
Abstract
The mental retardation-associated protein, srGAP3 is highly expressed in neurogenic sites. It is thought to regulate the key aspects of neuronal development and functions. Little is known about the interaction between srGAP3 and immature neural stem cells/neural progenitor cells (NSCs/NPCs). In the current study, the expression of srGAP3 in NSCs/NPCs was detected. Then, survival, proliferation, differentiation, and morphological alteration of NSCs/NPCs were assessed after a lentivirus-mediated knockdown of srGAP3. The results showed that srGAP3 is highly expressed in NSCs/NPCs both in vitro and in vivo. After knockdown of srGAP3 (LV3-srGAP3 infection), viability and proliferation of NSCs/NPCs dramatically decreased, approximately 85% displayed a similar morphology with type I cells that have no or only few indistinguishable processes. After 7 days culture in a differentiation medium, 62.5%±8.3% of cells in the srGAP3 knockdown group were nestin-positive and 24.8%±5.8% of them were β-tubulin III-positive, which are significantly higher (30.2%±9.9% and 14.6%±2.7%) than in the control group (LV3-NC infection). In addition, cells in the knockdown group had significantly fewer, but longer processes. Our results demonstrate that srGAP3 knockdown negatively regulates NSCs/NPCs survival, proliferation, differentiation, and morphological alteration, particularly, process formation. Taken together, our results provide strong evidence that srGAP3 is involved in the regulation of biological behavior and the morphological features in rat NSCs/NPCs in vitro.
Introduction
NSCs and NPCs are self-renewing and multipotent cells [17 –19]. They exist in neurogenic sites in the brain such as the VZ/SVZ and subgranular zone in the dentate gyrus of the hippocampus in both the developing brain and adult brain. They are responsible for generating the 3 main cell types of the nervous system [19,20]. During development, NSCs/NPCs assume different cellular morphologies and reside within changing microenvironments (the stem cell niche), while retaining their basic properties. NSCs/NPCs behave differently when exposed to various niche signals [21 –23]. Since srGAP3 is highly expressed in VZ/SVZ and influences neuronal differentiation and cell morphology, there must be some direct interaction between NSCs/NPCs and srGAP3. In other words, the influence of srGAP3 on neural development might be attributed to the regulation of NSCs/NPCs biological behavior.
To evaluate the effect of srGAP3 on the NSCs/NPCs, the expression of endogenous srGAP3 in NSCs/NPCs was first detected. Then, the survival, proliferation, differentiation, and morphological alteration of srGAP3 knockdown NSCs/NPCs were analyzed in vitro.
Materials and Methods
Culture and identification of rat embryonic NSCs/NPCs
NSCs/NPCs were isolated from the cerebral cortex of Sprague Dawley (SD) rat embryos on embryonic day 14 (E14) and cultured in a serum-free growth medium that contained the DMEM/F12 (the Dulbecco's modified Eagle's medium and Ham's F12, 1:1), 10 ng/mL of the basic fibroblast growth factor, 20 ng/mL of the epidermal growth factor, 1% penicillin, 1% streptomycin, 1% N2, and 2% B27 supplement (all from Invitrogen) following the protocol of Gage et al. [24] and optimized in the authors' laboratory [25]. The primary neurospheres were subcultured after 5 to 7 days in vitro (DIV). Upon passaging, the neurospheres were trypsinized and mechanically triturated into single cells. For observation of spontaneous differentiation, NSCs/NPCs were cultured on poly-
Construction of lentiviral vector and infection of NSCs/NPCs
The LV3 lentiviral vector carries a U6 promoter followed by a CMV-WPRE cassette that controls enhanced green fluorescent protein expression. The lentivirus encoding shRNA for srGAP3, LV3-srGAP3 was prepared by GenePharma. The shRNA sequence is as follows: 5′-GAATATGAAGCCCAAATAA-3′, which was demonstrated to knockdown srGAP3 expression dramatically [10]. The sequence for control shRNA (LV3-NC) was 5′-TTCTCCGAAC GTGTCACGT-3′. The viral solutions (5–10×108 TU/mL) were stored in aliquots at −80°C for later using.
NSCs/NPCs in passage 2 to 3 were processed for lentivirus infection. Neurospheres were trypsinized into single cells and were replaced at a density of 1×105 cells/mL 24 h before infection. LV3-NC and LV3-srGAP3 were added to the culture system at an multiplicity of infection of 20. The total protein was subjected for immunoblotting analysis to confirm the knockdown efficiency of LV3-srGAP3, which followed Chen's protocol [10]. The immunolabeled protein was detected by BM Chemiluminescence Western Blotting kit (Roche). Densitometric quantitation was acquired with Gel Doc 1000 system and analyzed using Quantity One software (BioRad). The antibodies, including srGAP3-3A1 (polyclonal, 1:4000) and α-tubulin (Santa Cruz, 1:1000) were used.
