
Abstract
Notch signaling plays critical roles in stem cells by regulating cell fate determination and differentiation. The aim of this study was to evaluate the participation of Notch signaling in neurogenic commitment of human periodontal ligament-derived mesenchymal stem cells (hPDLSCs) and to examine the ability to control differentiation of these cells using modified surfaces containing affinity immobilized Notch ligands. Neurogenic induction of hPDLSCs was performed via neurosphere formation. Cells were aggregated and form spheres as early 1 day in culture. In addition, the induced cells exhibited increased mRNA and protein expression of neuronal markers that is, β3-tubulin and neurofilament. During neuronal differentiation, a significant increase of Hes1 and Hey1 mRNA expression was noted. Using pharmacological inhibition (γ-secretase inhibitor) or genetic manipulation (overexpression of dominant negative mastermind-like transcription co-activators), neurosphere formation was attenuated and a marked decrease in neurogenic mRNA expression was observed. To confirm the role of Notch signaling in neuronal differentiation of hPDLSCs, the Notch ligand, Jagged-1, is bound to the surface using an affinity immobilization technique. The hPDLSC cultured on a Jagged-1-modified surface had increased expression of Notch signaling target genes, Hes-1 and Hey-1, confirming the activity and potency of surface-bound Jagged-1. Further, hPDLSC on surface-bound Jagged-1 under serum-free conditions showed multiple long and thin neurite-like extensions, and an increase in the expression of neurogenic mRNA markers was observed. Pretreatment of the cells with γ-secretase inhibitor, DAPT, before seeding on the Jagged-1-modified surface blocked development of the neurite-like morphology. Together, the results in this study suggest the involvement of Notch signaling in neurogenic commitment of hPDLSCs.
Introduction
Regarding neuronal differentiation, our group and other investigators have illustrated that hPDLSCs were able to differentiate toward neurogenic lineage under specific conditions [4 –7]. Neurite-like, long cellular processes and upregulation of neurogenic markers [β3-tubulin, nestin, and neurofilament (NF)] were noted after neurogenic induction [4,8]. Moreover, it has been demonstrated that hPDLSCs were able to integrate into brain tissues in vitro, evaluated by hippocampal organ culture model [9]. In addition to direct differentiation into neurons, hPDLSCs promoted survival, neurite outgrowth, and migration of neuronal cell line by secreting nerve growth factor [10]. Thus, hPDLSCs have been proposed as a candidate cell source for neurological regenerative treatment as they are capable of direct differentiation into neuron and indirect support nervous tissue regeneration.
Notch signaling has a critical role in maintaining stemness and directing differentiation in several types of stem cells [11 –13]. Notch signaling is activated upon the binding of Notch to its ligands on adjacent cells [14]. Subsequently, the cleavage of Notch protein occurs intracellularly, resulting in the translocation of Notch intracellular domain (NICD) into the nucleus and further regulating Notch target gene expression [14]. In periodontal ligament cells, Notch signaling was shown to participate in osteogenic differentiation process and induction of osteoclastogenesis [4,15,16]. However, evidence of Notch signaling in hPDLSCs differentiation toward neuronal lineage is yet unknown. Therefore, the aim of this study was to evaluate the role of Notch signaling during neurogenic differentiation of hPDLSCs in vitro. Here, we reported that attenuation of Notch signaling resulted in the decrease expression of neurogenic marker, while the activation of Notch signaling under serum-free condition promoted neurogenic phenotypes.
Materials and Methods
Culture and neurogenic differentiation hPDLSCs
hPDLSCs were isolated and cultured according to formerly published procedure [17]. The protocol for the isolation of hPDLSCs was previously approved by the Ethical Committee, Faculty of Dentistry, Chulalongkorn University. hPDLSCs were maintained in Dulbecco's modified Eagle medium (Gibco) containing 10% fetal bovine serum (Gibco), 2 mM
Neurogenic differentiation of hPDLSCs was performed using the sphere formation technique [18]. Briefly, hPDLSCs were cultured in neurobasal medium (Gibco) containing B27 (2%),
The percentage of neurospheres with given diameters was calculated as follows: group 1: spheres with a diameter <50 μm, group 2: spheres with a diameter 50–100 μm, and group 3: spheres with a diameter >100 μm.
To determine sphere formation efficiency, limited dilution assay was employed. Cells were seeded in 60 mm low attachment Petri dishes at various density (from 10,000–100,000 cells/plates) and maintained in neurogenic induction medium for 7 days. The number of sphere formation was evaluated under phase-contrast microscope.
For calcium imaging, cells were loaded with Fluo-4 AM dye (Invitrogen) for 30 min at room temperature according to manufacturing protocol. Cell were washed twice and maintained in calcium-free culture medium. Cells were stimulated with N-methyl-
Cell viability assay
Live/dead fluorescence staining and MTT assay were employed to evaluated cell viability. For live/dead fluorescence staining, calcein acetoxymethyl ester, and ethidium homodimer-1 were added into the culture and incubated at 37°C for 20 min and subsequently observed under a fluorescent microscope using Apotome.2 apparatus (Carl Zeiss). Live cells generated green fluorescence and dead cells exhibited red fluorescence. For flow cytometry analysis, a single-cell suspension was obtained from spheres using trypsin dissociation. Cell suspension was allowed to recover in culture medium for 30 min before staining with live/dead fluorescence. MTT assay was performed according our previous publication [19].
Transfection and retroviral transduction
The retroviral plasmid for expression of dominant negative mastermind-like transcriptional factor (DN-MAML; MSCV-Mam(12–74)-EGFP), a kind gift from Dr. Warren Pear (University of Pennsylvania) or the control vector (MSCV-IRES-GFP, Addgene plasmid 20672), was co-transfected with packaging construct pCL-Ampho (Imagenex) into HEK293 cells using the FuGene® HD transfection reagent (Roche) according to the manufacturer's instructions. Culture supernatants containing retroviruses were harvested twice at 48 and 72 h after transfection and were used to transduce hPDLSCs. Transduction efficiency was confirmed by fluorescent microscopy.
Immobilization of Jagged-1
Surface-bound Notch ligand, Jagged-1, was fabricated by an indirect affinity immobilization method previously reported by our group [4,20,21]. Briefly, recombinant protein G (50 μg/mL; Zymed) was incubated with the tissue culture plate surface for 16 h and subsequently, incubated with bovine serum albumin (10 mg/mL; Sigma) for 2 h. The surface was further incubated with recombinant human Jagged-1 (10 μM; R&D Systems) for 2 h. The surface was washed 3 times with sterile phosphate-buffered saline (PBS) between each step. An equal amount of human immunoglobulin G Fc fragment (Jackson Immunoresearch Laboratory) was employed as a control. Before seeding the cells, the surfaces were washed once with culture media. The presence of immobilized Jagged-1 was confirmed using enzyme-linked immunosorbent assay (ELISA) according to previously publish protocol [4,20,21].
