
Abstract
MicroRNAs are a class of short noncoding RNAs that are involved in various biological processes, including differentiation. MicroRNA-140 (miR-140) has been identified as a cartilage-specific microRNA with several targets involved in cartilage development and homeostasis. The aim of this study was to investigate the expression of miR-140 during chondrogenic differentiation of equine cord blood-derived mesenchymal stromal cells (eCB-MSCs). We demonstrate both that miR-140 is highly expressed in normal equine articular cartilage and that eCB-MSCs express significantly higher levels of this microRNA after 14 days of chondrogenic differentiation. Furthermore, miR-140 expression closely paralleled that of the cartilage-specific transcription factor Sox9, suggesting that miR-140 may be under the transcriptional regulation of Sox9 in these cells. The expression patterns of miR-140 targets the chemokine (CXC motif) ligand 12 (CXCL12), A disintegrin and metalloproteinase with thrombosponin motifs (ADAMTS)-5 and insulin growth factor binding protein 5 (IGFBP5) were also determined; however, only CXCL12 and ADAMTS-5 were repressed while miR-140 expression was upregulated. Together, these studies suggest that miR-140 is an important regulator of cartilage development and homeostasis in eCB-MSCs that may act, in part, through the regulation of CXCL12 and ADAMTS-5.
Introduction
MicroRNAs (miRNAs) are a class of endogenous noncoding RNAs, ∼20–25 nucleotides in length. These potent regulators of gene expression are thought to be involved in the regulation of one third of all mammalian genes [9]. They function at the post-transcriptional level by binding to complementary sequences in the 3′-untranslated region (3′-UTR) of target mRNAs and inhibiting their expression translation and/or facilitating their decay. miRNAs have profound effects on the cellular phenotype and biological function in many different tissues and numerous studies have shown that they are important regulators of diverse biological processes, such as differentiation, development, and tumorigenesis [10 –12].
During miRNA biogenesis, long primary transcripts (pri-miRNAs) are synthesized by RNA polymerase II, and then processed into small hairpin structures known as premiRNAs by the enzymes, Drosha and DGCR8. They are then exported into the cytoplasm where the RNAse III, Dicer, cleaves the loop region of the hairpin to generate a short RNA duplex. One strand is degraded leaving a mature single-stranded miRNA molecule that becomes incorporated into the RNA-induced silencing complex [13] and binds target mRNAs to either block translation or induce transcript degradation.
Evidence for the crucial role of miRNAs in cartilage development was provided by a study in which Dicer, an endoribonuclease required for the production of mature miRNA, was knocked out in the cartilage of mice. This resulted in a smaller proliferating pool of chondrocytes and accelerated hypertrophic differentiation, causing severe skeletal defects [14]. Findings by Wienholds et al. demonstrating that most conserved vertebrate miRNAs are expressed in a tissue-specific manner in zebrafish embryos and that microRNA-140 (miR-140) is only expressed in cartilaginous tissue, suggested an important role for this miRNA species in cartilage development [15]. Subsequent studies confirmed the cartilage-specific nature of miR-140 in mice [16] and humans [17].
Several miR-140 targets have been identified; the histone deacetylase 4 (HDAC4) [16], chemokine (CXC motif) ligand 12 (CXCL12) [18], insulin growth factor binding protein 5 (IGFBP5) [19], and a disintegrin and metalloproteinase with thrombosponin motifs (ADAMTS)-5 [20], Smad3 [21], Dnpep [22], and Sp1 [23]. Additionally, miR-140 was found to be directly induced by Sox9, the master transcriptional regulator of chondrogenesis. Furthermore, it is coexpressed with the host gene from which it is intronically transcribed, Wwp2-C, an E3 ubiquitin protein ligase [23].
Although miR-140 is the most frequently studied miRNA in cartilage biology and pathology, studies have not been described in equine cells. In this study, we demonstrate that miR-140 is highly expressed in normal equine cartilage and that expression increases after 14 days of chondrogenic differentiation of eCB-MSCs. Expression was directly correlated with the transcriptional regulator Sox9, and previously identified targets ADAMTS-5 and CXCL12 showed expression patterns that suggest possible regulation by miR-140. These findings provide further insight into the molecular mechanisms of chondrogenic differentiation in eCB-MSCs.
Materials and Methods
The study design is illustrated in Fig. 1. Briefly, a preliminary screening of 11 eCB-MSCs was performed by differentiating the cells for 2 weeks then staining with toluidine blue. Strong staining was associated with high CP; thus, strongly stained differentiated eCB-MSCs were selected (N=5) for further analysis. Chondrogenesis was induced for 7, 14, and 21 days before histology, immunostaining and quantitative real-time reverse transcription polymerase chain reaction (qPCR) were performed.