Assessment of NSCs/NPCs survival and proliferation
The survival and growth of NSCs/NPCs were analyzed by using a cell counting kit-8 (CCK-8, Dojindo). Cells at a concentration of 1×104/100 μL growth medium were added to each well of a 96-well plate. Cell viability in 3 wells was observed every day by using an UV-1750 spectrometer reader (Shimadzu) at 450 nm for 6 days. At the same time, the diameters of neurospheres obtained from 3 individual wells of the culture plate were measured using an eyepiece micrometer to reveal the survival and the proliferation of NSCs/NPCs. All spheroids were measured and the top 30 largest diameters were selected and averaged. The apoptosis of NSCs/NPCs was also examined by Hoechst 33342 (5 μg/mL, Sigma) staining. The percentage of apoptotic cells was compared. In addition, ki-67 expression and cell cycle were analyzed to further clarify the proliferation of NSCs/NPCs. NSCs/NPCs were cultured in 6-well plates at 5×105 cells/mL. Five days after infection, cell aggregates were dissociated into single-cell suspensions. Some of these cells were fixed with 4% paraformaldehyde (PFA) in a 0.1 M sodium phosphate buffer [phosphate-buffered saline (PBS, pH 7.4)] at room temperature for 30 min and plated onto the poly-
Morphological evaluation of NSCs/NPCs
The morphology of NSCs/NPCs cultured on coverslips in the differentiation medium was observed under a phase-contract microscope every day. Based on the various characteristics of cell morphology (numbers of processes), the attached cells were classified into 3 different types. Cells without or with few processes (<3) were identified as type I. Cells with 4 to 6 processes were identified as type II, and the cells with more than 6 processes were identified as type III. The proportion of the different types of cells in 5 individual coverslips was quantified by an observer blind to the treatment groups. Meanwhile, the number of processes of each cell was counted and the length of the longest process was measured using an eyepiece micrometer at different time points.
Tissue preparation and immunochemistry staining
Three embryonic brains were submersion fixed in freshly prepared 4% PFA in 0.1 M PBS at 4°C. Three adult SD rats were fixed by transcardiac perfusion with 4% PFA under terminal anesthetized by intraperitoneal injection of a 10% (W/V) chloral hydrate solution (300 mg/kg). Whole brains were dissected and postfixed in 4% PFA at 4°C overnight and cryoprotected in sucrose 30%, in a sodium phosphate buffer (pH 7.4). The cryosections containing VZ/SVZ were cut at 16 μm before immune-staining. Cells were cultured on poly-
Immunochemistry staining was performed following the standard protocol and optimized in the authors' laboratory [25]. Primary antibodies, including polyclonal rabbit anti-srGAP3-3A1 antibody [10], polyclonal mouse anti-ki-67 (Chemicon, 1:300), and mouse anti-nestin, mouse anti-β-tubulin III, mouse anti-GFAP, and mouse anti-O4 (described as above) were diluted in PBS with 2% normal goat serum (NGS). The blocking solution contained 5% NGS and 0.25% Triton X-100 in PBS. Tetramethyl rhodamine isothiocyanate- and fluorescein isothiocyanate-conjugated goat anti-mouse IgG/anti-rabbit IgG (1: 400; CoWin Biosciences) were used as secondary antibodies. Cell nuclei were counterstained with 4′,6-diamidino-2-phenylindole-containing mounting media (Vector Labs) and visualized under a fluorescent microscope (Olympus BX57) equipped with a DP70 digital camera and the DPManager (DPController, Olympus) software. For the negative control, the primary antibody was replaced by the blocking buffer.
Statistic analysis
All the quantitative data were obtained from 3 independent experiments. They were presented as mean±SE, and statistical analysis was performed with software Prism 4.0. The T-test and one-way ANOVA were used and a P value <0.05 was considered significant.
