
Abstract
Human embryonic stem cells (hESCs) are known for their potential usage in regenerative medicine, but also for handling sensitivity. Much effort has been put into optimizing the culture methods of hESCs. It has been shown that the use of Rho-associated coiled-coil kinase inhibitor (ROCKi) decreases the cellular stress response and the apoptotic cell death in hESC cultures that have been passaged enzymatically. These observations sparked a wide use of ROCKi in hESC cultures. We and others, however, noted that cells passaged enzymatically with the use of ROCKi had a different morphology compared to cells passaged mechanically. Here we show that hESCs that were enzymatically passaged displayed alterations in the nuclear size compared to cultures that were mechanically passaged. Notably, a dramatically decreased expression of the genes encoding common pluripotency markers, such as OCT4/POU5F1 and NANOG were revealed in enzymatically passaged hESCs compared to mechanically passaged, while such differences were not significant when assessing protein levels. The differences in gene expression did not correlate strongly with commonly analyzed histone modifications (H3K4me3, H3K9me3, H3K27me3, and H4K16ac) on the promoters of these genes. Surprisingly, the effects of enzymatic passaging were at least in part reversible as the gene expression profile of enzymatically passaged hESCs that were transferred back to mechanical passaging, showed no significant difference compared to those hESCs that were continuously passaged mechanically. Our results suggest that enzymatic passaging influences parameters associated with hESC characteristics, and emphasizes the importance of using cells handled in the same manner when comparing results both within and between projects.
Introduction
S
Various passaging techniques have been established [8 –13] to facilitate the use of hESCs, for example, mechanical passaging using a surgical scalpel or cell scraper [14], or enzymatic passaging using trypsin-like solutions to dissociate the cells into single cells [15]. It was earlier noted that hESCs do not grow as well when dissociated from each other [16]. This dissociation is an effect from passaging the cells enzymatically, where a high degree of cell death is believed to correlate to the loss of cell-to-cell interactions [15,17]. Therefore, the use of Rho-associated coiled-coil kinase (ROCK) inhibitor (ROCKi; Y-27632) emerged as a sound solution [15]. Rho is a member of the superfamily Ras and acts through a number of different proteins, including ROCK. Combining ROCKi with enzymatic passaging became a standard procedure in many laboratories and has been reported to yield an increase in cloning efficiency from ∼1% to ∼27% [15]. The chemical structure and pharmacological properties of Y-27632 have been described elsewhere [17,18].
When hESCs are dissociated into single cells, they are more prone to apoptosis and spontaneous differentiation [15,19] and using ROCKi is thus believed to minimize stress that occurs when using enzymatic passaging techniques. The effects seen in hESCs when using ROCKi is supposed to be due to the roles of Rho in the G-Protein Coupled Receptor and extracellular matrix (ECM)-induced intracellular signaling [20 –23]. It has further been shown in other cell types that ROCKi eliminates stress fibers and that ROCK is involved in the formation of actin stress fibers [18]. Many studies have investigated the effects of culturing conditions on hESCs [5,24] focusing on culturing media and matrices. In contrast, fewer studies have assessed the techniques used for passaging the cells. The aim of this study was therefore to investigate whether different passaging techniques, that is, mechanical or enzymatic, and the use of ROCKi resulted in any differences in parameters associated with hESC characteristics, such as the gene expression and histone modifications of commonly used pluripotency markers.
We used a set of four well-characterized hESC lines [25] that went through three common passaging techniques; (i) mechanical, (ii) enzymatic, and (iii) enzymatic with the use of ROCKi, all cultured on Matrigel. We noted a clear difference in the morphology of cells that were enzymatically passaged with ROCKi compared to the other techniques. Interestingly, there was a clear increase in the nuclear size in enzymatically passaged hESCs compared to mechanical irrespective of ROCKi treatment. Most notably, we found a decrease in relative gene expression in enzymatically compared to mechanically passaged hESCs of the most common markers for pluripotency in hESCs (OCT4/POU5F1, NANOG, and KLF4). This decrease in gene expression was not dependent on ROCKi treatment. The difference in gene expression profile correlated poorly with protein levels as assessed by immunocytochemistry as well as histone modifications in the promoters of the genes as assessed by chromatin immunoprecipitation (ChIP). The effects on gene expression seemed to be reversible as there was no difference between treatments in relative gene expression of the examined pluripotency markers when enzymatically passaged hESCs were transferred back to mechanical passaging for three passages. Our results suggest that enzymatic passaging with or without treatment with ROCKi influence the hESC state.
Materials and Methods
Culturing of hESCs
Four hESC lines were used; HS346 (46:XX), HS207 (46:XY), HS360 (46:XY), and HS401 (46:XY), all derived at Karolinska Institutet [25]. The hESCs were cultured in 37°C in 5% CO2 on mitotically inactivated (irradiation 40 Gy) human foreskin fibroblasts (hFFs) in a medium containing KnockOut Dulbecco's modified Eagle's medium (KO-DMEM; Gibco, Invitrogen Corporation, cat # 10829018) supplemented with 20% KnockOut Serum Replacement (KO-SR; Gibco, cat # 10828028), 2 mM glutamax (Gibco; cat # 35050), 0.5% penicillin–streptomycin (Gibco; cat # 15140122), 1% nonessential amino acids (Gibco; cat # 11140035), 0.5 mM 2-mercaptoethanol (Gibco; cat # 31350), and 8 ng/mL of the basic Fibroblast Growth Factor (bFGF; R&D Systems, cat # 234-FSE/CF), as previously described [1,11,26].
