
Abstract
Lifelong neurogenesis in the mouse olfactory epithelium (OE) is regulated by the response of stem/progenitor cells to local signals, but embryonic and adult OE progenitors appear to be quite different—with potentially different mechanisms of regulation. A recently identified progenitor unique to embryonic OE—the nestin+ radial glial-like progenitor—precedes some Mash1+ progenitors in the olfactory receptor neuron (ORN) lineage, which then gives rise to immediate neuronal precursors and immature ORNs. Neurogenic drive at each stage is governed largely by exogenous factors. Fibroblast growth factor 2 (FGF2) is believed to increase cell proliferation in both presumptive OE stem cells and immediate neuronal precursors in explants, but whether FGF2 directly acts on different target progenitors or stages in the embryonic OE is not known. Here we show that fibroblast growth factor receptor (FGFR)1 and FGFR2 are found in a variety of embryonic olfactory cells, including olfactory ensheathing cells and their precursors, and neuronal nestin+ and Mash1+ progenitors. Combining gain and loss of function for FGF2 activity in a novel in vitro clonal progenitor assay, we reveal that different colony phenotypes are formed by presumably different OE progenitors. FGF2 is essential for the survival and expansion of colony-forming cells of the OE, and also enhances the proliferation of embryonic Mash1+ progenitors, leading to long-lived enhancement of neurogenesis. Our data suggest that distinct OE progenitors yield different in vitro phenotypes with different potentials, that colony-forming activity is profoundly altered by laminin and collagen, that multiple ORNs can be produced from single colony-forming progenitors, and demonstrate a broader progenitor range of FGF action in the embryonic OE than previously demonstrated.
Introduction
T
Neurogenesis in the OE occurs in a stepwise manner, where stem/progenitor cells feed into the ORN lineage at different stages, depending on the feedback regulation provided by cells further along in the differentiation cascade. At the apex, stem cells produce Mash1+ neuronal progenitors, which normally produce Ngn1+ immediate neuronal precursors, which divide a limited number of times before producing committed ORNs [5]. More recent data from transgenic mice and in vitro assays suggest that nestin+ radial glial-like progenitors precede Mash1+ progenitors in the embryonic (but not adult) ORN lineage [15]. Elegant explant and transgenic mouse experiments have demonstrated multiple roles for the transforming growth factor-β (TGF-β) superfamily, leukemia inhibitory factor (LIF), epidermal growth factor/TGF-alpha (EGF/TGF-α), and FGFs in the regulation of olfactory neurogenesis [7,16 –23]. Although FGFs serve a variety of roles during nervous system development, we do not know which specific OE cell subtypes respond to FGF directly, and how FGF impacts neurogenesis or gliogenesis, during embryonic olfactory system development.
FGF1 and FGF2 activate each of the 4 FGF receptors (FGFR1–4), where the level of activation is dependent upon the specific FGF ligand and receptor subtypes [24,25]. FGF2 is largely proneurogenic in the CNS, where it can promote the proliferation, survival, and differentiation of neural stem and progenitor cells [26 –30]. In the E10.5 olfactory placode/pit, FGFR1, 2, and 3 have been detected by immunohistochemistry [31], with only FGFR1 and FGFR2 transcripts detected by RT-PCR by E14.5 [32]. Further, in embryonic OE explant (E14.5) FGF2 increases the number of divisions of immediate neuronal precursors up to 2-fold before producing ORNs, and may increase the proliferation of presumptive OE stem cells [32]. However, we do not know if FGF2 acts directly on nestin+ and Mash1+ embryonic OE progenitors, and previous assay systems have achieved only short-term survival of OE progenitors and neurons. Here, we identify the specific progenitors and cell types expressing FGFR1 and FGFR2 in the embryonic OE. Using gain- and loss-of-function studies with 2 different in vitro assays, we show how FGF signaling can enhance the expansion of nestin+ radial glial-like cells and Mash1+ neuronal progenitors, and thus enhance production of ORNs over an extended period of time in culture.
Materials and Methods
Tissue preparation
Pregnant mice were anaesthetized with Xylaket: 25 mg/mL Ketamine HCL (MTC Pharmaceuticals, Cambridge, ON), 2.5 mg/mL Xylazine (Bayer, Inc., Tarrytown, NY), 15% ethanol, and 0.55% NaCl (120 mg/kg Ketamine and 12 mg/kg Xylazine), embryos dissected out, and rinsed in phosphate-buffered saline (PBS) before being immersion-fixed in 4% paraformaldehyde (PFA) at 4°C overnight, and cryoprotected at 4°C in 10% and 30% sucrose/PBS, 24 h each. Embryos were embedded in the Tissue-Tek medium (OCT; Sakura Finetek, Torrance, CA) before freezing in liquid nitrogen. Coronal, sagittal, or transverse 12–14-μm sections were taken on a Microm HM 500 cryostat, mounted onto charged Superfrost glass slides (Fisher Scientific, Suwanee, GA), and stored at −20°C for subsequent analysis [33]. The day of vaginal plug was defined as E0.5.
Immunofluorescence
Standard conditions
Frozen sections were placed on a slide warmer (42°C) for 10 min, postfixed in 4% PFA, and PBS washed twice, 5 min each. Sections were permeabilized in 0.1% Triton X-100/PBS for 30 min, PBS washed twice for 5 min each, and blocked with 4% normal serum for 20 min before primary antibody incubation (see Supplementary Table S1; Supplementary Data are available online at
Exceptions
Before blocking, FGFR2 detection required a 60-s incubation of the sections with 0.12% trypsin/EDTA (Gibco Life Technologies, Burlington, ON), followed by washing in PBS. Nestin monoclonal antibody (Rat401), but not Nestin polyclonal rabbit antibody (Nestin clone 20), required antigen retrieval. MASH1 signals in vitro were amplified using the Vectastain ABC Kit (Vector Laboratories) and Amplex Red Elisa Kit#2, Horseradish Peroxidase conjugate (Molecular Probes, Eugene, OR), following the manufacturer's instructions, and developed for 5 min before washing in PBS. MASH1 detection in vitro was amplified using biotin secondary antibodies, followed by streptavidin fluorochrome-conjugated antibodies.
Immunocytochemistry
Immunocytochemistry was performed with cells plated onto collagen/laminin-coated glass coverslips. Substrates included collagen (2.5–5 μg/cm2; rat tail, mostly type I; Roche Diagnostics, Mannheim, Germany), laminin (1–2 μg/cm2; Roche Diagnostics, from EHS), or collagen combined with laminin. Immunocytochemistry was performed as outlined for immunohistochemistry; except that for nuclear antigen detection, cells were fixed for 3 min in −20°C methanol and washed in PBS before proceeding directly to serum blocking. For BrdU (bromodeoxyuridine) detection, coverslips were treated with 4 M HCl for 10 min before serum block and treated with primary and secondary antibodies sequentially for NST-Rat BrdU.