Results
SrGAP3 is highly expressed in NSCs/NPCs
NSCs/NPCs were isolated from the cerebral cortex of SD rat embryos and cultured in the growth medium. Neurospheres of different sizes developed over a period of 5 to 7 DIV and the majority of cells in neurospheres were found to be nestin-positive (Fig. 1A, B). After 7 days culture in the differentiation medium, β-tubulin III-positive neuronal progenitors/neurons, GFAP-positive astrocyte progenitors/astrocytes, and O4-positive oligodendrocytes were observed (Fig. 1C–E). SrGAP3 expressed in almost 100% of cultured cells, including nestin-positive NSCs/NPCs (Fig. 1F, srGAP3/nestin double labeled), and in differentiated neuronal progenitors and neurons (Fig. 1G, srGAP3/β-tubulin III double labeled) or astrocyte progenitors and astrocytes (Fig. 1H, srGAP3/GFAP double labeled). The expression persisted for periods of up to 14 days in vitro (data not shown). Expression of endogenous srGAP3 was also detected in brain tissue, results showed that, both in developing (Fig.1I, J) and adult brain (Fig. 1K, L), srGAP3 expressed predominantly in VZ/SVZ and colocalized with nestin-positive NSCs/NPCs.

Expression of srGAP3 in neural stem cells/neural progenitor cells (NSCs/NPCs). NSCs/NPCs isolated from the rat embryo cortex were cultured in the Dulbecco's modified Eagle's medium/F12 serum-free medium.
NSCs/NPCs viability and proliferation dramatically decreased after srGAP3 knockdown