All four hESC lines were transferred from hFFs to Matrigel, a basement membrane matrix extracted from Engelbreth-Holm-Swarm tumor (BD Bioscience, cat # 356234) [27] using three different passaging methods: 1. Mechanical dissociation (MECH) performed by scraping small pieces of the colonies from the feeder layer using a surgical scalpel and subsequently passed onto freshly coated Matrigel plates. 2. Enzymatic passaging using ROCKi (ENZ + ROCKi) was performed with TrypLE Select (Gibco; cat # A1217701) together with a selective Rho-associated coiled-coil kinase (ROCK) inhibitor Y-27632 (Merck KGaA; cat # 688001) in brief; the hESCs were incubated 1 h before passaging with 10 μM ROCKi. ROCKi is taken up by the cell by carrier-mediated facilitate diffusion [18]. To reach maximal inhibition, the cells need to be incubated for a minimum of 30 min in order for the intracellular concentrations of the compound to be comparable to the concentration outside of the cell. Therefore, after 1 h, the culture medium was removed and the plates were washed twice with Dulbecco phosphate buffered saline (DPBS) 1×(Gibco; cat # 14190), and 500 μL of TrypLE Select was added to the plate and incubated at 37°C for 5 min. Colonies were allowed to de-attach and the plates were washed twice with one volume of the culture medium, transferred in to a centrifuge tube and centrifugated at 1,300 rpm for 5 min. The supernatant was discarded and the pellet resuspended in a fresh mTeSR1 culture medium [28] (Stem Cell Technologies; cat # 05850) supplemented with 10 μM ROCKi and plated on a new Matrigel-coated culture dish [15]. 3. Enzymatic passaging without ROCKi (ENZ) was performed as (ENZ + ROCKi), but without treating the hESCs with ROCKi.
The hESCs were cultured in the mTeSR1 medium on Matrigel-coated plates [27] for at least three passages using these three different passaging methods; MECH, ENZ + ROCKi, and ENZ before collecting cells for analyses. For experimental setup, see Supplementary Fig. S1 (Supplementary Data are available online at
Doubling time (DT), measured in days, was calculated according to Miyazawa et al. [29] using the following equation: DT=(T − T0) log 2/(log N − log N0), where (T − T0) is the incubation period for the duration of experiment (in days), N is the number of cells at the end of the incubation period, and N0 is the number of cells seeded initially.
Morphology
The morphology of the hESCs was monitored using an inverted phase-contrast microscope (Nikon ECLIPSE TE2000-S). Pictures were taken using 40×and 400×magnification (Nikon Digital sight camera) at each passage. The nuclear size was analyzed by Zeiss Axiovision software in 158–232 nuclei of cells. The cells were fixed before analysis using a formaldehyde solution based on 4% paraformaldehyde 48 h after the last split in each condition in three replicates.
Immunocytochemical staining
The hESC lines were passaged according to the three different methods (MECH, ENZ + ROCKi, and ENZ) to coverslips (Nunc Thermanox Coverslips, tissue culture treated; Thermo Scientific; cat. no. 12-565-27) precoated with Matrigel and cultured for 2 days in the mTeSR1 medium before fixation. The fixating of the cells in brief; the medium was aspirated and the cover slips were washed twice with DPBS. About 4% of paraformaldehyde (PFA) was added in a volume to cover the slips and then incubated at room temperature for 10 min. After incubation, the PFA was removed and fixed cells were washed twice with DPBS and stored in DPBS at 4°C before performing immunocytological staining. The hESCs were permeabilized in 0.2% Triton X-100 in DPBS for 5 min at room temperature and washed three times in DPBS. The hESCs were blocked in 5% fetal bovine serum (FBS; Gibco, cat # 16000044) in DPBS for 30 min at room temperature and washed again 3 times in DPBS. The primary antibody specific for the pluripotency marker NANOG (goat, polyclonal, and FITC conjugated; R&D systems, cat # NL1997G) was diluted 1:100 in DPBS containing 5% FBS. TRITC-conjugated phallodin (Sigma-Aldrich; cat # P1951) was diluted 1:300 from a stock solution of 0.5 mg/mL in methanol. The hESCs were incubated with NANOG and phallodin for 1 h and washed three times with DPBS. Nuclear staining was performed with 4,6-diamino-2-phenylindole dihydrochloridine (DAPI; Sigma-Aldrich, cat # D9542) diluted 1:1,000 from a stock solution of 1 mg/mL in H20. The cover slips were mounted on object slides by the use of Fluoromount-G.
The hESC line HS346 was stained using POU5F1, NANOG, and SOX2 (all from Cell signaling Technology; StemLight™ Pluripotency Transcription Factor Antibody Kit, cat # 9093). The primary antibodies were diluted 1:400 and performed according to the manufacturer's protocol. Alexa Fluor 594 goat anti-rabbit (Invitrogen; cat # A11012) was used as the secondary antibody, diluted 1:300. Cells were also mounted on object slides by using Fluoroshield with DAPI (Sigma; cat # F6057). Immunostainings were carried out on MECH, ENZ + ROCKi, and ENZ as well as reversed conditions.
