
Abstract
The technology to reprogram human somatic cells back to pluripotency allows the production of patient-specific induced pluripotent stem cells (iPSCs) and holds a great promise for regenerative medicine. Choosing the most suitable cell type for induction and reducing the risk of viral transgene activation, especially oncogene activation, are important for iPSC research. To date, human dermal fibroblasts (HDFs) are the most frequent cell source used for iPSC generation, but they have several limitations. An invasive skin biopsy must be performed to obtain HDFs, and HDFs must be cultured for a prolonged period before they can be used for experiments. Thus, in an effort to develop a suitable source for iPSC studies to avoid the limitations mentioned above, we have here identified stromal cells derived from menstrual blood (MenSCs) as suitable candidates. In the present study, we found that MenSCs can be reprogrammed to pluripotent status by doxycycline-inducible lentiviral transduction of OCT4, SOX2, and KLF4. Additionally, we found that MenSCs have a significantly higher reprogramming efficiency than HDFs. The combination of OCT4 and SOX2 is sufficient to reprogram MenSCs into iPSCs without the use of c-MYC or KLF4. The resulting MenSC-iPSCs showed the same characteristics as human embryonic stem cells with regard to morphology, pluripotent markers, gene expression, and the epigenetic status of pluripotent-cell-specific genes. These cells were able to differentiate into various cell types of all 3 germ layers both in vitro and in vivo. Therefore, MenSCs may be a preferred candidate for generation of iPSCs.
Introduction
Recent studies have demonstrated that human iPSCs can be generated from various cell origins, such as neonatal or adult dermal fibroblasts [1,3,8 –11], keratinocytes [12], neural stem cells [13], adipose stem cells [14], gut mesentery-derived cells [15], amnion-derived cells [16], and urine-derived cells [17]. An important observation from these studies is that the cell origin has a significant influence on reprogramming efficiency. Until now, human iPSCs have most commonly been derived from human dermal fibroblasts (HDFs). However, the need for an invasive skin biopsy and a prolonged growth period in cell culture before use will restrict the application of iPSCs derived from HDFs. For routine clinical application, an ideal cell source for iPSC generation is provided by discarded tissue, which contains cells with high proliferation capability and can be collected noninvasively in large amounts. To generate less-manipulated, safe iPSCs, it is important to reduce the number of reprogramming factors used, especially oncogenes.
Menstrual blood from the uterine lining has been recognized as a novel source of stem cells [18] with high regenerative capability after the menstrual cycle [19]. Additionally, stromal cells derived from menstrual blood (MenSCs) can be acquired without invasive procedures and avoid any ethical controversies. These cells display stem cell-like phenotypic markers, a propensity for self-renewal, high proliferative potential in vitro, and the ability to differentiate toward diverse cell lineages [18 –20]. Here, we showed that doxycycline (Dox)-inducible lentiviral transduction of OCT4, SOX2, and KLF4 can reprogram MenSCs into a pluripotent state with a significantly higher efficiency than that observed for HDFs. In particular, the combination of 2 factors, OCT4 and SOX2, was sufficient to reprogram MenSCs into iPSCs without the use of the oncogenes c-MYC or KLF4.
Materials and Methods
Derivation and culture of MenSCs
After informed consent was obtained, menstrual blood samples were collected from healthy, young donors (25–35 years of age) on the second day of menses. The samples were transferred to phosphate-buffered saline (PBS) containing penicillin/streptomycin and heparin. The supernatant of the samples was evaluated for bacteria. Mononuclear cells were fractionated in Ficoll (Sigma, St. Louis, MO) and cultured in Dulbecco's modified Eagle medium (DMEM): nutrient mixture F-12 (Ham's) (1:1; HyClone, Logan, UT) supplemented with 15% fetal bovine serum (FBS; Hyclone), 2 mM
Generation of human iPSCs
The generation and structure of Dox-controlled Tet-on-inducible lentiviruses expressing OCT4, SOX2, c-MYC, and KLF4 have been described in a previous report [8]. A constitutively active human ubiquitin C promoter-alpha promoter in the FUW lentiviral backbone (FUW-M2rtTA) drives the expression of reverse tetracycline transactivator (rtTA). The lentiviral vector maps are shown in Supplementary Fig. S1;(Supplementary Data are available online at
Flow cytometry analysis
FITC-conjugated mouse anti-human antibodies for CD45, CD105, CD73, and HLA-DR, as well as PE-conjugated mouse anti-human antibodies for CD34, CD44, CD38, and CD90 (Pharmingen, San Jose, CA), were used to characterize MenSCs. Stained cells were analyzed on a FACScalibur™ Flow Cytometer (Becton-Dickinson, Mountain View, CA) using CellQuest software.
Alkaline phosphatase and immunofluorescence staining of MenSC-iPSCs
Alkaline phosphatase (AP) staining was performed using an AP kit (Roche Applied Science, Mannheim, Germany) according to the manufacturer's instructions. For immunocytochemistry, cells were fixed with 4% paraformaldehyde for 10 min at room temperature. After washing with PBS, the cells were treated with PBS containing 10% normal bovine serum albumin (Sigma) and 0.1% TritonX-100 for 30 min at room temperature. The cells were then incubated with primary antibodies at 4°C overnight. The primary antibodies used in these incubations included mouse anti-SSEA-4 (1:100; Chemicon, Temecula, CA), mouse anti-TRA-1-60 (1:100; Chemicon), mouse anti-TRA-1-81 (1:100; Chemicon), rabbit anti-NANOG (1:100; Abcam, Cambridge, United Kingdom), rabbit anti-OCT4 (1:100; Santa Cruz Biotechnology, Santa Cruz, CA), and rabbit anti-SOX2 (1:100; Chemicon). Normal mouse or rabbit serum served as a negative control. Anti-rabbit or anti-mouse IgG secondary antibodies conjugated with a fluorescein (Santa Cruz Biotechnology) allowed the visualization of the interested antigens. Additionally, the nuclei were counterstained with DAPI. Immunofluorescent images were visualized under a fluorescence microscope (OLYMPUS BX51).
