
Abstract
The molecular mechanisms underlying spermine osteo-inductive activity on human adipose-derived stem cells (ASCs) grown in 3-dimensional (3D) cultures were investigated. Spermine belongs to the polyamine family, naturally occurring, positively charged polycations that are able to control several cellular processes. Spermine was used at a concentration close to that found in platelet-rich plasma (PRP), an autologous blood product containing growth and differentiation factors, which has recently become popular in in vitro and in vivo bone healing and engineering. Adipose tissue was surgically obtained from the hypodermis of patients undergoing hip arthroplasty. Patient age negatively affected both ASC yield and ASC ability to form 3D constructs. ASC 3D cultures were seeded in either non differentiating or chondrogenic conditions, with or without the addition of 5 μM spermine to evaluate its osteogenic potential across 1, 2, and 3 weeks of maturation. Osteogenic medium was used as a reference. The effects of the addition of spermine on molecular markers of osteogenesis, at both gene and protein level, and mineralization were evaluated. The effects of spermine were temporally defined and responsible for the progression from the early to the mature osteoblast differentiation phases. Spermine initially promoted gene and protein expression of Runx-2, an early marker of the osteoblast lineage; then, it increased β-catenin expression and activation, which led to the induction of Osterix gene expression, the mature osteoblast commitment factor. The finding that spermine induces ASC to differentiate toward mature osteoblasts supports the use of these easily accessible mesenchymal stem cells coupled with PRP for orthopedic applications.
Introduction
A
During development, the skeleton is formed by 2 different types of ossification depending on the type of bone: intramembranous (the skull bones and the lateral halves of the clavicles) and endochondral ossification (most long bones). The process starts in both cases from the condensation of mesenchymal precursors.
In intramembranous ossification, bone is deposed directly into the mesenchymal anlagen by the newly differentiated osteoblasts. In endochondral ossification, bone is deposed on a template of cartilage defined as “temporary,” because its default fate is terminal differentiation and matrix mineralization, unlike articular cartilage, which is programmed to remain “permanent” and whose functional derangement into a degenerated tissue with many features resembling the changes seen in endochondral ossification is the result of a “loss of maturational arrest” [7,8].
Polyamines are naturally occurring, positively charged polycations that are able to interact with negatively charged compounds and structures within the living cell, thus controlling several cellular processes, including cell differentiation and already shown to be implicated in bone growth and development [9]. Functional genomics in rodents targeting key biosynthetic or catabolic enzymes of polyamines metabolism have revealed a series of phenotypic changes, many of them related to human diseases [10] underlying their ubiquitous involvement in cellular physiology.
The authors previously reported the ability of polyamines to support and enhance endochondral ossification in in vitro 3D cultures of articular osteoarthritic chondrocytes, which retain a “phenotypic plasticity,” recapitulating their designated differentiation programming [11]. It should be noted that at equimolar concentrations, spermine appeared to be more active than spermidine in accelerating endochondral ossification. Indeed, while spermidine was more active in inducing the expression of Sox9 (a transcription factor with a key role in chondrocyte differentiation), spermine was better at inducing both the expression and nuclear localization of Runx-2 (the transcription factor supporting differentiation toward the osteoblastic lineage). It was previously reported that polyamines are more concentrated in the ossifying area of epiphyseal cartilage and accumulate outside the cells where it was suggested that they are directly involved in the mechanism of calcification [12]. In addition, tissue tranglutaminase, which can use polyamines as a substrate, is present in growth plate cartilage, but not in articular cartilage [13]; its expression correlates with events of the terminal differentiation of chondrocytes, and its externalization occurs before matrix mineralization. The osteogenic promoting ability of spermine on ASC has already been reported along with the modulation of spermine metabolism genes during osteogenic differentiation in goat-derived ASC [14], and various differentiation protocols are available for osteogenic differentiation of ASC [15]: spermine seems to be able to enhance the differentiation of precursor cells along the osteogenic lineage, so that the combined use of this molecule with either ASC or SVF might be useful in the development of a one-step surgery approach [16].
In the present study, the effects of exogenously added spermine were investigated during human ASC differentiation recapitulated in 3D micromass cultures, an experimental model closer to in vivo conditions, where differentiation events are also modulated by cell condensation. The object of the study was to determine the effects of spermine on gene and protein expression of key differentiation regulatory transcription factors, markers, and effectors.
The concentration of spermine used in the present study was chosen in accordance with that obtained by high-performance liquid chromatography (HPLC) quantification of PRP samples. PRP is a blood product with high levels of growth factors and cytokines (naturally present in platelets) that promote tissue healing and has been applied in several surgical settings [17]. The main rationale for its use in orthopedics pertains to the ability of platelets to release factors that can recruit mesenchymal cells to an injury site and promote their proliferation and differentiation [18].
The finding that spermine is present in PRP at almost the same concentration used in vitro may help explain the osteo-inductive ability of this blood product [6].
A tridimensional culture model was selected to better assess the effects of the addition of spermine on matrix properties and mineralization and because of its more physiological nature, closer to what occurs in vivo, as it allows the mechanical and biochemical interplay of cells with their surrounding extracellular matrix (ECM) to be reproduced. Moreover, ASC were cultured in 2 different types of media to better determine whether the osteogenesis-promoting activity of spermine is present in different culture conditions. Another aim of the study was to gain further insight into the molecular mechanisms underlying this enhanced osteogenesis, with particular reference to the β-catenin signaling pathway.
The addition of spermine-boosted β-catenin activation thus promoted a selection of the osteogenic rather than the chondrogenic lineage from these multipotent cells.
Materials and Methods
Materials
Spermine and all other biochemical reagents were obtained from Sigma Chemical Company.
Cell isolation and culture
With approval from the local Ethics Committee, adipose tissue was obtained from the surgical subcutaneous area of 24 patients (age 26–74), 15 men and 9 women, undergoing hip arthroplasty. Tissue from these patients was harvested consecutively in a 2-year period without any a priori selection except obvious exclusion criteria such as the presence of rheumatic diseases, diabetes, hemocoagulation diseases, chronic infective diseases, or cancer. Supplementary Table 1 (Supplementary Data are available online at
Flow cytometry
To monitor phenotypic homogeneity, a flow cytometric analysis of the cells at p1 passage was carried out to evaluate the expression of CD 31, 34, 45, 271, 44, 73, 90, and 105. All the antibodies were from Biolegend except for the anti-CD73 antibody (Biosource). Analyses were performed using an FACS Canto II flow cytometer (BD).
Osteogenic differentiation
To evaluate the osteo-inductive effect of spermine addition in 3D culture, ASC were seeded into micromasses [19]. Briefly, 250,000 cells per cryotube were centrifuged (740g, 10 min at 4°C) so that they formed a flattened pellet which was left to mature and release ECM proteins over 3 weeks with medium change every second day. Within a few days, the cell pellet showed a thickened lip and became spherical [20]. The spermine effect was assessed in 2 different culture conditions, and compared with osteogenic conditions (reference). Overall, 5 μM of spermine (S) was added at the time of micromass seeding in the presence of 1 mM of aminoguanidine to control any toxicity of the polyamines due to their oxidation by the amino oxidase present in the bovine serum. Common additives were represented by 100 U/mL penicillin and 100 μg/mL streptomycin.