Quantification of antigen-expressing cells
The percentage of antigen-positive cells [(antigen+ cell #/total DAPI cell #)×100] was determined by counting ∼20 fields of view per coverslip under 400×magnification for 2 coverslips per experiment, in 3 independent experiments. The total number of cells counted with and without SU5402, a chemical that blocks intracellular FGF signaling through all 4 of its receptors [35,36], ranged 878–4839 for nestin+ cells; 247–633 for MASH1+ cells; 85-344 for NST+ cells, as outlined in the Results section. The number of MASH1+ cells per semiadherent colony was determined in at least 3 experiments, by counting 20 to 55 colonies with a total of 112 to 2526 MASH1+ cells for each colony subtype.
Image analysis
All images were visualized with an Axioskop 2 MOT microscope (Zeiss, Jena, Germany) using a SPOT camera (Diagnostic Instruments, Inc., Sterling Heights, MI) with Northern Eclipse software (Empix Imaging, Inc., Mississauga, ON) and compiled using Adobe Photoshop 7.0.
Embryonic OE cell isolation
E13.5 OE was dissected from head primordia at the level of the first brachial arch, with the lower jaw removed and the OE isolated in its entirety. Pooled E13.5 OE was transferred to PBS with 100 U/mL penicillin and 100 μg/mL streptomycin plus 0.6% glucose. OE tissue was minced into 1-mm2 pieces with an ethanol-sterilized razor blade in a 60-mm petri dish, transferred to a 50-mL tube, and dish washed in PBS/glucose/penicillin–streptomycin (up to 20 mL) before spinning at 160 rcf (relative centrifugal force, or G force), 10 min. All, but 3 mL, of liquid was removed before triturating the cells 25 times with an FBS-prewetted P1000 tip, followed by triturating 30 times with a prewetted polished Pasteur pipette. Tube sidewalls were rinsed with PBS/glucose/penicillin–streptomycin (up to 20 mL) and cells spun at 460 rcf, 5 min. All liquid was removed, and cells were resuspended in 2 mL of Neurocult media (see below) before counting cells on a hemocytometer with trypan blue exclusion used to identify live cells. Cells were resuspended in the medium (0.7–3×105 cells/mL) and centrifuged at 100 rcf for 1 min before plating, to pellet tissue aggregates.
In vitro progenitor assays
Cells from E13.5 OE were cultured in the Neurocult medium: Neurocult, 1×proliferation supplement (Stem Cell Technologies, Vancouver, BC), 20 ng/mL growth factors, EGF or FGF2 (both Sigma, St. Louis, MO), 100 U/mL penicillin, 100 μg/mL streptomycin, and 2 mM
OE cells were plated in 12- or 24-well plates with 1.5 mL or 750 μL media, respectively, and 0.7–2×105 cells/mL, cell densities tested for their clonality [15]. Cells were grown on substrate-coated glass coverslips, with either collagen plus laminin (Figs. 3 and 4), or collagen alone, and then transferred to collagen plus laminin for immunocytochemistry (Figs. 5 and 6). The total numbers of semiadherent colonies or OE neurospheres (dependent upon the substrate) were counted in each well before fixation and processing for immunocytochemistry. OE-nonadherent neurospheres were transferred onto collagen–laminin-coated glass coverslips for immunocytochemistry and allowed to adhere for 3–6 h before fixation. Primary cultures were grown for 7–10 days before counting E13.5 OE neurospheres or semiadherent colonies. An OE neurosphere was counted if >25 μm in diameter; semiadherent colonies comprised of >8 cells were counted.
FGF signal blockade and BrdU labeling
Blockade of FGF signaling in vitro was accomplished by adding 10 μM SU5402 (Calbiochem, Merck KGaA, Germany), to block receptor phosphorylation, or 0.12–4.8 μg/mL bFM-1 (Cell Signaling, Danvers, MA), an antibody specific for FGF2 that blocks ligand binding to the receptor, together with 20 ng/mL FGF2. Controls included FGF2 alone or FGF2 with either vehicle or isotype control antibodies (IgG). FGF signal blockers were added at the time of plating or after 7 days, as indicated in the text. To test for cell proliferation in culture, 0.4 μM BrdU (Sigma) was added into individual wells in 10 μL medium after 7 days and cultured for 1–3 more days before immunodetection of BrdU+ cells.
Statistics
Values shown are presented as±standard error of the mean (SEM). Variability between individual groups was determined using analysis of variance with a student's t-test, and additional statistical significance across multiple groups calculated using the Tukey HSD post hoc test.
Results
FGF receptor 1 and 2 expression in E13.5 OE
To gain insight into how different progenitors and precursors may use FGF receptors to mediate the actions of FGF2 during embryonic OE development, we first used cell phenotype-specific antibodies to test for the coexpression of FGFR1 and FGFR2. Expression of FGFR1 was detected in cells throughout the embryonic OE, spanning from the basal to apical surface (Fig. 1A), but immunoreactivity was more readily detected in discrete apical regions, where subsets of more primitive progenitors reside (Fig. 1A–C). FGFR1 was detected mainly in proliferating cells (expressing proliferating cell nuclear antigen (PCNA)) in the apical, intermediate, and basal OE (Fig. 1B), and some nestin+ and MASH1+ progenitors (Fig. 1C–H). FGFR1 immunoreactivity was rarely detectable as progenitors differentiated into TuJ1+ (β-III neuron-specific tubulin; NST) immature neurons (Fig. 1I).

Fibroblast growth factor receptor (FGFR)1 expression in the E13.5 olfactory epithelium (OE).
FGFR2 immunoreactivity is most highly detected in proliferating progenitors in the olfactory bulb (OB) ventricular zone, and olfactory ensheathing cells (OECs) of the OB nerve fiber layer (ON) (Fig. 2A, B) and olfactory nerve (Fig. 2I, J). Likewise, the majority of chondrocytes in the septal cartilage also express FGFR2 (Fig. 2A, B). Immunoreactivity for FGFR2 was higher in the apical than basal OE, both regions that are enriched in dividing progenitors (Fig. 2C). FGFR2 can be readily detected in nestin-expressing radial glial-like OE progenitors (Fig. 2D), MASH1+ neuronal progenitors (Fig. 2E–H), and partially in some subpopulations of NST+ immature neurons (Fig. 2I). FGFR2-immunopositive cells were also strongly detected in OECs directly surrounding NST+ axon bundles in the lamina propria (Fig. 2I, J) and in nestin-rich presumptive OEC precursors immediately subjacent to the basement membrane in the lamina propria (Fig. 2D). These results suggest that FGFR2 may mediate pleiotropic effects in both neuronal and glial lineages in the embryonic olfactory system.
