
Abstract
Muscle-derived stem cells (MDSCs) are known to promote angiogenesis, but have never been studied in vascular diseases. We differentiated MDSCs into endothelial lineage cells in vitro by stimulation with shear stress and vascular endothelial growth factor. Such differentiated MDSCs (diff-MDSC) showed strong angiogenic potential in vitro. When tested in ischemic hindlimbs of mice, diff-MDSCs increased perfusion and decreased necrosis of the ischemic limbs, by promoting new vessel formation and by upregulating genes involved in endothelial expression. Such effects were not observed with native MDSCs (without endothelial stimulation in vitro). Diff-MDSCs were also injected into carotid arteries of rats after balloon denudation of the intima layer to induce intimal hyperplasia. The cell-treated group had significantly reduced intima-to-media thickness ratio compared to control, thus attenuating intimal hyperplasia by early re-endothelialization of the intima layer. Our findings suggest that MDSCs are a potential source of stem cell therapy for treatment of various vascular diseases, by inducing angiogenesis to improve perfusion in sites of ischemia, and by preventing intimal hyperplasia in sites of vessel injury.
Introduction
M
Intriguingly, it has been reported that MDSCs are not restricted to the myogenic or mesenchymal lineage. Many studies have shown that these cells exhibit angiogenic potential when differentiated into the endothelial lineage by stimulation with vascular endothelial growth factor (VEGF), and that this feature is crucial for the successful implantation of these cells into their respective target systems [5 –9]. MDSCs are known to express CD34 in vitro, a hematopoietic marker, but also a marker of endothelial expression, which implies that they can be differentiated into endothelial lineage cells [10,11]. However, there have been no studies up to date reporting the role of these cells in vascular diseases. Endothelial differentiation, and consequent angiogenesis, can theoretically be used to treat sites of ischemia. Such therapeutic angiogenesis has been proposed as an alternative to currently used treatment modalities for vascular diseases [12 –14]. However, it is not certain whether the angiogenic potential of these cells is strong enough to treat vascular diseases. Another important aspect of vascular diseases is intimal hyperplasia, especially after the introduction of endovascular therapies [15,16], and it is well known that early re-endothelialization of the diseased segment attenuates thickening of the intima [17,18].
In this study, we describe a method to differentiate isolated MDSCs into endothelial lineage cells, its characteristics, and its behavior in vitro. Diff-MDSCs were injected into ischemic hindlimbs of mice to investigate their angiogenic effect in vivo as well as their potential for use in the treatment of ischemic diseases. These cells were also injected into injured carotid arteries of rats to identify their role in the prevention of intimal hyperplasia.
Materials and Methods
Isolation and characterization of MDSCs
Under the approval of the Institutional Animal Care and Use Committee (IACUC) at our institution for the whole study, the gastrocnemius muscles from male Sprague-Dawley rats and Balb/c mice (3–6-week old) were removed and minced with a scalpel. After several times of washing with phosphate buffer saline (PBS; Gibco BRL), the tissues were treated with a serum-free medium containing 0.2% collagenase II (Sigma-Aldrich) at 37°C for 1 h and centrifuged at 3000 rpm for 5 min. The collected cells were then treated with PBS containing 3% dispase (Gibco BRL) for 30 min and 0.125% trypsin–ethylenediaminetetraacetic acid (EDTA)/PBS (Gibco BRL) for 10 min sequentially. After enzymatic dissociation, the cells were passed through a 70-μm stainer and cultured in a proliferation medium, consisting of a low-glucose Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum (FBS), 10% horse serum (Gibco BRL), and 1% penicillin–streptomycin (Sigma-Aldrich). The modified version of the preplate technique [2] was used to isolate MDSCs based on the differences in adhesion of the different populations to collagen-coated flasks. After 6 passages (pp6), the cells were plated in collagen-coated dishes and cultured for 1 week. The remaining cells represented our MDSCs, which were trypsinized with trypsin-EDTA to detach them from the dishes. These cells were sorted with MiniMACS (Miltenyi Biotec Inc.) to isolate cells with CD34 (+) features. Isolated MDSCs (1–5×104) were cultured in immunocytochemistry (ICC) plate slides (Nalge Nunc) and blocked with 5% blocking solution (Dako) at room temperature for 30 min. The primary antibodies for CD34 (1:50), Sca-1 (1:50), CD45 (1:50), and desmin (1:50 Abcam) were diluted and incubated with MDSCs at 4°C overnight. The cells were rinsed with PBS, incubated with their respective secondary antibodies (1:100) conjugated with fluorescein isothiocyanate (FITC) at 4°C for 3 h, and mounted with DAPI solution (Vector).
Shear stress and VEGF application
Isolated MDSCs were stimulated for endothelial differentiation by 2 different methods: shear stress application and VEGF supplementation. Shear stress was applied using an orbital shaker (SH30t; Finepcr), and the shear stress was calculated using the following formula:
Analysis of stimulated MDSCs
The effect of different stimulation methods of MDSCs was assessed by investigating the endothelial phenotypes from 3 different groups: MDSCs stimulated by shear stress (10 dyne/cm2) alone (SS group); MDSCs stimulated with VEGF (50 ng/mL) alone (VEGF group); and MDSCs stimulated with both shear stress and VEGF (SS-VEGF group); a last group consisted of MDSCs expanded without any stimulation (control group). Markers specific for endothelial phenotype were detected by immunocytochemistry: von Willebrand factor (vWF), VE-cadherin, Fetal liver kinase-1 (Flk-1), and CD31. This was done by incubating the cells with peroxidase-blocking solution (Dako) for 30 min at room temperature and adding the primary antibodies anti-vWF (1:100; Dako), VE-cadherin (1:100; Abcam), Flk-1 (1:100; Abcam), and CD31 (1:100; Abcam) for overnight incubation at 4°C. FITC-conjugated secondary antibodies were added to the cells, and the fluorescent stains were viewed under microscopy.