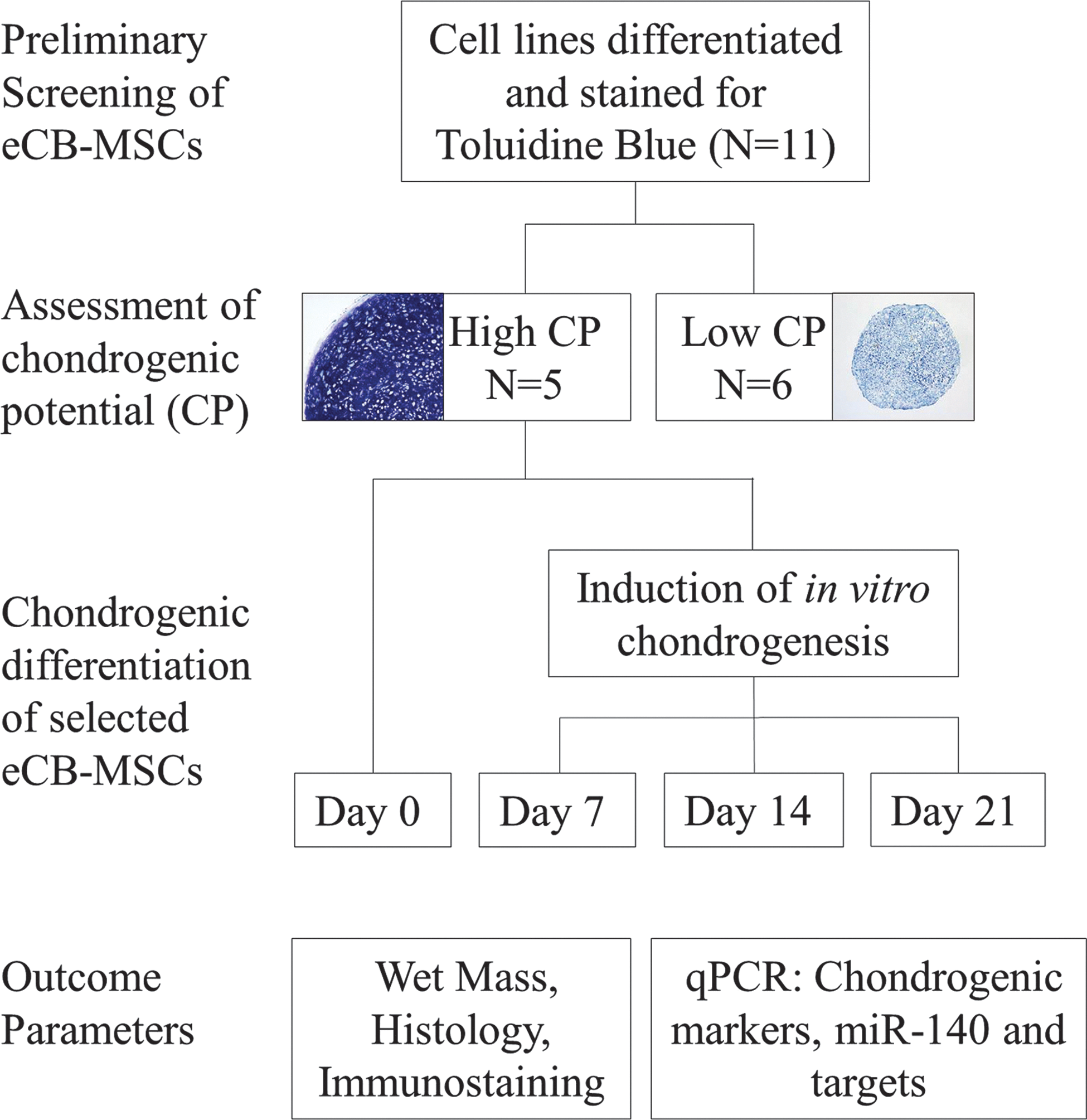
Study design. All equine cord blood-derived mesenchymal stromal cell (eCB-MSC) lines were screened for chondrogenic potential (CP) before the study by differentiating pellets for 2 weeks, then staining with toluidine blue. Strongly stained pellets with hyaline-like morphology were grouped as high CP eCB-MSCs, while weakly stained pellets were grouped as low CP eCB-MSCs. High CP eCB-MSCs were subjected to chondrogenic differentiation for 7, 14, or 21 days before assessment of outcome parameters. Color images available online at
eCB-MSC isolation and culture
The isolation of equine MSCs from umbilical cord blood was performed as previously described [24]. Briefly, the nucleated cell fraction was isolated from 11different cord blood samples using the PrepaCyte®-WBC medium (BioE, Inc.) according to the manufacturer's instructions. The cells were suspended in an isolation medium [the low-glucose Dulbecco's modified Eagle's medium (DMEM; Lonza), 30% FBS (Invitrogen), dexamethasone (10−7 M; Sigma), penicillin (100 IU/mL; Invitrogen), streptomycin (0.1 mg/mL; Invitrogen), and L-glutamine (2 mM; Sigma)], seeded in polystyrene culture flasks, and incubated at 38.5°C with 5% CO2 in a humidified incubator. When cells reached 70%–80% confluence, they were detached using a 0.04% trypsin/0.03% EDTA solution and reseeded at 5,000 cells/cm2. For cryopreservation, trypsinized cells were washed, resuspended in 10% dimethyl sulfoxide (Sigma), stored overnight at −80°C, and transferred to liquid nitrogen for long-term storage. Following preliminary screening, 5 eCB-MSC lines were selected, thawed, and expanded; experiments were performed in passages 5–6.
Chondrogenic differentiation
High-density micromass pellet cultures were prepared by centrifuging 2.5×105 cells/pellets at 150 g in 96-well V-bottom polypropylene plates (Phenix). 20 pellets were prepared for each cell line for each time point and cultured at 38.5°C in a hypoxic environment of 5% O2 for 7, 14, or 21 days in chondrogenic differentiation media containing the DMEM-HG, 200 mM Glutamax-I, 100 mM Sodium pyruvate, 1× ABAM (Invitrogen), 0.1 mM dexamethasone, 100 μg/mL ascorbic acid-2 phosphate, 40 μg/mL proline (Sigma-Aldrich), 1xITS (BD Biosciences), and 10 ng/mL transforming growth factor-β3 (TGF-β3; R&D Systems). The medium was changed every 2–3 days.
Chondrocyte isolation
Articular cartilage was collected from the fetlock joints of 3 horses without evidence of joint or systemic disease immediately after euthanasia. Tissue was minced with a scalpel blade and digested with 1.5 mg/mL collagenase type II (Sigma-Aldrich) overnight at 37°C. Digested samples were strained and isolated chondrocytes were snap frozen in the mirVanalysis/binding buffer (Ambion) and stored at −80°C until RNA isolation.