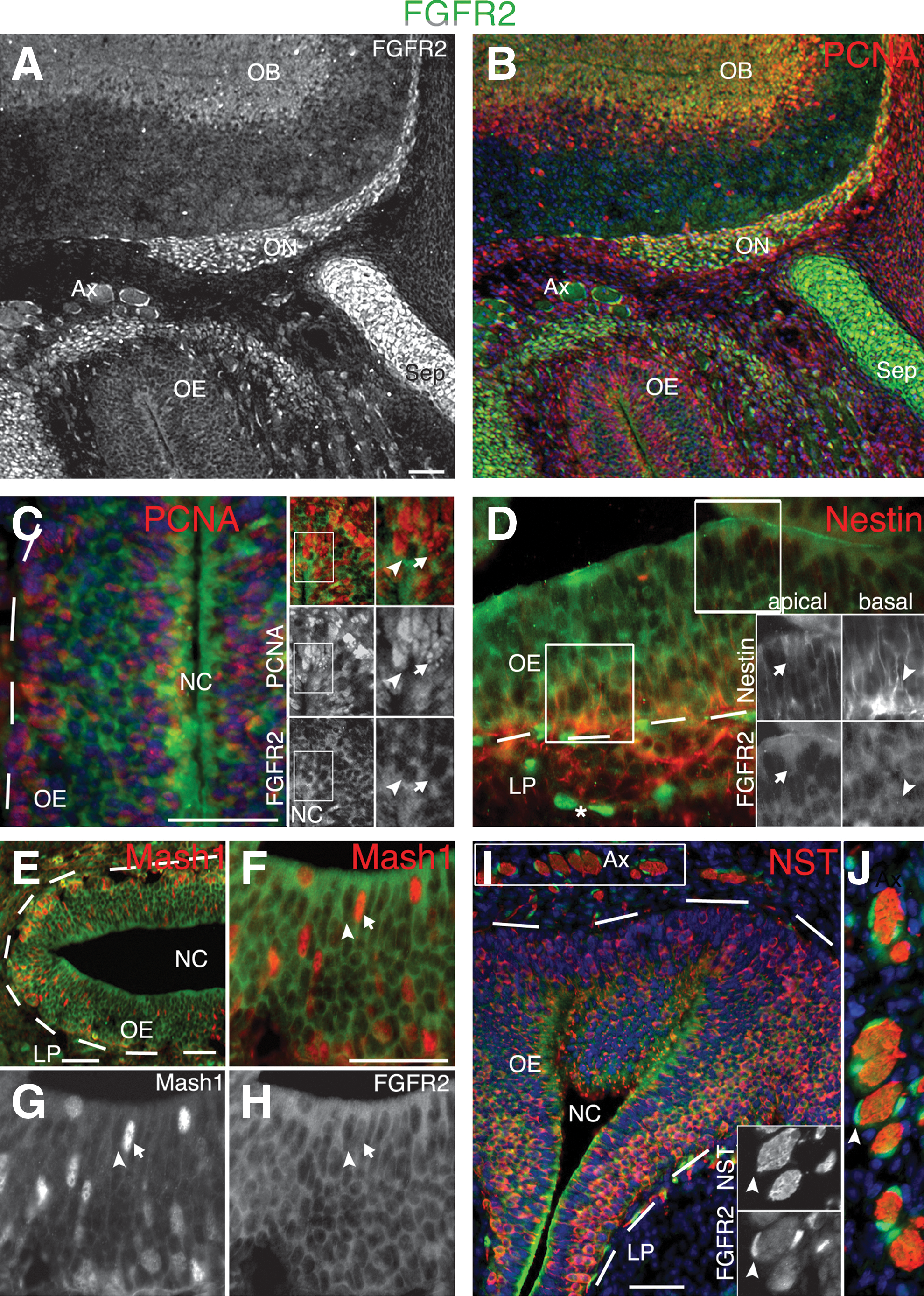
FGFR2 expression in the E13.5 OE.
FGF signaling regulates E13.5 OE colony number and subtypes
To assay the direct impact of exogenous FGF2 on different OE progenitors, we developed an assay for testing the colony-forming ability of dissociated single cells from the E13.5 OE. When plated onto collagen plus laminin under serum-free conditions, OE progenitors at E13.5 form 3 phenotypically distinct semiadherent colony subtypes—spherical, fusiform, and polygonal (Fig. 3A) [15]. All 3 colony subtypes contain nestin+ cells in their cores, and can produce different combinations of neurons and glia [15]. Each colony subtype contains cells expressing FGFR1 and FGFR2, which have a distinct distribution within colonies (Fig. 3B–K; spherical colonies shown). FGFR1 is expressed in cells clustered within the cores of some colonies (Fig. 3B, C), in cell processes on the periphery of colonies (Fig. 3D, E), or restricted to a subpopulation of cells on the outer colony edge (Fig. 3F, G). FGFR2 can be expressed in cell somas and in the processes of core cells (Fig. 3H, J, K). Unlike FGFR1, FGFR2 can also be localized to the nucleus of colony core cells and peripheral cells with large, oval nuclei, similar to S100β+ OECs [15] (Fig. 3I–K).
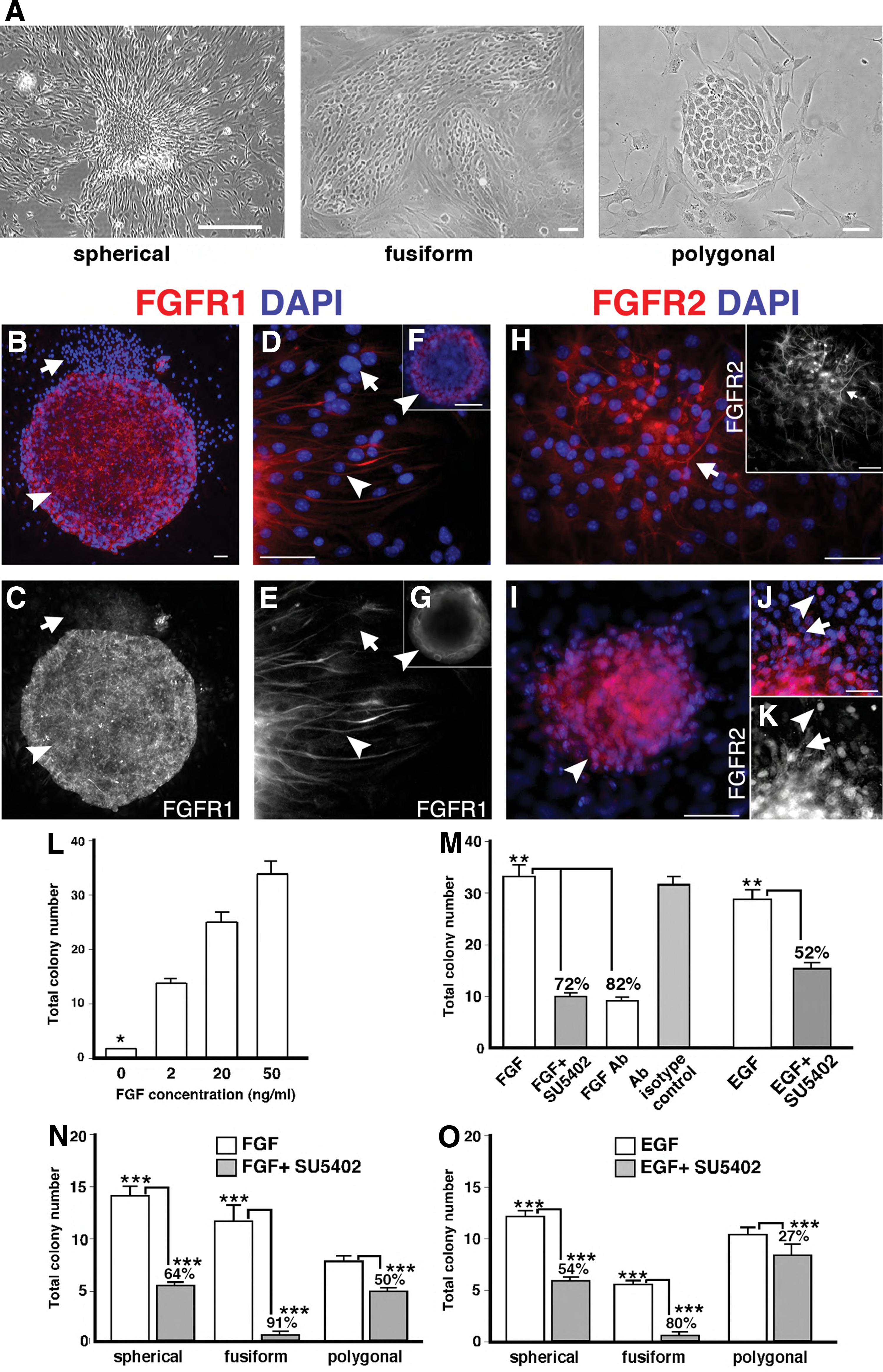
FGF signaling regulates E13.5 OE colony number and subtype. A single-cell suspension of E13.5 OE was cultured in a serum-free medium supplemented with FGF2 or epidermal growth factor (EGF), with or without FGF signal blocking, for 10 days, after which colony number and subtype were assayed.