RNA isolation and reverse transcription
Total RNA was purified using an RNeasy Mini Kit (Qiagen, Valencia, CA) according to the manufacturer's instructions. Approximately 1 μg of total RNA from each sample was used for oligo(dT) 20-primed reverse transcription (SuperScript™ III First-Strand Synthesis System for RT-PCR; Invitrogen). Polymerase chain reaction (PCR) conditions included an initial denaturation step of 5 min at 95°C, followed by 35 cycles of 30 s of denaturation at 95°C, 30 s at annealing temperature, and 1 min of extension at 72°C, with a final elongation step at 72°C for 10 min. Amplified PCR products were resolved on (1.5%) agarose gels and visualized by ethidium bromide staining. Images were obtained using a Bio-Rad Gel document system. Primer sequences are shown in Supplementary Table 1.
Western blot analysis
The antibodies used in the western blots were rabbit anti-OCT4 (1:1000; Santa Cruz Biotechnology), anti-SOX2 (1:1000; Chemicon), rabbit anti-NANOG (1:250; Abcam), mouse anti-KLF4 (1:500; Chemicon), mouse anti-c-MYC (1:1000; Chemicon), and rabbit anti-β-Actin (1:10000; Sigma). IRDye700/800-conjugated secondary antibodies (Rockland Immunochemicals, Inc., Gilbertsville, PA) were used for immunodetection according to the manufacturer's instructions. Imaging was then performed using an Li-Cor Odyssey Infrared Imaging System.
Bisulfite sequencing
Genomic DNA (1 μg) was treated with the CpGenome DNA modification kit (Chemicon) according to the manufacturer's recommendations. The treated DNA was purified using a QIAquick column (QIAGEN). The promoter regions of the human OCT4 and NANOG genes were amplified by PCR. The PCR products were subcloned into pCR2.1-TOPO. Five clones of each sample were verified by sequencing using the M13 universal primer.
Microarray experiments
The gene expression profiles of H1 human ESCs, 3F MenSC-iPSCs, 2F MenSC-iPSCs, and MenSCs were obtained by using human U133 Genome 430 2.0 GeneChip arrays (Affymetrix, Santa Clara, CA). All experiments were performed and analyzed at CapitalBio Corporation. Briefly, total RNA from each sample was subjected to probe preparation, and cRNA was hybridized. Arrays were scanned using an AffymetrixGCS3000 device, and images were analyzed using GCOS software. The raw data for gene expression were extracted by using GeneChip Operating Software. The signal intensities from the mRNA channel in all arrays were normalized by using dChip Analysis software. Additionally, the signal intensities from the mRNA channel in all arrays were normalized by using the Robust Multiple-chip Analysis (RMA) algorithm.
In vitro and in vivo differentiation
For spontaneous differentiation through EB formation, MenSC-iPSCs were treated with collagenase type IV (Invitrogen) and cultured in a human ESC medium without bFGF in nontissue-culture-treated plates. After 8 days in suspension culture, embryoid bodies were transferred to gelatin-coated plates and cultured in a differentiation medium (high-glucose DMEM supplemented with 15% FBS) for another 8 days. The cells that underwent spontaneous differentiation were then detected with appropriate markers by immunofluorescence.
To test in vivo pluripotency, MenSC-iPSCs were harvested, suspended in PBS, and injected subcutaneously into the testis of SCID mice (3×106 cells per mouse). Approximately 8 weeks after the injection, the tumors that formed were dissected, fixed in 4% paraformaldehyde, embedded in paraffin, and examined histologically by hematoxylin and eosin staining and immunohistochemistry. Primary antibodies, including mouse anti-β-III-tubulin (1:100; Chemicon, Temecula, CA), mouse anti-smooth muscle actin (1:1000; Sigma), mouse anti-pan-cytokeratin (1:100; Santa Cruz Biotechnology), and rabbit anti-PDX-1 (1:100; Chemicon), were applied.
Karyotyping
Standard G-banding chromosome analysis was carried out in the Cytogenetics Laboratory at Peking University.
Results
Characterization of MenSCs
Cultured MenSCs showed stromal cell morphology after the primary and secondary passages (Fig. 1A, B). Flow cytometry analysis showed that these cells did not express CD34, CD45, CD38, or HLA-DR, but did express high levels of CD44, CD73, CD90, and CD105 (Fig. 1C). Immunofluorescence staining showed that ∼60% of these cells were positive for SSEA-4 (Fig. 2A), which was similar to the ratio of SSEA-4-positive cells (65.86%) obtained from flow cytometry analysis (Fig. 2B). The reprogramming genes OCT4, NANOG, KLF4, and c-MYC were detected by reverse transcription (RT)-PCR, but SOX2 was under detectable level (Fig. 2C), which was consistent with the protein expression profile. Moreover, OCT4, NANOG, and KLF4 had a lower protein expression level in MenSCs than in ESCs, whereas the c-MYC protein level was not significantly different among ESCs, MenSCs, and HDFs (Fig. 2D).