Expression of srGAP3 in cultured NSCs/NPCs significantly decreased after the infection of LV3-srGAP3 (Fig. 2A). Shortly after infection, some of NSCs/NPCs started to attach to the surface of tissue culture flasks and the cell debris was clearly observed on the second day, especially in the knockdown group (LV3-srGAP3 infection). Although cells in both groups proliferated and grew into neurospheres at 7 DIV, the diameters of neurospheres in the knockdown group were significantly smaller than that in the control group (Fig. 2B, P<0.001). There were significantly more apoptotic cells with condensed chromatin and stained more heavily (Fig. 2C, arrow, P<0.001) in the knockdown group. The CCK-8 assay further confirmed that the number of viable cells dramatically decreased after LV3-srGAP3 infection (Fig. 2D). Approximately 58.7%±1.8% of cells in the knockdown group were ki-76-positive, which is significantly less than that in the control group (83.2%±2.1%, P<0.001, Fig. 2E). Cell cycle analysis by FACS similarly showed a significantly lower percentage of cells in phases G2/M and S in the srGAP3 knockdown group (5.1%±2.0%, 15.5%±3.1%) as compared to the control group (13.0%±3.4% and 26.9%±5.7%) (Fig. 2F, P<0.01).
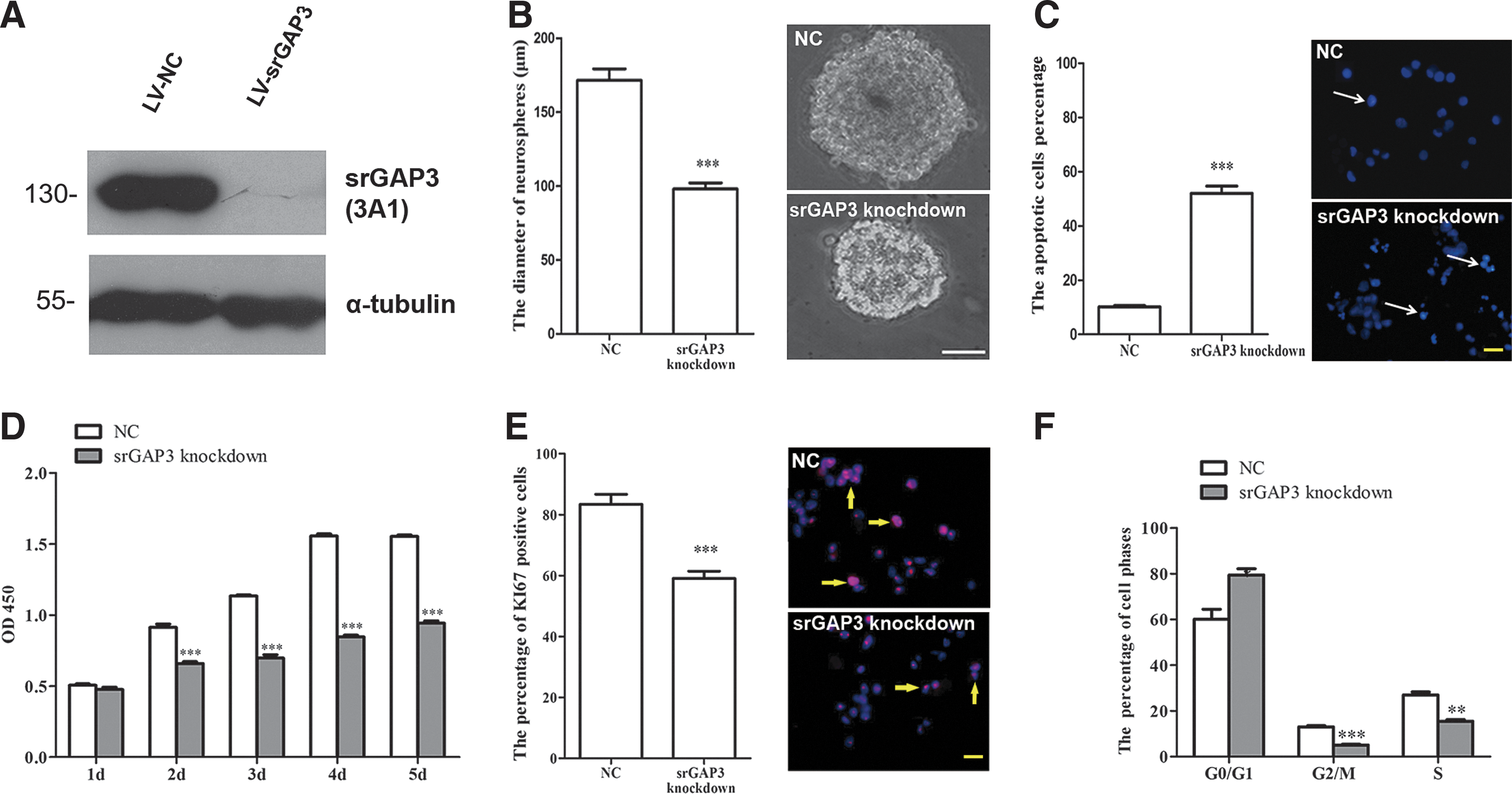
Assessment of survival and proliferation of NSCs/NPCs after SrGAP3 knockdown.
Most of cells in the srGAP3 knockdown group appeared as type I cells
The number of cells showing different morphological features (Fig. 3A) was counted at different time points. In the knockdown group, with the culturing in the differentiation medium, the percentage of type I cells increased (Fig. 3B), whereas the proportion