
Abstract
Multiple sclerosis (MS) is an irreversible and demyelinating disease of the central nervous system, in part influenced by chronic inflammation. There is no proven effective therapy to stop the pathological progression of MS, although suppressing the immune system to control the inflammatory response may improve the clinical performance acutely. Here, we found that mesenchymal stem cells from human umbilical cord (hUC-MSCs) could restore behavioral functions and attenuate the histopathological deficits of experimental autoimmune encephalomyelitis mice over the long term (ie, 50 days) by suppression of perivascular immune cell infiltrations and reduction in both demyelination and axonal injury in the spinal cord. These findings suggest that transplantation of hUC-MSCs may be a potential therapy for MS.
Introduction
Stem cell therapy has emerged as a potential treatment for MS. Hematopoietic stem cell transplantation (HSCT) is the first candidate for cell-based therapy, including both autologous and allogeneic HSCT. However, a consensus has not been reached on HSCT, as patients with MS have to follow a series of cytotoxic drugs and radiotherapy to inhibit cell immunity, and the mortality rate is still ∼40% due to graft-versus-host disease [3]. Moreover, HSCT is only effective for patients with acute malignant MS, but not those with diffuse white matter damage [4]. In the last decade, transplantation of neural precursor cells (NPCs) has been explored as an alternative cell transplant source for MS due to the cells' therapeutic plasticity [5]. However, logistical issues in propagating ample supply of NPCs make them challenging for clinical application. In parallel, mesenchymal stem cells (MSCs) are multipotent cells found in almost all organs and tissues, including bone marrow (BM), embryo, adipose tissue, and umbilical cord (UC) [6]. To date, BM has been regarded as the most common source for MSCs, and various preclinical data have demonstrated the beneficial effects of BM-MSC transplantation in alleviating the symptoms of experimental autoimmune encephalomyelitis (EAE). However, the invasive procedure associated with harvesting BM-derived MSCs (BM-MSCs) and the low yields of viable stem cells limit their usage in clinic [7]. Recently, MSCs derived from human UC (hUC) have attracted much attention because of a sufficiently high cell yield without a painful, invasive harvesting procedure. Several studies have provided compelling evidence that hUC-MSCs had the higher proliferation and differentiation ability than BM-MSCs [8 –10]. Moreover, hUC-MSCs have significantly lower human leukocyte antigen-1 (HLA-1) expression, stronger immune tolerance, and a faster proliferation rate than BM-MSCs [11].
In this study, we tested our hypothesis that hUC-MSCs exerted therapeutic effects on an EAE mouse model through the grafted cells' immunomodulatory properties, which may involve both anti-inflammation and remyelination via leucine-rich repeat and Ig domain containing NOGO receptor-interacting protein 1 (LINGO-1)-signaling pathways.
Materials and Methods
EAE model and behavioral test
All mice were from the Drum Tower Hospital Animal Center, and the protocols were approved by the Committee of Experimental Animal Administration of Nanjing University. EAE was induced in 6–7-week-old female C57BL/6 mice by immunization with an emulsion containing 300 μg of purified myelin oligodendrocyte glycoprotein (MOG) peptide (MEVGWYRSPFSRVVHLYRNGK, corresponding to residues 35–55) in phosphate-buffered saline (PBS) and an equal volume of complete Freund's adjuvant (Sigma). About 0.2 mL of the inoculum was injected subcutaneously on the day of induction (day 0) and day 7. In addition, 300 ng of Bordetella pertussis toxin (Merck) in 0.2 mL PBS was administrated intraperitoneally at days 0 and 2, as described previously (n=20 per group) [12]. Mice were scored daily for neurological symptoms up to 50 days after EAE induction as follows: 0=asymptomatic; 1=partial loss of tail tonicity; 2=atonic tail; 3=hind-leg weakness and/or difficulty to roll over; 4=hind-leg paralysis; 5=foreleg paralysis; 6=death due to EAE. Clinical symptoms of EAE animals usually appeared on the day 9–10 post-EAE induction, arriving at the peak score after day 15 with no spontaneous restoration (as shown in Fig. 1A).