Gene expression analysis using reverse transcriptase-quantitative polymerase chain reaction (qRT-PCR)
In brief, total RNA from all four hESC lines was extracted by the RNeasy Mini kit (QIAGEN; cat # 74104) according to the manufacturer's recommended protocol, and converted to cDNA using the High-Capacity cDNA Reverse Transcription Kit [Applied Biosystems (ABI); cat # 4368814] also according to the manufacturer's recommended protocol. Quantitative polymerase chain reaction was performed by using TaqMan® Universal PCR Master Mix (ABI; cat # 4369510) together with TaqMan gene expression assays (Table 1). The gene expression was calculated according to the ddCt method, where the mean of triplicates from each sample was normalized to the mean of 18S.
Chromatin immunoprecipitation (ChIP)
ChIPs were carried out using a ChIP assay kit (ChIP-IT express, Active Motif and Diagenode; cat # 53008) and by a LowCell# ChIP kit (Diagenode cat # kch-maglow-A48), both according to the manufacturer's protocol. Cells were fixed in 4% PFA for 10 min at room temperature. Antibodies used (Table 2). SYBR Green PCR master mix (ABI; cat # 4309155) was used for analysis of samples as previously described [30] with primers specifically designed for POU5F1, NANOG, and KLF4 (Table 3). The levels of histone acetylation and methylation were assessed as the ratio of the PCR product after ChIP toward the signal generated by PCR of the input DNA before ChIP.
ChIP, chromatin immunoprecipitation.
Reversed culturing
To study whether the possible effects on the pluripotency of different passaging methods (MECH, ENZ + ROCKi, and ENZ) on hESCs would remain or revert when transferred back to their standard culture condition (MECH), these conditions were transferred back from enzymatically (ENZ + ROCKi and ENZ) to mechanical passaging. After being cultured and passaged mechanically for at least 3 passages, the cells were reanalyzed by gene expression using markers for pluripotency; POU5F1 and NANOG.
Immunoblotting
Cells were lysed in the ice-cold RIPA lysis buffer (150 mM NaCl, 1% Triton-X-100, 0.5% Na-deoxycholate, 0.1% SDS, and 50 mM Tris-HCl, pH 8.0) with the addition of a complete, Mini, EDTA-free Protease Inhibitor Cocktail (Roche) according to the manufacturer's recommendations. The samples were resolved by the Novex tris-glycin gel 4%–20% (Invitrogen) and transferred to Hybond-P membranes (GE Healthcare). After blocking in Amersham ECL Prime Blocking (GE Healthcare), the membranes were incubated with appropriate antibodies and washed in PBS-T; the blots were visualized with Amersham ECL Select (GE Healthcare) using Image Quant LAS 4010 (GE Healthcare).
Primary antibodies: monoclonal mouse anti GAPDH (G8795; Sigma Aldrich) diluted 1:5,000, monoclonal rabbit anti NANOG, and monoclonal rabbit anti OCT-4 (9093S; Cell Signaling), diluted 1:1,000. Secondary antibodies: anti mouse IgG HPR-linked (NA931; GE Healthcare) diluted 1:100,000 and anti rabbit IgG HPR-linked (NA934; GE Healthcare), diluted 1:50,000.
Flow cytometry
Flow cytometry analysis was carried out for two hESC lines, HS346 and HS401, using the monoclonal liver alkaline phosphatase antigen TRA2-54 [5]. An FITC tagged secondary antibody was used recognizing mouse IgG and IgM. Quantitative analysis of cell surface antigen expression was carried out for MECH, ENZ, and ENZ + ROCKi, as well as for reversed conditions.
Statistical analysis
One-way repeated measures analysis of variance (ANOVA; SigmaPlot 11.0; Systat Software, Inc.) was used to compare the difference between the three passaging methods. A difference was considered statistically significant when the P value was ≤0.05 and highly significant (***) when the P value was ≤0.001.
Results
Different passaging techniques and ROCKi treatment yield differences in hESC morphology
We initially observed that mechanically or enzymatically passaged hESCs displayed different morphologies when cultured on hFFs and on Matrigel. On hFFs, the hESCs formed nice compact colonies guided by the feeder layer, giving rise to an elongated appearance (Supplementary Fig. S2). When grown on Matrigel, the cells formed colonies with a circular appearance (Fig. 1A–C), where the individual cells were less tightly packed together (Fig. 1D–F). When passaged using the three different methods (MECH, ENZ + ROCKi, and ENZ), it was observed that hESCs passaged enzymatically with or without ROCKi displayed a different morphology than cells passaged mechanically (Fig. 1A–C). A difference in the growth rate was also noted. When analyzing the DT as described previously [29], we found the DT to be slower, 0.95±0.04 days, for cells passaged enzymatically without ROCKi compared to 0.71±0.07 days for cells passaged mechanically as well as passaged enzymatically with ROCKi.

Inhibition of Rho with the downstream target ROCK (Y27632) steers hESCs into structures not resembling typical hESC colonies.
