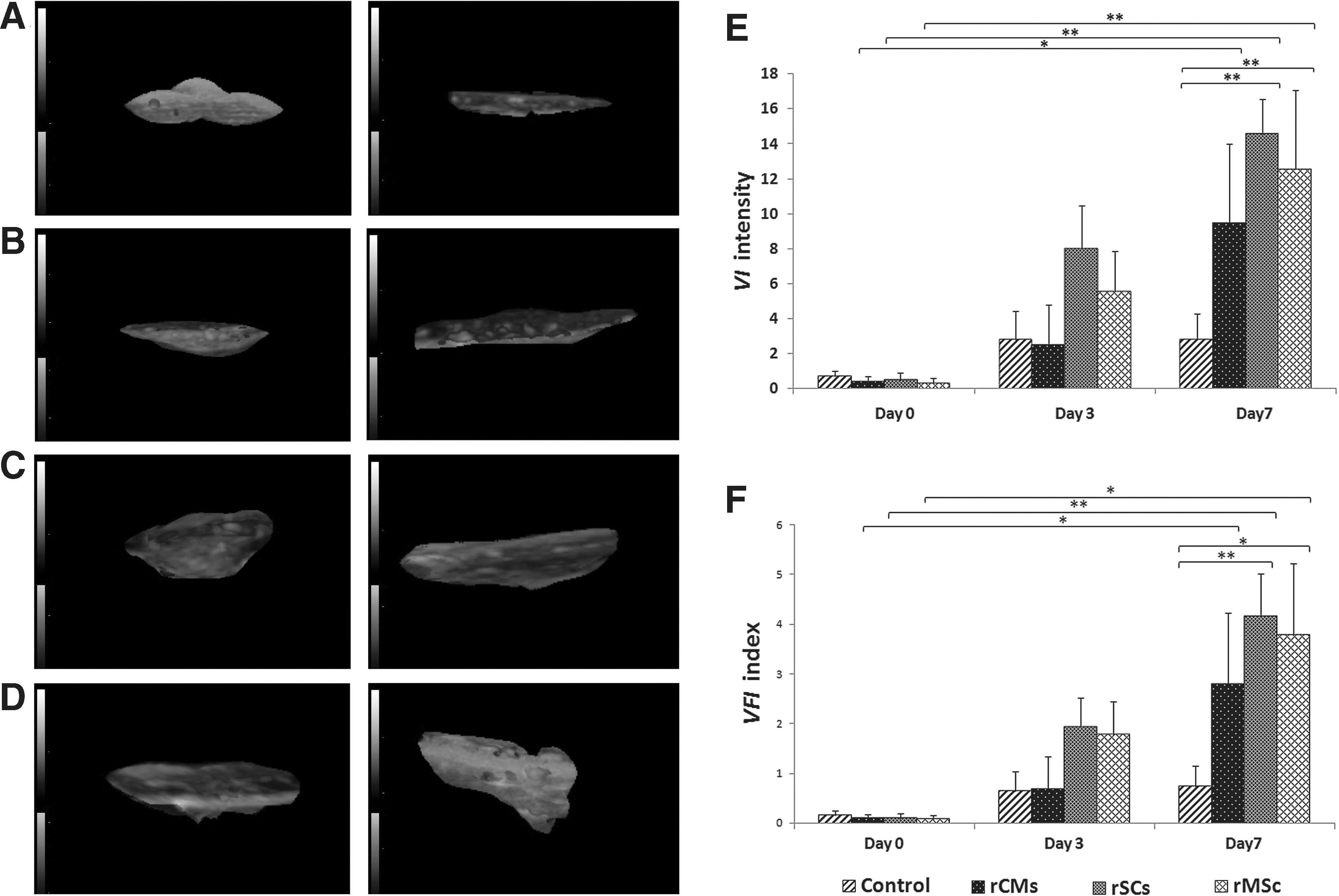
Representatives of the 3D views of Matrigel plugs by Vocal® analysis in the 4 groups at day 3 (left column) and day 7 (right column). [
Seven days post injection, the rats were sacrificed and the Matrigel plugs were removed. On histological review, a robust network of new, functional blood vessels with intraluminal erythrocytes had formed in the groups that had received Matrigel with the rMSC-derived CM and with the rSC-derived CM (Fig. 8A–H). A diffuse cellular infiltration, including a heterogeneous population of macrophages, lymphocytes and plasma cells, was seen in all plugs (Fig. 8A–H), which is consistent with a previous study that used Matrigel in rats [18]. Statistical analysis proved the rMSCs CM to be the most potent to increase capillary and arteriolar density, but this CM had less effect on myofibroblast recruitment into the Matrigel plugs (Fig. 8I–K). While the rCM-derived CM had a comparable effect on the in vivo angiogenesis process as did the control DMEM, the rSC-derived CM increased the numbers of vWF-positive capillary ECs, α-SMA-positive arterioles and myofibroblasts (Fig. 8I–K).