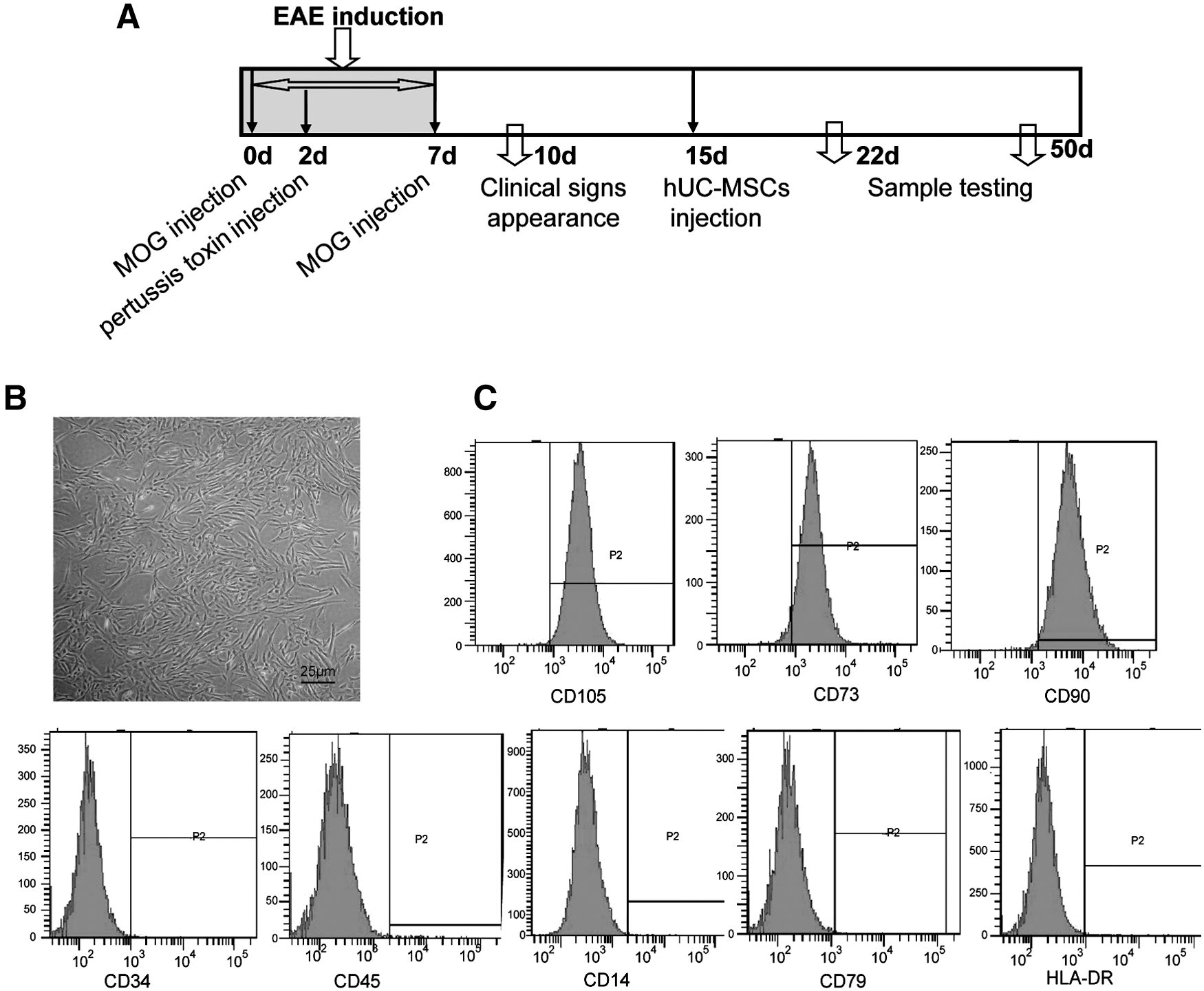
The schematic diagram of the experiment and properties of hUC-MSCs in vitro.