Characterization of stromal cells derived from menstrual blood (MenSCs). Phase-contrast microscopic images of MenSCs at primary passage
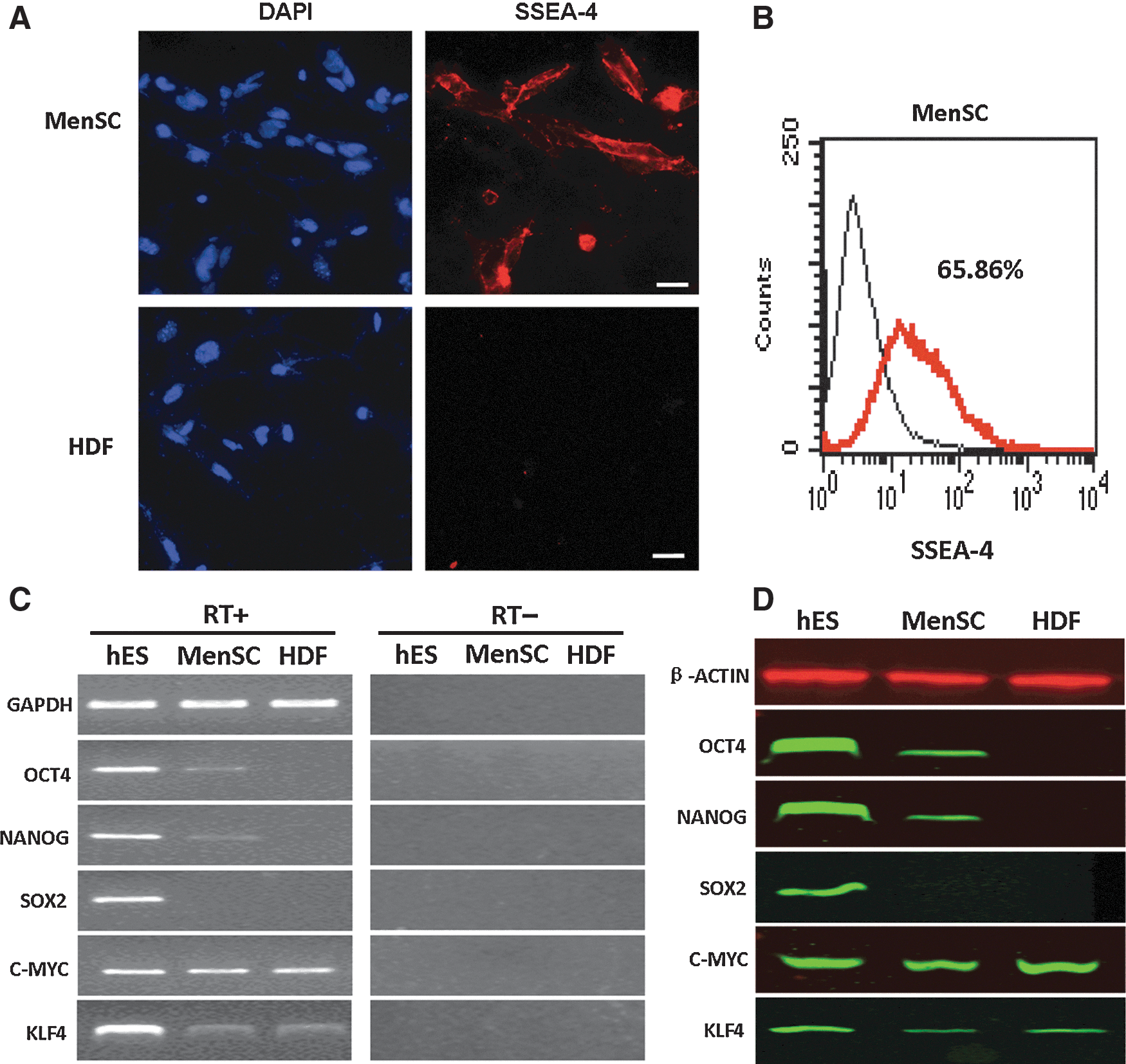
Expression of embryonic stem cell (ESC)-specific factors in MenSCs. Immunofluorescence staining demonstrated that these cells (∼60%) were positive for expression of SSEA-4. By contrast, human dermal fibroblasts (HDFs) were negative for SSEA-4
Generation of MenSC-iPSCs
A flow diagram for MenSC induction is shown in Fig. 3A. MenSCs and HDFs were transduced at a density of 1×105 cells per 35-mm plate using lentiviruses that encoded the 3 factors OCT4, SOX2, and KLF4 (OSK) or the 2 factors OCT4 and SOX2 (OS). Two days post-transduction, the cells were trypsinized, plated onto mitomycin C-treated feeder cells, and cultured in an ESC medium supplemented with 2 μg/mL Dox. At day 20, we performed AP staining and counted the number of AP-positive colonies, whose morphology resembled that of ESCs. We found that the number of AP-positive colonies generated from MenSCs