Given the widespread (and not unexpected) expression of FGFR1 and FGFR2 in olfactory colonies, we next used gain- and loss-of-function studies in a highly defined medium to test how FGF signaling may regulate the production of phenotypically distinct colonies. Without exogenous FGF2, rare colonies were obtained, but FGF2 addition stimulated significant increases in total colony numbers, in a dose-dependent manner (Fig. 3L; no FGF2 vs 2–50 ng/mL FGF2, P<0.001, n=3 representing independent experiments with each condition represented by 3–6 replicates per experiment). After blocking FGF signaling with either SU5402 (10 μM), which blocks phosphorylation of all 4 FGF receptors [35,36], or an anti-FGF2 antibody (1.2 μg/mL) that specifically prevents FGF2-receptor binding, we found significant reductions in colony output (∼80%; P<0.001, n=3; Fig. 3M). Vehicle and isotype antibody controls were similar to FGF2 alone (Fig. 3M, and data not shown). Interestingly, when EGF-treated cultures were simultaneously treated with SU5402 (which does not affect EGF receptor phosphorylation) [35], there was also a significant 52% reduction in total colony number (P<0.01, n=3; Fig. 3M). These results suggest that EGF-treated cultures may be producing paracrine FGF to enhance colony formation (Fig. 3M).
To determine if FGF2 acts on the expansion of colony-forming cells (as opposed to their initial survival in vitro), colony numbers were assessed in FGF2-treated cultures, where the addition of SU5402 was delayed until day 2 of culture. If FGF2 acts directly on colony-forming cells, FGF2 together with the latent blockade of FGF signaling should allow for some colonies to be produced. Compared to the day 1 addition of SU5402, we found a 50% recovery in colony number when SU5402 was instead added on day 2 (data not shown). In addition, the colony sizes were smaller, suggesting that FGF acts on colony-forming cells and also the expansion of their progeny.
In addition to the total colony number, we assessed the effects of EGF and FGF signaling on the types of colonies produced (Fig. 3N, O). In either FGF2 or EGF alone (Fig. 3N,O), spherical colonies were the most abundant (P<0.05, n=3 FGF2; P<0.001, n=3, EGF), followed by fusiform (P<0.05, n=3, FGF2; P<0.001, n=3, EGF). Blocking FGF signaling with SU5402 in the presence of either FGF2 or EGF produced a significant reduction in all colony subtypes (Fig. 3N, O), but especially in fusiform colonies. In FGF2 with SU5402, fusiform colonies demonstrated an 91% decrease (P<0.001, n=3) compared to 64% (P<0.01, n=3) for spherical and 50% (P<0.05, n=3) for polygonal colonies (Fig. 3N). A similar, but less pronounced, reduction was seen when SU5402 was added to cultures with EGF alone (Fig. 3O). These results indicate that FGF signaling regulates the formation of embryonic OE colonies, and the expansion of different colony subtypes is promoted by both FGF2 and EGF.
FGF signaling increases the number of colonies containing Mash1+ neuronal progenitors
Previous studies using embryonic OE explants suggest that FGF2 supports neurogenesis by acting on immediate neuronal precursors and rarely on presumptive OE stem cells [32]. Given that the in vivo expression profile (Fig. 1) suggests that multiple progenitor subtypes express FGFR1 and 2, we directly tested if FGF2 directly activates Mash1+ neuronal progenitors, which are downstream of neural stem cells and precede immediate neuronal precursors. When grown in either FGF2 or EGF, ∼60% of all colonies contain detectable MASH1+ cells (Fig. 4A), which are found in 75%–78% of fusiform, 62%–68% of spherical, and 53%–41% of polygonal colonies. Blocking FGF signaling with SU5402 reduces the total number of colonies containing MASH1+ cells 67% in EGF alone (P<0.01, n=3, representing independent experiments with each condition represented by 3–6 replicates per experiment) and 85% in FGF2 alone (P<0.001, n=4) (Fig. 4B, C). When SU5402 is added to FGF2, the number of fusiform colonies is dramatically reduced, and they fail to produce any detectable MASH1+ progenitors (Fig. 4B). Additionally, the number of colonies containing MASH1+ cells is reduced for both spherical (75%; P<0.01, n=3) and polygonal (90%; P<0.001, n=3) colonies (Fig. 4B). When FGF signaling is blocked in EGF-treated cultures, significant reductions in the number of colonies containing MASH1+ progenitors are found only for spherical (P<0.01, n=3) and fusiform colonies (P<0.05, n=3), but not polygonal (Fig. 4C). These results demonstrate that FGF signaling directly impacts the development of MASH1+ progenitors within all types of colonies, but that spherical and fusiform neurogenic colonies are the most affected by FGF loss of function.