Preparation and transplantation of hUC-MSCs
hUC-MSCs were prepared by the Jiangsu Provincial Stem Cell Engineering Research Center (Beike Bio-Technology) under an approved clinical protocol, including informed consent forms from donors. Fresh UC was obtained from healthy mothers in a local maternity hospital after normal deliveries and were processed as quickly as possible. The cords were rinsed twice in PBS in penicillin and streptomycin, and the cord blood was removed. The cords were cut into 1-mm2 pieces and floated in the Dulbecco's modified Eagle's medium/F12 medium with human platelet-derived growth factor-BB, basic fibroblast growth factor, transforming growth factor-β1, and epidermal growth factor and then incubated at 37°C in a humidified atmosphere consisting of 5% CO2. When well-developed colonies of fibroblast-like cells appeared after 10 days, the cultures were trypsinized and passaged into a new flask for further expansion.
All of the infused hUC-MSCs were derived from passages 2–3 and got the concentration of 2×107/mL with rigorous purification and quality control (Table 1). For EAE (induction 15 days), a single injection of 2×107/mL×0.1 mL hUC-MSCs or medium via the tail vein was employed.
HBsAg, hepatitis B surface antigen; HBcAb, hepatitis B core antibody; HCV-Ab, hepatitis C virus antibody; anti-HIV (I+II), human immunodeficiency virus antibodies I and II; CMV-IgM, cytomegalovirus IgM; HTLV-I/II, human T lymphocytic leukemia virus; Anti-EBV antibody, Epstein-Barr virus antibodies; ALT, aspartate aminotransferase; HLA, human leukocyte antigen.
Histological assay
All mice were deeply anaesthetized with chloral hydrate (400 mg/kg i.p) and were perfused transcardially with ice-old PBS, followed by perfusion with 4% ice-cold phosphate-buffered paraformaldehyde on day 22 (acute phase) after MOG induction. Tissues were postfixed for 48 h in paraformaldehyde, and then 8-μM-thick spinal cord longitudinal frozen sections were performed. The sample size was n=10 per group.
Hematoxylin–eosin staining
To reveal perivascular inflammatory infiltrates, 6 different rostrocaudal levels of longitudinal spinal cord sections were detected at 22 days post-EAE induction after hUC-MSCs or vehicle injection. A blood vessel surrounded by 10 nuclei or more was considered as a positive perivascular infiltrate. All sections were studied under fixed illumination of a light microscope (Olympus BX51; at 40× magnification). The total average numbers of infiltrations per mm2 in the spinal cord were counted with the software Image-pro Plus (IPP).
Luxol fast blue staining
Luxol fast blue can differentiate the fiber tracts (white matter) from cell bodies (gray matter). For assessing demyelination of the spinal cord, 5 serial spinal cord sections per mouse were stained at 22 days post-EAE induction. Briefly, frozen sections were placed directly into a 1:1 alcohol/chloroform solution for a few hours, and then hydrated back with 95% ethyl alcohol, and left in luxol fast blue solution in 56°C overnight. The slides were differentiated in lithium carbonate solution for 30 s and then in 70% ethyl alcohol for 30 s. After the samples were rinsed in distilled water, they were checked microscopically to see if the gray matter was clear, and the white matter was sharply defined. When differentiation was complete, the samples were counterstained in cresyl violet solution for 30–40 s. All sections were studied under fixed illumination of a light microscope (Olympus BX51; at 40× magnification). The areas of luxol fast blue loss were calculated with software IPP, representing areas of demyelination (% demyelination/section) [12,13].
Amyloid precursor protien immunohistochemistry and Bielschowsky silver staining
For accessing the axonal pathology, the spinal cord was sectioned into 5 longitudinal serial sections at 22 days post-EAE induction. Amyloid precursor protein (APP) was accumulated within acutely injured axons. Sections were incubated with an antibody against APP (rabbit anti-APP, A8717; 1:1,000; sigma) at 4°C overnight, and followed by biotinylated anti-rabbit secondary antibody and ABC kit (Vector Laboratories) and visualized using 3,3′-diaminobenzidine (Dako). Bielschowsky silver staining was performed according the standard procedure [14]. All sections were studied under fixed illumination of a light microscope (Olympus BX51). Axonal damage was evaluated microscopically (at 40× magnification) throughout the spinal cord by counting APP+ axonal swellings and spheroids (APP+ profiles/mm2) and calculating the area of silver loss (%), respectively [12].