Cytoskeleton organization and nuclear size are affected by passaging techniques and/or ROCKi
Cells cultured with all three passaging techniques (MECH, ENZ + ROCKi, and ENZ) were fixed after 48 h, and subsequently stained for stress fibers using phalloidin, which binds to F-actin, NANOG, for pluripotency, and DAPI for nuclear staining. Cells that were mechanically passaged and cells that were enzymatically passaged without ROCKi throughout the experiment stained positive for F-actin, indicating that the cells were under stress (Fig. 2A). The cells were immunoreactive for NANOG after passaging with the three different methods. Generally, the nuclei of cells passaged enzymatically appeared bigger. We therefore performed a careful calculation of the nuclear area of a total of 163 nuclei for cells passaged mechanically, 232 nuclei for ENZ + ROCKi, and 158 nuclei for ENZ. This analysis confirmed that cells passaged enzymatically (with or without ROCKi) displayed a significantly larger nuclear area compared to those passaged mechanically (Fig. 2B). The morphological differences seemed to remain as long as the cultures were maintained.
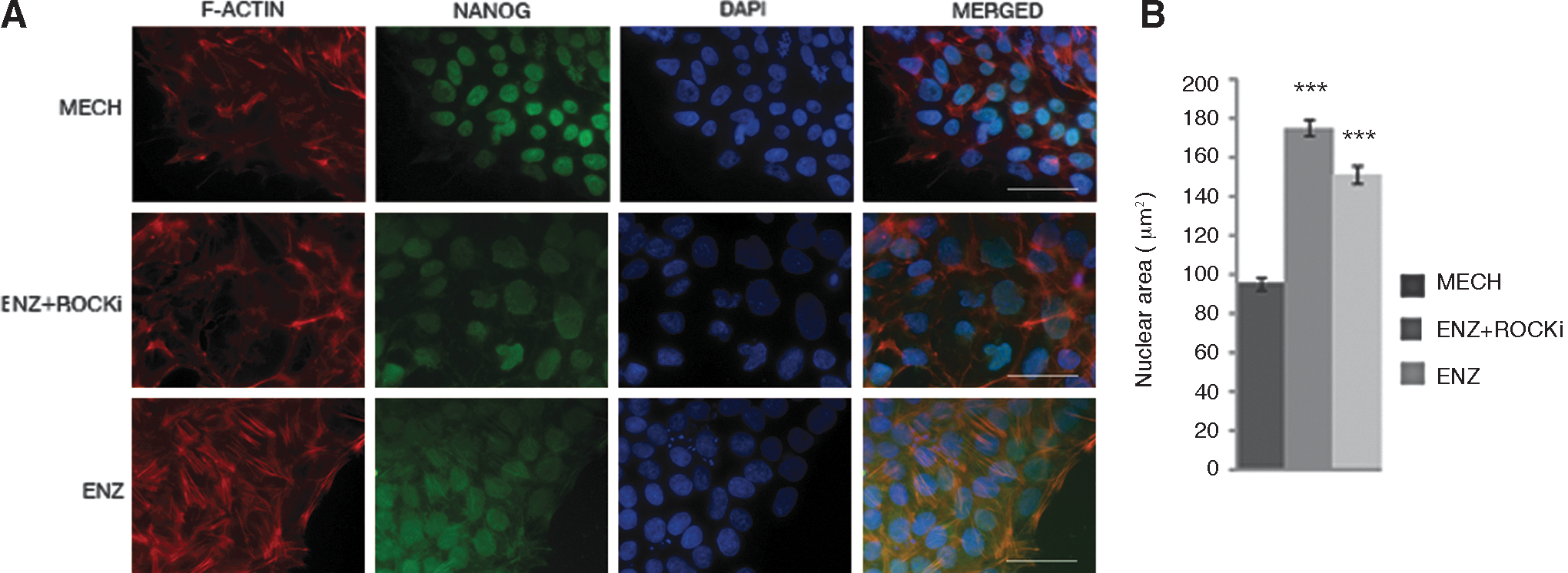
Immunostaining and nuclear size.
Differences in gene expression of pluripotency markers after mechanical versus enzymatic passaging of hESCs
We next investigated the gene expression of various markers that previously have been demonstrated to be essential for hESC characteristics. Surprisingly, the relative mRNA levels of the pluripotency markers POU5F1 and NANOG were dramatically and significantly (P≤0.001) reduced for hESCs that were enzymatically passaged and treated with ROCKi (ENZ + ROCKi; POU5F1 4.2% and NANOG 2.3%) and for enzymatically passaged cells without ROCKi (ENZ; POU5F1 3.1% and NANOG 2.1%) when compared to cells passaged mechanically (MECH; 100%) (Fig. 3). KLF4, which is a transcription factor known for its role in cell proliferation, differentiation, and survival in hESCs [23,31], showed a similar expression pattern as the gene expression was significantly (P≤0.001) downregulated when cells were passaged enzymatically with ROCKi (ENZ + ROCKi; 17.0%) or without (ENZ; 12.3%) compared with mechanically passaged cells (MECH; 100%) (Fig. 3). The percentage was calculated as mean relative quantity of all four hESC lines for the three different markers (Fig. 3). For individual cell lines, see Supplementary Data (POU5F1: Supplementary Fig. S3; NANOG: Supplementary Fig. S4; KLF4: Supplementary Fig. S5).