Reverse transcription (RT)–polymerase chain reaction (PCR) was performed by extracting RNA using the RNeasy Mini Kit (Qiagen) according to the manufacturer's instructions. The forward and reverse primers for vWF, VE-cadherin, Flk-1, CD31, and GAPDH (control) were as follows: vWF forward 5′-GACTTTAGGACGCAGGAGGG-3′, reverse 5′-GCCGTCTTCAGTAACTGGCA-3′; VE-cadherin forward 5′-GAGGCCACAGACCCCACTAT-3′, reverse 5′-TGGAAGTGGTAGAAGCGTCG-3′; Flk-1 forward 5′-CGAAATCTAGGCCTCAGCAC-3′, reverse 5′-CTTTTTGTCCACGGTCACCT-3′; CD31 forward 5′-GGCGGTGGTGACAGTATCTT-3′, reverse 5′-GTCACTGACAGAGGCGATGA-3′; GAPDH forward 5′-TGCCACTCAGAAGACTGT-3′, reverse 5′-GCATGTCAGATCCACAATGG-3′. PCR amplification was performed with a PTC-200 PCR Thermo cycler (MJ Research). The quantity and quality of the extracted RNA were determined by absorbance at 260-nm and 280-nm wavelengths using the Smartspec Plus Spectrophotometer (Bio-rad). PCR products were subjected to electrophoresis on 2%–3% agarose gels containing ethidium bromide. Images were captured with a camera, and the relative band intensities were calculated after being normalized with the band intensity of GAPDH.
In vitro angiogenesis assay
The angiogenic potential of the 4 groups of MDSCs was evaluated by investigating their ability to form tubules using the Matrigel Tube Formation Assay (Becton Dickinson). Matrigel was spread onto 24-well chamber slides (Nalge Nunc) and cells (3×104) from each group were plated and incubated at 37°C for 24 h in an M119 culture medium containing 2% FBS (Gibco BRL). The areas and lengths of the circles formed by the cells, as well as branch points (points with at least 3 or more branches sprouting), were calculated with Image-Pro Plus 4.5 (Media Cybernetics, Inc.) from 5 random high-power (×100) microscope fields per well.
Hindlimb ischemia model with cell treatment
MDSCs differentiated to exhibit endothelial regeneration potential by stimulation with both shear stress (10 dyne/cm2) and VEGF (50 ng/mL) were tested in vivo. Such differentiated MDSCs (diff-MDSC group) were compared with native MDSCs without stimulation (MDSC group) by injection of cells into hindlimb ischemic mice. Thirty-two Balb/c mice were divided into 3 groups: the diff-MDSC group, the MDSC group, and the control (sham operation) group. A hindlimb ischemic model was created by meticulous dissection and surgical resection (about 1 cm in length) of the femoral artery. The respective cells (1×107 of diff-MDSCs and native MDSCs) were injected intraoperatively and after 2 days in 4 different locations of the left hindlimb. The same amount of saline was inserted in the control group. The rate of limb necrosis was compared between the 3 groups, and the perfusion was assessed at 2 and 7 days using a laser Doppler flowmeter (Perimed). The perfusion unit ratio was calculated by comparing the perfusion of the ischemic left hindlimb with the normal right hindlimb for each mouse.
Carotid artery injury model with cell treatment
A carotid injury model was made on Sprague-Dawley rats by meticulous exposure of the common, internal, and external carotid arteries, after which black silk ties were placed around the proximal and distal ends for vascular control, and a small arteriotomy was made at the external carotid artery. A 2 Fr. Fogarty catheter (Edwards Lifescience) was inserted through the incision site, and balloon-induced endothelial denudation was performed by passing the balloon catheter 3 times along a 1-cm length of the common carotid artery. The denuded site was injected with either 1×107 cells of diff-MDSCs or saline (control) inside the lumen for implantation to occur for 30 min before restoration of blood flow.
In this model, we used diff-MDSCs previously transfected with lentivirus (Microgen) tagged with green fluorescent protein (GFP). This was done by adding 1 mL of lentivirus into 6-well plates containing MDSCs (at 70%–90% confluence) and 4–8 μg of polybrene (Sigma). The whole mixture was gently mixed together and incubated overnight in a 37°C and 5% CO2 atmosphere. Twenty-four hours after transfection, the cells were moved to a new culture medium. The cells were injected into the denuded carotid arteries 48–72 h after transfection.
Immunohistochemical staining, RT-PCR analysis, and intima-to-media thickness ratio measurement
Muscles from the hindlimbs were harvested at 4 weeks and fixed with 4% formalin for 24 h. The tissues were sectioned into 7-μm thicknesses and incubated with the primary antibody CD31 (1:500; Abcam) overnight. Secondary antibodies were added using an immunodetection kit (K5007; Dako) at 4°C for 3 h and mounted using Histomount (National Diagnostics). The sections were also incubated with a BCIP/NBT-buffered substrate (Sigma-Aldrich) at 37°C for 30 min for determination of alkaline phosphatase activity. The CD31-positive vascular structures and capillary densities were counted from 4 random fields at 200×magnification with Image Pro Plus 4.5 (CyberMetics, Inc.), and the mean value was calculated.
RT-PCR was performed for the hindlimb ischemia model using the RNeasy Mini Kit, (Qiagen) mentioned before. The forward and reverse primers for Vegf, Egr-1, and HIF-1α were as follows: Vegf forward 5′-GCCCATGAAGTGGTGAAGTTC-3′, reverse 5′-ACTCCAGGGCTTCATCATTG-3′; Egr-1 forward 5′-CATGCGTAACTTCAGTCGTA-3′, reverse 5′-ATGTGGTATGGCTGATTATG-3′; HIF-1α forward 5′-TGCTCATCAGTTGCCACTTC-3′, reverse 5′-CCATCCAGGGCTTTCAGATA-3′.