Histology and immunohistochemistry
One representative pellet from each cell line was selected (N=5), fixed in 10% formalin, and embedded in paraffin. Five micrometer sections were prepared and stained with hematoxylin and eosin or toluidine blue. For collagen type I and type II immunostaining, paraffin-embedded sections were rehydrated and digested with 20 μg/mL proteinase K and 1600 U/mL hyaluronidase, blocked with 3% bovine serum. and incubated with an antibody reactive to collagen type I (Calbiochem) or collagen type II (Developmental Studies Hybridoma Bank) overnight at 4°C. Subsequently, samples were incubated with the HRP-conjugated anti-mouse secondary antibody (DAKO) and immunoreactivity was detected using diaminobenzidine chromagen (DAKO) with hematoxylin counterstain. QCapture software was used for imaging.
RNA extraction, reverse transcription, qPCR
Total RNA, including small RNAs, was isolated from equine articular chondrocytes (N=3), undifferentiated eCB-MSCs (N=5), and pellet cultures (N=5 with 5 pellets per cell line) using the mirVana miRNA isolation kit according to the manufacturer's protocol (Ambion). RNA concentration and purity were determined using the Nanodrop ND-1000 spectrophotometer. To determine miRNA expression, 100 ng of RNA was reverse transcribed using the TaqMan miRNA reverse transcription kit with the appropriate miRNA-specific stem-loop primer, mmu-miR-140. This primer, while designed for mouse, is identical to the homologous Equus caballus sequence as confirmed by the BLASTN sequence alignment of the probe on the equine genome. qPCR was then performed using the TaqMan PCR Master Mix and TaqMan microRNA assay (Applied Biosystems). To assess mRNA expression, complementary DNA was synthesized from total RNA using SuperScript II and oligo (dT)18 primers (Invitrogen) and qPCR was performed using Roche LightCycler 480, Fast Start DNA Master SYBR Green I (Roche Diagnostics), and the equine-specific primers found in Table 1. All reactions were run in triplicate. Crossing point (Cp) values for miR-140 were normalized to U6 snRNA, while Cp values for mRNAs were normalized to GAPDH. Relative gene/miRNA expression was determined using the delta–delta Ct method.
Primers were designed by Dr. Lise C. Berg.
Primers were designed by Laurence Tessier.
Statistical analysis