was significantly higher than generated from HDFs transduced with OSK (Fig. 3B). When transduced with 2 factors (OCT4 and SOX2) devoid of KLF4, MenSCs demonstrated the formation of AP-positive colonies; however, no positive colonies were seen in HDFs (Fig. 3C).
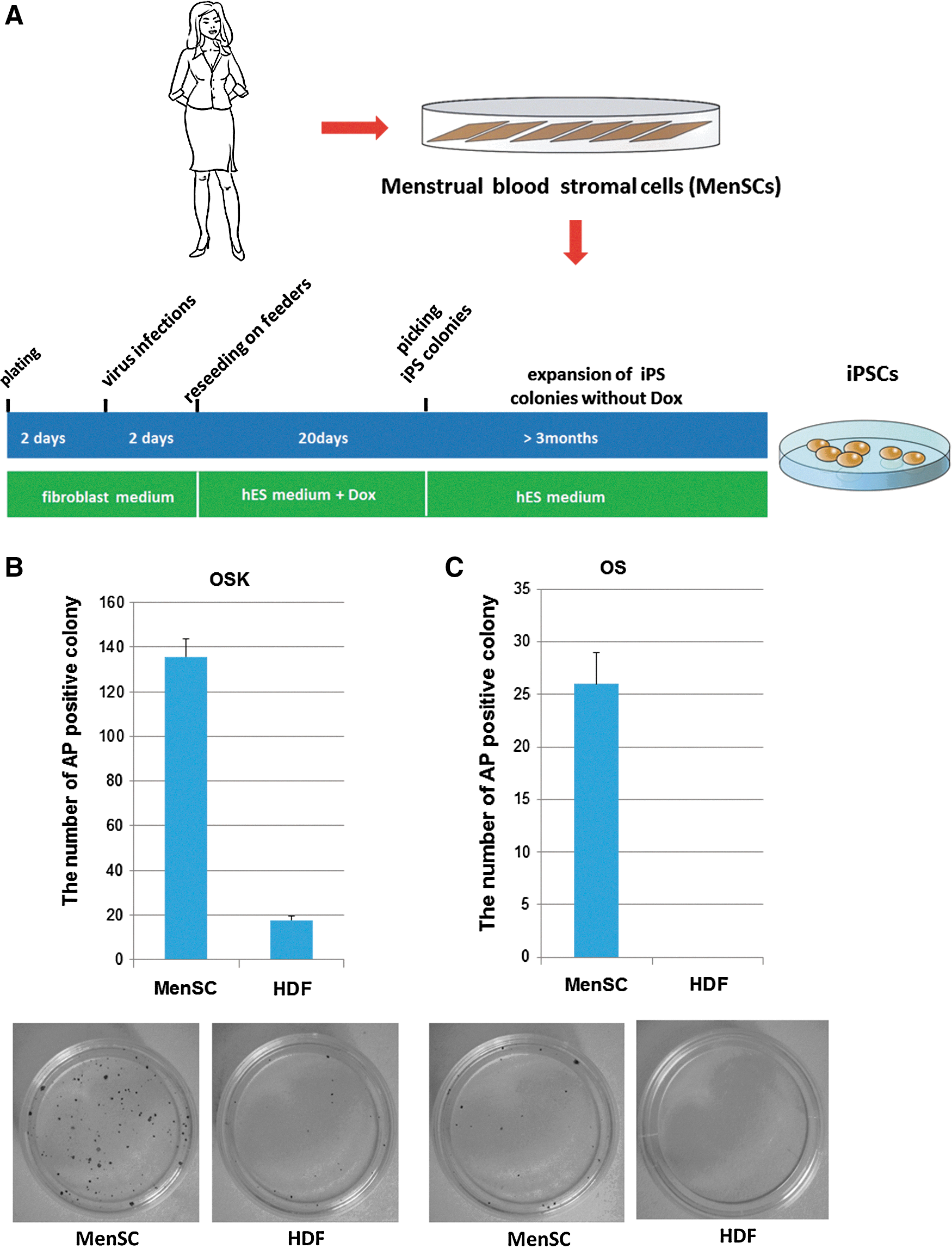
Induction of induced pluripotent stem cells (iPSCs) from MenSCs. Flow diagram for the generation of human iPSCs from MenSCs by Dox-inducible lentiviral transduction
Morphology and expression of pluripotent genes in MenSC-iPSCs
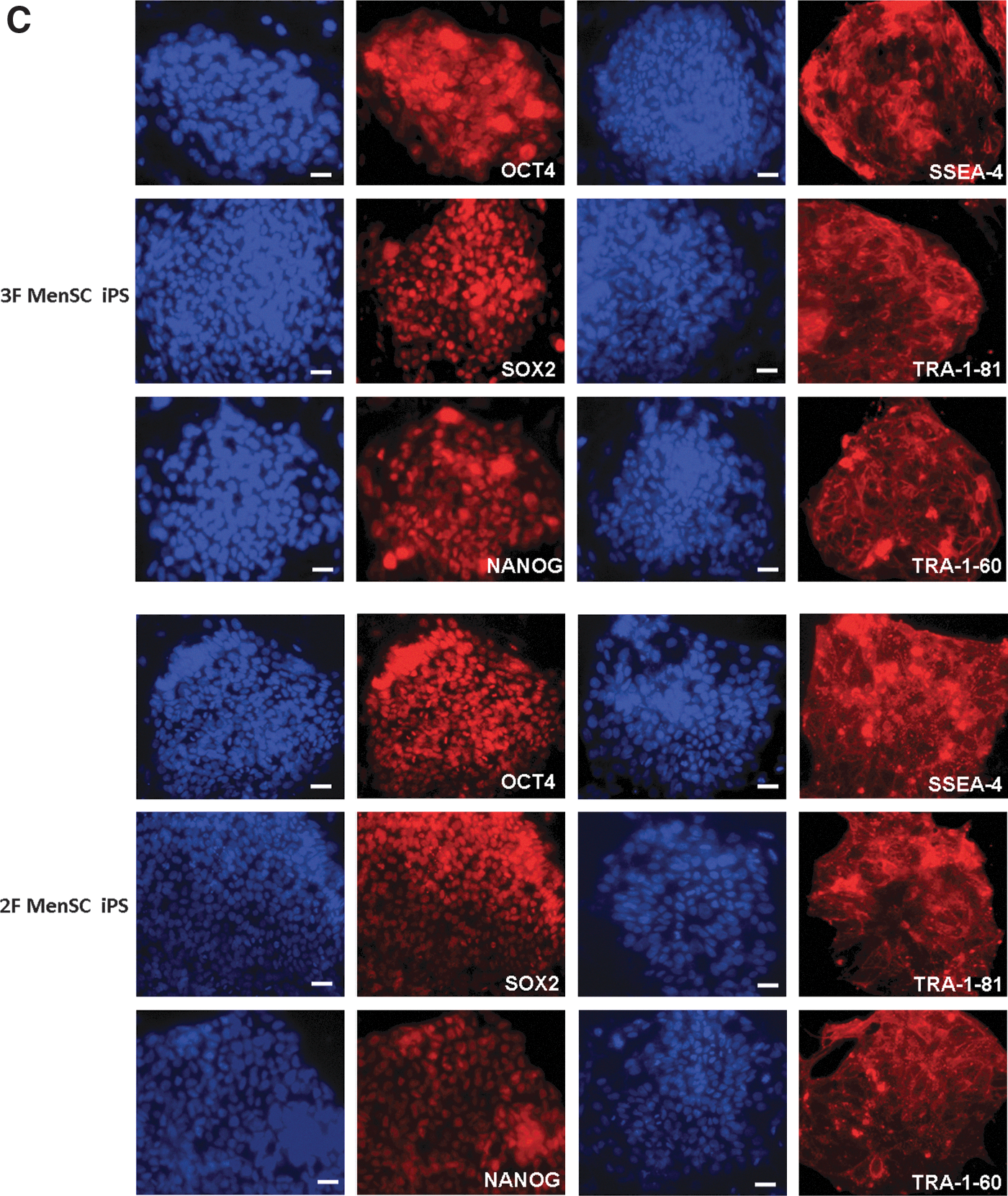
MenSC-iPSC colonies that were transduced with 3 factors (3F MenSC-iPS) or 2 factors (2F MenSC-iPS) were picked, cultured for several passages, and then checked for potential characteristics of human ESCs. These colonies exhibited typical human ESC-like morphology and stained positively for AP (Fig. 4A). Immunofluoresence staining confirmed the expression of pluripotency genes, including OCT4, SOX2, NANOG, SSEA-4, TRA-1-60, and TRA-1-81 (Fig. 4B), in these MenSC-iPSC colonies.