The carotid arteries of the study animals were harvested at 1, 2, and 4 weeks and fixed with 4% formalin for 24 h. The tissues were then sectioned (7-μm thickness) and stained with hematoxylin and eosin. After mounting with Histomount (National Diagnostics), the degree of intimal hyperplasia was determined by measuring the intima-to-media thickness ratio (IMTR). This was done by using a Leica application suite (Leica Microsystems) at 100×magnification, and analyzed with Image Pro Plus 4.5 (CyberMetics Inc), by calculating the mean after measuring the thicknesses at the 12 O'clock direction and every 60 degrees counterclockwise.
Immunohistochemical staining with CD31 for the carotid injury model was done using the same methodology mentioned previously. Immunofluorescent staining was done by treatment of the harvested tissue in sucrose solution and fixation with tissue-TEK using an OCT compound (Sakura Finetek). The primary antibody CD31 (1:100; Abcam) was added and cultured overnight, and the respective secondary antibody conjugated with rhodamine (Abcam) was added and cultured for 3 h at 4°C. The tissues were mounted with a mounting medium conjugated with DAPI (Vector).
Statistical analysis
All data are presented by the mean and standard error of the mean. For statistical analysis involving 3 groups or more, the one-way analysis of variance (ANOVA) was performed with post hoc test using the Tukey method. For comparisons between 2 groups, the independent variables Student's t-test was used. SPSS release 18.0 (SPSS Inc.) was used for analysis, and differences were considered statistically significant when P<0.05.
Results
Characterization of MDSCs
MDSCs isolated using the preplate technique were morphologically round and smooth bordered compared to earlier preplates (EP). We expanded our population of MDSCs up to 30 population doublings (PD30) and found that the properties of MDSCs in terms of morphology, marker expression, and growth rate were maintained without showing features of cell senescence (data not shown).
Immunocytochemical analysis to verify the properties of these cells showed a high expression of Sca-1 (90%) and CD34 (70%) compared to EP cells (pp2). Expression of desmin was indeterminate (30%–40%), although it showed a higher expression than EP cells while CD45 was negative (Fig. 1A, B). RT-PCR also showed that the expression of CD34 and MyoD was increased in MDSCs compared to EP cells, while the expression of desmin and CD45 was indeterminate (Fig. 1C).
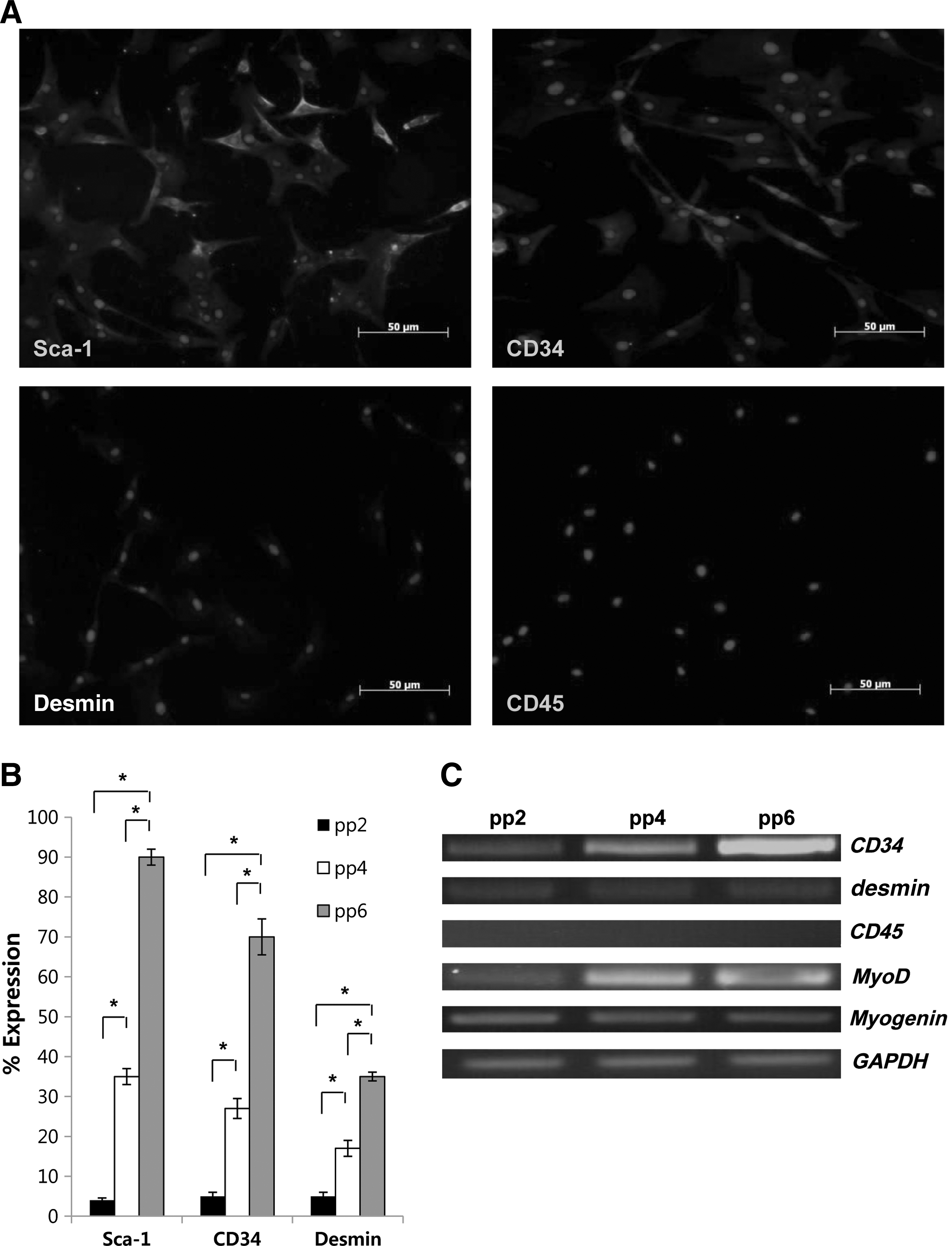
Characteristics of isolated MDSCs.