GraphPad Prism 5 Software was used to perform statistical analyses. One-way analysis of variance with the Bonferroni post-test was applied to determine the differences between the gene expression levels and wet mass. The Student's unpaired t-test was used to compare the expression of miR-140 between MSCs and chondrocytes. Gene expression data are expressed as fold differences±SEM compared with undifferentiated eCB-MSCs. Significance was determined using raw data and assigned at P<0.05.
Results
Chondrogenic differentiation of eCB-MSCs
eCB-MSCs formed white, glistening pellets, an appearance consistent with hyaline cartilage (Fig. 2A). They increased in size and weight over 21 days of differentiation (Fig. 2B), with the average wet mass increasing from 0.56 mg on day 7 to 0.93 mg on day 21. Additionally, pellet sections were stained with hematoxylin and eosin to assess their morphology and with toluidine blue to determine the presence of proteoglycans (Fig. 2C). Histological examination clearly showed cartilage tissue development; cells with round morphology inside lacunae were visible at day 14 and became more defined at day 21. toluidine blue staining was visible as early as day 7 and showed a progressive increase in the proteoglycan content in the accumulated matrix at days 14 and 21. Immunostaining was also performed to assess the protein expression of collagens type I and II (Fig. 2C). As expected, the staining intensity for collagen type II increased from day 7 to day 21, while decreasing for collagen type I.
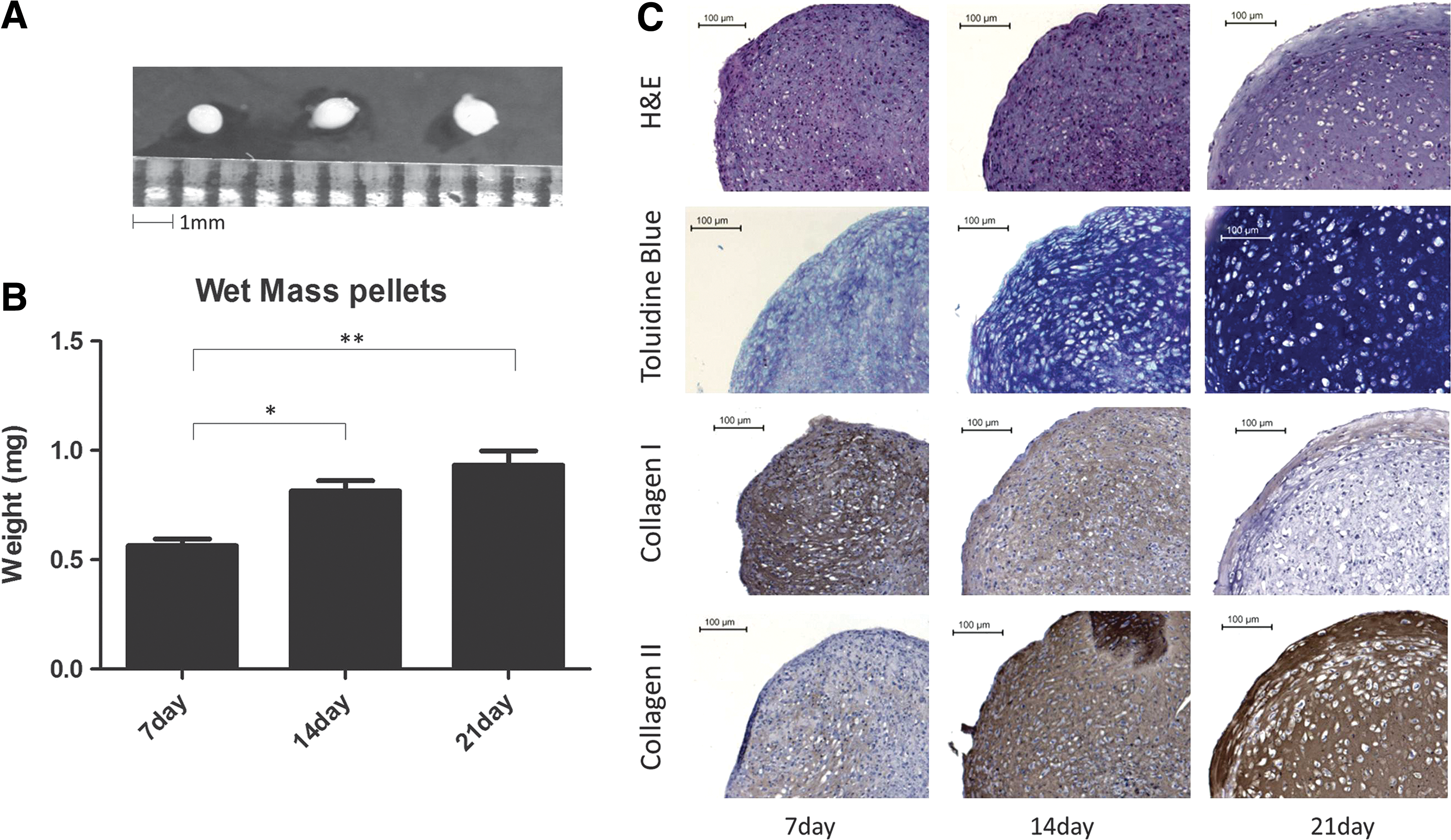
Chondrogenic differentiation of eCB-MSCs.
Chondrogenesis was also evaluated over the 3-week period through the assessment of gene expression for collagens I, II, and X and aggrecan using qPCR (Fig. 3). Expression levels of collagens I and II, and aggrecan increased significantly from day 0 to day 7, then began to decrease. The expression of aggrecan at 7 days of chondrogenesis was similar to articular chondrocytes, while expression levels of collagens I and II were similar to articular chondrocytes at 21 days. Collagen X expression peaked at day 14, then decreased at day 21, at which point the expression level was not significantly different from that of articular chondrocytes.
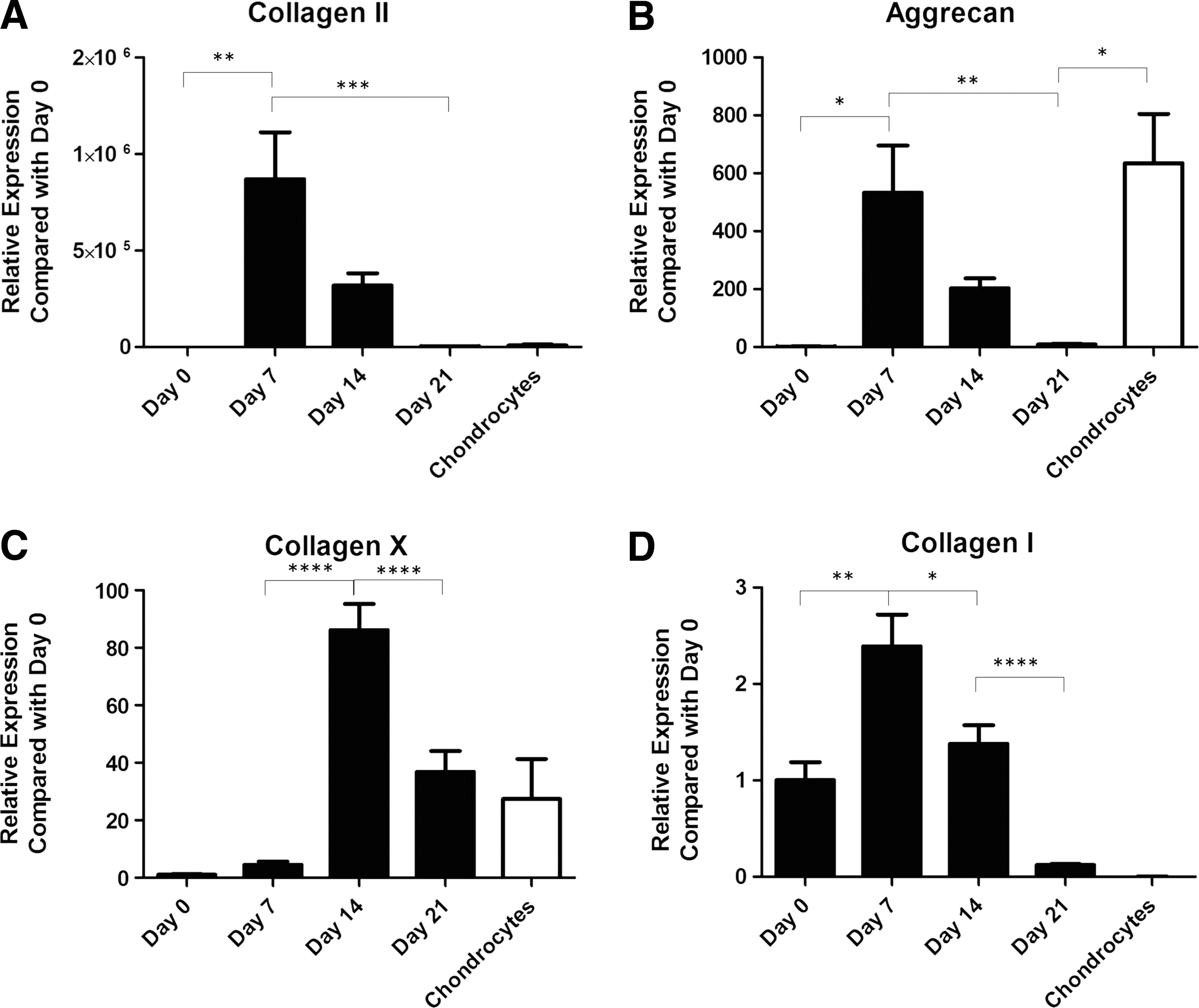
Gene expression of cartilage markers by eCB-MSC pellets cultured up to 21 days in chondrogenic media. mRNA levels were assessed by qPCR with GAPDH as a reference gene.