Polymerase chain reaction
For RT-PCR, total cellular RNA was extracted with Trizol reagent. RNA samples (1 μg) were converted to cDNA by reverse transcriptase enzyme (Promega). RT-PCR was performed using Taq polymerase (Invitrogen). The amplified DNA was then electrophoresed on a 1.8% agarose gel and visualized by ethidium bromide staining. In some experiments, the band density was determined using ImageJ software. For real-time quantitative PCR experiment, PCR was performed in a LightCycler® Nano (Roche) with LightCycler480 SYBR Green I Master kit (Roche Diagnostic). Reaction product was quantified with GAPDH as the reference gene. The primer sequences were shown in Supplementary Table S1 (Supplementary Data are available online at
PCR array
Total RNA was extracted using RNeasy Mini kit (Qiagen). The contaminating DNA was digested by RNase-free DNase I (Qiagen). One microgram of total RNA was employed to synthesize first-strand cDNA using RT2 First Strand kit (Qiagen). Human Notch Signaling Pathway PCR array (PAHS-059; Qiagen) was performed with real-time PCR using RT2qPCR Master Mixes (Qiagen) in LightCycler480 (Roche). The data were analyzed using RT2 Profiler™ PCR array data analysis software.
Immunocytochemistry
The cells were fixed in 10% buffered formalin at room temperature for 30 min, permeabilized with 0.15% Triton®-X100 in PBS and 10% horse serum for 1 h at room temperature. The cells were then incubated with mouse anti-β3-tubulin (Promega) or mouse anti-NF, medium chain (NF) (Invitrogen) at a 1:200 dilution for 18 h at room temperature. After washing with PBS, the cells were incubated with biotinylated rabbit anti-mouse antibody (Zymed) for 30 min at 1:500 dilution. Subsequently, the antibody was detected using Strep-Fluorescein isothiocyanate (Sigma) or Rhodamine Red™-X (Invitrogen) and the nuclei were counterstained with 4′,6-diamidino-2-phenylindole (0.1 μg/mL). The cells were analyzed with a fluorescent microscope using Apotome.2 apparatus (Carl Zeiss).
Statistical analyses
Data are reported as mean±standard deviation. Statistical analyses were performed using 2-independent Student t test for 2-group comparison. A one-way analysis of variance followed by Dunnett test was employed to compare in experiments containing 3 or more groups. Differences at P<0.05 were considered to be statistically significant.
Results