Endothelial differentiation and enhanced angiogenesis in vitro
Endothelial differentiation by application of shear stress and VEGF showed that the combined SS-VEGF group showed strongest expression of endothelial markers VE-cadherin, vWF, Flk-1, and CD31 on fluorescent immunocytochemical staining, as shown by the strong expression of FITC (green) compared to the SS and control groups (Fig. 2A). RT-PCR analysis confirmed such findings, with a higher intensity of Flk-1 and CD31 and a higher tendency (although not statistically significant) of VE-cadherin and vWF expression in the combined group compared to the other groups (Fig. 2C). Its endothelial function was demonstrated by assessing the ability to form capillary-like tubular structures in vitro using Matrigel (Fig. 3A). The length and area of the tubules, as well as the number of branch points, were quantified for the 4 different groups (Fig. 3B). The SS group showed similar tubule formation to control while the VEGF group showed increased formation of tubular structures. However, the SS-VEGF group showed a significantly higher degree of tubule formation compared to all other groups, demonstrating that this combination group had the highest potential for angiogenesis in vitro.

Characteristics of MDSCs after endothelial differentiation with shear stress and VEGF.
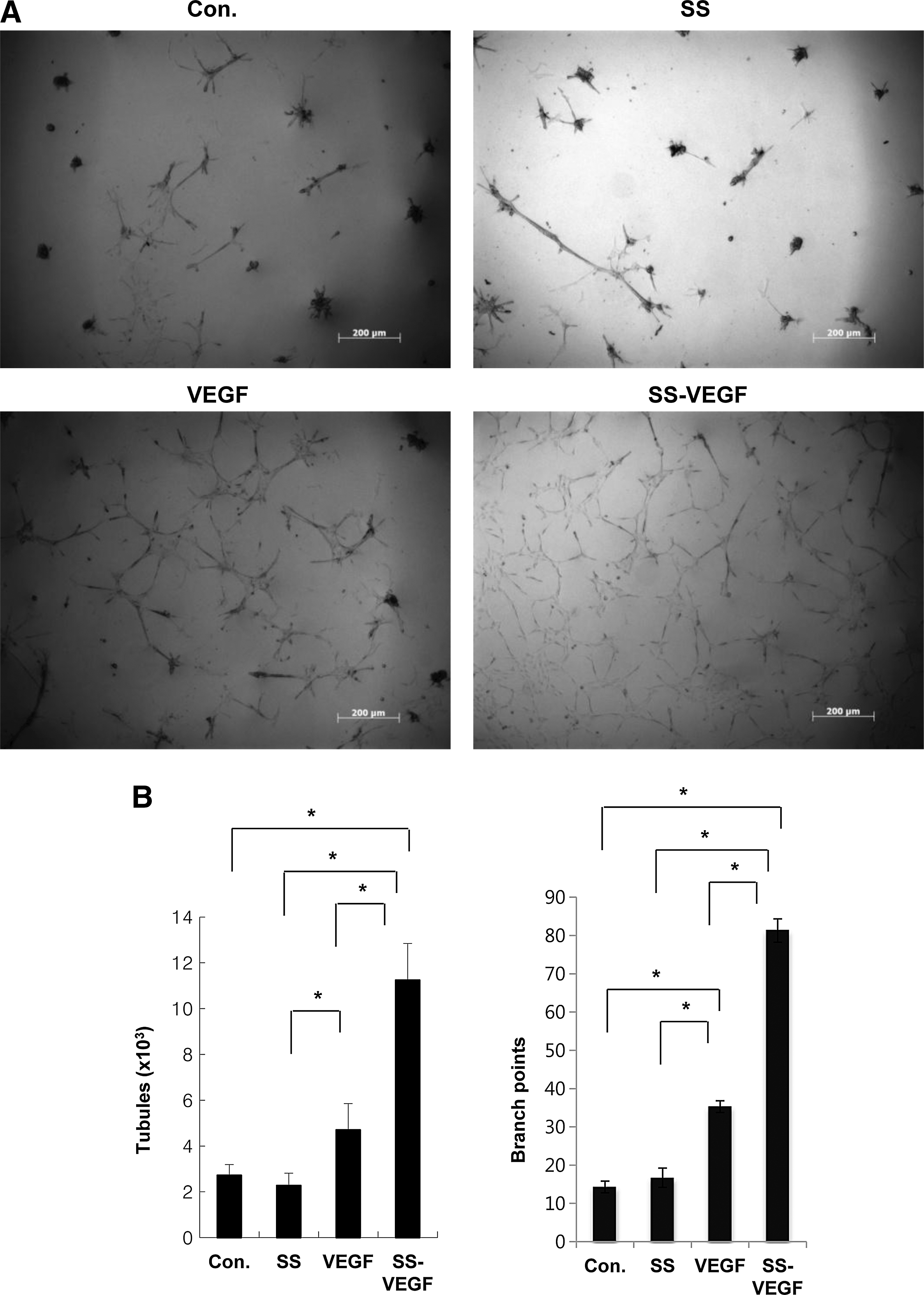
Angiogenic potential of diff-MDSCs in vitro.