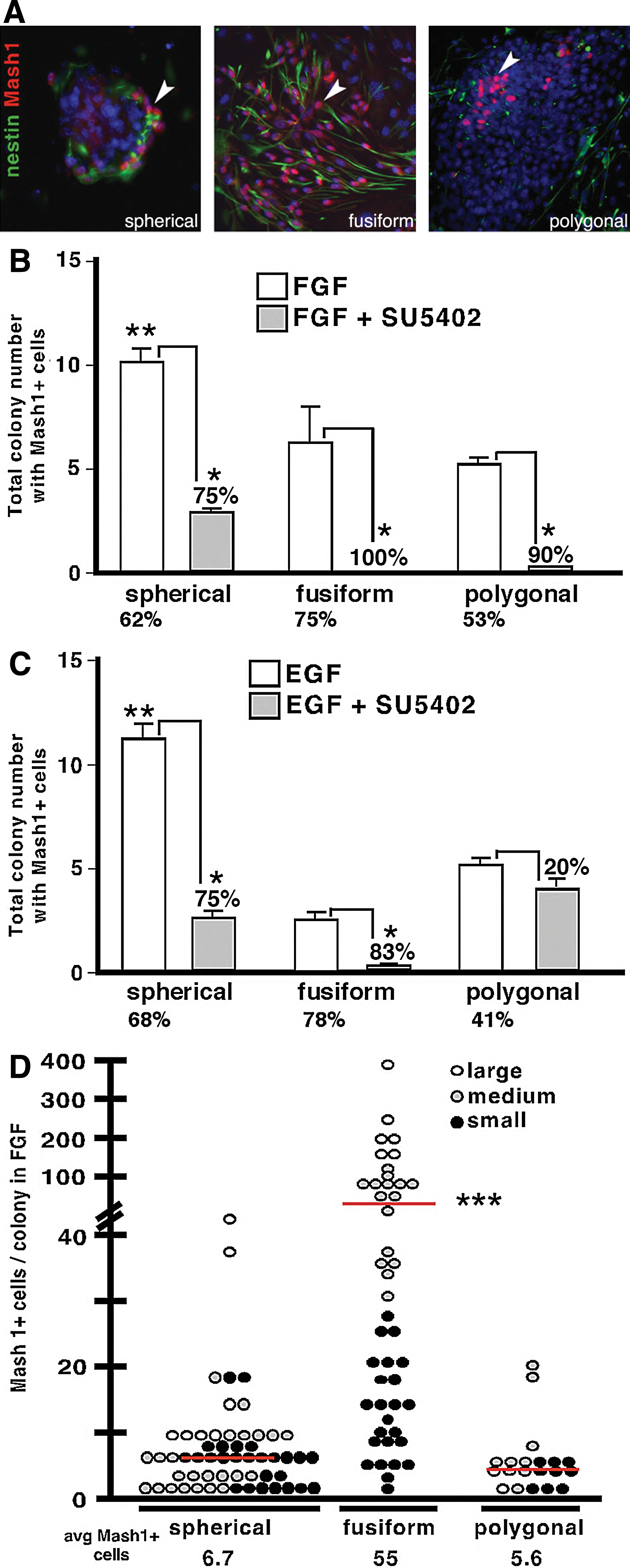
FGF signaling increases the production of neurogenic colonies and MASH1+ neuronal progenitors. E13.5 OE was clonally cultured in a serum-free medium supplemented with FGF2 or EGF, with or without FGF inhibition, for 10 days before assaying the total colony number containing at least one MASH1+ cell.
Given that FGF can alter the number of colonies containing MASH1+ cells, we tested if FGF blockade also decreased the frequency of MASH1+ cells per colony, and thereby limits the neurogenic potential of any colony subtype. The average number of total detectable MASH1+ cells/colony was highest in cultures treated with FGF2 (24.9±4.9, P<0.001, n=3) compared to EGF (13.8±1.8), and reduced by blocking FGF signaling, 76% (P<0.001, n=3) and 62% (P<0.01, n=3) in FGF2 and EGF, respectively.
Remarkably, fusiform colonies in the presence of FGF contained almost 10 times more MASH1+ cells (55 cells/colony; P<0.001, n=3), than either spherical (6.7 cells/colony) or polygonal colonies (5.6 cells/colony; Fig. 4D). In fusiform colonies, the MASH1+ progenitor number increased in direct proportion to increasing colony size, a trend not seen with the other colony subtypes (Fig. 4D). These results show that the highest neurogenic potential, as measured by MASH1+ output, is achieved via FGF stimulation of fusiform colonies.
FGF regulation and phenotype of E13.5 OE neurospheres
In multiple tissues, extracellular matrix (ECM) molecules control adhesion and contribute to a niche microenvironment that regulates progenitor proliferation, self-renewal, and differentiation [37]. This is the case in the postnatal OE, where laminin and collagen in the OE basement membrane differentially regulate mature progenitors [7]. Here, we tested if collagen or laminin substrates could similarly enhance E13.5 OE colony production in FGF2-treated cultures. Surprisingly, alone neither collagen nor laminin produced adherent or semiadherent colonies, but instead appeared antiadhesive and promoted the production of nonadherent spheres (Fig. 5A), similar to CNS neurospheres [30]. The production of embryonic OE neurospheres was also stimulated by FGF2 in a dose-dependent manner, similar to semiadherent OE colonies. We found significant increases (P<0.001, n=3) in the number of OE neurospheres at high (20 and 50 ng/mL), compared to low FGF2 concentrations (0.2 ng/mL; Fig. 5B). Blocking FGF signaling in FGF2-treated cultures with either SU5402 (data not shown) or function-blocking antibodies significantly reduced the number of neurospheres produced (P<0.01 to P<0.001, n=3), and depended upon the antibody concentration (Fig. 5C). Compared to FGF2, no significant change in the OE neurosphere number was seen with either vehicle (not shown) or antibody isotype controls (Fig. 5C), providing both gain-of-function and loss-of-function evidence that FGF2 can also regulate the production of embryonic nonadherent OE neurospheres.
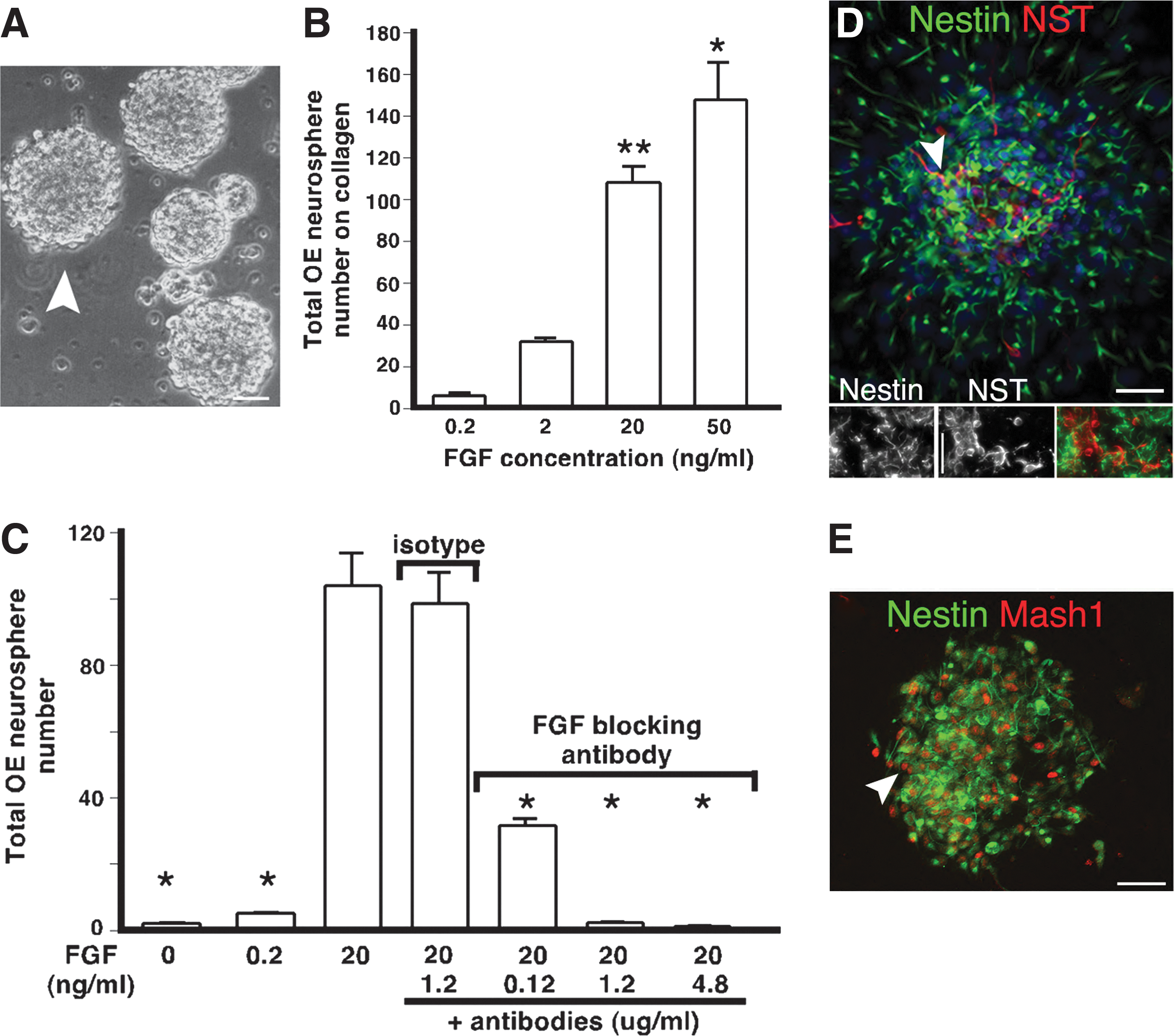
FGF regulation and phenotyping of E13.5 OE neurospheres. A single-cell suspension of E13.5 OE was cultured in an FGF2-treated serum-free medium on collagen substrate and grown for 10 days before counting the total number of nonadherent OE neurospheres or performing immunocytochemistry.