hPDLSCs differentiated into neuronal linage
Neurosphere formation assay was employed to evaluate neurogenic differentiation ability of hPDLSCs. Similar to our previous reports [4], neurosphere formation was noted at day 1 after culturing cells in neurogenic medium (Fig. 1A). Size and density of the spheres were increased at day 3 and 7 compared to day 1 (Fig. 1B–D). However, the efficiency of the sphere formation was 2–4 spheres per 10,000 cells at day 7. After 7 days, the spheres were transferred to collagen IV coated dishes and maintained in neurogenic medium supplemented with retinoic acid for another 7 days. Cells migrated out from the spheres and exhibited neurite-like cellular processes (data not shown). β3-tubulin and NF protein expression were observed in the floating and adhered spheres (Fig. 1E–H). More cells expressed β3-tubulin than expressed NF in both floating and adhered conditions. The NF expression was noted in cells located at the out border of the spheres. Cell viability in floating and adhered condition was evaluated using live/dead fluorescence (calcein acetoxymethyl ester and ethidium homodimer-1) staining. The results illustrated that most of the cells were positively stained with green fluorescence, indicating cell survival (Fig. 1I, J). In addition, neurogenic differentiation was confirmed by the mRNA expression of neurogenic markers. The mRNA expression of Sox2 and Sox9 was significantly upregulated in floating spheres (Fig. 1K–P). For adhered spheres, it was also noted that Sox2 and Sox9 mRNA levels were decreased, while β3-tubulin and NF was increased when compared to the floating spheres. Moreover, the upregulation of GABA receptor β3 was noted in both sphere and adhered condition. Intracellular calcium imaging was performed comparing calcium at baseline and after treatment with NMDA or DPH (Fig. 1Q). The results showed that levels of intracellular calcium were increased immediately after cells were exposed with the stimuli, implying neuronal function. The intracellular calcium was decreased to baseline within 30 s after stimulation. Together, these results suggest that hPDLSCs were able to differentiate into the neurogenic lineage.
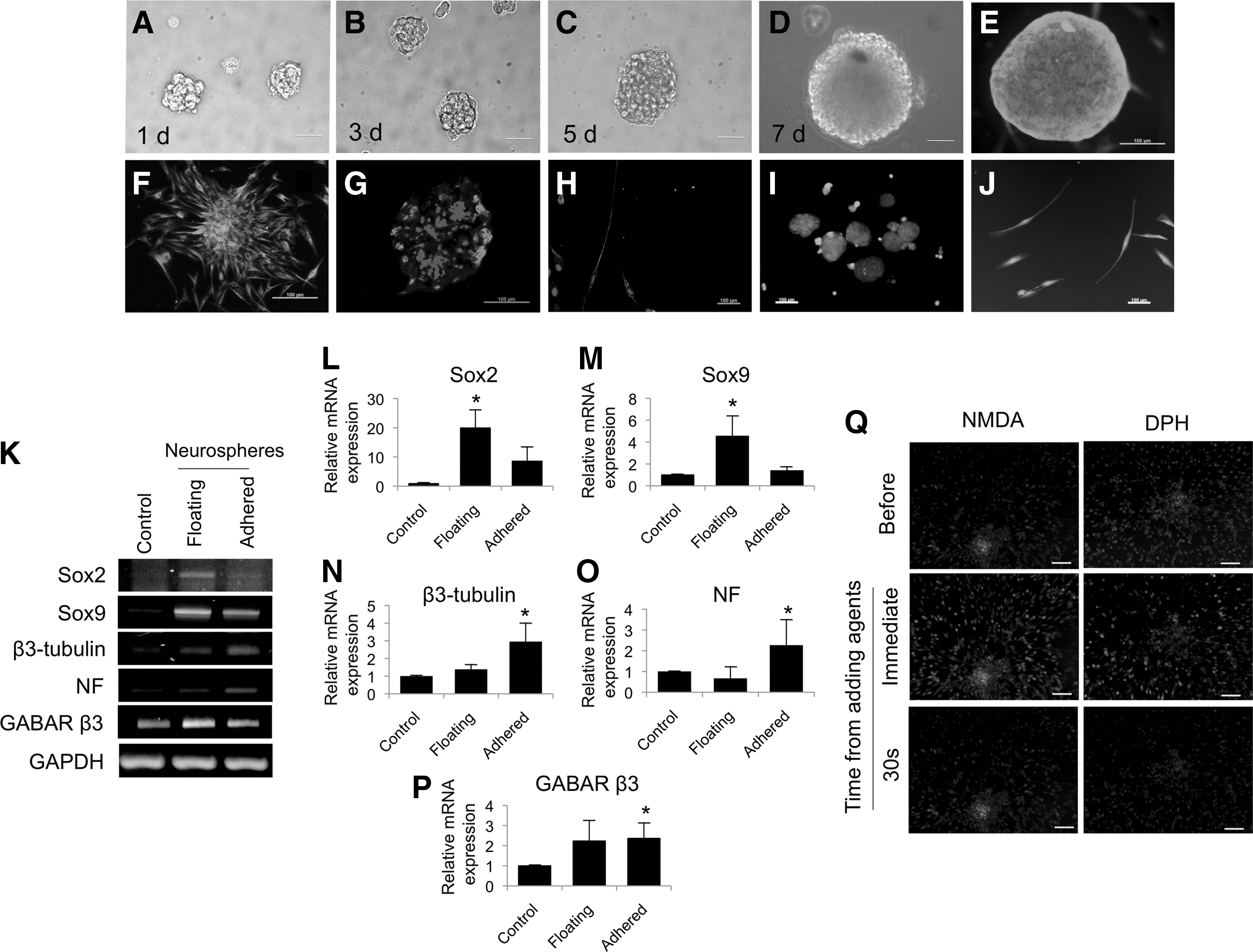
Human periodontal ligament-derived mesenchymal stem cells (hPDLSCs) were able to differentiate into neurogenic lineage. Neurosphere formation was observed at day 1