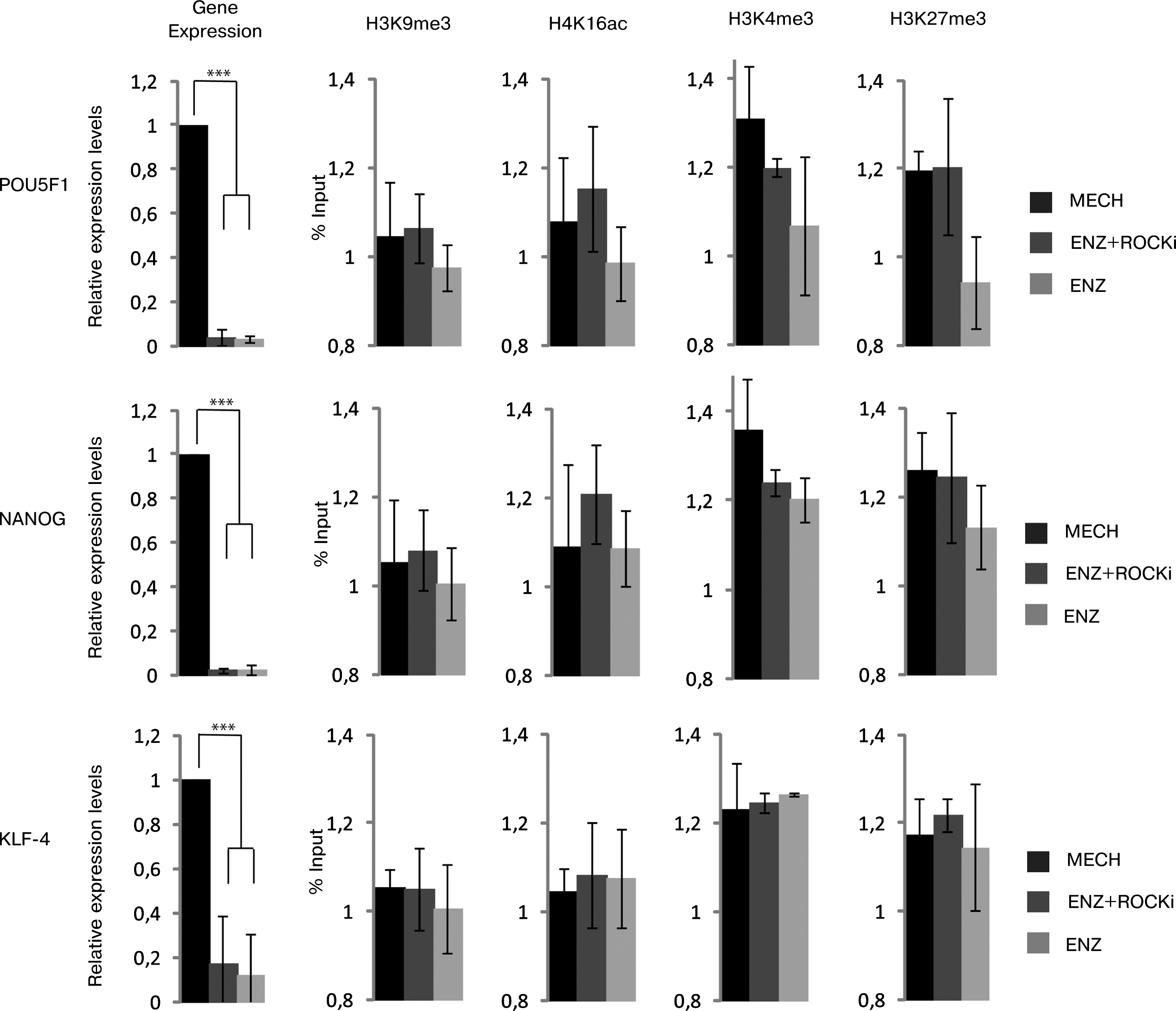
Regulation of pluripotency genes in mechanically and enzymatically treated hESCs. Gene expression was calculated using the Ct (ΔΔCT) method and presented as mean relative quantifications across all four hESC lines. Gene expression of pluripotency-related genes in hESCs for POU5F1, NANOG, and KLF-4 decreased significantly when treated with (ENZ + ROCKi) or without (ENZ) ROCKi, as compared to mechanically passaged cells. Histone modification (H3K9me3, H4K14ac, H3K4me3, and H3K27me3) for pluripotency markers POU5F1, NANOG, and KLF4 showed no correlation with the decreased gene expression. ***P<0.001.
To elucidate whether the enzymatically passaged cells were gaining a cancer cell-like profile, gene expression analysis was also performed for other markers associated with stem and cancer cell characteristics, namely, SRY (Sex determining region Y)-box2 (SOX2), Insulin-like growth factor 2 (IGF2), C-MYC, and H19 [32 –34]. SOX2 and IGF2 followed the same pattern as the other markers, whereas both the oncogene C-MYC and the imprinted gene H19 were not expressed in any of the cell lines (data not shown). These results thus did not support the idea that the enzymatically passaged hESCs gained cancer cell-like characteristics.
Different passaging techniques do not affect histone modifications analyzed in hESCs
Transcription and the regulation of gene expression are coupled to histone modifications, and transcriptional complexes regulating histone methylation such as the Polycomb complex, and acetylation such as the NuRD complex, have been shown to be important for embryonic stem cell characteristics [35 –37]. We therefore investigated the relative levels of methylation and acetylation on a subset of lysines on histone H3 and H4 in the promoters of the above-analyzed genes using ChIP–qRT-PCR. We investigated trimethylated lysine 4 on histone H3 (H3K4me3), which when located to a promoter is a mark of transcriptional activation due to recruitment of initiation factors, and trimethylated lysine 9 and 27 on histone H3 (H3K9me3, H3K27me3), which are regulated marks for transcriptional repression and associated with heterochromatin [30 –32]. We could, however, not detect any significant differences in trimethylation of the analyzed histone lysines between the groups of different treatments (Fig. 3).