We tested the antigenic profile of individual OE neurospheres using immunocytochemistry for a combination of neuronal, glial, and progenitor markers, to test their neurogenic potential. OE neurospheres are highly enriched for NST+ neurons, MASH1+ neuronal progenitors, and nestin-expressing progenitors or glia (Fig. 5D, E), indicating that OE neurospheres, like semiadherent spherical colonies, have the capacity to support neurogenesis and gliogenesis.
FGF2 enhances the production of Mash1+ progenitors and nestin-expressing cells
Since embryonic OE neurospheres appeared to be highly enriched for neurons and neuronal progenitors, we tested if FGF signaling was an essential component of OE neurogenesis. E13.5 OE was first plated onto a collagen substrate in a medium containing FGF2 (as in Fig. 5), and colonies allowed to develop. About 50% of the FGF2-treated cultures were treated at day 7 with the inhibitor SU5402, to block ongoing FGF signaling, and BrdU, to identify cells undergoing DNA synthesis. The next day (day 8), after brief adhesion to collagen plus laminin, spheres were fixed, and we used immunofluorescence to test if we had altered the proliferation of cells that were nestin+ or MASH1+ and had recently divided (BrdU+). Of the total cells in OE neurospheres, 99% are nestin+ (of total1542 cells), 61% are dividing (BrdU+, of total1553 cells), and 42% are MASH1+ (of 1501 cells total; Fig. 6A and data not shown). The majority of MASH1+ cells (99%) are also BrdU+. When FGF signaling is blocked for just 24 h, the percentage of BrdU+ cells drops to encompass only 54% of the nestin+ cells (P<0.001, n=3), and 43% of the MASH1+ cells (P<0.01, n=3; Fig. 6C). We further tested the effects of FGF inhibition on new ORN production by quantifying at day 10 the percentage of newly generated NST+ ORNs that were BrdU+ (from labeling on day 7). In exogenous FGF2, 35%±2.5% of NST+ ORNs are BrdU+ by day 10, which decreases by 50% with FGF signal blockade (Fig. 6B, C; >335 NST+ cells counted for each). These results show that FGF signaling stimulates the proliferation of MASH1+ neuronal progenitors, which leads to the production of new ORNs. FGF2 also regulates proliferation of nestin+ cells, which appear to represent both neuroglial progenitors and immature (more rapidly dividing) OECs.
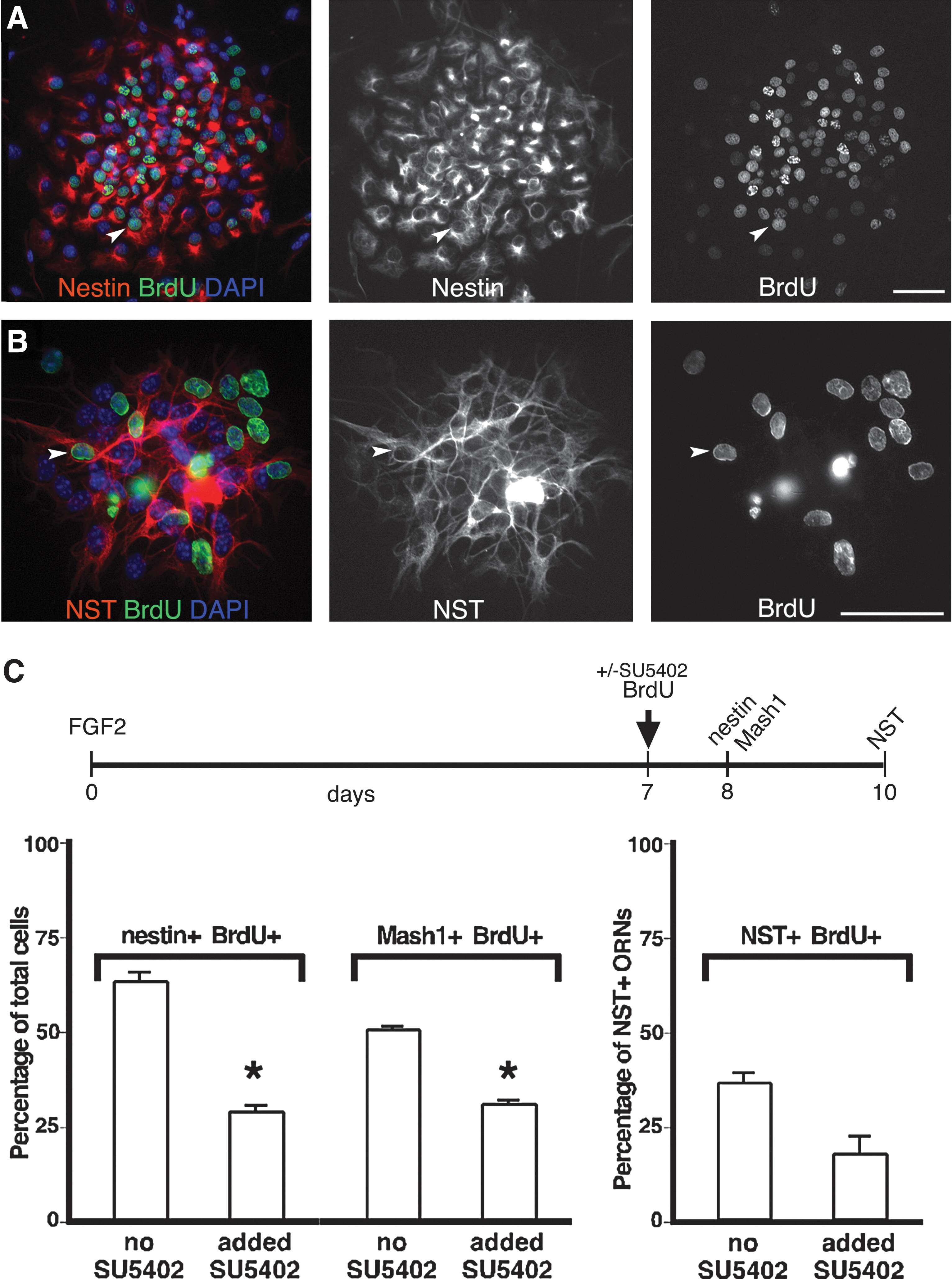
FGF signaling enhances olfactory neurogenesis by increasing nestin+ and MASH1+ progenitor proliferation. E13.5 OE was cultured at clonal density in a FGF2-treated serum-free medium on a collagen substrate and grown for 7 days before adding BrdU±SU5402. After 1 or 3 more days, nonadherent OE neurospheres were transferred to collagen/laminin substrates and fixed for immunocytochemistry.