of type II cells slightly decreased (Fig. 3C). There was no significant alteration in regarding type III cells (Fig. 3D). In comparison with the control group, there was significantly more type I cells, but less type III cells in the srGAP3 knockdown group (Fig. 3B, D, P<0.05).
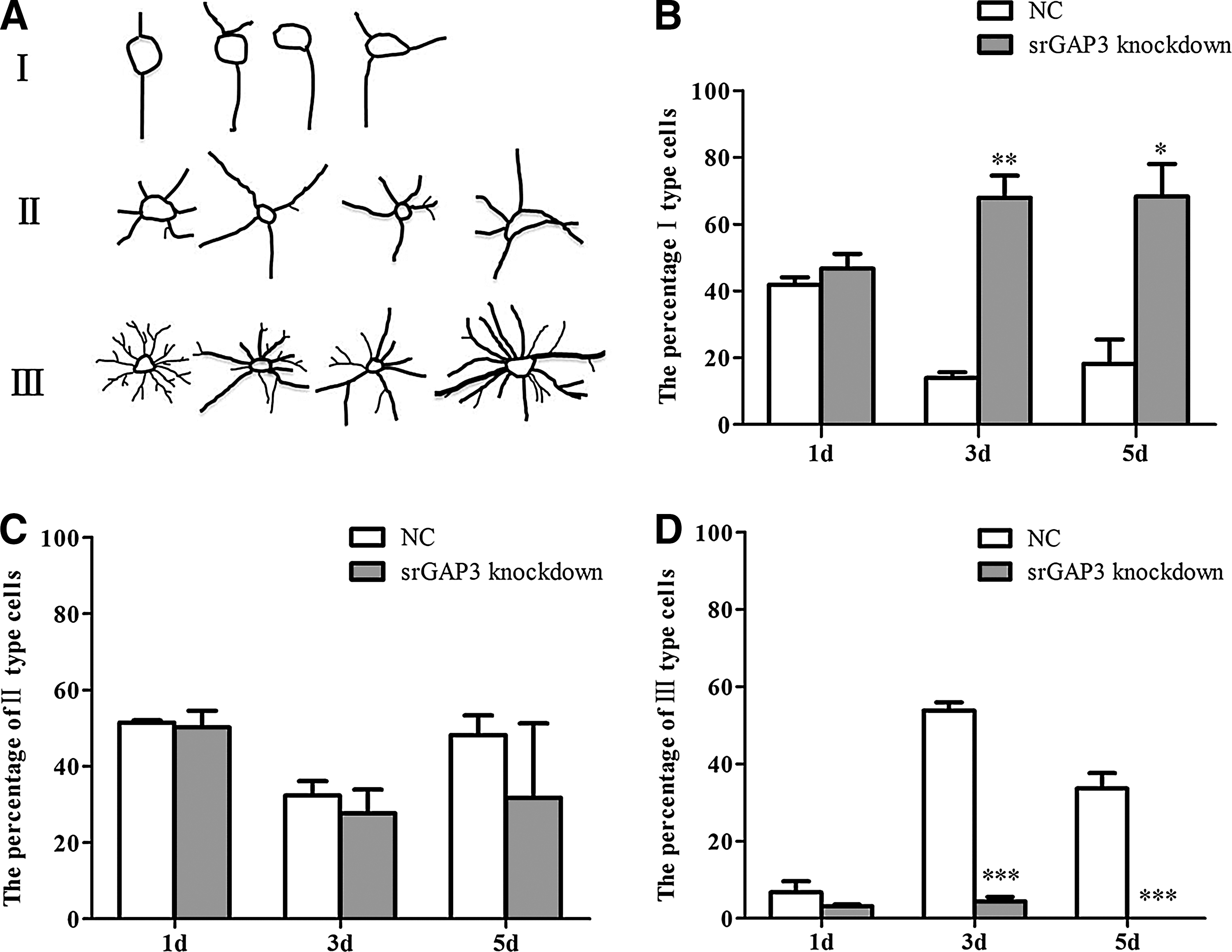
Morphological observation of NSCs/NPCs after srGAP3 knockdown.
Cells in the srGAP3 knockdown group predominantly remained as immature cells
After 7 days culture in the differentiation medium, in the knockdown group, 62.5%±8.3% of the cells were nestin-positive undifferentiated cells and 24.8%±5.8% were β-tubulin III-positive neurons. This represented a significantly higher percentage than in the control group (30.2%±9.9% and 14.6%±2.7%, respectively). However, the percentage of GFAP-positive astrocytes was significantly lower in the knockdown group (26.4%±1.6%) than in the control group (40.3%±9.7%, Fig. 4A, P<0.05). No significant difference was noticed between these 2 groups in regarding to the ratio of O4-positive cells. In addition, a small proportion of these cells were nestin/β-tubulin III or nestin/GFAP double-positive progenitors (data not shown). Interestingly, in the srGAP3 knockdown group, cells displayed a similar morphology having less than 3 processes per cell, which differed from the control group and was independent of the specific phenotypes (Fig. 4B).
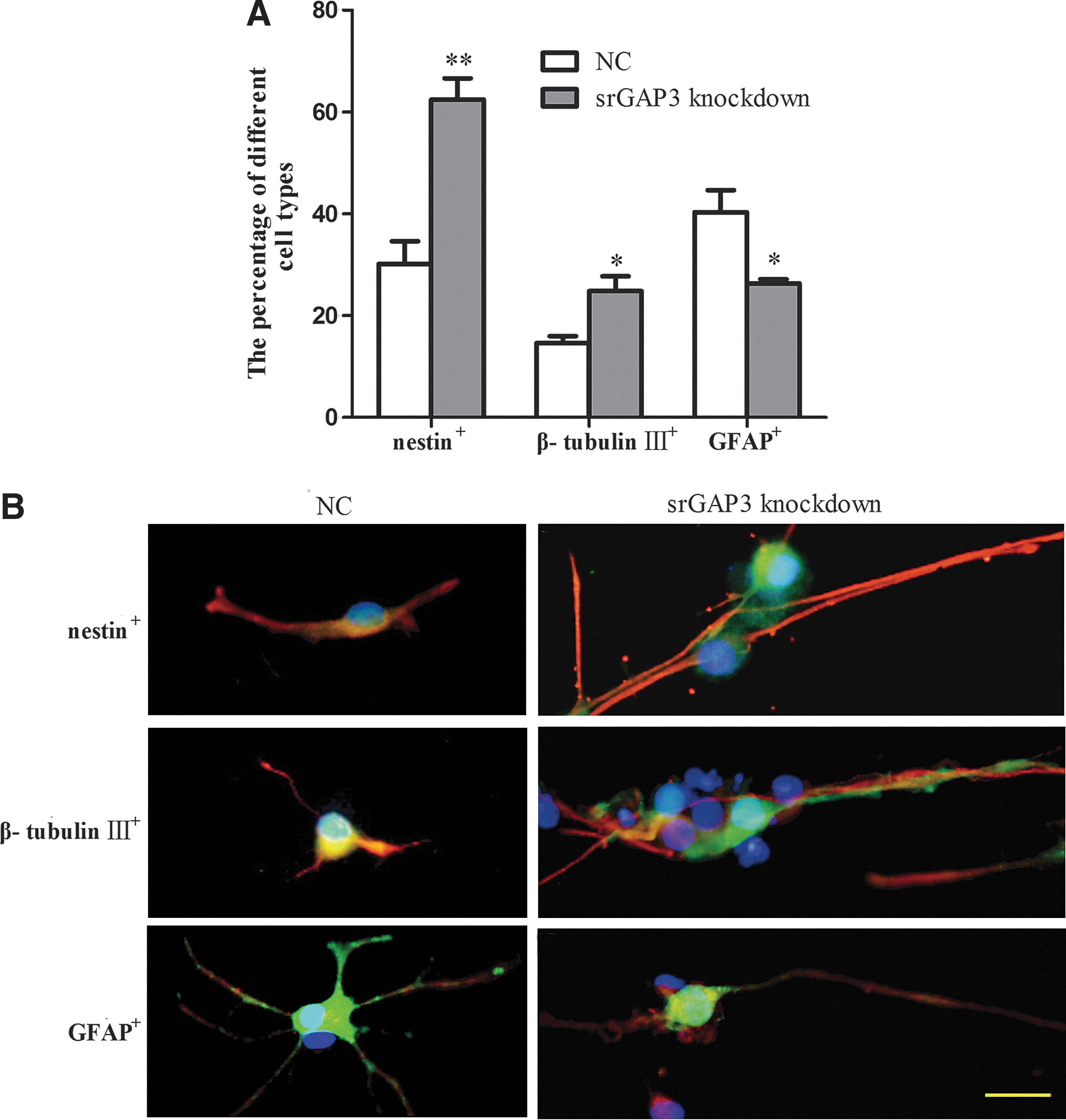
Differentiation of NSCs/NPCs after srGAP3 knockdown.
Cells in the SrGAP3 knockdown group had significantly fewer, but longer processes