Due to the increased nuclear area in hESCs passaged enzymatically, we speculated that there may be a more general effect on the chromatin structure that obscured more specific differences in histone lysine methylation between the different treatments. We therefore included analysis of acetylated lysine 16 on histone H4 (H4K16ac), a modification influencing histone–histone interactions, nucleosome organization, and a higher order chromatin structure [38]. We did not detect any significant differences in H4K16ac between the groups of hESCs receiving different treatments (Fig. 3). These negative results only suggest that there were no differences in these particular lysines at these locations, but could point to a role for other, less studied, histone modifications as well as epigenetic regulators beyond the chromatin structure, such as noncoding RNA.
Different passaging techniques do not affect protein levels of pluripotency markers in hESCs significantly
Due to decreased levels in gene expression of the pluripotency markers, we speculated whether the protein levels also would decrease depending on the different passaging methods. One cell line, HS346, was stained for the pluripotency markers POU5F1, NANOG, and SOX2. We found that the hESC lines expressed immunoreactivity for these pluripotency markers to a similar extent regardless of the different passaging techniques used (Fig. 4A, Supplementary Fig. S6). Similar results for POU5F1 and NANOG were obtained by immunoblotting (data not shown). To determine the fraction of undifferentiated hESCs, we performed FACS analysis of TRA-2-54-positive cells. TRA-2-54 is an antibody detecting alkaline phosphatase, a general marker of pluripotent cells [39]. A total of 50,000 cells were calculated for all three passaging methods, MECH, ENZ + ROCKi, and ENZ, as well as for the reversed condition (see further below). Although a trend similar to the gene expression differences was noted, no significant differences between the different passaging methods could be detected (Fig. 4B).
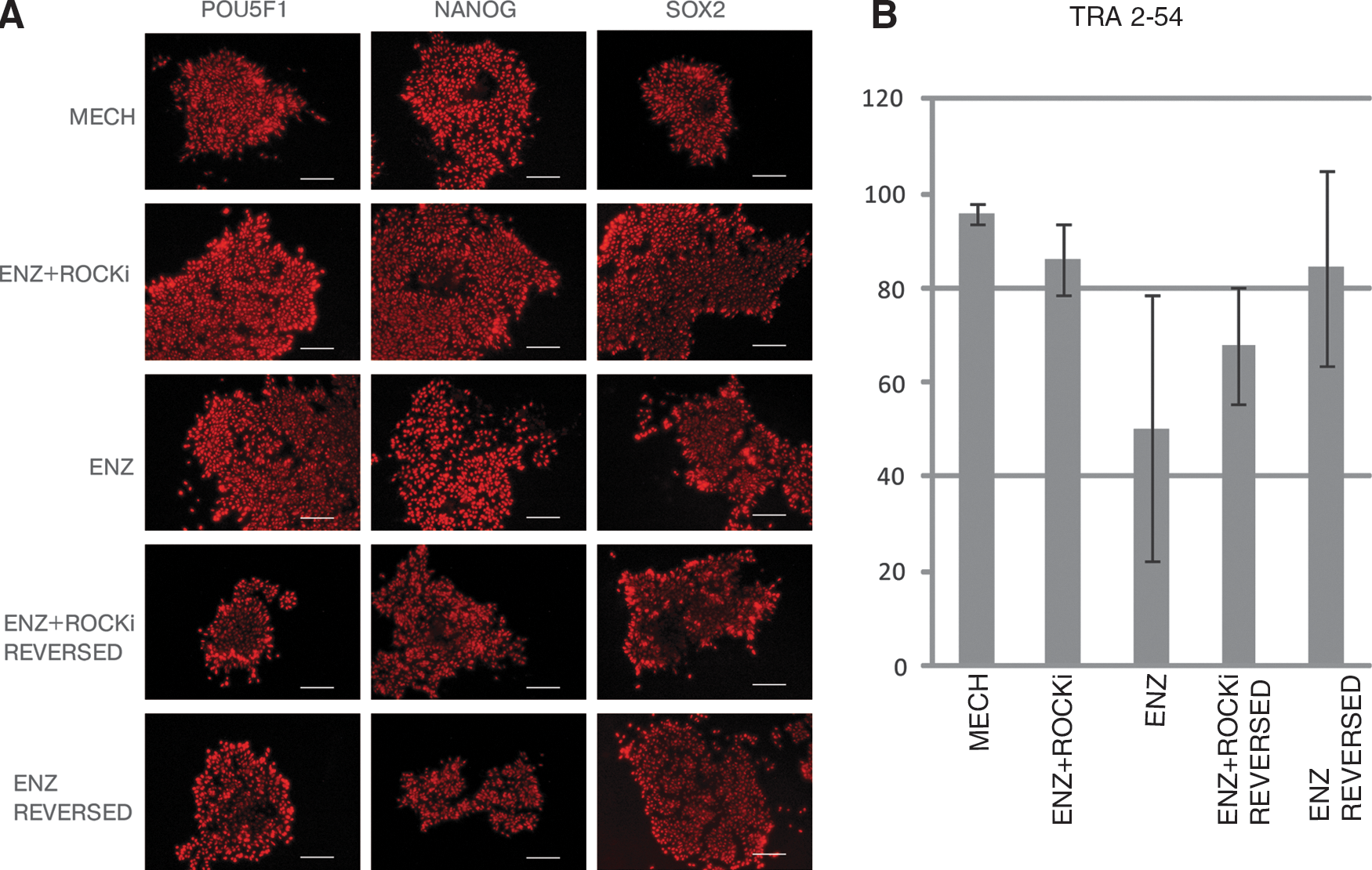
Protein levels.