Discussion
Although FGF signaling has been implicated in the regulation of neurogenesis throughout the nervous system and in OE explants, here we have tested specifically how it impacts the production of embryonic OE radial glial-like nestin+ and MASH1+ progenitors, and ultimately OE neurogenesis. By mapping embryonic expression of its receptors in different OE subcompartments, we first established different cellular sites of FGF2 action in vivo. By using novel, clonal in vitro assays together with gain- and loss-of-function studies, we were able to more tightly define FGF action to distinct cells and stages of the ORN lineage than is possible in explant assays. These data reveal the existence of phenotypically distinct OE precursors that FGF signaling has 3 major sites of action in the embryonic OE, and allows us to establish their differential dependence on FGF and ECM for neurogenesis. FGF appears to first act on colony-initiating cells to increase the production of phenotypically different subsets of neuroglial progenitors, and secondly acts on MASH1-expressing progenitors to enhance their mitosis and production of immature ORNs. Finally, FGF also regulates the mitosis and production of presumptive OECs from glial precursors. Our studies reveal—for the first time—different neuronal and glial progenitor targets for FGF action that can support long-lived olfactory neurogenesis during extended culture.
FGFRs are expressed in different progenitor phenotypes in the embryonic OE
Multiple FGF receptors have been detected in the embryonic OE and/or placode, but their cell-specific expression has not been elucidated [31,32]. Here, we show that both FGFR1 and FGFR2 are expressed in the embryonic OE in different subtypes of proliferating neurogenic cells, including radial glial-like nestin+ progenitors and MASH1+ neuronal progenitors, and with differentiation into immature neurons are progressively lost (Figs. 1 and 2). FGFR2, but not FGFR1, is readily detected in developing OECs (Fig. 2), which proliferate in response to FGFs [38].
FGF2 can enhance the proliferation of immediate neuronal precursors in the embryonic OE explants [32]. However, FGFRs are expressed in a number of different cells in the embryonic OE (Figs. 1 and 2), revealing a number of potentially different cellular sites of action for FGFs. Although neurosphere assays have been useful in testing FGF dependence of embryonic and adult CNS progenitors, these same conditions do not produce classic neurospheres from the developing OE. To overcome this, we developed a unique assay system, to clonally assay different subtypes of colony-forming progenitors from embryonic OE to test where in the ORN lineage, FGF may most exert its action. By combining both gain and loss of FGF action, we were able to test if FGF2 may also act on a more apical, primitive OE progenitor, and if EGF can also independently regulate embryonic olfactory neurogenesis in different colony subtypes.
FGF stimulates clonal colony formation and expansion from phenotypically distinct embryonic OE progenitors
Subpopulations of dividing embryonic OE nestin+ radial glial-like cells appear to coexpress FGFR1 and FGFR2 in vivo (Figs. 1 and 2) and are thus candidate founder cells for at least some OE embryonic colonies. Indeed, embryonic OE semiadherent colonies enriched in FGFR1 and FGFR2 are enriched in nestin+ cells (Figs. 3 –5), where elongated, bipolar mitotic nestin+ cells, lacking expression of MASH1, can be detected as early as 2.5 h after plating [15]. FGF2 differentially promotes the production and expansion of 3 different colony phenotypes—fusiform (primarily neurogenic), polygonal (primarily gliogenic), and spherical (more multipotent)—which we postulate are likely to be founded by distinct classes of progenitor present in the same time window of OE development. Exogenous FGF2 enhances the production of MASH1+ neuronal progenitors within all semiadherent colonies, but neurogenic fusiform colonies produce 10 times more MASH1+ neuronal progenitors in response to FGF than their spherical or polygonal counterparts (Fig. 4). EGF can also enhance the production of MASH1+ progenitors, but less significantly than FGF2. To ensure that we could effectively and specifically block FGF action, we used 2 independent approaches—the SU5402 inhibitor and FGF function-blocking antibodies [39]. SU5402 blocks the phosphorylation of all 4 FGFRs without affecting the phosphorylation of EGFRs [35,36], and can block FGF signaling in postnatal olfactory cells [40,41]. When added at initial culture, both treatments profoundly inhibited the ability of OE neurogenic progenitors to adhere and proliferate, reducing the initiation of fusiform and spherical colonies by 99% (Figs. 3 and 5).
How is FGF acting on different OE progenitors?
FGF2 thus appears to regulate the initiation of all semiadherent colonies (and OE neurospheres, Fig. 6), where FGF inhibition alters number, size, and subtype of colony, and the resulting cell phenotypes produced, irregardless of the presence of EGF (Figs. 3 –6). Blocking FGF signaling in either FGF2- or EGF-treated cultures also significantly reduces the production of MASH1+ cells, where fusiform colonies are the most FGF-sensitive neurogenic colony subtype, becoming completely inhibited in their ability to produce MASH1+ progenitors (Fig. 4). In this case, FGF signaling could synergize with bone morphogenetic protein (BMP) inhibition to initially stabilize MASH1 in ORN precursors, thus deterring MASH1 proteolysis and opposing MASH1 degradation by BMPs [21]. FGF2 could also regulate the subcellular location of the transcription factor FoxG1 [42]. Neural progenitor cell lines from the olfactory placode retain FoxG1 in their nucleus in the absence of FGF, but in the cortex, FGF2 stimulates FoxG1 to translocate to the cytoplasm, stimulating neuronal differentiation [42]. If blocked by FGF signal inhibition, neuronal precursors accumulate and block further neural differentiation, as we see here in the embryonic fusiform colonies.
Polygonal colonies are primarily EGF responsive, contain the lowest number of MASH1+ progenitors and only sporadic neurons, and are the least affected by FGF signal inhibition, (Figs. 3 and 4). They are also most similar in morphology and growth factor responsiveness to P5 OE ICAM-1+-adherent colonies, which are predominantly gliogenic (expressing S100β or GFAP) [7]. Spherical colonies appear to be equally neuro- and gliogenic. Their frequency is similar in either EGF or FGF2 (Fig. 3), and they contain glia, neurons [15], and MASH1+ neuronal progenitors (Fig. 4). FGF2 appears to favor the neurogenic, rather than gliogenic, potential of cells within spherical colonies, although we cannot rule out that a number of the nestin+ dividing cells in spherical colonies might also be OEC precursors that express high levels of FGFR2. Given that we have never produced spherical colonies similar to these from postnatal OE (even in control experiments performed in parallel), spherical colonies suggest the existence of more primitive embryonic bipotent OE progenitors that may not persist into the adult [11].