Rescue effect of diff-MDSCs in hindlimb ischemia model
The angiogenic potential of diff-MDSCs (SS-VEGF) was tested in vivo using a murine ischemic hindlimb model. The perfusion unit ratio, measured as a ratio of the perfusion in the ischemic limb to that of the normal contralateral limb, was significantly higher in the diff-MDSC group compared to both native MDSC and saline-treated groups (Fig. 4A). This increased perfusion in the diff-MDSC group correlated with the overall necrosis rate, with a lower necrosis rate in the diff-MDSC group compared to the other groups (diff-MDSC 10% vs. MDSC 64%, control 73%) (Fig. 4B).

Angiogenic potential of diff-MDSCs in vivo.
Immunohistochemical staining with CD31 and alkaline phosphatase showed a higher degree of CD31-positive vascular structures in the diff-MDSC group (Fig. 5A). Native MDSCs also showed significantly higher CD31-positive vascular structures compared to control, but the diff-MDSC group showed significantly higher formation compared to both native MDSC and control groups. Alkaline phosphatase-stained capillary density also showed similar results, with diff-MDSCs showing a significantly higher capillary density compared to the other 2 groups (Fig. 5B).
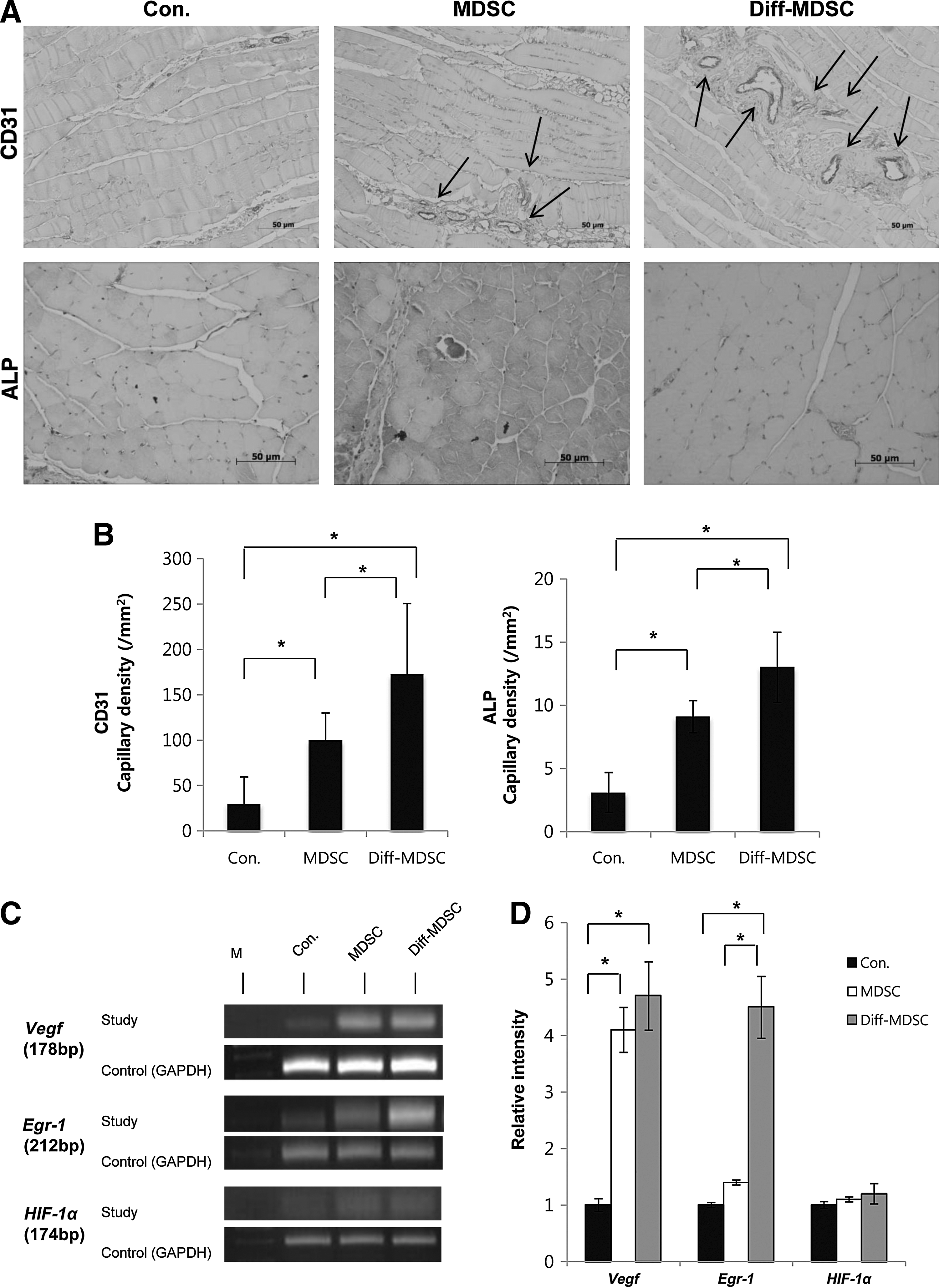
Immunohistochemical analysis and RT-PCR from hindlimbs of ischemic mice.
RT-PCR analysis for the master genes Vegf, Egr-1, and HIF-1α, known to play an important role in angiogenesis, showed an overall upregulation of Vegf and Egr-1 in diff-MDSCs compared to the other groups, but the difference in intensity was not observed with HIF-1α (Fig. 4D, E).