miR-140 expression in undifferentiated eCB-MSCs and equine articular chondrocytes
qPCR analysis using TaqMan miRNA assays revealed that the expression of miR-140 is almost 300-fold higher in equine chondrocytes compared to undifferentiated eCB-MSCs (Fig. 4).
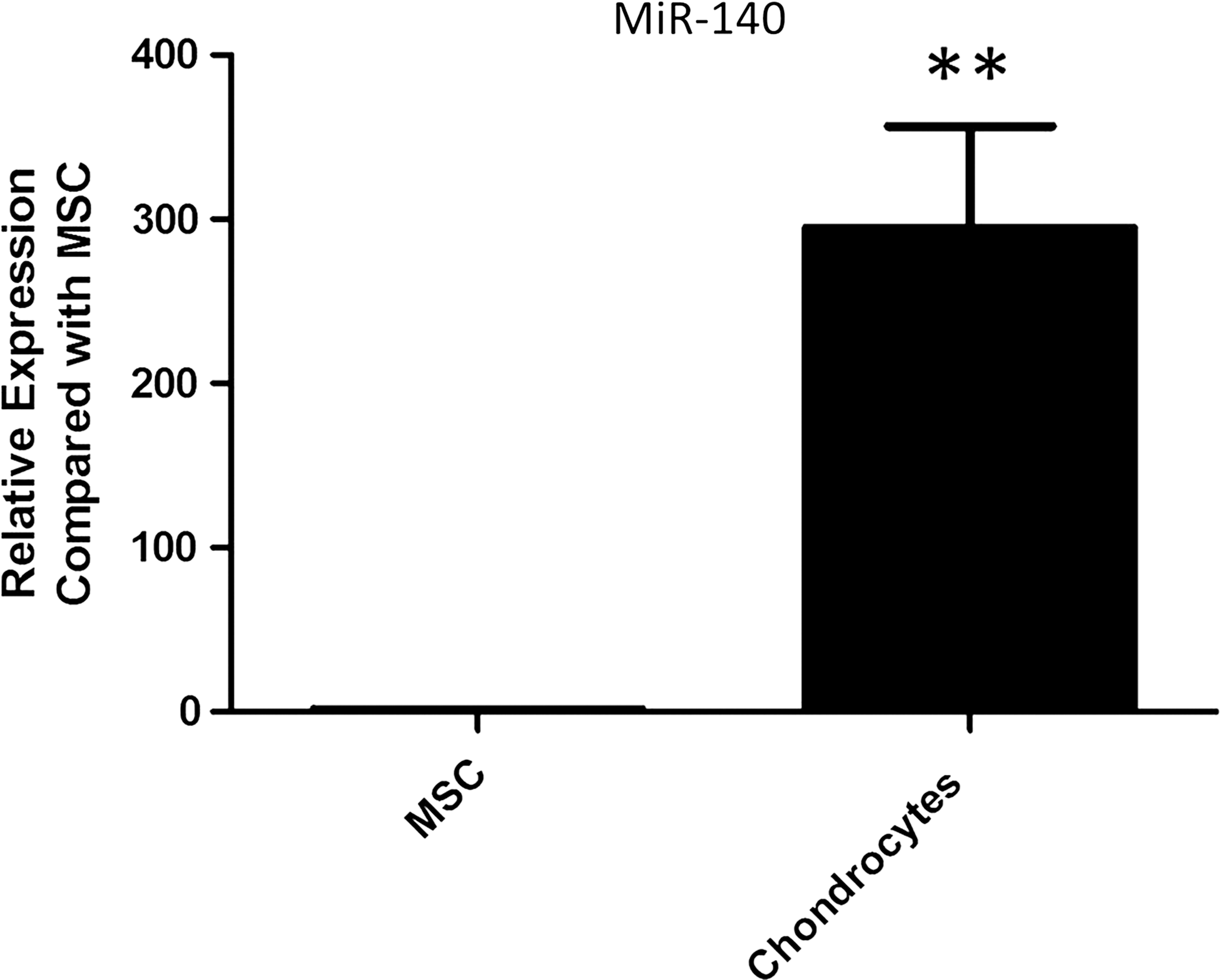
Expression of microRNA-140 (miR-140) in undifferentiated eCB-MSCs and articular chondrocytes. MiR-140 expression was significantly higher in chondrocytes (N=3) compared to eCB-MSCs (N=5). Data are shown as mean±SEM. **P<0.01.