“Non differentiating medium” (N) corresponded to Dulbecco's minimal essential medium (D-MEM) high glucose, 10% fetal calf serum (FCS), and 50 μg/mL ascorbic acid; “chondrogenic medium” (C) corresponded to the chondrogenic medium by Miltenyi Biotec (Miltenyi Biotec S.r.l.) and “osteogenic medium” (O) corresponded to D-MEM low glucose, 10% FCS, 0.1 μM dexamethasone, 0.2 mM ascorbic acid, and 10 mM glycerol 2-phosphate solution.
For each culture condition, parallel micromass samples were used for either western blot or real-time polymerase chain reaction (PCR) analysis. Other samples were snap frozen in optimal cutting temperature compound and dedicated for immunohistochemistry (IHC) or confocal microscopy essentially as described in [21].
Polyamine analysis of PRP
To assess the polyamine content of PRP, this blood derivative was obtained from 5 different subjects (31–65 years of age, 2 men and 3 women). PRP was prepared according to procedures established at the Istituto Ortopedico Rizzoli [22]. Overall, 500 μL of PRP was added to an equal volume of 0.6 N perchloric acid and stored at −80°C until further analysis. Polyamines were separated and quantified in acidic cellular extracts by HPLC after derivatization with dansyl chloride [23].
Immunoblotting
Evaluation of the expression levels of selected transcription factors, markers, and regulators was carried out by western blotting. To achieve effective extraction of proteins, including those bound to DNA, radioimmunoprecipitation (RIPA) buffer with the addition of benzonase and protease inhibitor cocktail (PIC; Sigma-Aldrich) was used to extract proteins. The composition of the buffer was as follows: Tris-HCl 50 mM pH 7.4, NaCl 150 mM, Nonidet P-40 1%, SDS 0.1%, Na deoxicolate 0.5%, NaF 1 mM, Na3VO4 1 mM, PMSF 1 mM, 1:200 PIC, and 100 U/mL benzonase. Briefly, total cellular lysates were obtained by solubilizing micromasses with RIPA buffer in addition to a vigorous homogenization with disposable pestles (Sigma) and sonication. Aliquots of micromass lysates corresponding to one or half micromass were loaded in the wells of Nu-Page precast 4%–10% polyacrylamide gels (Invitrogen), which were subsequently transferred onto polyvinylidene fluoride membranes by a dry electroblotting method using I-Blot (Invitrogen) and then subjected to immunodetection exploiting the SNAP-ID device (Merck Millipore). Signals were detected with appropriate secondary antibodies and revealed with ECL Advance kit (Amersham), using the CCD camera acquisition system of Image Station 4000 MM and Carestream Molecular Imaging Software 5.0. (Carestream Health, Inc.). Runx-2 was stained with goat polyclonal antisera (R&D Systems). Cyclin D1 and caspase 3 were stained with rabbit polyclonal antibodies (Santa Cruz Biotechnology). γH2AX (phospho-Histone H2A.X, Ser139) was detected by means of a rabbit polyclonal antibody (Upstate–Millipore). Monoclonal anti-GAPDH (clone 6C5, Chemicon–Millipore) or beta-actin (Sigma) served as loading controls. At least 4 different experiments were carried out for each analysis.
Quantification of mineralization
To carry out a quantitative determination of the mineralizing ability of cultures on treatment with spermine, the insoluble pellet left after protein extraction of the micromasses was further treated overnight at room temperature (RT) on agitation with 50 μL of 0.5 M HCl, and then calcium was quantified by the Quantichrom DICA-500 assay (Bioassay Systems). In 2 cases, ASC were also seeded at high density and grown in monolayers, 6,000 cells per well. Overall, 50 μL of 0.5 M HCl was directly added to the cultures in the wells, and calcium was subsequently evaluated according to Quantichrom DICA-500 assay protocol.
Micromasses were also scored for mineralized areas by alizarin red staining as previously described [19].
Real Time PCR
Real-time PCRs were performed as previously described [21,24] with Forward (F) and Reverse (R) PCR primers, including GAPDH (NM_002046) 579-598F and 701-683R); RUNX-2 variant transcript 3 (NM_004348) 864-883F and 968-949R; RUNX-2 variant transcripts 2 (NM_001015051) and 1 (NM_001024630) 716-735F and 820-801R; Sox9 (NM_000346) 952-968F and 1069-1054R; alkaline phosphatase (ALP) variant transcript 3 (NM_001177520.1) 1080-1097F and 1231-1214R; variant transcript 2 (NM_001127501) 1196-1213F and 1347-1330R; variant transcript 1 (NM_000478.4) 1361-1378F and 1512-1495R; β-catenin (NM_001904) 1031-1052F and 1313-1293R; Osterix variant transcript 2 (NM_152860.1) 1204–1224F and 1538–1517R and variant transcript 1 (NM_001173467.1) 1219-1239F and 1553-1532R. Most primers were annealed at 56°C, except for Sox9 at 60°C and Osterix at 65°C. Eight different experiments were analyzed.
Immunohistochemistry
Safranin-O and Alizarin Red staining in 5 μm sections of micromasses fixed with 4% paraformaldehyde were performed as previously described [21].
Micromasses were prepared and analyzed by IHC along with hematoxylin counterstaining of nuclei essentially as described, and signals were developed with a biotin-/streptavidin-amplified, alkaline-phosphatase-based detection system (Biogenex) with fuchsin as the substrate. Antigen unmasking was carried out with 0.02 U/mL chondroitinase ABC 20′ at RT. Runx-2 was detected by overnight incubation with a rat monoclonal antibody (R&D). Sox9 expression was evaluated after antigen unmasking with a rabbit polyclonal antiserum (R&D). Active Caspase 3 was detected by a rabbit polyclonal antibody (R&D). Collagen type I was detected with a mouse monoclonal antibody (clone 5D8-G9, Millipore).
Confocal microscopy
Nuclear translocation of Runx-2 or active β-catenin was evaluated in 4 different experiments by confocal microscopy using double immunofluorescence in 4% paraformaldehyde-fixed 5 μm sections of micromasses established with ASC under different culture conditions. Runx-2 staining was performed with 5 μg/mL rat anti-hRUNX-2 antibody (R&D) with signals revealed by an anti-rat TRITC antibody (1:100) (SIGMA), while active β-catenin was stained with an anti-active (unphosho) β-catenin (Millipore, clone 8E7) detected with a Donkey anti-mouse Dy Light 647 (Jackson Laboratories). In samples derived from 3 different donors, evaluation of the extent of DNA oxidative damage as a function of donor age was carried out by confocal evaluation of the nuclear signal of γH2AX by a mouse monoclonal antibody (clone JBW301, Upstate–Millipore). Nuclear staining was observed with Sybr green (1:10,000-fold dilution) (Molecular Probes). The acquisition of TRITC-labeled anti-hRUNX-2 antibody or Dy Light 647-labeled anti-active β-catenin and Sybr green-labeled DNA signals was performed using a Radiance 2000 confocal laser scanning microscope (BioRad), equipped with argon-krypton and red diode laser (excitation at 488, 568, and 637 nm for Sybr Green, TRITC, and DyLight 647 fluorescence, respectively). The Sybr green emission signal (rendered in green) was detected by a photomultiplier tube preceded by an emission filter (HQ 515/30 nm), the TRITC emission signal (rendered in red) was detected by a photomultiplier tube preceded by an emission filter (E580LP), and the Dy Light 647 signal (rendered in red) was detected by a photomultiplier tube preceded by an emission filter (HQ660LP). 200 μm×200 μm fields (acquired with a Nikon plan apo 60x/A 1.40 oil objective) of sections derived from micromasses grown in different culture conditions were analyzed. Optical sections were spaced ∼0.5 μm along the z axis, and were digitized with a scanning mode format of 512×512 pixels and 256 gray levels. Only sections passing through nuclei were submitted to confocal analysis. This was done to avoid confocal sections acquired either above or beneath the nuclear planes, which could give somewhat misleading information on fluorophore co-localization. Image processing was performed with ImageJ 1.42q software (NIH) and NIS (Nikon Imaging Software) elements (Nikon, Inc.); image rendering was performed with Adobe Photoshop. Co-localized pixels appeared as yellow-orange with varying degrees of luminosity depending on the intensity of the fluorophore signal and the extent of co-localization. Both Runx-2 and β-catenin nuclear localization were reported as Mean Intensity (ie, usual statistical mean of intensity values of pixels, NIS Elements AR–Image Analysis) of objects identified by a binary mask created on Sybr green-stained nuclei areas.