To further clarify the effect of srGAP3 on the morphological features of NSCs/NPCs during spontaneous differentiation, the average number of processes per each cell and the length of the longest process were quantified. Result showed that the density of cell processes in the control group increased with respect to time in culture, a similar pattern was not found in the srGAP3 knockdown group (Fig. 5A). Quantification of cell processes showed that after 3 days, each cell in the knockdown group had significantly less processes than those of the control group (Fig. 5B, P<0.01). Nevertheless, we noticed that with the increasing culture time, the length of the longest processes gradually increased, particularly in the srGAP3 knockdown group and were found to be significantly longer than those of the control group (Fig. 5C, P<0.01). No difference was found in the cell soma size.
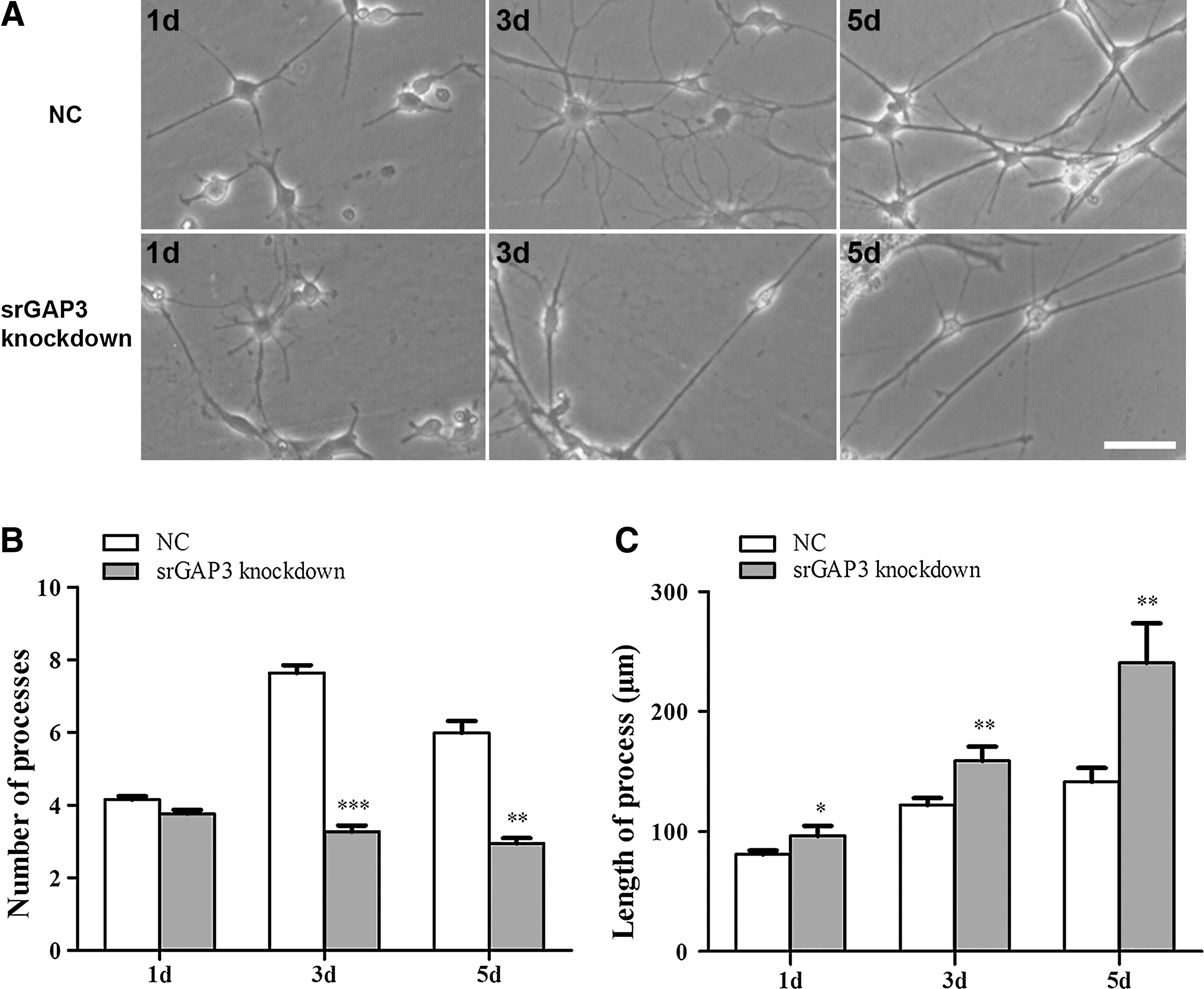
Quantification of the number of processes and the length of axons.
Discussion
The results presented here shown that srGAP3 is expressed predominately in NSCs/NPCs both in vitro and in vivo. After srGAP3 knockdown, NSCs/NPCs viability and proliferation is dramatically reduced, the differentiation of NSCs/NPCs is delayed, cell morphology altered, and the genesis of process was attenuated, but the extension was significantly enhanced.