Expression of miR-140, Sox9, and miR-140 target genes during chondrogenesis
During chondrogenic differentiation, miR-140 increased significantly from day 0–14, then decreased slightly, although not significantly, at day 21 (Fig. 5A). The expression of Sox9, the master transcriptional regulator of chondrogenesis, also followed this pattern (Fig. 5B). The expression patterns for previously identified miR-140 targets IGFBP5, CXCL12, ADAMTS-5, and HDAC4 [16, 18–20] were also examined. IGFBP5 expression levels increased from day 0–14, and then decreased at day 21 (Fig. 5C). In contrast, CXCL12 expression was highest at day 0, decreasing significantly by day 7, and remaining repressed throughout the differentiation period. CXCL12 expression appeared higher in the chondrocytes compared with the differentiating MSCs, although statistical significance was not achieved as there was considerable variability (Fig. 5D). A similar pattern was observed for ADAMTS-5, its expression decreased significantly from day 0–7 and remained low until day 21; however, this repression was not significant at day 14 and day 21 compared to day 0, likely due to interdonor variation Surprisingly, ADAMTS-5 mRNA expression appeared relatively high in normal articular chondrocytes, although significant variability was present (Fig. 5E). Expression of HDAC4, a corepressor of Runx2, was also examined, but mRNA levels in most samples were below the detection limit (data not shown). These results suggest that miR-140 may play an important role in the chondrogenic differentiation of eCB-MSCs through selective targeting of specific mRNAs, such as CXCL12 and ADAMTS-5.
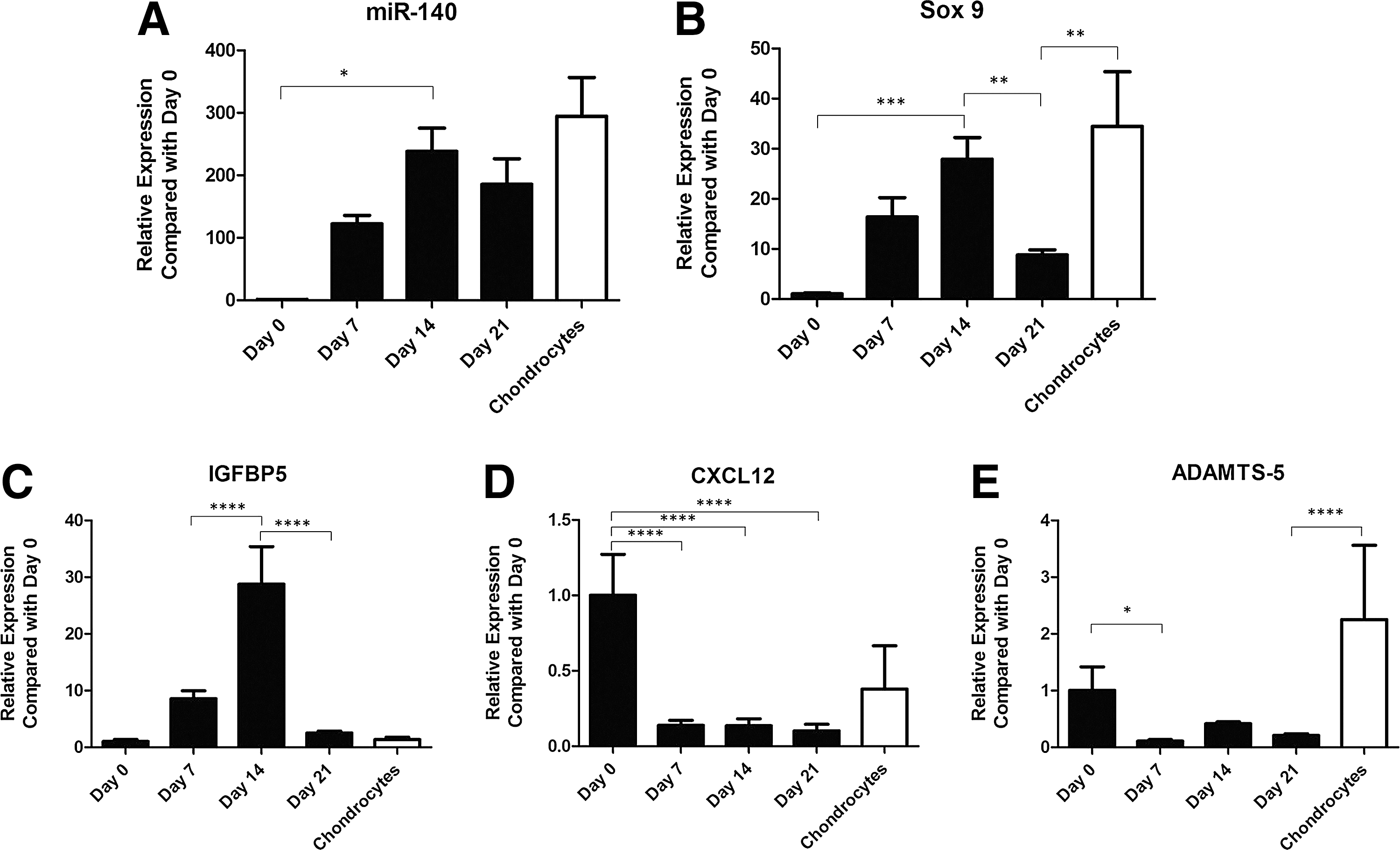
Expression of miR-140, Sox9 and previously identified targets insulin growth factor binding protein 5 (IGFBP5), chemokine (CXC motif) ligand 12 (CXCL12), and A disintegrin and metalloproteinase with thrombosponin motifs (ADAMTS)-5.