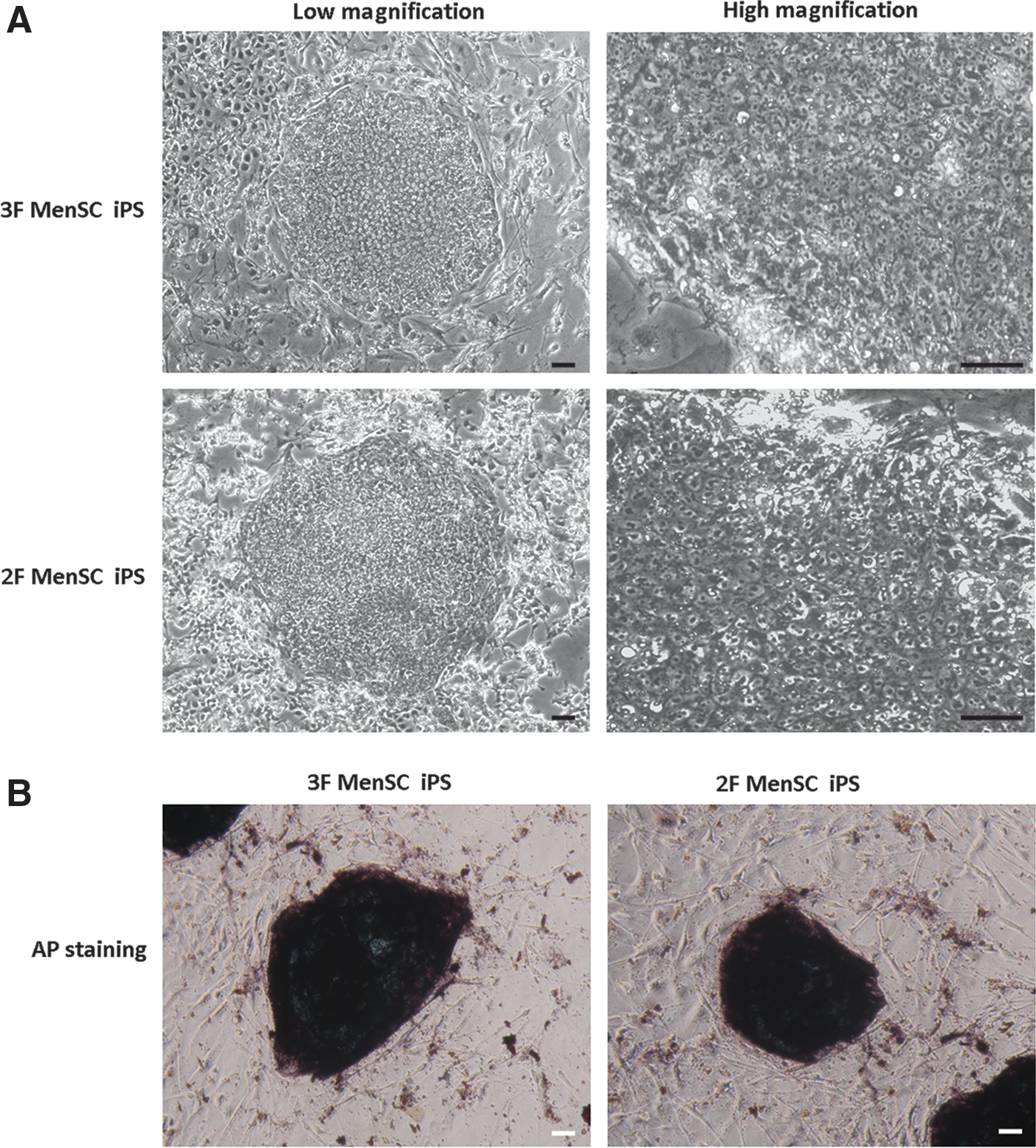
Characterization of MenSC-iPSCs. Morphology of 3F and 2F MenSC-iPSC colonies at low and high magnification
DNA microarray analysis
Global gene expression was analyzed among MenSCs, 3F and 2F MenSC-iPSCs, and H1 human ESCs using oligonucleotide microarrays to compare the gene expression profiles of MenSC-iPSCs and human ESCs. A hierarchical clustering analysis clearly indicated that 3F and 2F MenSC-iPSCs clustered closely with human ESCs and were distinct from parental MenSCs (Fig. 5A). Analysis of scatter plots further demonstrated that 3F and 2F MenSC-iPSCs are highly similar to human ESCs and hence completely different from MenSCs at the global transcriptional level (Fig. 5B).