Notch signaling involved in neuronal differentiation of hPDLSCs
To examine a possible role for Notch signaling in neuronal differentiation of hPDLSCs, PCR array of human Notch signaling was performed to compare mRNA expression between control hPDLSCs those undergoing neurogenic differentiation. Thirteen genes were upregulated (data not shown). Among those, 2 Notch signaling target genes, Hes1 and Hey1, were noted. The upregulation of Hes1 and Hey1 mRNA expression was confirmed by RT-PCR and real-time quantitative PCR (Fig. 2A–C). Hes1 and Hey1 mRNA levels were higher in neurospheres than in controls. In time course experiments, Hes1 and Hey1 mRNA levels increased upon culturing cells in neurogenic medium as early as 1 day compared to the control. Both Hes1 and Hey1 mRNA expression tended to decrease at later time points, although the expression was still relatively higher than the control (Fig. 2D–F). In addition, the decrease of Hes1 and Hey1 mRNA levels was also noted in adhered neurosphere-derived neuronal cells compared to floating spheres (data not shown), suggesting the reduction of Notch signaling may required in neuronal maturation process. Together, the upregulation of Notch signaling target genes upon neurogenic differentiation suggest a role of Notch signaling in neurogenic commitment process in hPDLSCs.
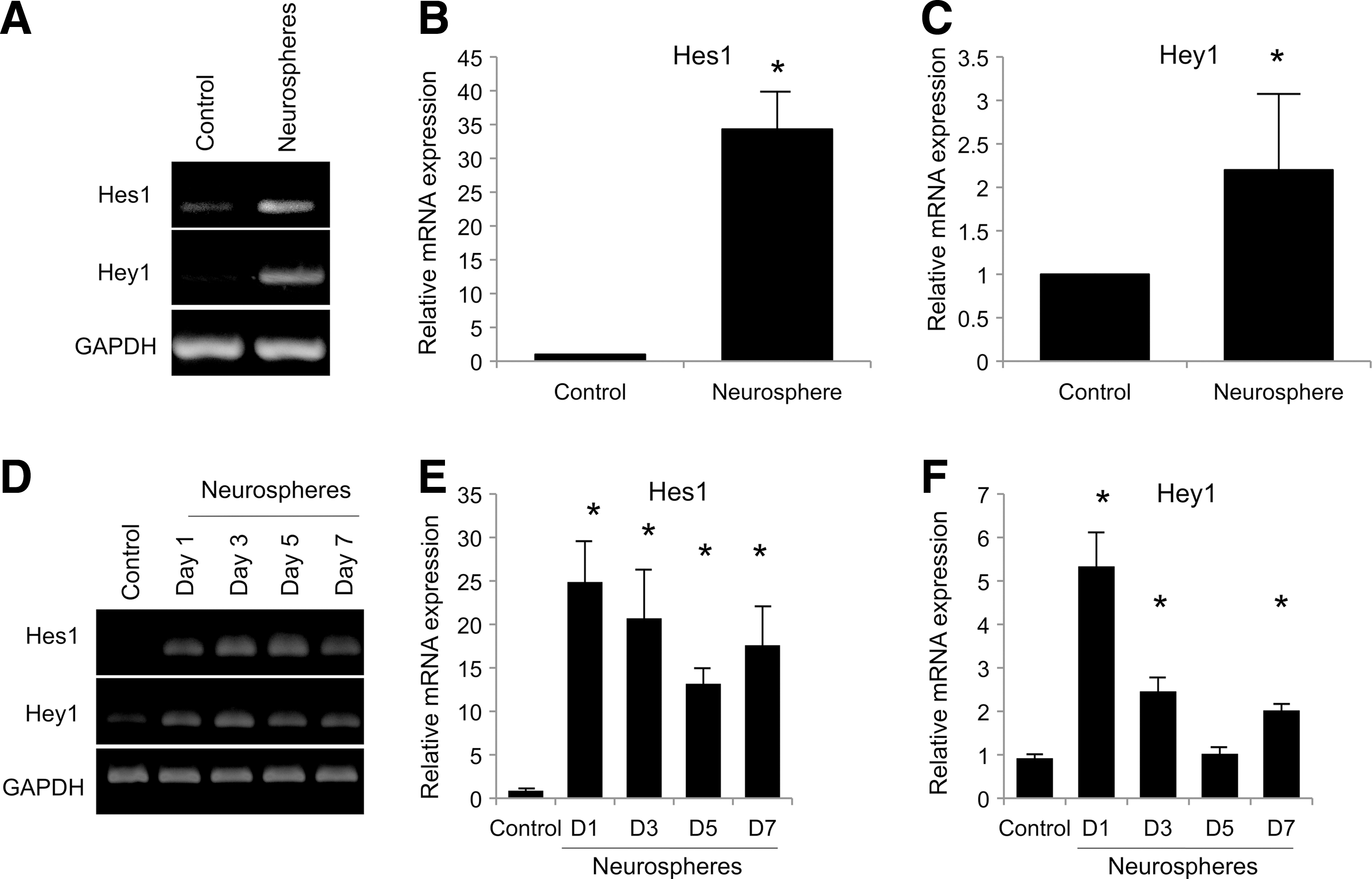
hPDLSCs expressed Notch signaling target genes during neurogenic differentiation. The Hes1 and Hey1 mRNA levels were significantly upregulated after neurogenic induction for 7 days
To further confirm the involvement of Notch signaling in the cell fate decision toward the neurogenic lineage, a γ-secretase inhibitor, DAPT, was applied to inhibit Notch signaling initiation by impeding the cleavage of NICD. In the presence of DAPT, the spheres appeared smaller than the control (Fig. 3A–D). An average sphere diameter was decreased when cells were exposed to DAPT compared to control cells, but a statistical significant difference was not observed (Fig. 3E). A higher percentage of small-size neurospheres (<100 μm) was observed with the addition of DAPT compared to the control at day 1 and 7 (Fig. 3F). Cell viability was not significantly different between the control and DAPT treated group as determined by flow cytometry and MTT analysis (Fig. 3G, H, respectively). At