Discussion
During vertebrate development, chondroprogenitor cells undergo morphological changes and begin to express cartilage-specific genes, leading to chondrocyte maturation and ECM production [30]. Chondrogenesis is initiated by the recruitment and condensation of mesenchyme, a process which is regulated by cell–cell interactions. Similarly, in vitro chondrogenesis involves the aggregation of mesenchymal cells into high-density pellets [31]. TGF-β is one of the earliest genes expressed in in vivo mesenchymal condensations, whereas TGF-β is applied exogenously in vitro. Sox9 is a key transcriptional regulator in early chondrogenesis, with a crucial role in regulating cartilage formation by activating the expression cartilage-specific genes, including collagen type 2, aggrecan, and cartilage oligomeric matrix protein [32,33]. Sox9 is also expressed during chondrogenic differentiation of micromass pellets. As previously reported, eCB-MSCs can be successfully differentiated toward a chondrogenic fate, as indicated by a gradual increase in size and weight and by histological evaluation of morphology, proteoglycan content, and expression of chondrogenic markers [4,7,24]. Gene expression patterns for collagen II and aggrecan in this study showed an initial increase after 1 week, then began to decrease. This suggests that upon chondrogenic stimulation, the eCB-MSCs rapidly accumulate transcripts to produce ECM. Collagen II mRNA levels did not correlate with the protein abundance seen in the immunostaining. One possible explanation for this is that, as differentiation progresses and the ECM is established, collagen II mRNA levels decrease to baseline expression as the chondrocytes take on a maintenance role. However, the significantly lower expression of aggrecan compared to chondrocytes and the increased expression of collagen X indicates that the differentiation method does not perfectly recapitulate in vivo cartilage development.
Studies demonstrating the loss of the miRNA processing enzyme Dicer in the cartilage of mice revealed an overall role for miRNAs in chondrogenesis and skeletogenesis. The absence of mature miRNAs caused severe growth defects due to a decrease in proliferating chondrocytes and accelerated hypertrophy [14]. Many miRNAs with roles in cartilage-specific processes have since been identified. For instance, miR-199a* was identified as an inhibitor of early chondrogenic differentiation. While overexpression of miR-199a* in murine MSCs decreased the expression of early chondrogenic markers during BMP2-induced chondrogenesis, anti-miR-199a* increased their expression. This was mediated through Smad1 signaling, which was identified as a direct target of this miRNA through 3′-UTR luciferase reporter assay [34]. In a similar study, miR-145 was reported to directly repress Sox9 during TGF-β3-induced chondrogenic differentiation of murine MSCs [35]. MiR-140 has been the focus of many cartilage studies due to its specificity in this tissue. It is transcribed from an intron within the Wwp2-C gene and thought to be induced by Sox9 [23]. Identified targets include HDAC4, IGFBP5, ADAMTS-5, Cxcl12, Smad3, Dnpep, Sp1, and MMP-13 [16,18 –22,36]. The role of miRNAs in cartilage development and homeostasis has been reviewed elsewhere [37].
We demonstrate here that miR-140 is expressed in equine cells, specifically in chondrogenically differentiating eCB-MSCs and articular chondrocytes. This observation is in agreement with previous reports that miR-140 is cartilage specific [15] and highly expressed in human articular chondrocytes [17]. A significantly higher expression was observed in eCB-MSCs following 2 weeks of in vitro differentiation stimuli, compared to undifferentiated cells. The presence of miR-140 in undifferentiated eCB-MSCs suggests that this miRNA could be involved in the maintenance of CP. This is consistent with the detection of miR-140 in undifferentiated C3H10T1/2 cells that were subsequently differentiated into chondrocytes [18]. The expression pattern of miR-140 correlated with the expression of chondrogenic transcription factor Sox9. Yang and colleagues reported that suppression of Sox9 reduced miR-140 expression in ATDC5 cells and they utilized luciferase reporter assays to confirm the direct binding of Sox9 to the upstream region of miR-140 [23]. Our data are consistent with the potential for Sox9 to regulate miR-140 expression during eCB-MSC chondrogenic differentiation; however, additional studies are required to confirm this hypothesis.
miRNAs influence target gene expression by facilitating the decay of specific mRNA targets or by inhibiting translation of target genes. In this study, we also evaluated the expression of 3 previously identified miR-140 mRNA targets; IGFBP5, CXCL12, and ADAMTS-5. IGFBP5 is a protein that regulates the availability of IGF-1, an anabolic factor involved in matrix synthesis and chondrocyte survival [38]. IGFBP5 was reported to be directly targeted for mRNA degradation by miR-140 in human OA chondrocytes. Pre-miR-140 treatment caused a significant decrease in IGFBP5 mRNA expression, while knockdown caused a significant increase. These changes occurred early (24 h) and since a miR-140 seed sequence was predicted in the 3′-UTR of IGFBP5, a direct interaction between IGFBP5 and miR-140 was suggested; however, 3′-UTR luciferase reporter assays were not performed [19]. CXCL12, also known as the stromal-derived factor-1, functions by activating the G-protein-coupled receptor CXC chemokine receptor 4 (CXCR4). CXCL12 is a potent chemoattractant for CXCR4-expressing cells and may have a role in MSC migration [39], survival, and initiation of differentiation [40]. Using an overexpression/knockdown approach, CXCL12 was identified as 1 of 49 genes that were repressed by miR-140 overexpression and derepressed by miR-140 silencing. Northern blot analysis and 3′-UTR luciferase reporter assays validated CXCL12 as a miR-140 target [18]. ADAMTS-5 is a proteolytic enzyme that cleaves aggrecan and has been associated with the degeneration of osteoarthritic cartilage [41,42]. In a study by Miyaki et al., miR-140−/− mice and transgenic (TG) mice overexpressing miR-140 were generated. ADAMTS-5 expression was increased in the chondrocytes of miR-140−/− mice and reduced in the chondrocytes of miR-140 TG mice. Furthermore, ds-miR-140 decreased the expression of ADAMTS-5 in miR-140−/− mice, providing additional evidence that miR-140 targets this gene for degradation. Direct binding of miR-140 to ADAMTS-5 mRNA was demonstrated by luciferase reporter assay [20].