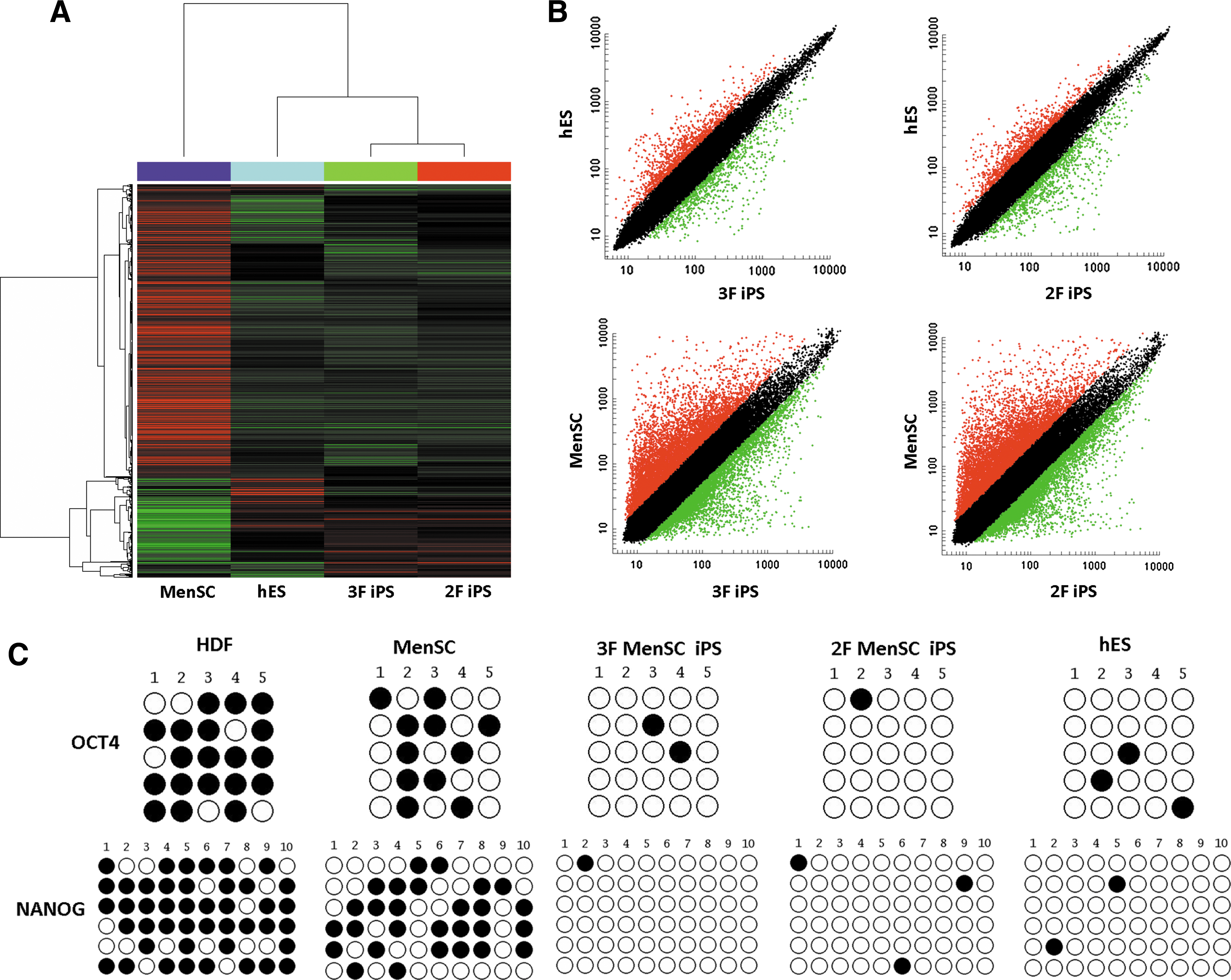
Gene expression profile and methylation analysis of the OCT4 and NANOG promoter regions of MenSC-iPSCs. Hierarchical cluster analysis of microarray data from MenSCs, MenSC-iPSCs, and human ESCs
Promoter methylation status of ESC-specific genes
To confirm epigenetic remodeling in reprogrammed cells, we also evaluated the methylation status of CpG dinucleotides in the OCT4 and NANOG promoters. Bisulfite genomic sequencing analysis showed that the OCT4 and NANOG promoter regions were demethylated in MenSC-iPSCs relative to the parental MenSCs and were thus similar to human ESCs. We also found that both promoter regions had slightly fewer methylated sites in MenSCs than in HDFs, which corresponds to the lower level of OCT4 and NANOG expression observed in MenSCs and not in HDFs (Fig. 5C).
Pluripotency of MenSC-iPS cells
To examine the differentiation potential of MenSC-iPSCs, we performed floating culture to demonstrate embryoid body formation. The embryoid bodies were transferred to gelatin-coated plates and cultured for an additional 8 days. The cells attached to the culture dish showed various types of cell morphology. Immunofluorescence staining was used to detect cells positive for β-III-tubulin (ectoderm), smooth muscle actin (mesoderm), and PDX-1 (endoderm) (Fig. 6A).
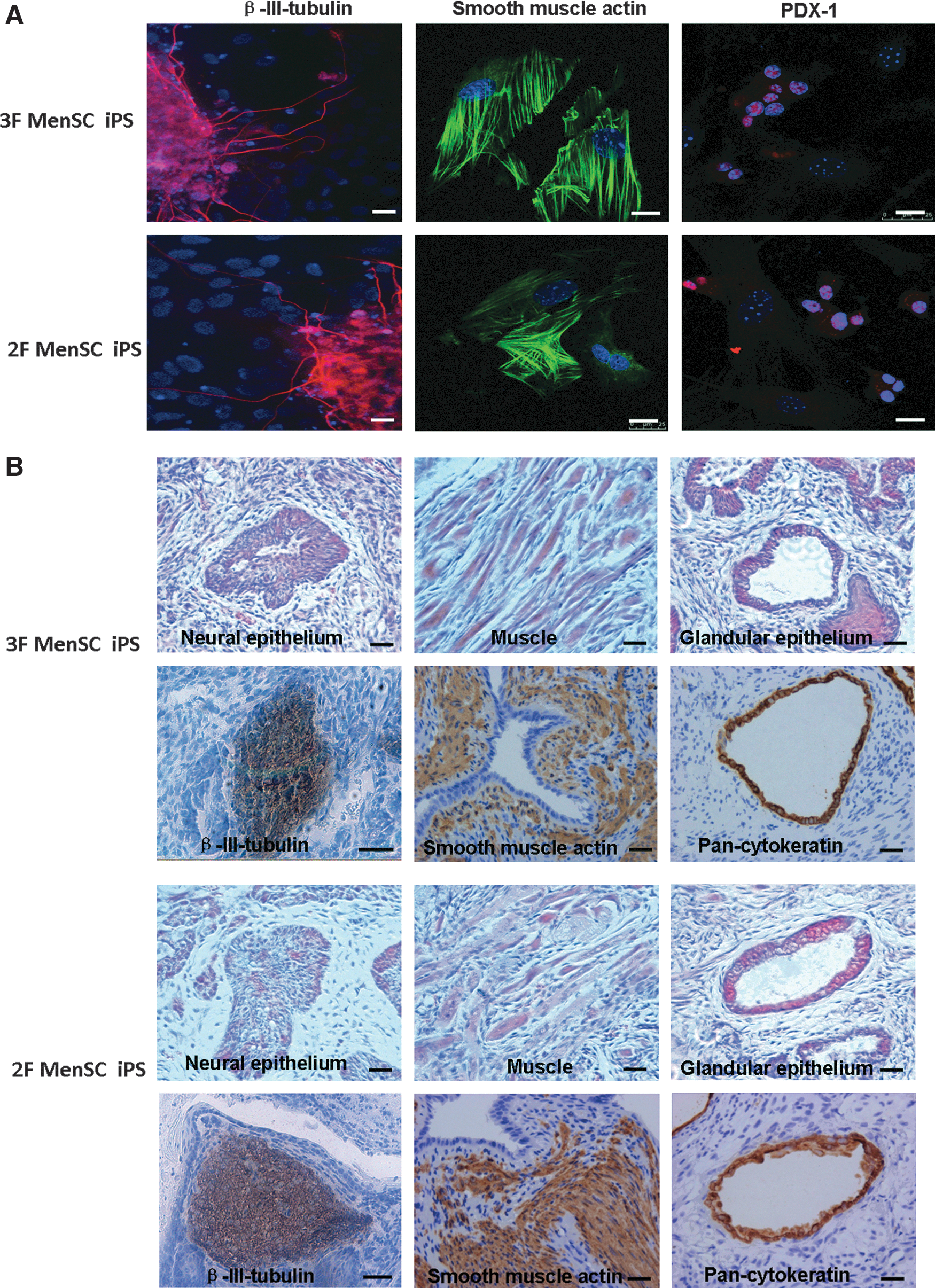
In vitro and in vivo differentiation of MenSC-iPSCs. Immunofluorescence staining showed differentiation of 3F and 2F MenSC–iPSCs into cells expressing markers of the 3 germ layers
To test for pluripotency in vivo, MenSC-iPSCs were transplanted into the hind limbs of SCID mice. Teratomas were found after 8 weeks after inoculation. Histological examination and immunohistochemistry staining revealed the presence of derivatives of all 3 germ layers, including neural epithelium, muscle, and glandular epithelium, within the tumors (Fig. 6B).
Normal karyotypes of MenSC-iPSCs
To assess genomic integrity, karyotyping was conducted on the iPSC population. Cytogenetic analysis showed a normal female karyotype for MenSC-iPSCs (Fig. 7). The results are consistent with previous reports, which showed that reprogramming using viral transduction does not alter the cell karyotype.
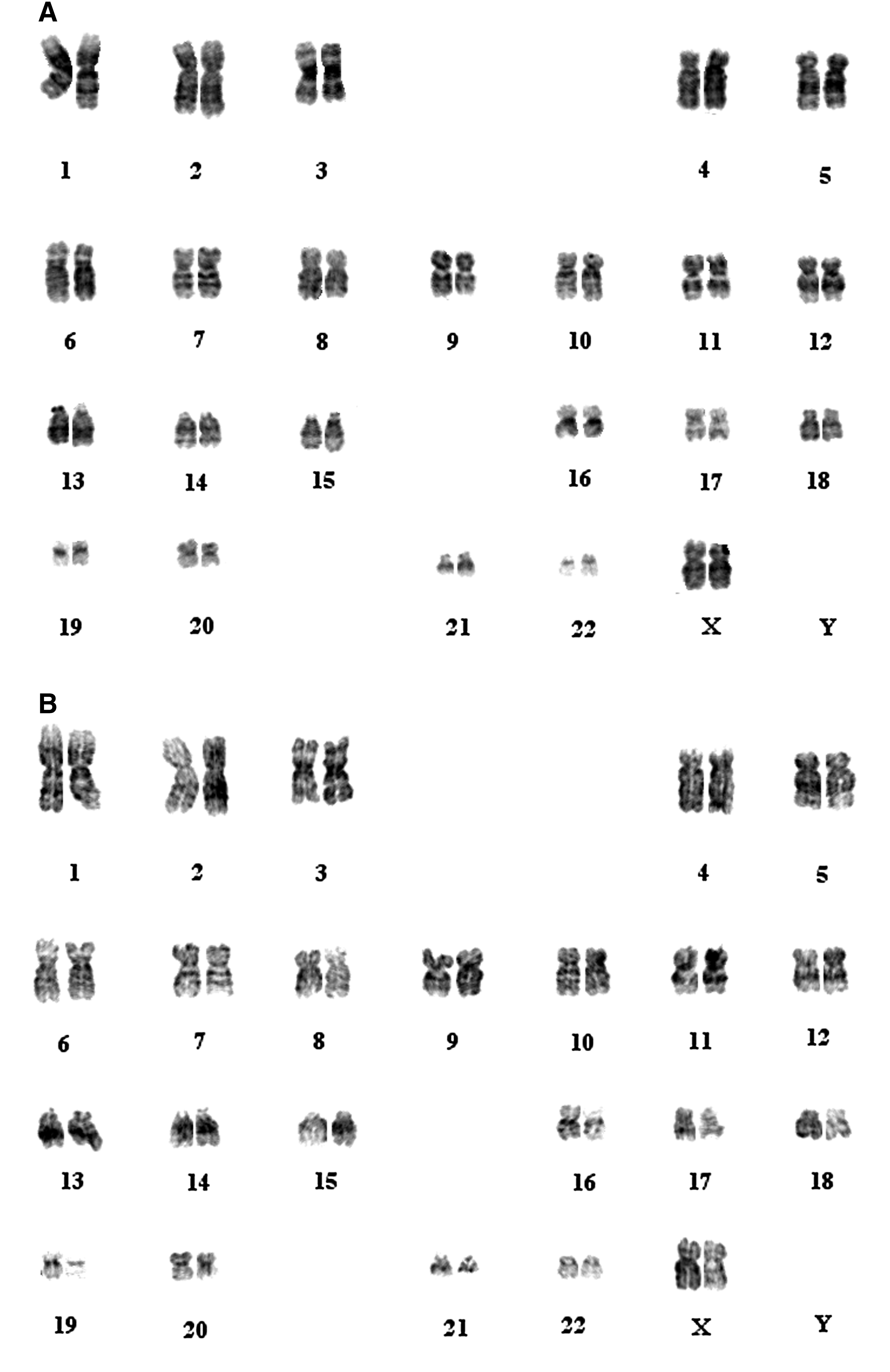
Karyotyping of MenSC-iPSCs. 3F MenSC-iPSCs at passage 21
Discussion
The iPSC technology holds great potential in regenerative medicine. To date, many different human cell types have been tested to be able to be reprogrammed into iPSCs. Here, a novel human cell source for iPSC production has been reported. In this study, we showed MenSCs to be negative for the expression of hematopoietic markers, such as CD34, CD45, and CD38, but positive for CD44, CD90, CD73, and CD105, which are generally considered as markers of mesenchymal stem cells [21]. SSEA-4, which