Attenuation of intimal hyperplasia with early re-endothelialization in carotid injury model
Carotid arteries stained with hematoxylin and eosin showed a well-preserved internal elastic lamina in all specimens. In one specimen harvested immediately after arterial injury, a well-denuded endothelium was observed (Fig. 6A). The control group showed a thicker intima compared to the diff-MDSC group (Fig. 6B), especially at 4 weeks, and quantification by measuring the IMTR demonstrated that the IMTR was statistically smaller in the diff-MDSC group compared to the control group at 2 and 4 weeks (Fig. 6C).
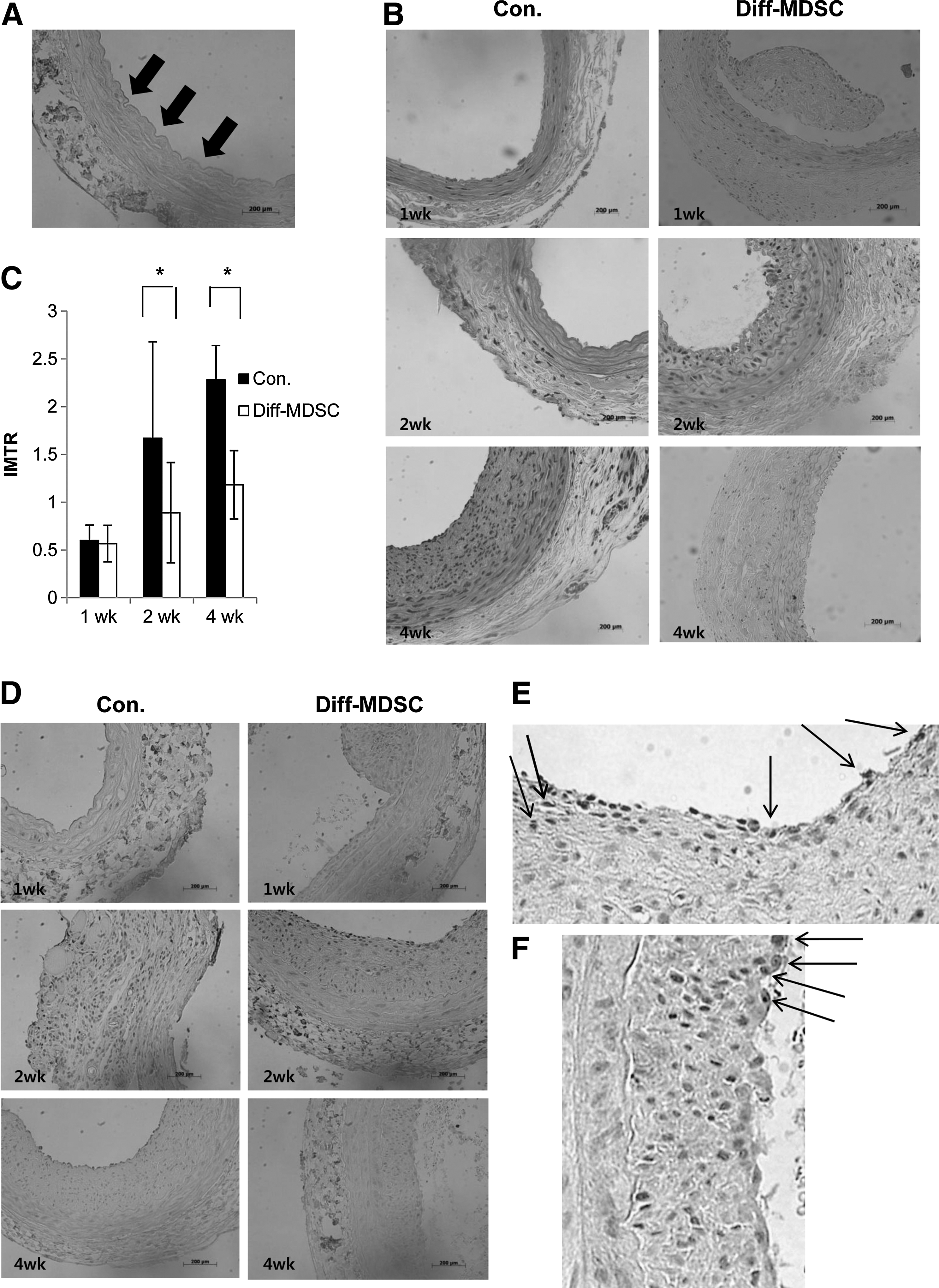
Attenuation of intimal hyperplasia by diff-MDSCs in vivo.
IHC staining with CD31 revealed that in the control group, there was no visible expression of CD31-positive endothelial cells at 1 week, around 30%–40% at 2 weeks, and 50%–70% at 4 weeks, while in the diff-MDSC group, around 50% of endothelial cell attachment was observed in the intima at 1 week, 80% at 2 weeks, and 90%–100% at 4 weeks (Fig. 5D–F). Therefore, re-endothelialization of the intimal layer was observed in both groups, but a higher amount of endothelialization was observed in the diff-MDSC group.
Diff-MDSCs transfected with lenti-hCMV-GFP were used to directly visualize the implantation of these cells. Characteristic green color of the GFP was found in the intima layer of the harvested tissues (Fig. 7A). When the specimens were costained with CD31, areas showing coexpression of GFP (green) and CD31(red) were visible, as shown by the merging of the 2 colors to show characteristic areas of higher yellow intensity, probably due to MDSC implantation (Fig. 7B).