Statistics
Cumulative representations of data are expressed as mean±standard error of the mean (SEM). PRP polyamine content, gene expression, and mineralization data refer to different patients, while confocal analysis data are relative to a representative patient and show the cumulative expression of the intensity of several hundreds of different cells for each condition.
Due to the small size of data sets, which is typical for primary patient samples, nonparametric statistics were employed in most cases. Correlation was tested by Spearman rank-order correlations. The Wilcoxon-Matched Pairs Test was used to compare the effect of spermine addition on gene expression or mineralization. The t-test for independent samples was used for evaluating spermine effect by confocal analysis data. All data were analyzed with STATISTICA 7 statistical software (StatSoft). Tests were considered significant when P<0.05, with *P<0.05; **P<0.01; ***P<0.001.
Results
Characterization and yield of ASC
To monitor phenotypic homogeneity of ASC, superficial stem cell markers were analyzed in flow cytometry using cells in culture at passage 1. This was done in 10 patients. While CD105 staining was dim, (data not shown), ASC samples were highly positive for CD44, CD73, and CD90. Age failed to reach a statistical significant correlation with either the percentage or the fluorescence intensity of the various markers. Staining of CD31, CD45, and CD271 (not shown) was almost negative, while CD34 expression was limited to <30% of the cells (Fig. 1A).
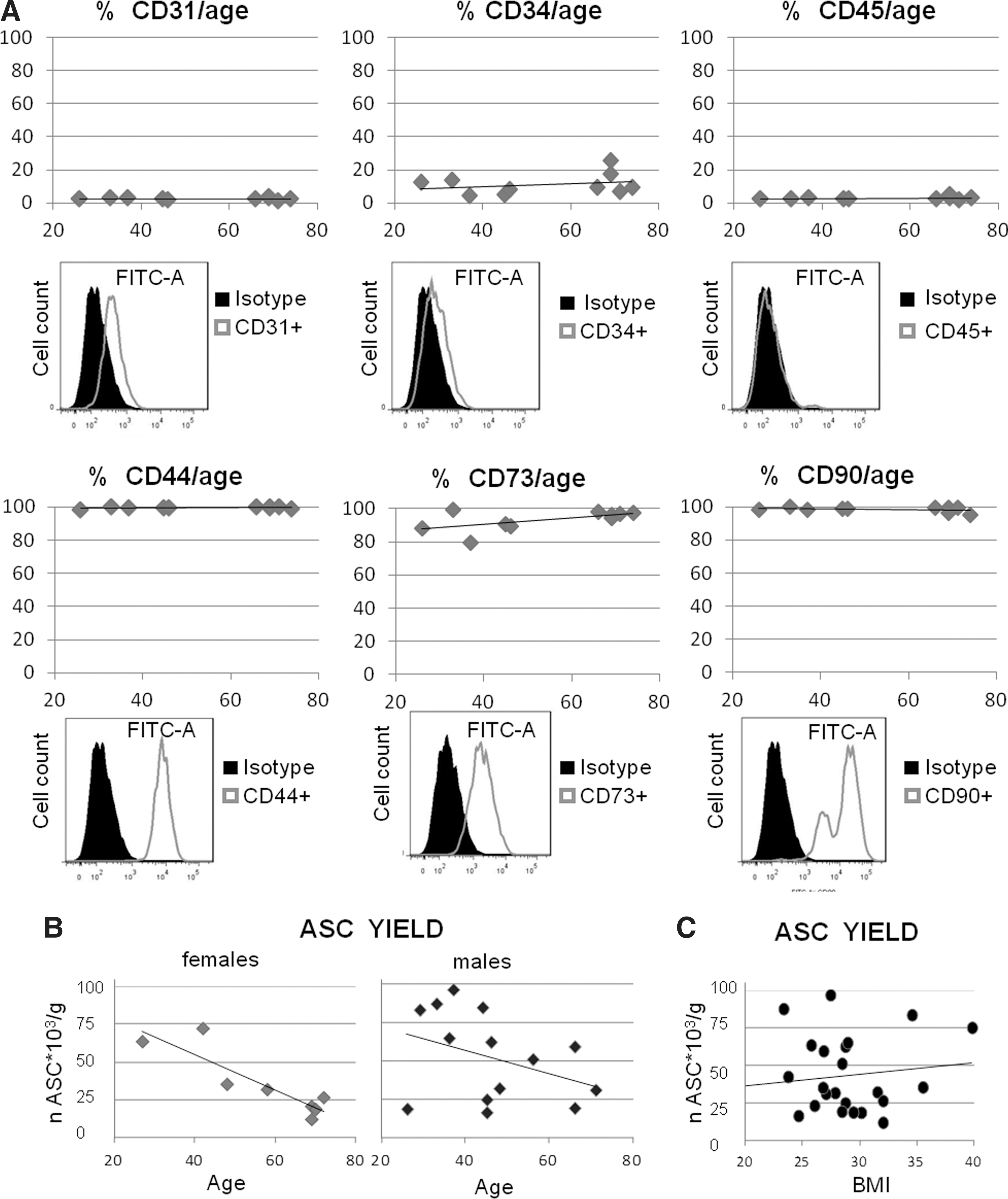
Flow cytometric analysis and yield of adipose-derived stem cell (ASC).
The Mean±SEM cell number of the SVF per gram of adipose tissue was 817,504±111,306. Considering that ASC account for about 5% of the SVF cells, the average yield was 42,088±5,617 per gram of tissue. On exclusion of the data obtained from severely obese patients (BMI>35), the number of ASC per unit (g) of adipose tissue was inversely correlated with the age only in the case of female patients (Sperman Rank-Order Correlations=− 0.79, P<0.05; Fig. 1B). No correlation was found between the number of ASC per unit (g) of adipose tissue and the BMI of the patients (Fig. 1C).
The evaluation of signaling pathways responsible for the osteo-inductive ability of spermine was carried out in 3D cultures with ASC obtained from patients below 50 years of age because of the poor quality of the tissue obtained from older patients.
Analysis of polyamine content of PRP samples
HPLC analysis of 5 PRP samples after acidic extractions indicated the following mean±SEM polyamine content (μM): putrescine: 0.39±0.21; spermidine: 8.26±1.11 and spermine: 7.70±1.96 (Fig. 2). Therefore, the concentration of 5 μM was chosen to evaluate the spermine effect on maturing micromasses, as it may fall within the range of spermine bioavailability on PRP delivery. In selected experiments, the dose-dependent effect of the addition of spermine or other polyamines alone or in combination with cells cultured either in a monolayer or in a micromass was also evaluated.