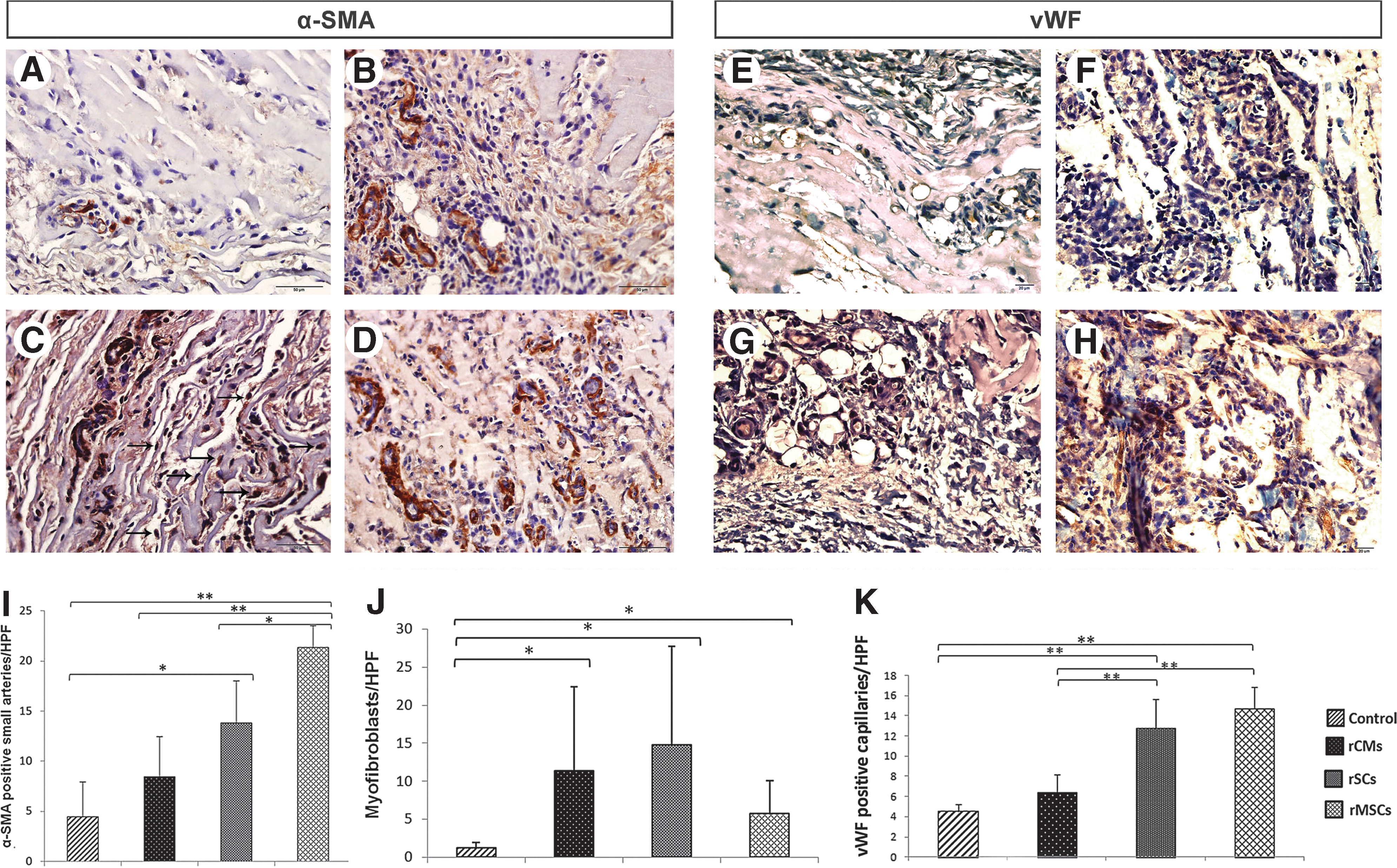
The effect of cell-derived CM on the development of α-SMA-positive arterioles, vWF-positive capillary ECs, and recruitment of myofibroblasts in the Matrigel plugs
Secretion of pro-angiogenic factors in cell-derived CM
EC tube formation is under the control of angiogenic soluble factors as well as direct cell–cell contact [1]. We examined the secretion of angiogenic soluble factors in the rMSC-, rSC- and rCM-derived CM via western blotting analysis. Western blotting confirmed the paracrine role of these cells in angiogenesis dynamics. VEGF was detected to be secreted into all of the 3 CM, with a strong immunoreactive band in the rMSCs CM (Fig. 9A). However, the expression of VEGF in the rCM-derived CM was barely detectable. Basic fibroblast growth factor (bFGF) was also found in both the rMSC- and rSCs derived CM (Fig. 9B). Tie2 was only detected in the harvested rMSC-derived CM (Fig. 9C). Soluble fragment of Tie2 (sTie2), which has been reported in human, is produced from proteolytic cleavage and shedding of Tie2, which was demonstrated to be induced by VEGF [19]. Here, we found that Tie2 is also shed from rat MSCs. In addition, stromal derived factor-1 alpha (SDF-1α) was detected in the rMSC-derived CM (Fig. 9D). None of the CM analyzed showed immunoreactive bands for Ang1 or angiopoietin 2 (Ang2).

Western blot analysis for angiogenic paracrine factors in cell-derived CM. VEGF was found in the CM from the rMSCs, rSCs, and rCMs with different levels. The highest VEGF level was detected in the rMSC-derived CM and the lowest level in the rCM-derived CM
Discussion
Stem cells are heralded as a source of great promise for augmenting angiogenesis during ischemia. Nonetheless, there is a controversy over their role in the angiogenesis process. This study was conducted to assess the juxtacrine interaction between rMSCs, rSCs, and rCMs with ECs undergoing tube formation angiogenesis, and to assess the paracrine effects of these cells on in vitro and in vivo angiogenesis. While monocultured rMSCs, rSCs, and rCMs were not qualified for tube formation angiogenesis on Matrigel, their cocultures with the ECs caused these cells to be aligned along the ECs during angiogenesis