is a marker used to characterize pluripotent stem cells [22], was also highly expressed in MenSCs. MenSCs did not express SOX2, and they expressed lower levels of OCT4, NANOG, and KLF4 than human ESCs. Additionally, the expression of c-MYC was similar to that of ESCs. Taking the results above into account, we developed strategies to reprogram MenSCs by overexpressing 3 factors (OCT4, SOX2, and KLF4) or 2 factors (OCT4 and SOX2) without the need for the oncogenes (i.e., c-MYC or KLF4). MenSCs and HDFs were transduced under the same experimental settings and infection protocols. Our results revealed that the 3 factors (OCT4, SOX2, and KLF4) can easily induce MenSCs into iPSCs at relatively high rates. While this manuscript was in preparation, Rodrigues et al. published findings in agreement with our study and demonstrated that menstrual blood-derived mesenchymal cells were reprogrammed with high efficiency when compared with HDFs through the forced ectopic expression of the 3 factors OCT4, SOX2, and KLF4 [23]. Compared with that study, we performed a more extensive investigation on this topic. In addition to the morphology, the expression of several pluripotent genes, and EB formation used by Rodrigues et al. to identify MenSCs-iPSCs, we also evaluated the global gene expression profile, DNA methylation status, and teratoma formation. Moreover, we found that the combination of OCT4 and SOX2 was sufficient to induce pluripotency in MenSCs, which was verified by the generation of AP-positive colonies post-transduction. Thus, these findings suggest that MenSCs can be easily reprogrammed into iPSCs, which may be partly attributed to their existing expression of pluripotent genes such as OCT4, NANOG, and SSEA-4, as these genes may facilitate reprogramming.
Even though many human cell types have been reprogrammed into iPSCs successfully, none of them demonstrate the complete combination of advantages that includes high efficiency, clinical-grade safety, and easy/simple accessibility. Human keratinocytes isolated from plucked hair can be rapidly and efficiently induced into iPSCs [12]. However, it remains unclear whether hair cells will be a faithful source for reprogramming, because they are directly exposed to UV radiation, which can result in mutagenesis. Additionally, human neural stem cells, which endogenously express SOX2, c-MYC, and KLF4, can be reprogrammed by OCT4 alone [13]. However, human neural stem cells are not an accessible source that can be easily isolated from patients. Reprogramming from human blood cells represents a novel way of establishing iPSCs from donor cells [24]. The procedure is minimally invasive and requires a small quantity of blood. However, the efficiency is low. Human iPSCs can also be obtained from urine, which is an even less-invasive and more convenient source