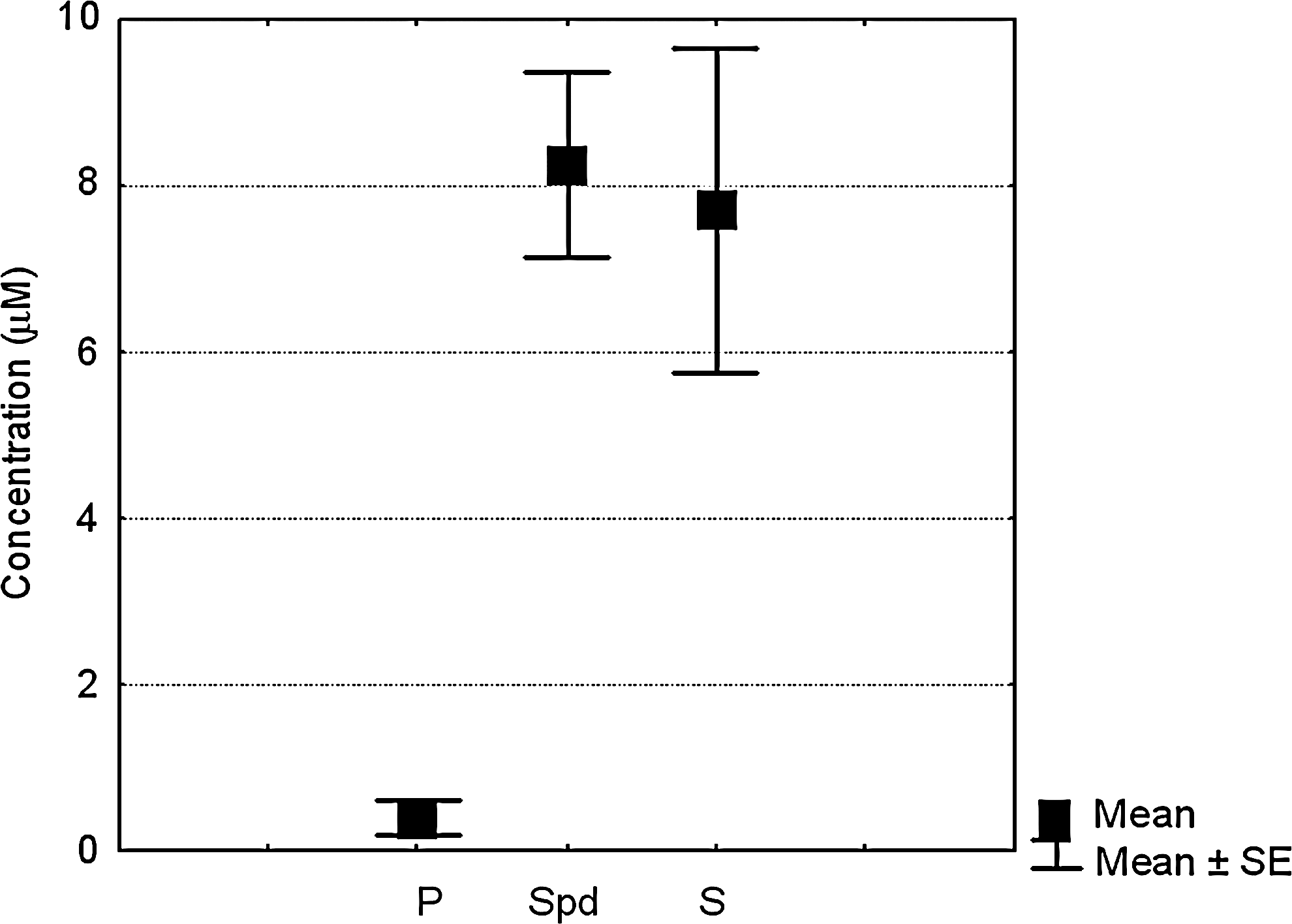
Polyamine content in platelet-rich plasma (PRP). High-performance liquid chromatography analysis of PRP derived from 5 patients, and obtained after acidic extraction. P: putrescin, Spd: spermidine, S: spermine.
Basal commitment of ASC micromasses
Within 2–3 subculturing passages, ASC were seeded in micromass cultures, and an evaluation of cell basal commitment was carried out at 1 week of maturation.
IHC performed to stain the expression of master transcription factors relevant for differentiation of skeletal precursors showed a much higher expression of Runx-2 compared with Sox9, in keeping with an already acquired ability by ASC to mineralize their matrix (Fig. 3, left panel). This prevalence was observed in all culture conditions (non differentiating medium, N; chondrogenic, C; osteogenic, O), with a more prominent positivity in the presence of exogenous chondrogenic or osteogenic stimuli compared with non differentiating conditions. Indeed, very rare reliable Sox9 signals were appreciable only in pellets from cells grown in chondrogenic medium (Fig. 3A, left, inset), whereas the Runx-2 signal was much stronger (Fig. 3A, right), at just 1 week of maturation, thus indicating an acquired osteogenic commitment of these cells. This is in keeping with reports highlighting an increased chondrogenic potential of ASC only after extended passaging [25]. Safranin-O and Alizarin Red stainings (Fig. 3B) corroborated the observation of a pre-existing commitment of ASC toward the osteogenic lineage.

Analysis of basal chondrogenic/osteogenic commitment of ASC micromasses. Within 2–3 subculturing passages, ASC were seeded in micromass cultures and an evaluation of their pre-existing commitment was carried out at 1 week of maturation.
Differentiation effects of spermine addition at the level of key differentiation transcription factors
To understand the molecular mechanisms underlying the osteoinductive properties of spermine, the effects of its addition to ASC micromasses were evaluated throughout their maturation. The effects of spermine on the expression of key differentiation transcription factors was evaluated at mRNA level (Fig. 4) in micromasses grown in chondrogenic medium to determine the ability of spermine to induce osteoblastogenesis also in the presence of alternative differentiative stimuli. The addition of 5 μM spermine enhanced osteoblastogenesis in chondrogenic medium as shown by the consistently increased expression of Runx-2 mRNA at 1 week, whose level approached that of cells cultured in osteogenic medium (data not shown) (Fig. 4A). The early effect on Runx-2 suggests that the compliance for Runx-2 modulation is something which occurs early in the maturation process, and might be needed to prompt subsequent processes. Although the present results partially agree with previous published observations [14], the novelty of the present study is the evaluation of the effect of spermine on human ASCs in 3D cultures, a condition that is closer to what may occur in vivo, where differentiation events are triggered by mesenchymal cell proliferation and condensation.