FGF signal blockers also attenuated the colony number and phenotype in EGF alone. Interestingly, unlike either spherical or fusiform colonies, polygonal colonies were virtually unaffected by FGF signal blockade, indicating that some colonies are more highly supported by EGF. This suggests 2 potential independent mechanisms of EGF: EGF could induce paracrine FGF production in vitro, as it does in the CNS, which enhances neurogenesis [26,43]. Second, EGF may act independently to support survival of distinct progenitors, since even after blocking FGF signaling, colonies are still formed, even if their expansion and differentiation are impaired. Paracrine regulation of neurogenesis and gliogenesis by factors other than FGFs is also likely, since a conditioned medium derived from 10-day-old FGF2-supplemented semiadherent cultures can support colony initiation from fresh clonal cultures, even after they have been freshly treated with antibodies to block FGF signaling (data not shown). Candidate paracrine factors include members of the TGF-β superfamily, since they and their receptors are expressed in the developing OE and in embryonic OE explants, and BMPs have been shown to modulate neuron and neuronal precursor numbers [20,21,23]. Enrichment for colonies containing neurons has also been achieved with a conditioned medium derived from embryonic OE stroma [20,44]. Differential responsiveness to external signals may allow different progenitors to respond to diverse biological needs during both development and repair, where fusiform-initiating progenitors (driven by FGF signaling) could be utilized for rapid production of many neurons, whereas more apical, multipotent progenitors would be used to initiate the development or restructuring of newborn (or in the adult, highly damaged) areas of OE. Together, these results suggest that FGF (and EGF) acts at different developmental stages on multiple progenitors within the embryonic OE, that these regulatory mechanisms may be shared between OE and CNS progenitors, and that the embryonic OE produces paracrine factors that may also support neurogenesis elsewhere in the CNS.
Matrix regulation of OE neurogenesis and gliogenesis
Perhaps, one of the most intriguing findings here was of the role of a matrix in directing the phenotype of embryonic colonies formed. Although a mixed collagen matrix selected for colonies with distinct physical and biological phenotypes, nonadherent OE neurospheres that cannot be individually distinguished phenotypically were consistently produced by plating cells on collagen or laminin alone. Since different ECM components can regulate stem/progenitor proliferation, self-renewal, and differentiation in neural and non-neural tissues [45 –47], the differential response of embryonic OE progenitors to mixed-versus-single substrates in vitro may reflect subtle microenvironments that regulate proliferation and migration as the olfactory system develops in vivo [48]. The majority of OE neurospheres, like semi-adherent colonies, contain proliferating nestin+ cells and MASH1+ progenitors, and immature neurons, all of which are increased in abundance by FGF2 (Figs. 5 and 6). This suggests that OE neurospheres may be founded by the progenitor that produces semiadherent spherical colonies, whereas a single ECM may not support the formation and expansion of embryonic polygonal and fusiform colonies. Taken together, the in vivo and in vitro data suggest that the most multipotent spheres/colonies may derive from FGFR-expressing nestin+ progenitors [15] that require both collagen and laminin to adhere and survive, but can proliferate independent of adhesion to substrate. Both semiadherent spherical colonies and OE neurospheres produce neurons and glia, but at very different frequencies to fusiform (almost exclusively neurogenic) and polygonal (almost exclusively gliogenic). Precursors that initiate different colony/sphere types may represent lineally related progenitors at different states of induction or commitment, where nonadherent OE neurospheres are less committed to differentiation than semiadherent colony precursors because of their increased frequency, greater number of MASH1+ cells per sphere (Fig. 5), and enhanced self-renewal capacity (data not shown).
Since most cells in OE neurospheres after 8–10 days were nestin+, and all MASH1+ progenitors were dividing (BrdU+), FGF2 likely acts directly on both nestin+ cells (glial precursors and neuroglial progenitors) and MASH1+ progenitors to increase their proliferation, leading to the production of more NST+ neurons (Fig. 6). FGF2 does not appear to act by changing the progenitor cell fate, since the proportion of dividing nestin+ and MASH1+ progenitors at 8 days compared to ORN output at 10 days is similar (56%–67%), even with FGF signal blockade. Since the majority of cells in OE neurospheres express nestin, of which a subpopulation express FGFR1 or FGFR2, and 42% of them also express Mash-1, we provide compelling additional evidence for the identification of an FGF-responsive nestin+ apical neuronal progenitor in the ORN lineage.
We propose the following model for the regulation of embryonic OE neurogenesis by FGF signaling (Fig. 7). FGF regulation of neurogenesis occurs whereby OE stem cells can either self-renew or give rise to FGF-responsive nestin+ neuronal progenitors lacking MASH1 expression, similar to those in the embryonic CNS ventricular zones [15,49]. As a subpopulation of nestin+ progenitors becomes committed to the ORN lineage, they induce Mash1 and form MASH1+ neuronal progenitors. Availability of FGF regulates the Mash1+ progenitor frequency and the mitotic index, and ensures that Mash1 expression is further downregulated before the production of immature ORNs.

Model of FGF2 regulation of neurogenesis via nestin+ and Mash1+ progenitors. Cells irreversibly pass through sequential progenitor stages, while losing their self-renewal, proliferative, and cell potential. Self-renewing (*) stem cells divide and give rise to a subpopulation of nestin+ neuronal progenitors that precede and develop into Mash1+ neuronal progenitors. Mash1+ progenitors initially express, but later in olfactory receptor neuron (ORN) development downregulate nestin, before becoming immediate neuronal precursors and finally postmitotic NST+ ORNs. In addition to acting on presumptive stem cells and immediate neuronal precursors, the present study shows that FGF2 acts to enhance neurogenesis by regulating the proliferation of nestin+ and MASH1+ progenitors and survival of newly generated neurons, leading to increased ORN production. EGF appears to act on more apical progenitors, and synergizes with FGF to enhance overall output of ORNs (and OECs).
Summary
In summary, here we developed in vitro assays that allowed us to test the ability of FGF2 to regulate production of olfactory neurons (and glia) from colonies founded by single-embryonic progenitors. FGF signaling appears to regulate embryonic OE neurogenesis not just by enhancing the number of Mash1-expressing precursors, but also by acting on colony-initiating cells, and that these progenitors are distinct from postnatal olfactory progenitors in their response to both growth factors and ECM. Our studies reveal for the first time multiple actions for FGF2 and ECM in OE development and novel progenitor targets for FGF action within the peripheral olfactory system that support long-lived neurogenesis in vitro.
Footnotes
Acknowledgments
We would like to thank Jane Johnson (University of Texas) for sharing the Mash1 antibody and Nicole Janzen for mouse colony maintenance. We are grateful for funding contributed by the Heart and Stroke Foundation of Canada, the Canadian Stroke Network and the Michael Smith Foundation for Health Research (B.M.), and Canadian Institutes of Health Research and Jack Brown Family Foundation (J.R.).
Author Disclosure Statement
The authors indicate no potential conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.