of cells that can be obtained from men or women at essentially any time [17]. However, this process used to generate iPSCs still requires KLF4 or c-MYC, which is associated with oncogenesis. Generation of iPSCs with high efficiency and safety may be necessary for the clinical use of iPSCs in the future. Compared with the other cell types mentioned above, the utilization of human MenSCs as a potential source for reprogramming into iPSCs offers several advantages. First, MenSCs may be more easily reprogrammed than terminally differentiated fibroblasts. Second, the procedure for isolating MenSCs is relatively simple, fast, and safe, and does not pose any ethics concerns. Third, it is convenient to obtain a large quantity of MenSCs as the starting population for reprogramming. Fourth, because the reprogramming process requires only 2 factors, opportunities for insertional mutagenesis are minimized. Furthermore, obviating the requirement for KLF4 and c-MYC reduces the risk of inducing tumorigenesis. However, there is one obvious limitation for MenSCs in that they are only obtained from menstrual blood samples of women of reproductive age, which may narrow their applications. However, if iPSCs indeed have memory of the donor tissue [25], MenSCs-iPSCs should be the best candidate for producing MenSCs to treat uterus-related problems [18].
We thus demonstrated that stromal cells from human menstrual blood can be reprogrammed into iPSCs by 3 factors (OCT4, SOX2, and KLF4) or 2 factors (OCT4 and SOX2), which provide an ideal source for iPSC research and potential application. Further questions will be addressed such as whether other approaches, including adenoviruses [26], recombinant proteins [27], episomal vectors [28], chemicals [29], microRNAs [30], or synthetic mRNAs [31], can be used for MenSC reprogramming to increase safety, as these approaches are integration free and completely avoid the risk of insertional mutagenesis, which make them preferable for future patient-specific cellular therapies when compared with the lentiviral method used in the study.
Footnotes
Acknowledgments
The authors thank Dr. Shiying Jin for critical reading of the manuscript. This study was financially supported by the Ministry of Science and Technology of China (2011CB966200), the National Natural Science Foundation of China (NSFC) projects (31000653, 31101060 and 31171417), and the Beijing Natural Science Foundation project (5112019), the Ph.D. Programs Foundation of Ministry of Education of China (20100001120044), the General Program of Science and Technology Development Project of Beijing Municipal Education Commission of China (11220129).
Author Disclosure Statement
There are no commercial associations that might create a conflict of interest in connection with this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.