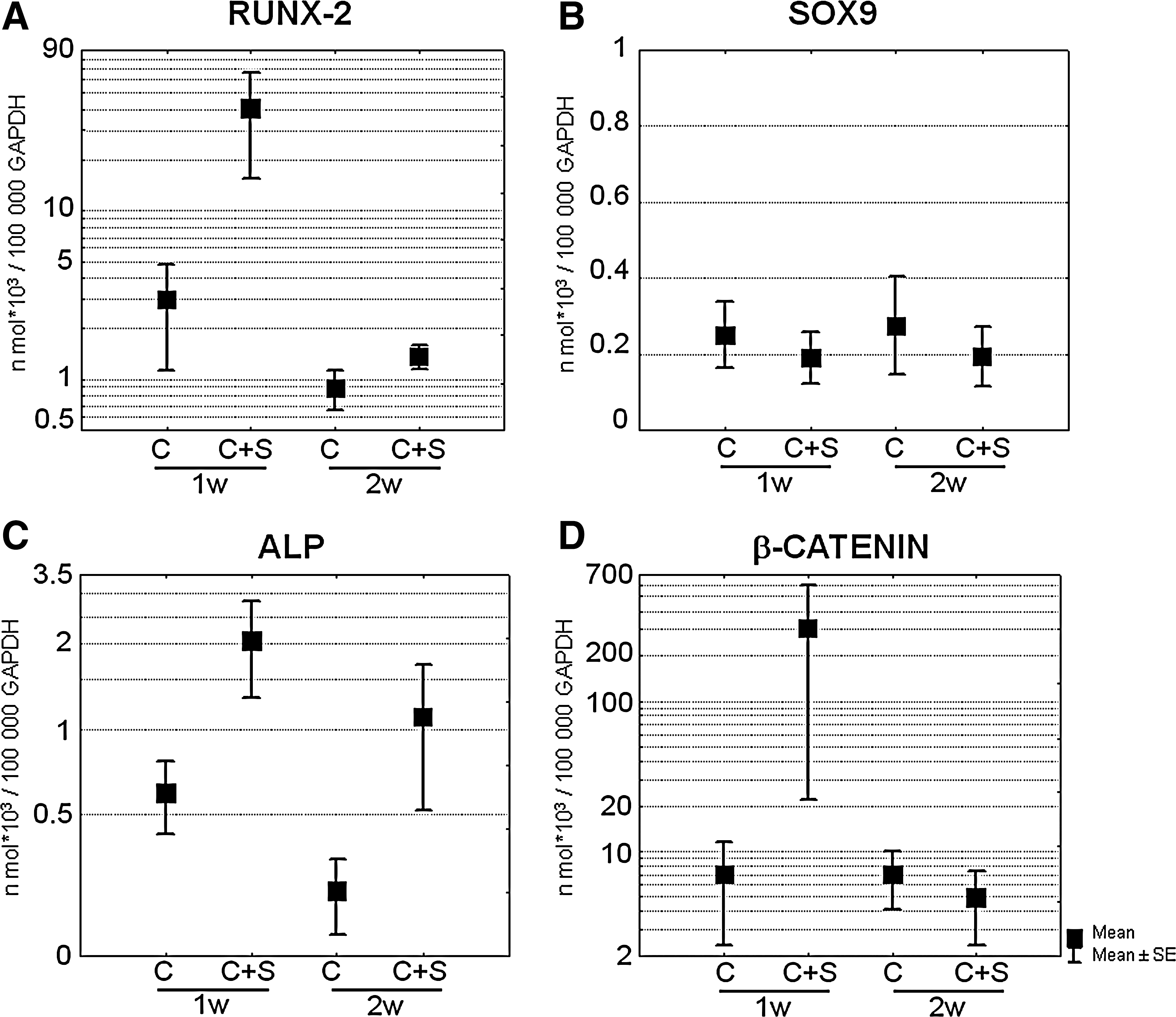
Effects of spermine on major differentiation transcription factors and markers. Cumulative data showing the effects of spermine addition on the expression level of Runx-2
In the primary culture of human chondrocytes, the authors previously reported that spermidine and spermine showed differential, complementary activities, with spermidine more active in inducing Sox9 protein, showing a stronger chondrogenic effect and spermine more active in inducing Runx-2 thus proving to be more osteo-inductive [11]. Real-time PCR analysis indicated a higher expression of Runx-2 (Fig. 4A) compared with Sox9 (Fig. 4B) in terms of number of molecules per 100,000 GAPDH molecules, which after spermine addition became more than 2 log higher at 1 week of maturation. Indeed, spermine addition selectively increased Runx-2 expression, without affecting Sox9 expression, and, therefore, its function is likely to serve as an osteogenic commitment stimulus to ASC (Fig. 4A, B), to support the lineage decision of these skeletal precursors.
Alkaline phosphatase mRNA, another early marker of the osteoblast lineage, paralleled the behavior of Runx-2, and the inductive effect of spermine was still evident at 2 weeks of maturation (Fig. 4C).
It has been reported that lineage decision occurs at an early stage but final osteoblast commitment requires β-catenin activity which leads to the expression of osterix, the osteoblast commitment factor [26,27]. Therefore, β-catenin mRNA expression was evaluated after the addition of spermine and, once again, it was found that at early time points, spermine led to an increased expression (Fig. 4D) which approached the level found in osteogenic medium (data not shown). Due to the high variability of the data obtained from different patients and the limited size of the sample, no statistical significant difference was found after the addition of spermine.
Nuclear translocation of transcription factors directing early and mature osteoblast differentiation phases
An analysis of the signaling pathways triggered by spermine using the confocal microscopy was performed to look at the phenomenon at the nuclear level. Despite a degree of biological variability that affected the timing of the effects at least to some extent, the maturation process of ASC micromasses derived from the 4 patients analyzed followed the mechanistically determined process described below.
Runx-2 is a pivotal factor for mammalian bone development. The regulation of its function occurs at multiple levels, including transcription, translation, and posttranslational modification [28]. Among the latter, a nuclear localization signal directs Runx-2 to the nucleus, so that it can exert its activity as transcription factor.
To assay the effect of spermine in increasing Runx-2 nuclear translocation, a confocal microscopy analysis of the intensity of Runx-2 signal in the nuclei was performed.
In the example shown in Fig. 5 (A and C) at 1 week of maturation, spermine significantly decreased the signal of nuclear Runx-2 compared with the unstimulated condition in both non differentiating and chondrogenic medium. The pattern of nuclear Runx-2 expression paralleled that of total Runx-2 as evaluated in terms of intensity per area unit or by western blotting (data not shown). It should be noted that this trend was the opposite to that of gene expression: despite the increase in mRNA, the portion of functional active Runx-2 was surprisingly lower after the addition of spermine, thus suggesting post-translational modifications. Indeed, maturation of osteoblasts requires β-catenin activity, which may then lead to the inhibition of Runx-2 transcriptional activity via phosphorylation-dependent ubiquitination and degradation mediated by its target gene Cyclin D1 [28,29]. Therefore, β-catenin activation was analyzed in the same samples, by evaluating its nuclear localization, and indeed a significant increase in this signal was observed in samples treated with spermine (Fig. 5B, C). These data suggest that spermine can speed up the progression from the early to the mature phase of osteoblast maturation.