dynamics. The best effect was from the rMSCs: not only significantly promoted tube formation by the ECs, but also organized to form tube-like spatial structures with sprouting tip cells. It has been suggested that EC crosstalk with surrounding stem cells and cell transdifferentiation are involved in the regulation of new blood vessel formation [20]. Specifically, MSCs play an active role in cellular processes involved in the formation and stabilization of tubular arrays by ECs [15,21]. In addition, MSCs have the potential to exhibit an endothelial phenotype under specific conditions with a capability for transdifferentiation into endothelial-like cells [22,23]. In this study, we showed through the coculture system that interaction of rMSCs with ECs is more potent than that of rSCs or rCMs, which indicates the importance of these cells for angiogenesis stem cell therapy. In contrast with the effects of MSCs observed in this study, Menge et al. recently reported that human MSCs exert stabilizing effects on ECs and inhibit endothelial angiogenesis via cell–cell contact [24]. The converse effects of MSCs on ECs in their study compared with our results can partially be due to the xenogeneic coculturing system that we employed in our study. Indeed, allogeneic and xenogeneic cells may employ different mechanisms when encountered with similar biological phenomena [25]. Dual-fluorescent cells seen in the combined cell culture further affirmed the cell-to-cell communication between adjacent, cocultured cells, which may partially be mediated by microvesicles that are able to transfer a number of proteins, RNAs, or microRNAs between adjacent cells [26 –28]. The juxtacrine interaction between rMSCs, rSCs, and rCMs with HUVECs might also be mediated via gap-junctions that were previously demonstrated to be formed between adjacent cells in cocultures [29 –32]; be that as it may, in a recent study where MSCs were cocultured with nucleus pulposus cells, the authors found that the bidirectional intercellular transfer of membrane-bound microvesicles is the predominant mechanism of interaction between cells, and that other phenomena such as gap-junctional communication were detectable only in a small proportion of cells [33].