Passaging dependent differences in morphology and gene expression in hESCs are reversible
To investigate whether the morphology of hESCs could be reversed, we transferred enzymatically passaged cells both with (ENZ + ROCKi) and without (ENZ) ROCKi back to their original mechanical passaging method (mechanically (MECH) using a surgical scalpel) in three of the hESC lines used and passaged with this technique for at least three passages to investigate whether the morphology of the cells was reversed to their original round sharp edged colonies observed in cultures where the cells had been mechanically passaged throughout the study. We further analyzed whether the decrease in pluripotency markers when passaged enzymatically would remain or be reversible. When compared with cells passaged using the mechanical method (MECH) throughout the entire study, it was shown that the cells transferred back from both enzymatic passaging methods (ENZ + ROCKi, ENZ) to the mechanical passaging method (MECH), did restore their original round sharp edged colony morphology and nuclear size (data not shown), and their pluripotency properties as both POU5F1 and NANOG mRNA levels were similar to the cells that were mechanically passaged only. Cells passaged with ROCKi had a mean expression between the three hESC lines of 93.0% of POU5F1 and 104.1% of NANOG. Cells passaged without ROCKi had a mean expression between the three different hESC lines of 125.6% for POU5F1 and 108.4% for NANOG [All compared with a 100% expression of the markers for cells passaged mechanically (Fig. 5)].
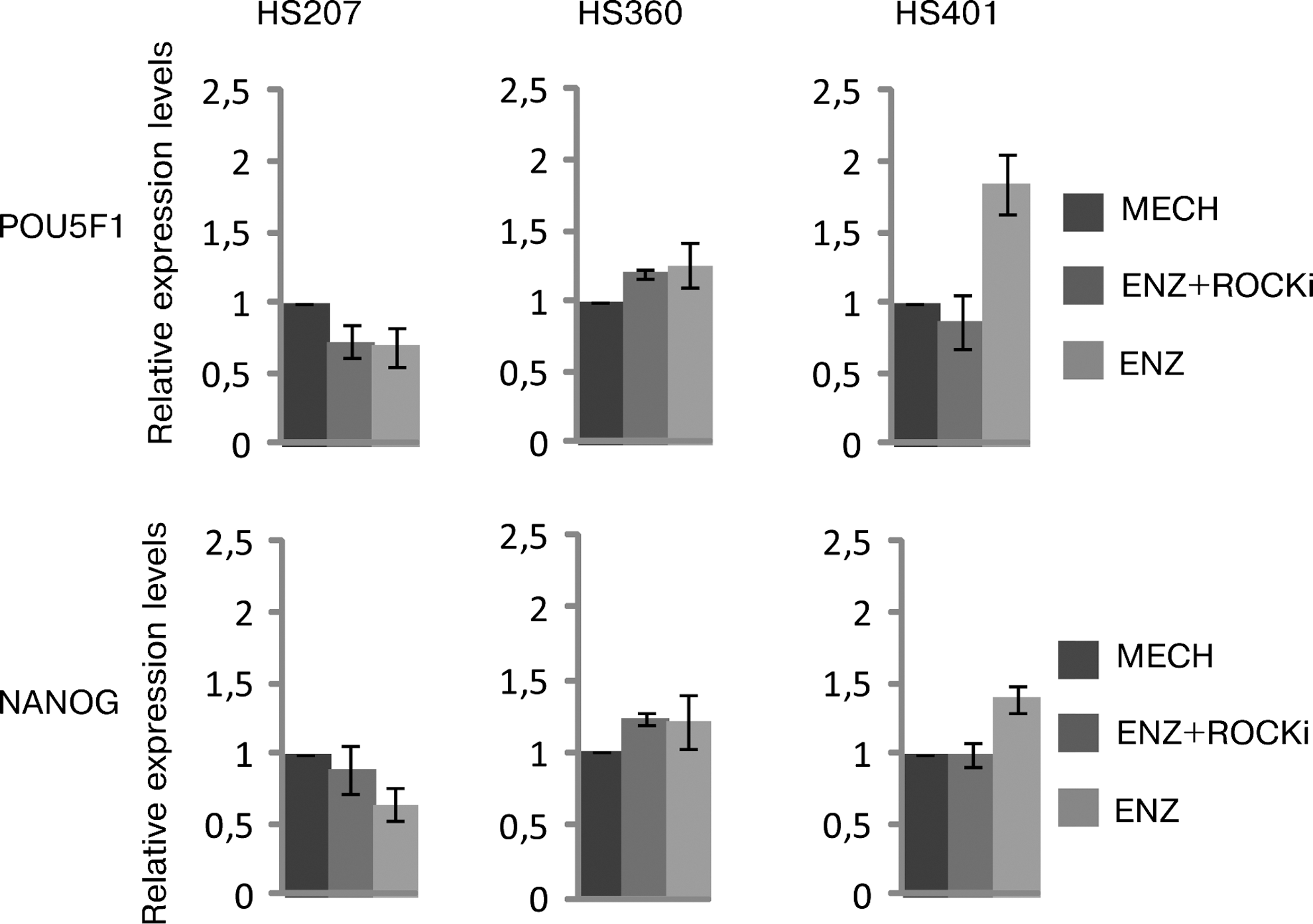
Gene expression profiles (qRT-PCR) of POU5F1 and NANOG in reversed samples. Cells were treated with (ENZ + ROCKi) or without (ENZ) ROCKi and transferred back using mechanical passaging for at least three passages. Data are presented as relative quantity and was done using the Ct (ΔΔCT) method. The expression of pluripotency-related genes POU5F1 and NANOG was reverted back when transferred back to the control method (MECH).