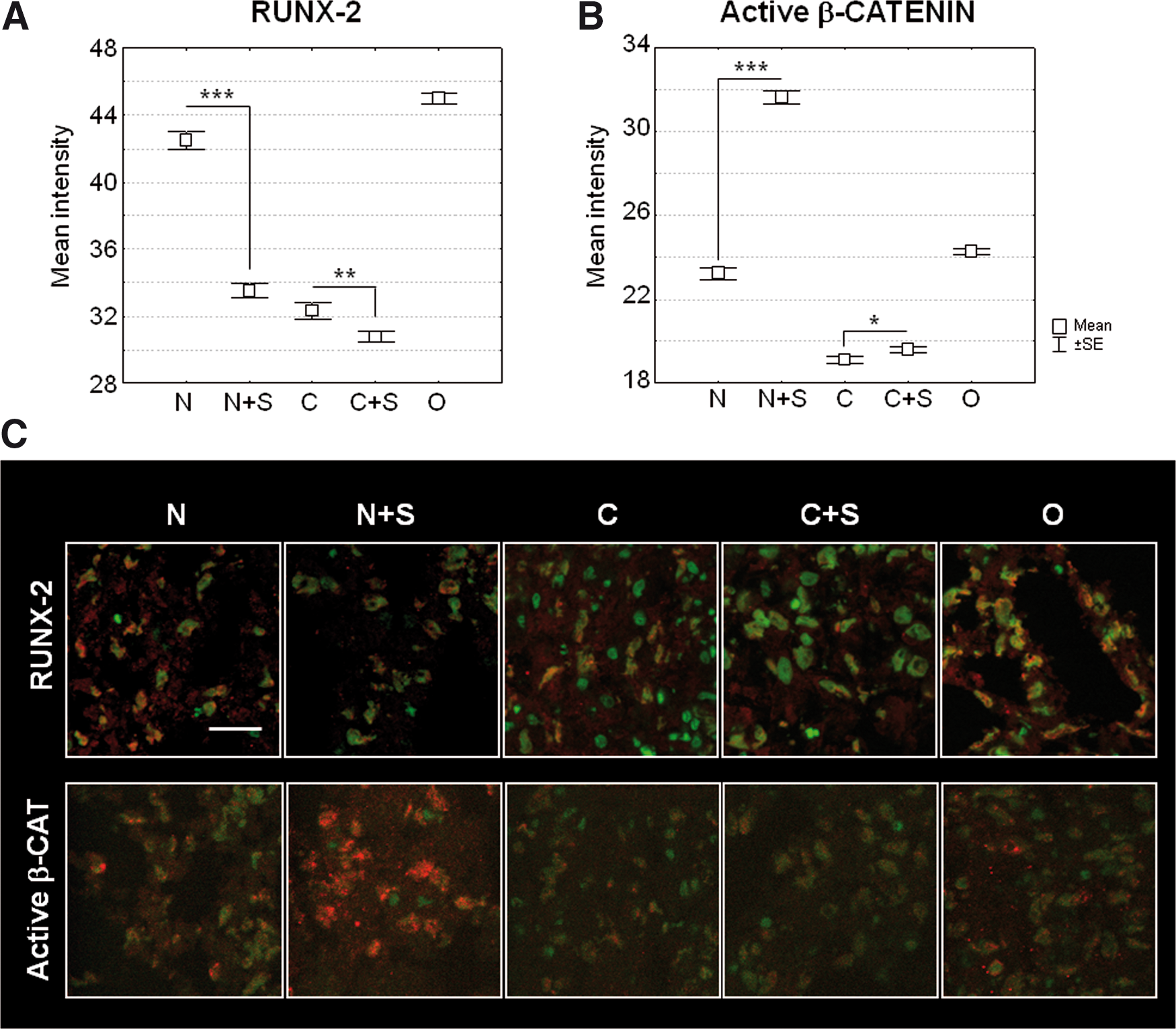
Nuclear translocation of transcription factors directing early or mature osteoblast differentiation phases.
Conversely, some patients presented a progressive increase in Runx-2 expression from 1 to 2 weeks (Fig. 6A) that was further significantly enhanced by spermine addition, and showed a spermine-dependent up-regulation of active β-catenin at 2 weeks of maturation (Fig. 6A, B). In the patient example shown in Fig. 6, spermine sustained β-catenin activation, and this resulted in a correlated increase of Runx-2 nuclear expression at 2 weeks. It has to be underlined that the results on Runx-2 protein expression might vary depending on the balance between the direct β-catenin stimulating effects [30] and the cyclin D1 ubiquitination activity [29].
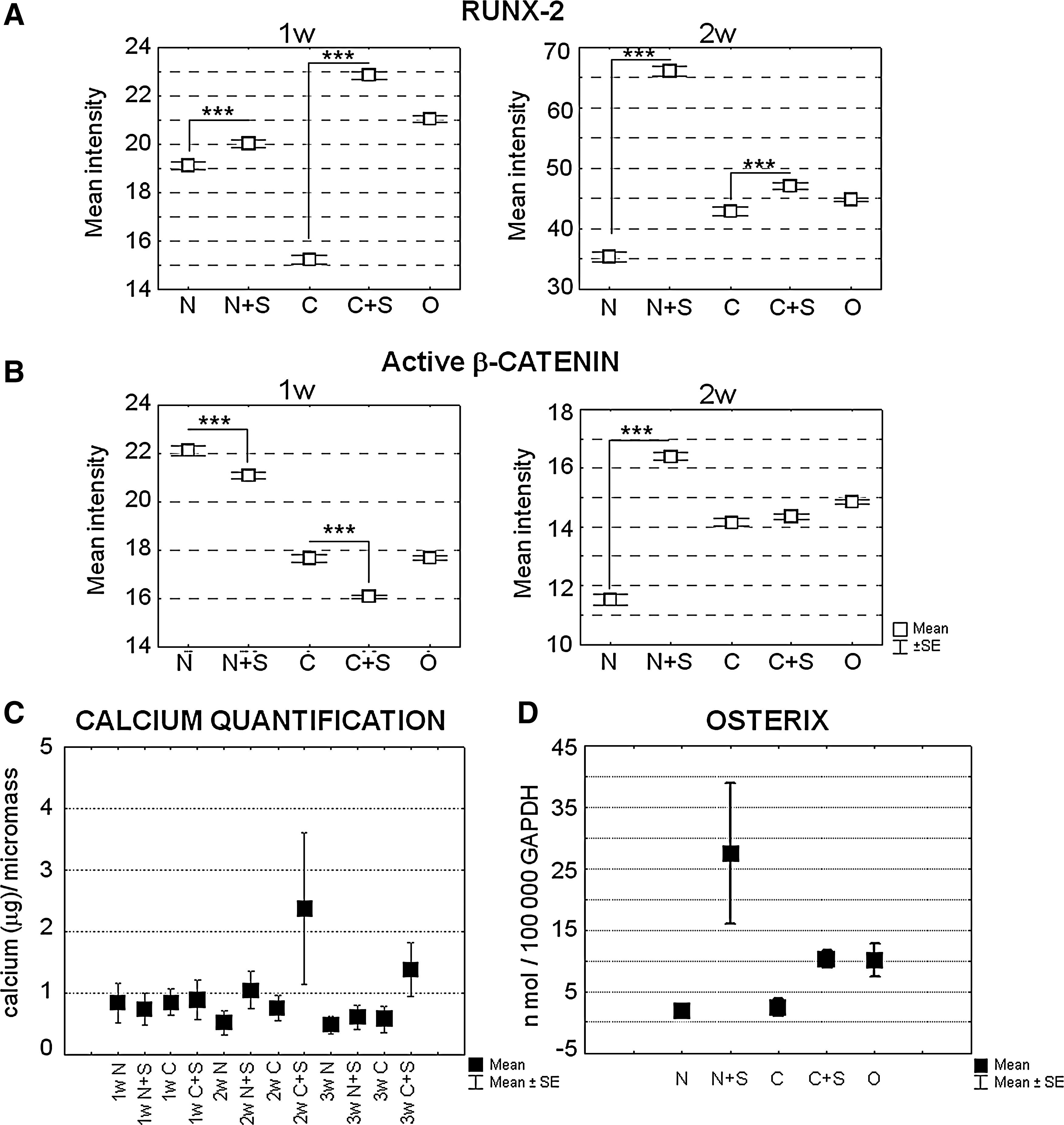
Kinetic analysis of spermine-dependent nuclear translocation of Runx-2 and active β-catenin and effects on matrix mineralization and osterix expression. Cumulative data of nuclear intensity of Runx-2
The effects of this anticipated osteoblast phenotype on mineralization are seen in Fig. 6C, which shows that the level of soluble calcium (μg) obtained from the acid treatment of micromass pellets was already higher in spermine-stimulated samples at 2 weeks of maturation. Supplementary Fig. 1 shows the dose-dependent effects of the addition of different polyamines (spermine, spermidine, and putrescine used at 1, 5, 7.5, and 10 μM and a combination of spermine and spermidine at 5, 7.5, and 10 μM) in the mineralization of ASC derived from the same patient, and tested in triplicate in monolayers, in chondrogenic medium. Overall, 5, 7.5, and 10 μM of spermine or spermidine were also tested in 3D culture. The dose-dependent effect of the addition of spermine or spermidine was markedly different in monolayers compared with the micromass condition. In the latter, the same level of mineralization was obtained with 5 or 7.5 μM concentrations of spermine or spermidine. This might depend on the differential occurrence of polyamine interconversion as a result of a different pattern of the enzymes of the polyamine pathway in the 2 culture conditions.