We also found that the MSC-derived CM had a greater angiogenic potential than the CM derived from the other cells. Chiming in with our findings, previous studies demonstrated that MSC-derived CM induced tube formation and sprouting of ECs, and could affect EC survival in vitro [15,34,35]. Specific factors such as vascular endothelial growth factor A (VEGF-A), hepatocyte growth factor (HGF), insulin-like growth factor 1 (IGF-1), and angiopoietins were reported to be upregulated in serum-deprived MSCs [22]. Here, we found a significant level of VEGF, bFGF, and SDF-1α secreted into the CM of the rMSCs; nonetheless, immune blotting of the CM showed neither Ang1 nor Ang2. Shedding of Tie2 into the rMSC-derived CM might be induced by a high level of VEGF in the CM. Inhibiting the proteolytic shedding of Tie2 through abrogation of VEGF signaling in uterine microvascular ECs has been suggested as an underlying cause of the failure of vascular remodeling in human pre-eclamptic pregnancies [36].
In the present study, the rMSC-derived CM increased the arteriolar density in Matrigel plug in vivo. Using ultrasonography analysis, we demonstrated that the new blood vessels were adequately functional to increase both the VI and VFI indices. VEGF as well as SDF-1α in the CM might also attract the endothelial progenitor cells (EPCs) circulating in peripheral blood. EPCs express VEGF receptors; hence, local VEGF plays a part in EPCs recruitment and subsequent formation of functional vessels [37]. Moreover, the combination of SDF-1α and VEGF can enhance EPC-mediated angiogenesis [38]. Despite the superior effects of rMSC-derived CM for angiogenesis induction in vivo, the population of myofibroblasts was inferior in the Matrigel plugs impregnated by rMSC-derived CM compared to the Matrigel plugs containing rSC- and rCM-derived CM. It seems that there are specific soluble factors that modulate myofibroblasts invasion. A previous study showed an inhibitory wound-healing effect of MSC-derived CM on corneal stromal fibroblasts in vitro [39]. Moreover, depending on concentration and release kinetics, MSCs exert inhibitory effects on inflammation, pathological remodeling, or scar formation [40].
An active, paracrine role of rSCs on in vitro and in vivo angiogenesis was also documented in this study. The stimulatory effect of the stem cell-derived CM on endothelial tube-like formation in vitro and arteriolar development in vivo confirmed the importance of paracrine signaling. It has been reported that a number of angiogenic inducers, including FGF, VEGF, IGF, and HGF, are produced by SCs [8]. Also, selected members of metalloproteases and extracellular matrix-degrading enzymes are secreted by human muscle SCs, which may participate in remodeling and in myogenesis [41]. In contrast to the above stem cells, the rCMs had no significant paracrine or juxtacrine effects during angiogenic dynamics.
In conclusion, this study demonstrated the paracrine and juxtacrine interaction between adult stem cells and ECs in angiogenesis dynamics. We showed that stem cells are actively and robustly involved in angiogenic processes. Future studies for understanding mechanisms mediating cell-to-cell communication between stem cells and ECs will provide new opportunities to interfere in the dynamics of angiogenesis for tissue engineering.
Footnotes
Acknowledgments
The authors wish to thank Mrs. L. Haghighi for her technical assistance and Dr. Mohseni, Mrs. M. Zakipour, and Mr. V. Siavashi for their help in this study. This study was supported by a grant from the University of Tehran, and a research grant (No. 89002374) from the Iran National Science Foundation (INSF).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.