day 1, the DAPT-treated neurospheres had decreased Hes1 and Hey1 mRNA expression, confirming the inhibition of Notch signaling (Fig. 3I, J). Correspondingly, Sox2 and β3-tubulin mRNA levels were decreased by treatment of the neurospheres with DAPT (Fig. 3I, J). In addition, overexpression of DN-MAML was employed. MAML is transcriptional co-activator and essential participant in Notch signaling. The DN-MAML overexpressed hPDLSCs resulted in a decrease of neurosphere's size at 1 day in neurogenic culture condition compared to the vector control transduced cells (Fig. 4A, B). Cell viability was not significantly different between the GFP vector control and DN-MAML group (Fig. 4C). In addition, a decrease in mRNA expression levels of Notch signaling target genes (Hes-1 and Hey-1) as well as neurogenic marker genes (Sox2 and β3-tubulin) was noted (Fig. 4D, E). Together, the data imply the involvement of Notch signaling in neuronal commitment of hPDLSCs.
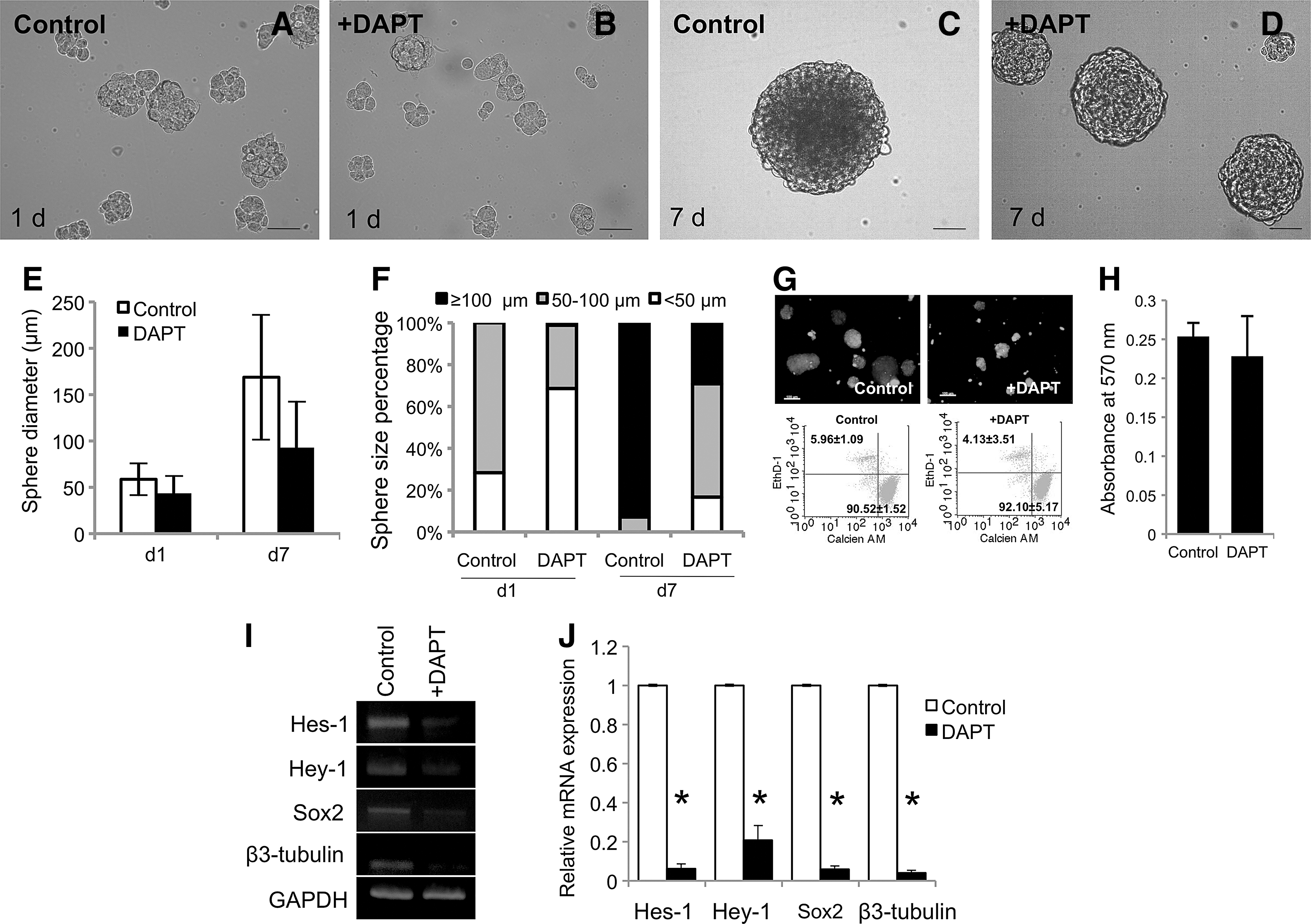
Pharmacological inhibition of Notch signaling attenuated neurosphere formation by hPDLSCs. A γ-secretase inhibitior (DAPT) was employed to inhibit the initiation of Notch signaling. Cells treated with DAPT resulted in small-sized neurospheres as noted at 1
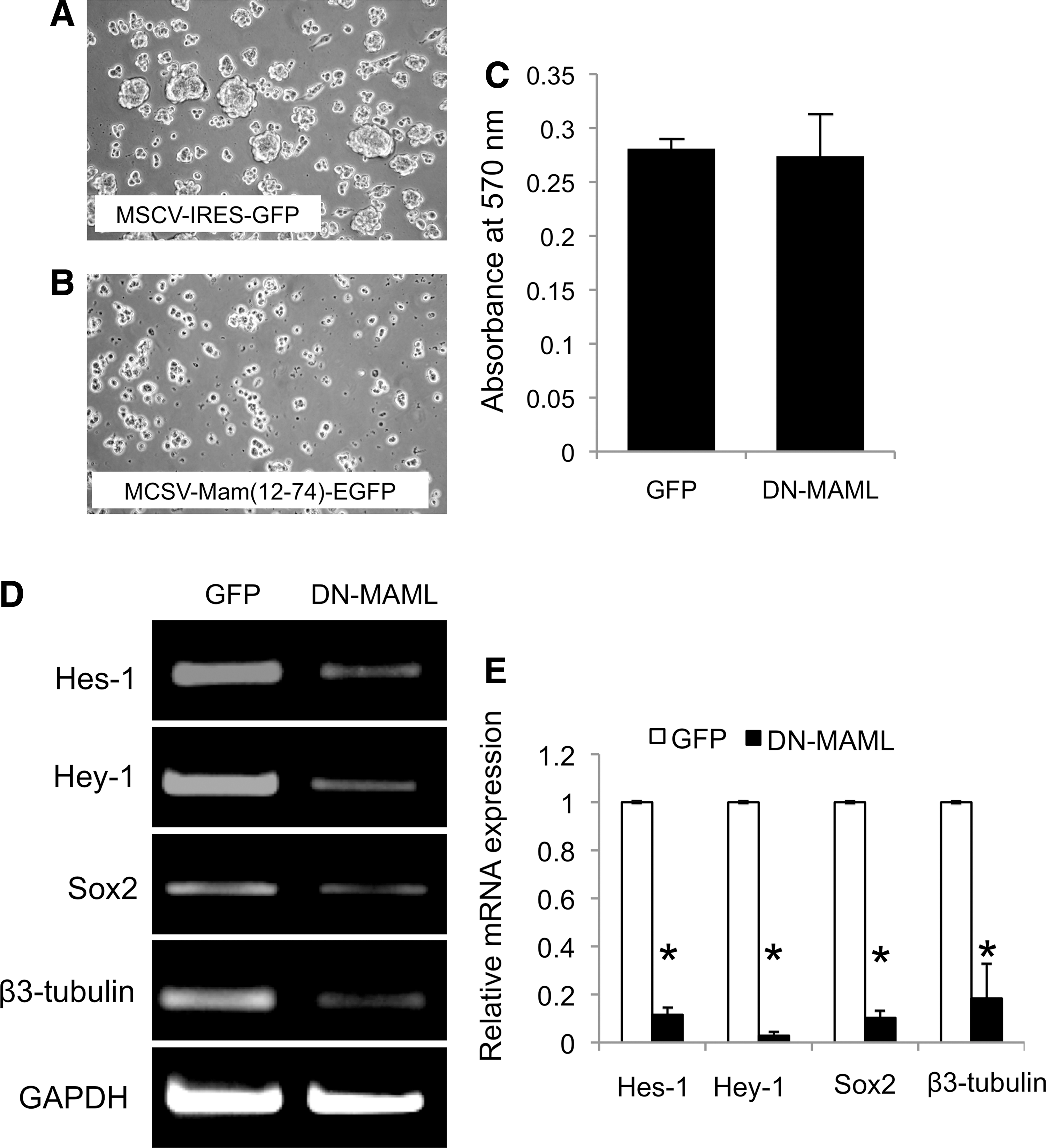
The suppression of Notch signaling by genetic manipulation impaired neurosphere formation by hPDLSCs. Overexpression of dominant negative mastermind-like transcriptional factor (DN-MAML) in hPDLSCs resulted in the reduction of neurosphere formation compared to those transduced with the control vector
Surface-bound Jagged-1 induces neuronal differentiation in hPDLSCs.
From PCR array analysis, Notch ligand, Jagged-1, was upregulated in neurospheres compared to the control (data not shown). The increased mRNA level of Jagged-1 was confirmed by real-time quantitative PCR (Fig. 5A). This result suggests a role of Jagged-1 in neurogenic commitment of hPDLSCs. We, therefore, utilized a Jagged-1-immobilized surface to further elucidate the role of Notch signaling in neurogenic commitment by hPDLSCs. The presence of bound Jagged-1 on the surface was confirmed by ELISA assay (Fig. 5B). The hPDLSCs seeded on the Jagged-1-immobilized surface exhibited a dramatic increase of Notch target gene (Hes-1 and Hey-1) expression compared to those exposed to normal culture surface and hFc bound surface (Fig. 5C, D). Moreover, the induction of Notch signaling by the Jagged-1-immobilized surface was inhibited by the addition of DAPT in culture medium, confirming the Notch signaling ability of the Jagged-1-bound surface.