Effects of spermine on β-catenin targets
As stated earlier, the addition of spermine enhanced the activation and nuclear localization of β-catenin compared with that of control samples. Among the β-catenin putative targets, the first gene investigated was osterix, the osteoblast-specific transcription factor that was considered a pivotal requirement for the differentiation of preosteoblasts into fully functional osteoblasts [26,27]. Spermine led to a marked increase in osterix mRNA at the time point, corresponding to maximum β-catenin nuclear translocation (Fig. 6D). This finding supports the enhanced osteoblast differentiation consequent to β-catenin activation after spermine addition, independent of the effects on Runx-2 expression.
Another effect of β-catenin activation was the increased expression of Cyclin D1 (Fig. 7C), which has been reported to be enhanced by spermine as both protein expression and transcriptional activity [31].
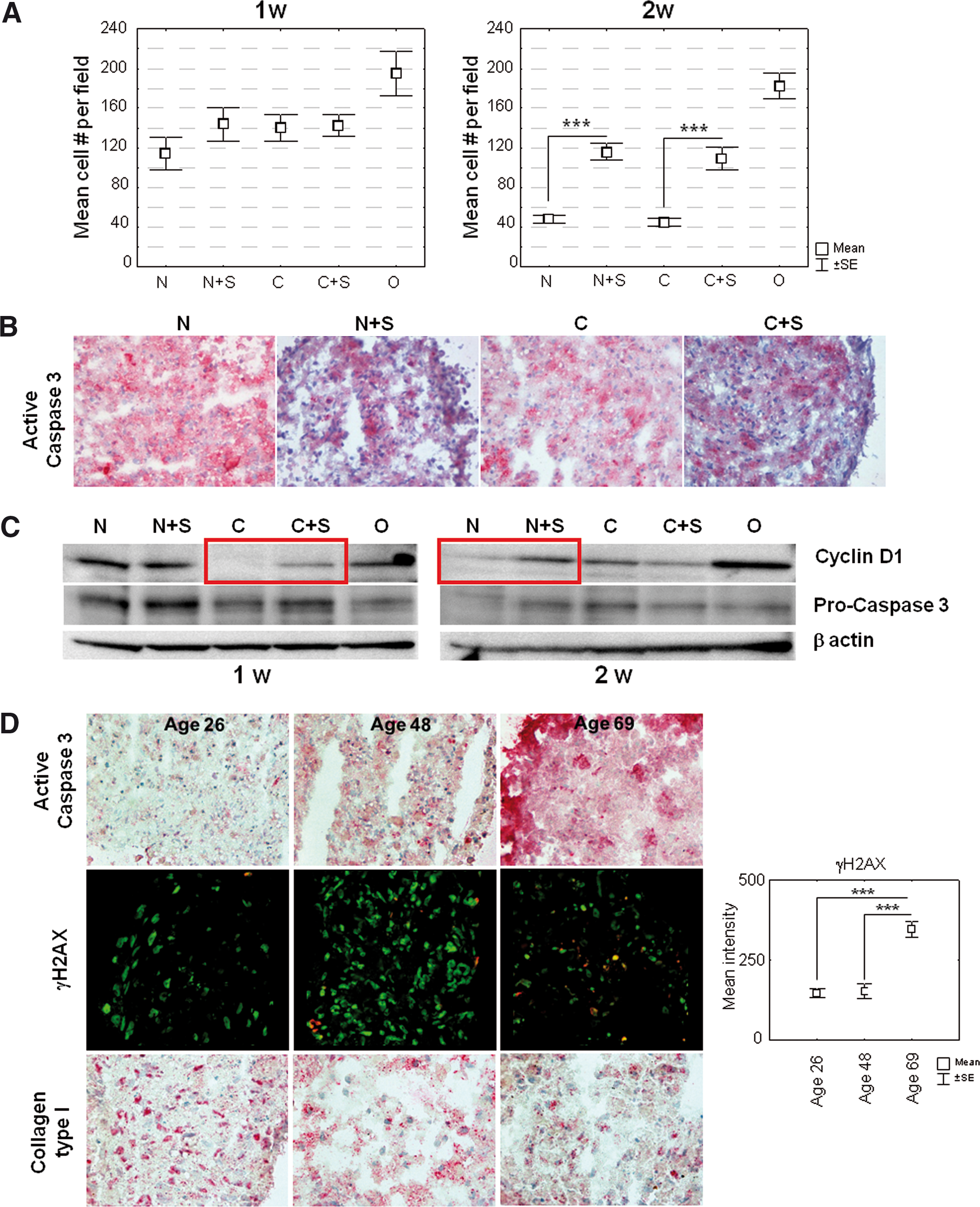
Effects of spermine on cell counts and apoptosis in maturing micromasses.
A nuclei count in 10 different fields randomly taken with a 60x objective in 2 different sections at 2 weeks of maturation showed a significantly higher number of cells in samples treated with spermine in both non differentiating and chondrogenic medium (Fig. 7A). Since the area of the sections examined was similar, the observation of higher nuclei counts in spermine-treated samples strongly supports the idea that spermine might promote ASC proliferation or protect ASC from cell death throughout their maturation in micromasses. The results of IHC and western blotting, the latter showing a higher cleavage of the pro-caspase 3 in the absence of spermine thus yielding more active caspase 3, indicated a lower level of apoptosis after spermine treatment, particularly in 1-week micromasses (Fig. 7B, C). Apoptosis as a function of asymmetric division in association with differentiation is a phenomenon that is better studied in 3D culture [20]. Therefore, the authors investigated whether a different degree of basal apoptosis might help explain the different quality of 3D cultures of ASC obtained from patients with different ages. Samples cultured for 1 week in non differentiating medium from a representative young (26-), adult (48-), and aged (69-years old) donor were assessed. Figure 7D shows that the basal level of apoptosis increases as a function of age (upper row), in parallel with a strong increase in the percentage of γH2AX-positive nuclei, as well as of their signal intensity. These findings strongly suggest that micromasses with cells from older donors have a higher susceptibility to oxidative stress. A higher (more than 2 fold) γH2AX signal in cells from older patients was also confirmed by western blot analysis of micromasses obtained from 2 patients younger and 2 patients older than 50, on normalization versus the hosekeeping gene with reference to a calibration sample run in each gel (data not shown). The increased apoptosis was also associated to an impaired ability to produce ECM proteins, in particular collagen type I, the main component of bone ECM (Fig. 7D).
Activation of the canonical β-catenin pathway has recently emerged as crucial in both intramembranous and endochondral ossification. The evidence collected by the authors' in vitro model suggested that spermine could promote intramembraneous ossification, thus pushing osteoblastogenesis at the level of ASC lineage decision.