Surprisingly, the expression pattern of IGFBP5 correlated directly with miR-140 levels rather than an inversely, which would be expected if miR-140 facilitated its decay. This data suggests that IGFBP5 mRNA may not be targeted for degradation by miR-140 during chondrogenic differentiation. The possibility remains that the translation of this gene is inhibited by miR-140; however, the small sample quantities and absence of reliably cross-reactive antibodies prohibits the evaluation of these possibilities at this time. In contrast, ADAMTS-5 and CXCL12 mRNA levels were high before differentiation, but low throughout differentiation, suggesting that these genes were targeted by miR-140. CXCL12 gene expression has been shown to decrease rapidly upon osteogenic and chondrogenic induction [43,44]. Blockade of CXCL12 signaling before BMP-2-stimulated osteogenesis decreased the expression of osteogenic markers and abrogated the accumulation of osteocalcin, suggesting that CXCL12 is involved in initiation of BMP-2-induced osteogenesis [43]. Regulation of CXCL12 expression appears to be important for chondrogenic induction, but whether its expression decreases because it would otherwise impede differentiation or whether its expression is required for initiation of chondrogenesis is unknown.
The roles of ADAMTS-5 suppression in cartilage development are not entirely clear. One hypothesis is that ADAMTS-5 is suppressed during cartilage differentiation and development to facilitate the accumulation of aggrecan in the ECM, but that it is increased in established articular cartilage to regulate baseline turnover of aggrecan [45]. This would help explain the seemingly high expression of ADAMTS-5 in our samples from articular chondrocytes (Fig. 5E). However, considerable variability in both CXCL12 and ADAMTS-5 expression was evident in those samples. The possibility of clinically undetectable joint disease remains a potential contributor to the variation, as it cannot be definitively excluded from the samples [41, 42, 46]. It will be important to study the effect of miR-140 overexpression and knockdown on the expression of both previously identified and novel targets in eCB-MSCs to provide further insight into the molecular pathways in these cells.
Another parameter that merits investigation is the direct effect of TGF-β on the expression of miR-140. The addition of soluble growth factors of the TGF-β superfamily to chondrogenic differentiation media is an established method for stimulating in vitro chondrogenesis [31]. However, a recent study by Davis et al. has revealed that a selective group of miRNAs are directly regulated by signal transducers of the TGFβ/BMP pathway, the Smads. The majority of miRNAs in this group contain a consensus sequence (R-SBE) in the stem region of their primary transcripts that is similar to Smad binding element (SBE). They propose a mechanism in which receptor-regulated Smads (R-Smads) bind directly to R-SBE in pri-miRNAs in response to TGFβ or BMP-4 stimulation to provide a platform for the recruitment of miRNA processing enzymes, Drosha and DGCR8, thus facilitating pri-miRNA cleavage by Drosha. MiR-140 was identified as one miRNA containing a putative R-SBE. However, this study was conducted in human pulmonary artery smooth muscle cells and the TGFβ or BMP-4 stimulation was applied for 24 h or less [47]. Determining whether this mechanism also applies to the expression of miR-140 during chondrogenic differentiation will improve our understanding of the complex network of molecular interactions that regulate chondrogenesis.
The finding that miR-140 expression correlates with both cartilage differentiation and with the suppression of CXCL12 and ADAMTS-5 mRNA levels suggests that miR-based manipulation of gene expression might represent a useful component of therapeutic approaches aimed at improving cartilage differentiation. It will be important to overexpress miR-140 during chondrogenic differentiation and determine whether it could improve or even induce chondrogenesis. Evidence for the potential of miRNAs to alter cell fate has been provided by recent studies in which specific miRNAs have been used to create induced pluripotent stem cells [48] and to transdifferentiate fibroblasts to neurons [49,50]. The induction of in vitro chondrogenesis using miRNA has not yet been shown, but the tissue specificity of miR-140 makes it an excellent candidate. This approach could be applied to cell-based cartilage repair strategies, such as chondrocyte implantation and osteochondral grafts.
Clinically, there are several problems associated with direct miRNA administration, such as delivery to targeted cells and immunogenic reactions; however, Ochi and colleagues have reported on intra-articular and intravenous administration of synthetic miRs in experimental models of arthritis with some success. The injection of double-stranded miR-15a into the joints of arthritic mice induced apoptosis in the synovium by suppressing Bcl-2 expression and although a therapeutic effect was not demonstrated, there was little evidence of toxicity [51]. More recently, the same group demonstrated that intravenous administration of miR-146a prevented joint destruction by inhibiting osteoclastogenesis in arthritic mice [52]. Whether or not double-stranded miRNA can be taken up by chondroprogenitors in vivo in the joint following cartilage damage has yet to be determined.
In conclusion, the results of this study show that miR-140 exhibits a dynamic expression pattern during chondrogenesis of eCB-MSCs that correlates with the expression of the transcription factor Sox9. Additionally, CXCL12 and ADAMTS-5 mRNA levels were suppressed during chondrogenic differentiation, while miR-140 expression was increased, suggesting that this miRNA may target these genes for degradation. Further studies, including overexpression and knockdown of miR-140, will clarify the role of miR-140 during chondrogenesis of MSCs and determine the utility of this miRNA as an enhancer or inducer of chondrogenesis.
Footnotes
Acknowledgments
This project was supported by the Danish Agency for Technology, Production and Innovation, Equine Guelph and Partners, NSERC (Canada, JL) and the Department of Biomedical Sciences at the University of Guelph.
The submitted material was presented at the Nordic Orthopaedic Federation 56th Congress in Tallinn, Estonia on May 3rd, 2012 and at the 3rd North American Veterinary Regenerative Medicine Association Conference in Savannah, Georgia on November 9th, 2012.
Author Disclosure Statement
M.E.B. and J.L. declare no competing financial interests. T.G.K. is the executive director, Scientific Affairs, of eQcell Therapies, Inc., and holds a noncontrolling share in the company.