It has been shown that srGAP3 is highly expressed in the neurogenic areas [8,16,26,27]. It is possible that it functions as a niche signal to regulate NSCs/NPCs biological behavior and fate decisions. Here we found that endogenous srGAP3 is not only widely expressed in the NSCs/NPCs in neurogenic sites in vivo, but also in cultured NSCs/NPCs. Interestingly, it was also detectable in β-tubulin III-positive and GFAP-positive cells. It may be because some of these cells were neuronal progenitors or glial progenitors. It also suggests that srGAP3 might have an effect on the neuron and glia.
In mammals, srGAP3 attenuates the activity of Rho GTPases resulting in the alteration of the cytoskeleton proteins. It therefore plays important roles in cell surviving and proliferation [2 –5]. We found here, after knockdown of srGAP3, more NSCs/NPCs went into apoptosis and less than a quarter of cells were in active mitosis, which further resulted in the dramatically decreased cell viability and reduced size of neurospheres. This result further confirms that srGAP3 is essential in regulating NSCs/NPCs survival and proliferation. The Slit-Robo signaling pathway and Rac1 may be involved [2,10,28]. In addition, we noticed that approximately two thirds of cells in the srGAP3 knockdown group were nestin-positive progenitors after 7 days culturing in the differentiation medium. In other words, differentiation of NSCs/NPCs was attenuated after srGAP3 knockdown. This result suggests that srGAP3 is also crucial for NSCs/NPCs differentiation. The mechanism regarding how srGAP3 is critically needed for maintaining the NSCs/NPCs population and also plays crucial roles in NSCs/NPCs differentiation still remain unknown.