Discussion
The appeal of using ASC in regenerative medicine is very high, because adipose tissue samples can be easily obtained by a minimally invasive procedure. Recent data have unambiguously indicated that combined with PRP the ability of ASC to repair critical size defects is similar to that of BM-MSC [6]. The need for supplemental osteo-inductive growth factors has been known for a long time, with convincing data obtained in animal models [32]. However, safety rules preclude the use of recombinant proteins and only allow the use of materials derived from the patients themselves, such as autologous cell transplantation without no manipulation in vitro combined with autologous PRP. Therefore, a future step in bone tissue engineering might involve the use of SVF obtained from patients by dedicated devices, combined with their autologous PRP.
Among various putative growth factors and molecules with regenerative capability contained in PRP, the present study focused on spermine, as this polyamine has already shown to have osteo-inductive ability [14]. Moreover, the pivotal role of spermine in osteogenesis is also supported by the evidence that the Snyder–Robinson syndrome caused by a genetic defect of spermine synthase comprises skeletal defects and osteoporosis [33].
The feasibility of the clinical use of ASC combined with spermine, a component of PRP, was tested for bone tissue engineering by evaluating in vitro the osteo-inductive ability of this molecule in 3-D cultures. These constructs, allowing for MSC condensation, are closer to the in vivo situation compared with most available studies mainly performed in monolayers and with animal-derived cells. It has recently been reported that 3D cultures are a useful tool for investigating the effect of seeding density and nutrients on cell growth and survival and/or apoptosis [34]. Furthermore, the cells were derived from the same anatomical sites that in the future might be used by surgeons to harvest adipose tissue for regenerative purposes during open surgery biopsy or lipoaspiration.
To test the osteo-inductive efficacy of spermine, we combined evaluations at the level of gene and protein expression, matrix mineralization along with an insight into the molecular mechanisms involved in the “ASC-to-osteoblast” differentiation processes to understand both the early and mature osteoblast differentiation phases [26].
The plasticity of the ASC derived from the present series of patients was evaluated in relationship with sex and age. Aging negatively affects the number of ASC obtained per unit of tissue, in women. Moreover, aging affects the efficiency of ECM deposition and the level of oxidative stress, which causes increased DNA damage and apoptosis throughout micromass maturation. Although some studies reveal contrasting results [35 –40], on the basis of our findings we suggest caution in the use of ASC for therapeutic application in elderly patients in keeping with recent reports that highlight impaired functional properties, such as increased DNA damage and senescence [35] and the reduced osteogenic potential [38] of ASC from aged patients.
The present results indicate that spermine consistently leads to increased Runx-2 gene expression in chondrogenic medium. It has already been reported that in human BM-MSC, contrary to what happens in rodent systems, osteoblast differentiation is primarily associated with increased phosphorylation and activity of Runx-2 rather than an overt increase in gene expression and protein [41]. In both non differentiating and chondrogenic medium, an increase in Runx-2 gene expression level was found after addition of spermine, although not statistically significant. However, in both these conditions, we found in some patients an increase in nuclear localization. More precisely, a kinetic observation of the samples revealed a spermine-dependent progressive maturation in the osteoblast lineage, because while some patients showed a progressive increase in the intensity of nuclear Runx-2, in other patients already at just 1 week, spermine caused an evident activation of the β-catenin pathway, as indicated by increased nuclear localization of active β-catenin, along with decreased Runx-2 nuclear localization.
β-catenin activation is indeed the critical event that is used for regulating the fate of osteo-progenitors according to many studies performed both in vitro [42,43] and in vivo [44,45], as the ablation of β-catenin results in a maintained chondrogenic phenotype, despite the ability to express early markers of the osteoblast lineage such as Runx-2.
The findings of increased cyclin D1 in spermine-treated samples suggests that the decreased Runx-2 nuclear localization might be due to cyclin-D1-dependent Runx-2 ubiquitination [28,29], as cyclin D1 is a β-catenin target gene which increases in canonical β-catenin activation.
The present data also indicate that spermine plays an anti-apoptotic role. This might be highly relevant in the perspective of bone tissue engineering, because a higher number of viable precursors can better withstand the apoptotic pressure associated with the process of asymmetric division ongoing in the differentiating 3-D constructs [46]. Indeed, recent studies performed in axin-2 deficient mice have pinpointed the dual role of β-catenin activation in cell proliferation and differentiation, which speeds up the process of intramembranous ossification, thus resulting in craniosynostosis [47]. The anti-apoptotic effect of spermine in ASC is a new finding to the present authors' knowledge, and might be explained by the recently reported ability to down-modulate nitric oxide production in these cells [48]. Indeed, nitric oxide is a highly reactive radical species that along with superoxide can lead to the production of peroxynitrite (ONOO−), a potent cytotoxic oxidant [49].
The importance of polyamines in ASC biology is also supported by evidence that on viable freezing the growth of cultures and the ability to form 3-D constructs is severely impaired. It has been reported in non-animal cells that freezing decreases polyamine content [50]. The low cryotolerance of ASC is well known [51], and these cells lose much of their osteogenic capability after freezing, which also impairs their proliferation rate and the ability to repair critical-sized defects. These functional properties are partially rescued by treatment with growth factors [51], and in particular with insulin-like growth factor, known to increase the activity of ornithine decarboxylase [52], the rate limiting enzyme in polyamine biosynthesis [53].
A pivotal effect of β-catenin activation was the increase in the gene expression of osterix, the final osteoblast commitment factor [27], which enhances the osteogenesis of adipose stem cells [54], further reinforcing the osteoblast lineage decision rather than eventual chondroblast transdifferentiation [26]. It has been clearly shown that osterix acts downstream of Runx-2, but only osterix functions as a no return point in the osteoblast lineage decision [27]; whereas Runx-2-expressing preosteoblasts are still bipotential cells [26,27].
Interestingly, the key role of β-catenin in promoting osteoblastogenesis in stem cells at the expense of other lineages has recently been shown in C3H10T1/2 MSC. On mechanical stimulation mimicking physical exercise [55,56], nuclear translocation of β-catenin has the ability to reduce adipogenesis and to increase osteoblastogenesis at the same time. The beneficial effect of mechanical stimulation might be suggestive of the efficacy of rapid mobilization of patients undergoing bone tissue engineering with ASC.
In conclusion, this study identifies some mechanisms whereby spermine boosts the osteogenesis of ASCs by accelerating progression first through the early and then the mature osteoblast differentiation phases. These maturation effects occur alongside the preservation of viable osteo-progenitors due to the anti-apoptotic ability of spermine on ASC in maturing 3-D cultures.
Since spermine is contained in the PRP, the results of the present study might help explain the efficacy of this blood derivative as a useful additive to combine with ASC or SVF for bone tissue engineering.
Footnotes
Acknowledgments
This work was supported by FIRB (MIUR, Italy) RBAP10KCNS and Fondi cinque per mille (Ministero della Salute, Italy).
The authors wish to thank Prof C. Ventura (Laboratorio di Biologia molecolare e ingegneria delle cellule staminali, Istituto Nazionale di Biostrutture e Biosistemi, Bologna, Italy) for helpful discussion during the early phase of this work; Dr L. Cattini (Laboratorio di Immunoreumatologia e Rigenerazione Tessutale, Istituto Ortopedico Rizzoli, Bologna, Italy) and Dr. Maddalena Zini (Dipartimento di Biochimica, Università di Bologna, Bologna, Italy) for their technical assistance; Dr. Elettra Pignotti for helpful suggestions in the use of statistical tests; and Keith Smith for revising the English language.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.