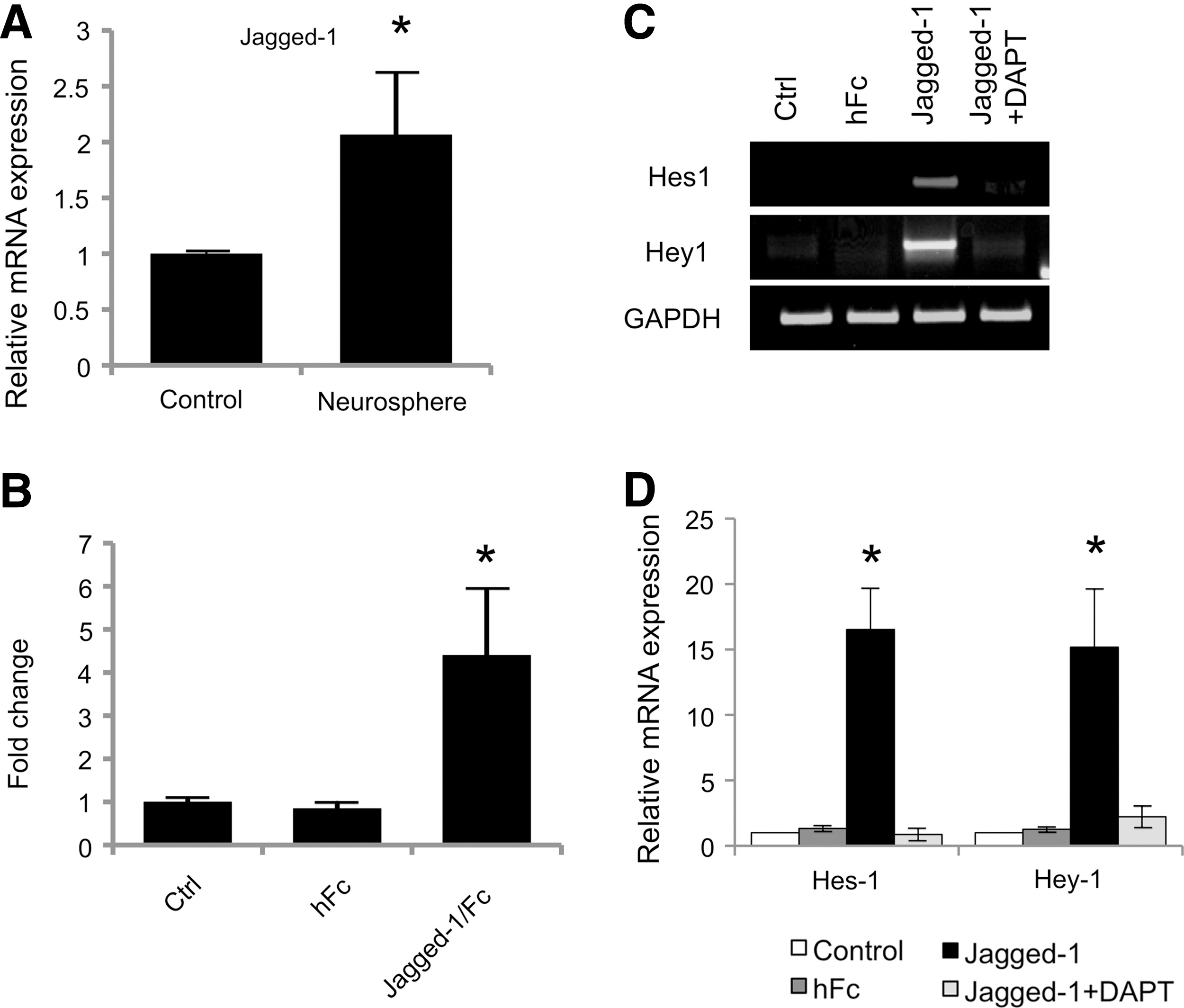
Surfaced-bound Jagged-1 initiated Notch signaling in hPDLSCs. The Jagged-1 mRNA expression was significantly increased after culturing cells in neurogenic medium for 7 days, as evaluated by real-time PCR
To examine the role of Jagged-1 on neurogenic differentiation of hPDLSCs, the cells were seeded on a Jagged-1-modified surface in serum-free condition for 72 h. These cells exhibited multiple long, thin neurite-like extensions in both 1 and 10 nM concentration of Jagged-1 (Fig. 6C, D). On the contrary, hPDLSCs on untreated and bound Fc control surfaces had a fibroblast-like morphology (Fig. 6A, B). By immunocytochemical analyses, cells seeded on Jagged-1-bound surface were also positively stained with an antibody to β3-tubulin (Fig. 6G, H). However, β3-tubulin positive cells were not observed on either of the control surfaces (Fig. 6E, F). Moreover, an increase in the expression of neurogenic mRNA markers, Sox2 and β3-tubulin, was observed in cells seeded on surface-bound Jagged-1 in a dose-dependent manner (Fig. 6I, J). These data indicate that Jagged-1 regulates neuronal differentiation of hPDLSCs. In addition, pretreatment of the cells with DAPT before seeding on the Jagged-1-modified surface blocked development of the neurite-like morphology and β3-tubulin protein expression (Fig. 7C, G, respectively). Moreover, the cells treated with soluble Jagged-1 displayed similar morphology and β3-tubulin protein expression to those on untreated surface and bound Fc surface (Fig. 7D, H, respectively). Sox2 and β3-tubulin mRNA expression were significant lower on DAPT pretreated cells on surface-bound Jagged-1- and soluble Jagged-1-treated cells compared to those on surface-bound Jagged-1 surface (Fig. 7I, J).
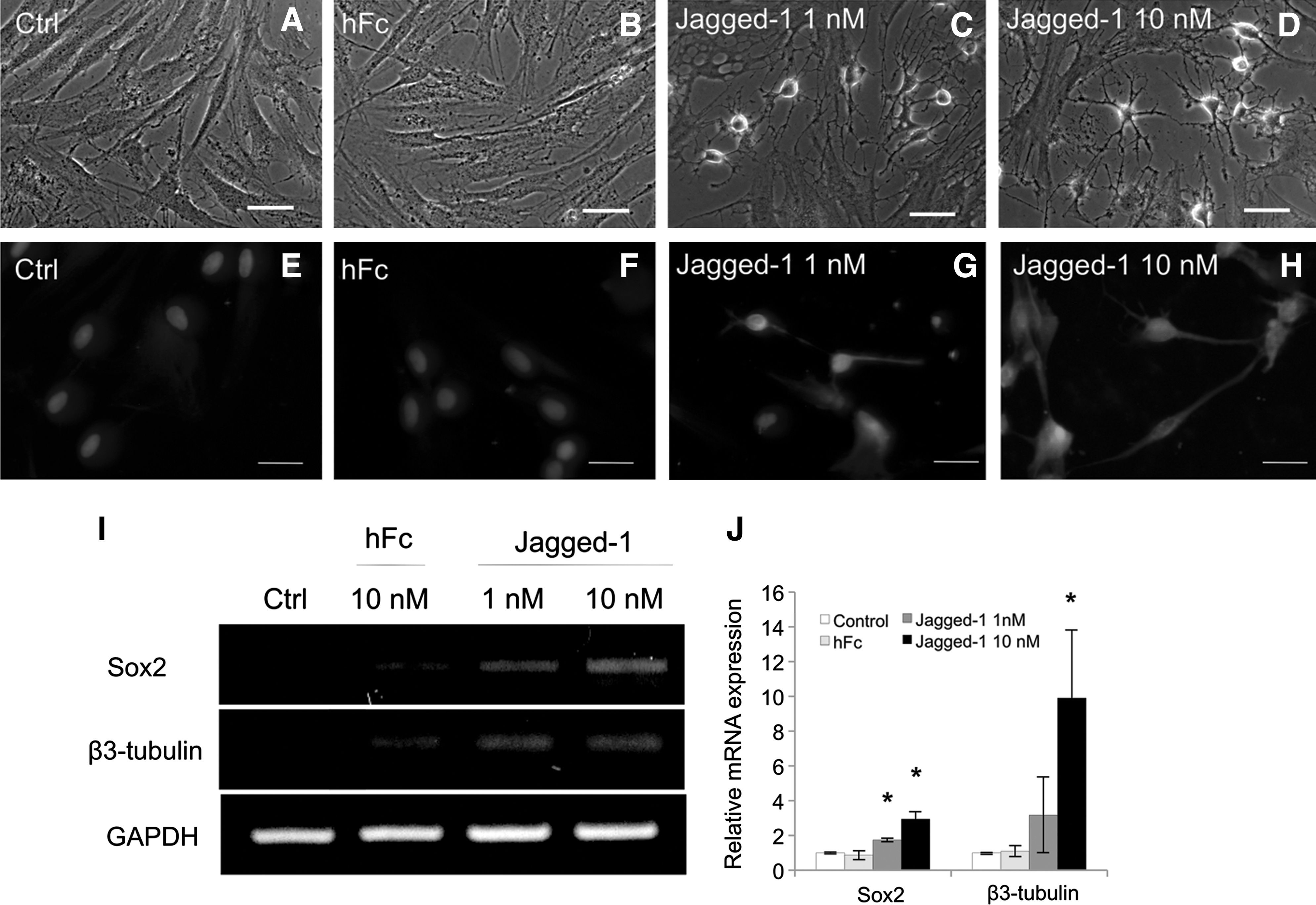
Surface-bound Jagged-1 initiated neuronal differentiation by hPDLSCs in serum-free condition. At 72 h, the hPDLSCs on untreated surface and bound Fc surface appeared fibroblast-like morphology
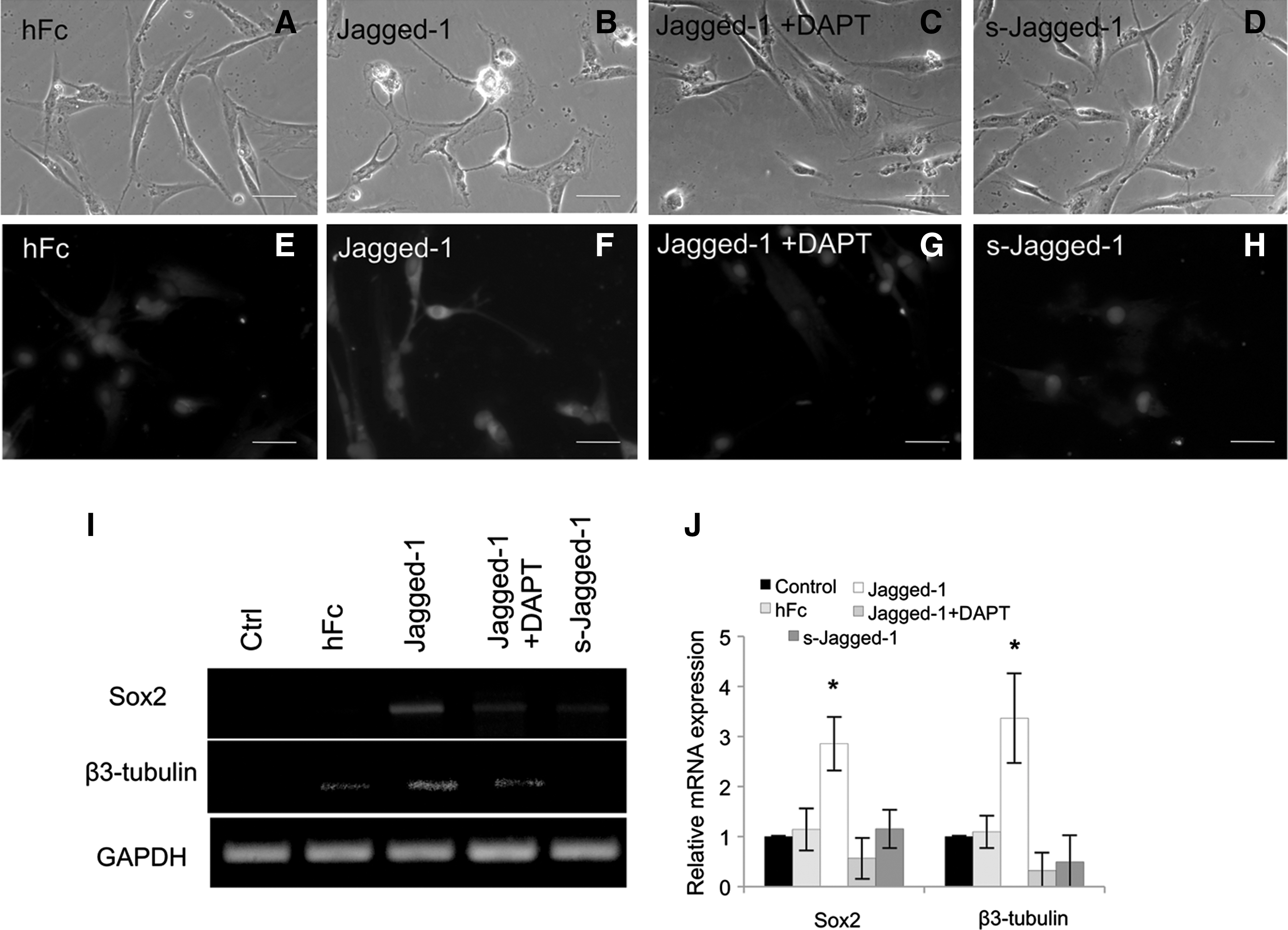