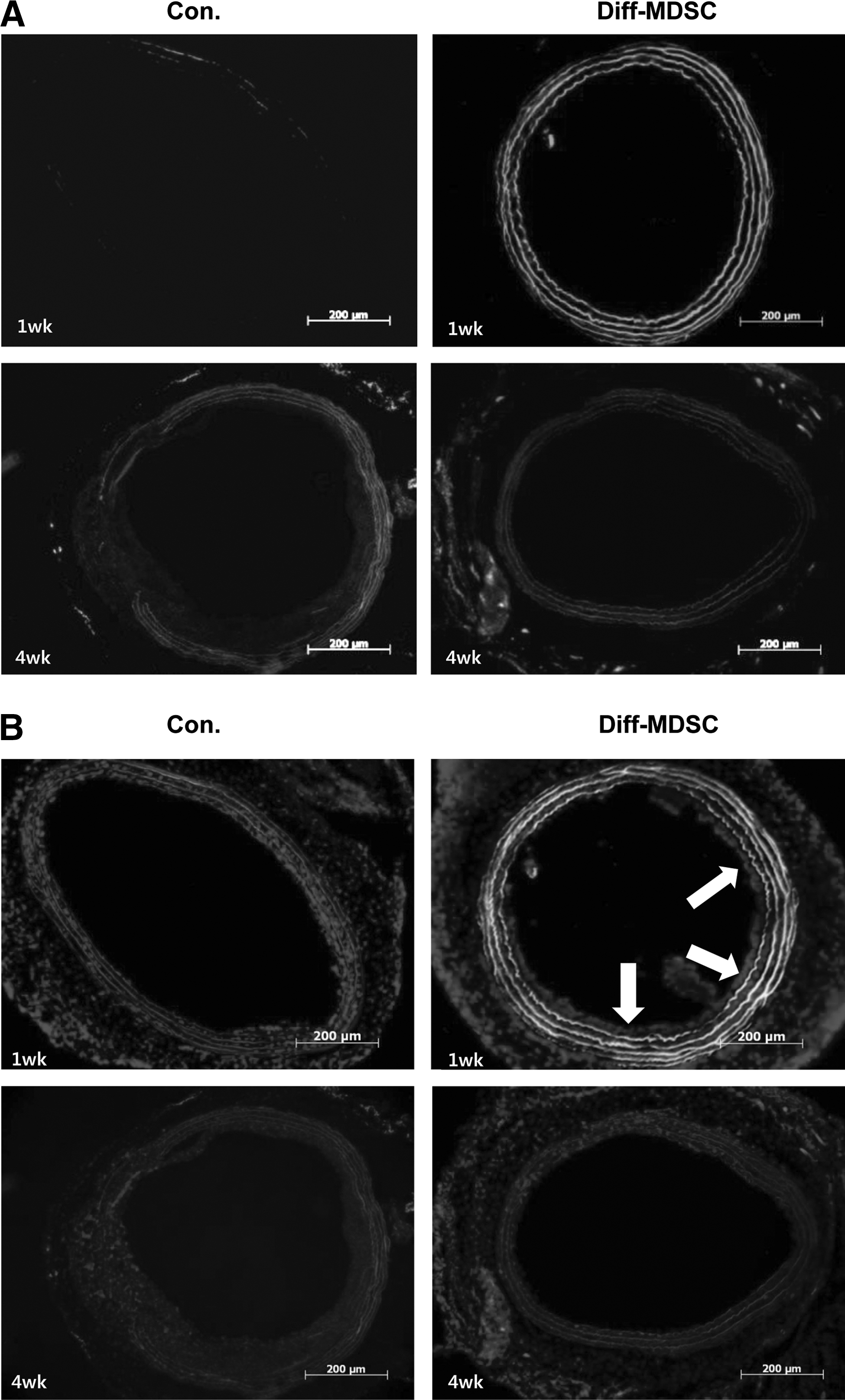
Lenti-hCMV-GFP-infected diff-MDSCs in carotid arteries.
Discussion
Our findings demonstrate that MDSCs can be differentiated in vitro to endothelial lineage cells using shear stress and VEGF, and that the combination of the 2 was most effective, as shown by the highest expression of endothelial markers Flk-1, CD31, vWF, and VE-cadherin. This was further supported by the stronger angiogenic potential of the cells in vitro, indicating that a higher degree of endothelial differentiation leads to a higher drive for angiogenesis. Shear stress has been recognized as an important modulator of endothelial phenotype, morphology, gene expression, and differentiation [20] and has been demonstrated to induce endothelial differentiation in different types of stem cells [21,22] and progenitor cells [23,24]. VEGF is a widely known factor to induce vasculogenesis and angiogenesis, and plays a fundamental role in the growth and differentiation of vascular endothelial cells [25,26]. In fact, there have been reports showing that in other types of cells, both shear stress and endothelial growth conditions are needed to induce endothelial differentiation, and stimulation with only one of these methods failed to show expression of endothelial markers [22,27].