Our previous studies observed the expression of endogenous srGAP3 was upregulated in the early stages of neuronal differentiation of Neuro2A cells, however, low levels of srGAP3 enhanced the Neuro2A neuronal differentiation. This paradoxically phenomenon may due to the membrane relocalization of srGAP3 [10]. Similarly, we found that srGAP3 is detectable in β-tubulin III-positive neuronal progenitors/neurons, and there were more β-tubulin III-positive, but less GFAP-positive cells in the srGAP3 knockdown group compared to the control group. Combining these results, it suggests that the effect of srGAP3 on neuronal differentiation might lie in a spatiotemporal sequence. It is relevant therefore that we should not ignore that some of the β-tubulin III-positive cells that we have counted as neurons as being neuronal progenitors (nestin/β-tubulin III double-positive), as neuronal differentiation precedes glial differentiation during development. However, the significant reduction of GFAP-positive cells in the srGAP3 knockdown group, including the nestin/GFAP double-positive glial progenitors, indicates that glial differentiation of NSCs/NPCs was attenuated dramatically by low levels of srGAP3. No significant effect was observed on oligodendrocyte differentiation. This phenomenon has not been reported previously, and it therefore suggests that srGAP3 may also play important roles in gliogenesis. Whether this is related to the activity of the Rho GTPase activity or neurogenin 2 needs to be further studied.
Rho proteins are key regulators of the cellular cytoskeleton. Different extracellular and intracellular signals converge on the regulation of the cytoskeleton. Rho GTPase is one of the major intracellular regulators [2,10,29 –33]. Immature NSCs/NPCs are normally round and symmetrical without any process in the undifferentiated state. The appearance of processes sprouting from the cell body indicates the start of differentiation/maturation. As development proceeds, the symmetry is broken, and cell polarity is established alongside the NSCs switch to lineage restricted progenitors. Finally, mature neurons and glia acquire their specific morphological features [32,33]. Breaking the symmetry, in particular axon specification depends on cytoskeletal rearrangements [32]. Cells with low levels of srGAP3 might have difficulties in the cytoskeletal remodeling that resulted in the arrest of morphological changes. This suggestion is confirmed by our results and showed that a high percentage of the cells in the srGAP3 knockdown group were of type I cells, which have no or very few indistinguishable processes. Furthermore, cell differentiation always accompany with morphological alteration. Therefore, the type I cells in the srGAP3 knockdown group might be undifferentiated progenitors or in a very early stage of differentiation. This is consistent with our immunochemistry staining.
SrGAP3 has also been implicated in axon guidance, neuronal migration, neurite outgrowth, and dendritic morphogenesis during neuronal development [11 –15]. In the current study, we found that the cell process development was attenuated by low levels of srGAP3. However, no corresponding changes in the process outgrowth were observed. The average length of the longest processes increased gradually with increasing time in culture and was significantly higher in the knockdown group compared to the control group. This is consistent with existed data [2,10], and indicates that srGAP3 may have a different influence on process genesis and extension. In addition, due to the distinct cytoskeletal reorganization, the morphology of neurons and astrocytes altered differently in response to insults. After stimulation, the processes of astrocytes retract as the cell soma enlarges and progresses to cell division. On the contrary, processes of neurons, particularly axons extend primarily to form synapses. In the current study, a dramatic decrease in the number, but a significant increase in the length of cell processes was observed. It should be pointed out that some astrocytic processes might have also been included in the evaluation, especially in the earlier period. However, after 3 days in culture, a significantly higher percentage of cells in the srGAP3 knockdown group could be identified as neurons as seen under the phase-contrast microscope and displayed typical neuronal morphology having 1 axon and few dendrites. The number of neurites dramatically decreased and the length of the longest neurite (axon) significantly increased. The question regarding the difference of the srGAP3 effect on the cytoskeletal reorganization of neurons and astrocytes needs to be further studied.
In conclusion, we found that knockdown of srGAP3 expression attenuated NSCs/NPCs survival, proliferation, and differentiation and arrested their morphological alteration. It suggested that srGAP3 may play an important role in the biological behavior and fate decision of rat NSCs/NPCs. It may also in some way direct the polarity formation of NSCs/NPCs. SrGAP3 is closely related to mental retardation. It might have a significant bearing on the ability of neurons to make appropriate synaptic contacts in the formation of neural networks. Failure to achieve this would lead to inappropriate or missing neural circuitry. Further in vivo studies regarding the relationship between srGAP3 and neural development as well as the cellular molecular mechanism are needed.
Footnotes
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (31070943, 81070998, and 31171033).
Author Disclosure Statement
The authors indicate there is no conflict of interest in connection with this article.