Surface-bound Jagged-1 initiated neuronal differentiation by hPDLSCs in serum-free condition. The hPDLSCs on the control bound Fc surface had a fibroblast-like morphology and were not stained with anti β3-tubulin antibody
Discussion
In the present study, we identified a role for Notch signaling on neurogenic commitment of hPDLSCs. The results showed that Hes-1 and Hey-1 were upregulated in hPDLSC-derived neurospheres. The inhibition of Notch signaling by either pharmacological or genetic approaches was able to attenuate neurosphere formation and neurogenic mRNA expression by hPDLSCs. Further, a transcriptional activity of the Notch/CSL complex was required during neurogenic commitment of hPDLSCs as the overexpression of DN-MAML attenuated this process. In addition, hPDLSCs exposed to a Jagged-1-immobilized surface exhibited neuronal-like morphology with an increase in neurogenic mRNA and protein expression. Together, these results suggest the involvement of Notch signaling in neuronal differentiation of hPDLSCs.
Upon induction, hPDLSCs were able to differentiate into neuronal-like cells. These induced cells exhibited neurite formation and expressed neurogenic markers in both mRNA and protein levels. In addition, calcium ions initiate intracellular signaling that correspond to neuronal function [22]. NMDA receptor and DPH receptor are expressed in neuronal cells. Thus, the change of intracellular calcium resulted from NMDA or DPH treatment in the present study, implying the expression of particular receptors and their function. Similar to the present study, neuronal differentiation of hPDLSCs has previously been reported [5,23]. Undifferentiated hPDLSCs expressed baseline mRNA levels of neurogenic markers (ie, β3-tubulin), but the protein expression was not observed [24]. Keeve et al. hypothesized that this phenomenon may be due to the translation repression of β3-tubulin mRNA by miRNA [24]. Agreeing with our observation, the β3-tubulin mRNA expression was noted in the control and hFc control surface. However, the protein expression was not observed by immunocytochemistry staining. In addition, it has been identified that periodontal ligament tissues contained a subpopulation that is derived from neural crest cells [10,25,26]. Thus, these cells may contain the intrinsic characteristics of neuroectodermal lineage. Together, hPDLSCs may be an alternative source for cell therapy in regenerative neuronal disease.