Our in vivo studies with ischemic hindlimb mice showed that diff-MDSCs increased perfusion of the ischemic limbs by inducing angiogenesis (shown by increased capillary densities) and by increasing the expression of genes Vegf and Egr-1, which led to an overall decrease in necrosis rate. Induction of angiogenesis in MDSCs by VEGF and its importance in graft implantation as well as improvement of ischemia in cardiac myocytes have been previously described [5 –9]. In these studies, MDSCs were engineered with retroviral vectors containing genes encoding VEGF, so that VEGF could be continuously expressed after injection into study animals. VEGF produced by these retrovirally transduced genes in MDSCs, as well as the ischemic conditions in vivo, may induce angiogenesis by stimulating endothelial differentiation of MDSCs. However, it is has been demonstrated that an overexpression of VEGF is detrimental due to excessive angiogenesis, which hinders engraftment of MDSCs by development of disorganized vascular structures [5] or leads to malignant transformation such as formation of hemangiomas [28,29]. We have induced endothelial differentiation of MDSCs in a controlled, ex vivo environment. This may probably be more efficient in maintaining an adequate VEGF microenvironmental dosage, which has been demonstrated to be important in normal induction of angiogenesis [30]. Furthermore, by providing the drive for endothelial differentiation ex vivo, it is possible to reduce the likelihood of differentiation into other undesired lineages, which could be detrimental in vascular diseases. As these cells are derived from muscles, it is possible for these cells to have a stronger drive toward myogenesis, which under the right conditions may be necessary to undergo angiogenesis by formation of vascular smooth muscle cells, but is definitely undesirable in most vascular diseases (including our second model of intimal hyperplasia). Additionally, the mechanism, as well as the degree of angiogenesis required to improve perfusion of a whole limb, may be different from angiogenesis in other systems, like in the ischemic hearts for improvement of contractility in cardiac myocytes, or in skeletomuscular systems for more efficient cell implantation. This may be why native MDSCs in our study showed some angiogenic features at the cellular level, but were not enough to improve limb perfusion or avoid necrosis. In fact, other studies have shown that native MDSCs produced the desired angiogenic effects under hypoxic conditions, probably by the self-upregulation of VEGF secretion and consequent paracrine effects under the hypoxic milieu [5]. However, the same hypoxic conditions in our study were not enough to prevent necrosis, suggesting that a higher degree of angiogenesis is probably required in vascular diseases. Therefore, we suggest that endothelial differentiation of MDSCs ex vivo with both shear stress and VEGF are a necessary drive to induce differentiation into a homogenous type of endothelial lineage cells, and the expression of VEGF as well as its paracrine effects due to the hypoxic conditions maintains the environment needed to produce a sufficient degree of angiogenesis needed for prevention of ischemia in vascular diseases.
Our second model showed that diff-MDSCs decreased intimal hyperplasia formation in carotid injured rats by deposition of CD31-positive cells in the intima. There are several theories regarding the mechanism underlying intimal hyperplasia formation, but the traditional theory is that after endothelial injury, vascular smooth muscles proliferate and migrate from the media toward the intima layer, causing a thickening of the intima [31 –34]. Furthermore, it has been demonstrated that early re-endothelialization of the injured intima prevents migration of vascular smooth muscle cells, preventing formation of intimal hyperplasia [17,18]. Immunofluorescent staining of lenti-hCMV-GFP-infected diff-MDSCs costained with CD31showed that the cells successfully deposited in the intima layer, with strongest expression at 1 week, but a relatively reduced expression at 4 weeks. This has 2 implications: first, not all diff-MDSCs may be able to survive even after successful implantation, or secondly, the infection of lentivirus may be lost during the replication process; therefore, expression of GFP may decrease even though MDSCs are still present. This latter finding is usually observed in vitro, where a selection process of cells expressing GFP has to be performed after a certain number (around 10 passages) of replications. In either case, early implantation of diff-MDSC is the mechanism by which the process of early re-endothelialization begins. Subsequently, multiple signaling pathways (the importance of paracrine effects has been previously mentioned) may further recruit host cells and thus increase endothelial cell deposition, leading to a decrease in intimal hyperplasia. In fact, it has been previously demonstrated that after MDSC transplantation, most of the cells in the neovasculature are host-derived, with very few donor-derived MDSCs fusing into the blood vessels [5]. The same mechanism may apply to intimal hyperplasia.
This is the first study to demonstrate the potential role of MDSCs in the treatment of vascular diseases. The angiogenic role of MDSCs in other organs has already been reported [5 –9], but the degree of angiogenesis required to treat vascular diseases is presumably much larger, and the mechanism underlying vascular diseases is different and more complex. Our method has the advantage of manipulating the cells in vitro before applying them to target organs, which may not only increase the content of the desired cells, but also allow for cells to have the most optimal features, thus reducing possible side effects. Use of MDSCs and differentiating them to exhibit endothelial progenitor-like features may be more advantageous than using endothelial progenitor cells (EPCs), since EPCs have been shown to originate from different origins [35 –37], and different types of subpopulations (early and late cells) exist even from the same origin [38,39]. Therefore, MDSCs can provide a more homogenous population of cells with possibly a higher differentiation potential.
Our results suggest that MDSCs can be clinically applied to a variety of vascular diseases. It may be used to increase formation of collateral vessels in patients with critical limb ischemia to treat an ischemic ulcer, or to prevent formation of intimal hyperplasia after endovascular intervention of an arterial occlusion. A tissue-engineered vascular graft with MDSCs has been developed previously and has shown promising results in vivo [40]. Angioplasty balloons or stents covered with MDSCs might be used in the future to prevent restenosis and increase patency rates after intervention.
This study has the limitation of having a short follow-up period, since our in vivo studies were observed for only 4 weeks. The long-term effects of diff-MDSCs are not determined, especially the fate of the cells, since one of the most important characteristics of stem cells is their self-renewal ability. Further studies with long-term follow-ups are needed if these cells are to have better clinical applicability. Another limitation is that the hindlimb ischemia mouse model used in this study is a model for acute limb ischemia, but most of the patients with vascular diseases are chronic in nature, and probably, the ischemic conditions are different in chronic limb ischemia. Therefore, future studies with chronic limb ischemia models are needed.
In conclusion, this study describes a method of differentiating MDSCs into endothelial progenitor-like cells ex vivo, capable of inducing angiogenesis, and attenuating intimal hyperplasia in vivo. These findings are very promising, since it may be applied for the treatment of various vascular diseases.
Footnotes
Acknowledgments
This study was funded by the Korea Research Foundation (KFR-2006-331-E00155) and by the Seoul National University Bundang Hospital (SNUBH 02-2005-015), Korea.
We would like to thank Hanmi Yun, RVT for her technical assistance in perfusion measurement.
This study was presented orally at the 12th Annual Meeting of the Asian Society for Vascular Surgery (2011) in Taipei, Taiwan.
Author Disclosure Statement
No competing financial interests exist.