Discussion
During this study it has become clear that, in our hands and with the cell lines that we have used, there are differences between hESCs in the morphology, stress fiber organization, and the gene expression of pluripotency markers depending on which passaging techniques are being used, whereas we found little or no difference in pluripotency marker protein levels or histone modifications of the residues we investigated. Passaging cells enzymatically using ROCKi has been considered to be a relatively easy and efficient strategy for hESC propagation, compared to the more technically difficult mechanical passaging procedure. The mechanical passaging method using various techniques (e.g., surgical scalpels) has been thought to induce less stress to the cells, but instead risking breakage of the cell itself, compared to enzymatical passaging were the cells lose their cell–cell interaction, and undergo more stress when introduced to different enzymes. The mechanical passaging methods are also time-consuming resulting in the cultures being out from the incubator for long periods, while enzymatical techniques are considered faster.
Our results suggest that there are additional effects on hESC morphology and gene expression due to different passaging techniques and ROCKi treatment than previously understood. ROCK is a Rho-binding protein involved in regulating stress fiber formation and focal adhesion assembly, and thereby, the cell adhesion to the ECM [22] in line with our results that the actin organization is different in enzymatically passaged cells treated with ROCKi. Indeed, previous studies have raised caution regarding prolonged cultivation of hESC lines in the presence of ROCKi [40]. The dramatic decrease in mRNA levels using pluripotency markers POU5F1, NANOG, and KLF4 (Fig. 3 and Supplementary Fig. S3–S5) further emphasizes the importance of cautious interpretation of hESC results obtained using different passaging techniques.
Although there was a significant decrease in the gene expression of several factors, changes in protein levels were not clear. The results from immunocytochemical experiments suggest that there was no significant decrease in the number of cells immunoreactive for POU5F1, NANOG, or SOX2 (Fig. 4A), but a change in protein levels in a subset of immunoreactive cells is harder to detect with immunocytochemistry as well as with immunoblotting. We therefore performed FACS analysis using the TRA-2-54 antibody, and although there was no statistically significant difference in the numbers of TRA-2-54-positive cells between the different groups, we noted a large variation in the number of TRA-2-54-positive cells in those cultures that were passaged enzymatically compared to mechanically passaged cultures (Fig. 4B). The enzymatically passaged cells further grew slower as assessed by calculating the DT. These observations should be interpreted with caution, but a possible cause for such variations as well as the discrepancy between gene expression and protein levels could be that the cells are compensating for a loss in the gene expression via a change in the protein turnover and metabolic rate. The importance of post-translational regulation of pluripotency factors as well as cell metabolism for ES cell characteristics is well-documented [41,42], and it will be of considerable interest to investigate whether such cellular events are affected by passaging techniques.
It should be noted that the results generated from enzymatic passaging in this study using TrypLE Select may differ from results from other methods of enzymatic passaging. It is further of great importance to consider the substrate used to grow the hESCs on. We and others have previously demonstrated the influence of substrate stiffness on tissue stem cell characteristics [43], and culturing hESCs on other substrates may have a more dominant influence on hESC characteristics than passaging techniques [44]. Lastly, the hESCs were still positive for pluripotency markers when assessing protein levels and thus should still be considered pluripotent [45]. However, the results of the present study indicate the importance of using cells that have been handled in the same manner when comparing data both within and between projects.
Conclusion
Different passaging methods (MECH, ENZ + ROCKi, and ENZ) affect the morphology, nuclear size, actin staining, and gene expression of commonly used stem cell markers associated with pluripotency. However, the effects of the enzymatic passaging technique were found to be reversible. These findings demonstrate the importance of comparing cells that have been treated with the same passaging techniques and should in future studies be considered equally important to the ECM and culture media effects.
Footnotes
Acknowledgments
The authors are grateful to Outi Hovatta and colleagues for providing cells and technical support, to members of the Hermanson laboratory for input, and to GE Healthcare for expertise and access to Amersham ECL Select and Image Quant LAS 4010. KF is supported by grants from the Stockholm County Council. PA is supported by grants from the Ludwig Institute, Karolinska Institutet, Swedish Cancer Society (CF) and the Swedish Research Council. OH is supported by grants from Karolinska Institutet, the Swedish Research Council (VR-MH, DBRM), the Swedish Childhood Cancer Foundation (BCF), and the Swedish Cancer Society (CF). RBT is supported by grants from Karolinska Institutet.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.