Notch signaling has been shown to regulate neural stem cell maintenance [27 –29]. Inhibition of Notch signaling resulted in an abundance of self-renewal stage and entry to neurogenic differentiation of neuronal stem cells [28]. However, an overexpression of Notch target gene, HeyL, promoted neuronal differentiation of neural progenitor cells both in vitro and in vivo [30]. Together, these data underscore the complex regulation of Notch signaling in neural stem cell maintenance and differentiation. In regard to mesenchymal stem cells, mRNA levels of Notch signaling pathway components were decreased in human adipose-derived mesenchymal stem cells upon neurogenic differentiation [31]. The inhibition of Notch1 expression enhanced neurogenic differentiation in murine bone marrow-derived mesenchymal stem cells [32]. On the contrary, some investigators reported that an activation of Notch signaling in mesenchymal stem cells promoted neuronal commitment. In this respect, overexpressing NICD in bone marrow stromal cells led to an increase in neurogenic markers, and these cells demonstrated action potentials as recorded by whole-cell patch clamp [33]. In addition, these Notch-induced mesenchymal-derived neuronal cells significantly improved behavioral responses upon implantation in a rat stoke model [34,35]. For embryonic stem (ES) cells, overexpression of NICD resulted in accelerated differentiation toward neuronal lineages [36]. NICD overexpression in ES cells resulted in the inhibition of non-neurogenic differentiation [36]. In addition, it was demonstrated that the co-culture of ES cells with stromal cells overexpressed Delta-like1 were able to promote the expression of neurogenic markers [36].
The discrepancies in observations regarding the role of Notch signaling in neurogenic differentiation may have several explanations. First, it has been shown that function of Notch signaling is cell type and developmental stage specific [37,38]. In this regard, a differential expression of Notch receptors was noted at different stages of development, which may influence intracellular signaling and cell fate decision [39,40]. Second, differences in neuronal induction protocols are likely to influence differentiation mechanism and efficiency [41]. Various reports employed β-mercaptoethanol as a component in neuronal induction medium [42]. Although, the protein expression of neuronal markers was upregulated, the neurogenic mRNA levels were not changed. Some studies showed that these β-mercaptoethanol-induced neurons did not exhibit neuron-like electrophysiological characteristics, suggesting pseudoneuronal differentiation [43,44]. In the present study, the neurosphere formation method was employed. Neurosphere-derived neuronal cells exhibited similar characteristics to those of bonafide neurons in term of mRNA and protein expression as well as electrophysiological properties [45,46]. Thus, the evaluation of Notch signaling using neurosphere formation technique could give a better understanding the mechanisms controlling true neuronal differentiation.
In the present study, we described the upregulation of Hes1 and Hey1 during neurogenic differentiation by hPDLSCs. However, the reduction of Hes1 and Hey1 mRNA levels was noted at later stages. These data suggest that Notch signaling may be required in neurogenic commitment of hPDLSCs and but that Notch signaling should fade out to permit neuronal maturation processes. Corresponding with our study, it was demonstrated that Hes1 mRNA levels were increased after exposing human bone marrow stem cells in neuroinduction medium [47]. However, it has to be noted that these genes have other potential functions, that is, cell proliferation and stemness maintenance, which should be considered [48]. In this regard, it has been reported that Hes1 prevented irreversible cell cycle exit in human fibroblast and rhabdomyosarcoma cells [49]. Thus, specific function of the upregulation of these genes in hPDLSCs should also be further investigated.
A role of Jagged-1 in cell fate decision control has been reported in several cell types. Jagged-1 promoted neuroectodermal commitment of ES cells [12]. Similar to that study, we illustrated that exposing hPDLSC to Jagged-1-modified surface promoted neurogenic commitment in serum-free culture medium as determined by the expression of neurogenic markers, Sox2 and β3-tubulin. However, our previous publication observed that Jagged-1 enhanced osteogenic differentiation in hPDLSCs in normal growth medium as well as in osteogenic medium [4]. These contrary results may have occurred for several reasons. First, the activation of Notch signaling prompts hPDLSCs to differentiate. However, the specific lineage determination is regulated by other external stimuli in the culture environment. In this respect, it has been reported that the activation of Notch target genes was differently regulated by extrinsic factor. Different culture medium supplementation influenced other intracellular signaling, which further controlled and interacted with Notch signaling [50]. Moreover, a withdrawal of trophic factors resulted in the reversible change of Numb isoforms that further controlled the activation of Notch signaling target genes [51]. Together, these results suggest that culture environment has considerable influences on the different regulation of Notch signaling target gene expression and cell's behaviors. Corresponding to our observation, Jagged-1 promoted neurogenic commitment of hPDLSCs in serum-free condition, but in the present of 10% serum, osteogenic differentiation was enhanced, implying the role of culture condition of Notch signaling-influenced cell function. Second, the difference in cell fate determination upon exposure to Notch ligand might have occurred due to different Notch receptor interactions as well as downstream intracellular signaling cascade. In this regard, it has been illustrated that an activation of canonical Notch signaling resulted differences in regulation and cell behavior compared to the noncanonical pathway [52]. In addition, it has been demonstrated that an alteration of Notch receptor expression may impact control of specific cell lineages as shown in olfactory epithelium [53]. In our laboratory, we observed the differential Notch receptor expression after hPDLSCs was induced to osteogenic, adipogenic, and neurogenic lineage (unpublished data). The mechanism for this requires further investigation.
In summary, we illustrated that the expression of Notch signaling target genes was changed during neuronal induction of hPDLSCs. Exposing cells to a Jagged-1-immobilized surface could promote morphological changes and expression of neurogenic markers. Together, these results suggest the potential role of Notch signaling in neuronal commitment of hPDLSCs. Further investigation on molecular mechanism is needed in order to elucidate a method to precisely control differentiation of these cells for clinical utilization.
Footnotes
Acknowledgments
This study was supported by “Integrated Innovation Academic Center: IIAC” Chulalongkorn University Centenary Academic Development Project. T.O. was supported by DRU Research Fund, Faculty of Dentistry, Chulalongkorn University and Thailand Research Fund. P.P. was supported by the Research Chair Grant 2012, the National Science and Technology Development Agency (NSTDA), Thailand. The authors thank Dr. Anjalee Vacharaksa for her advice regarding plasmid preparation and viral transfection. In addition, we thank Dr. Cecilia M. Giachelli for critical review of the manuscript.
Author Disclosure Statement
No benefit of any kind will be received either directly or indirectly by the author(s). No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.