Electron microscopy
For evaluating remyelinated axons, samples were cut into cubes (<1 mm3) and postfixed for an additional 5 days in 2% gluteraldehyde+2% paraformaldehyde, and then processed as previously described [15]. For quantification, normally myelinated, demyelinated, and remyelinated axons were counted at 1200× magnification using the Olympus AX80 microscope and imaging software as described previously [15] with slight modifications. For analyses, all demyelinated, remyelinated, and normally myelinated axons within the grid were touch-counted and then averaged with all grids for animal averages. Each animal average was subsequently averaged within their group for the total axon counts and graphed.
Flow cytometry assays
Spleens were simultaneously harvested at day 22 post-EAE-induction (n=6), and single-cell suspensions were obtained [16]. For the Th1/Th2 ratio and Th17 analysis, samples were stained with a combination of the following mAbs (eBiosciences): CD3-APC (14502C11), CD4-FITC (GK1.5), and CD8-PE (5306.7), and then cells were stimulated with 25 ng/mL phorbol12-myristate13-acetate (Sigma), 1 μg/mL lonomycin, and 10 μg/mL brefeldin A (ALEXIS), and incubated at 37°C and 5% CO2 for 4 h. After stimulation, fluorescent-labeled anti-mouse monoclonal antibodies [CD3-APC, CD8-FITC, IL-4-PE (11B11), IFN-γ-PE (XMG1.2), and IL-17-PE (17B7)] were used for FACS assay. For CD4+CD25+ regulatory T cell (Treg cells) analysis, samples were stained with a mouse regulatory T cell staining kit (eBioscience; Cat. No. 88–8111). Briefly speaking, splenocytes were surface stained with anti-mouse CD4-FITC (RM4-5) and anti-mouse CD25-APC (PC61.5), then fixed and permeabilized using the Foxp3 staining buffers, and subsequently stained with 0.5 μg of anti-mouse Foxp3-PE (FJK-16s). T-lymphocyte phenotypes were identified and Th1/Th2 was analyzed by 3-color flow cytometry on an FACSCanto™ machine using BD FACS Dvia Software (BD Biosciences). In each experiment, cells were stained with an isotype control Ab to establish background staining and to set quadrants before calculating the percentage of positive cells [17].
Splenocyte culture in vitro
Using a sterile technique, the spleens were segregated into the 3 groups: control and EAE mice with or without hUC-MSC injection (n=8). The manipulation was followed by procedures described previously [16]. Briefly speaking, a single-cell suspension was prepared by passing the tissue through a 400-micron nylon mesh screen. The cells were washed using RPMI 1640 (Gibco-Invitrogen), and red cells were lysed using a red-cell lysis buffer [8.3 g NH4Cl in 0.01M Tris–HCl (pH 7.4)] and incubated for 5 min. The supernatant was produced by centrifugation (4°C, 1,200 revolutions/min). Splenocytes were then washed with RPMI 1640, counted, and adjusted to 1×106 cells/mL with the RPMI medium. Splenocyte cultures (1×106 cells/well in 2 mL) were stimulated with or without MOG35-55(10 μg/mL) in 6-well tissue culture plates (Corning). The culture plates were incubated for 72 h at 37°C and a 5% CO2 incubator. After 3 days, the supernatant was harvested and stored at −80°C for cytokine detection by ELISA assay.
In vitro proliferation assays
Cell proliferation was assessed by addition of 20 μL (10 μM/mL) BrdU in the culture medium for a period of 24 h after MOG (10 μg/mL) or the non-T-cell receptor-mediated mitogen ConA (2.5 μg/mL) stimulation. Cells were then programmed with an FITC BrdU Flow Kit (BD Pharmingen™; Cat. No. 559619) staining protocol. Briefly, BrdU-pulsed cells were added to flow cytometry tubes and washed by adding 1 mL of staining buffer per tube, centrifuged for 5 min at 200–300 g/s, and discarded the supernatant; the cells were fixed and permeabilized with a BD cytofix/cytoperm buffer, incubated with a BD cytoperm plus buffer for 10 min, and refixed again; then, cells were treated with DNase to expose incorporated BrdU, and stained with BrdU using a fluorescent antibody. Stained cells were analyzed with a flow cytometer (run at a rate not >400 events/s) and acquired as multiparameter data files.
ELISA assays for cytokines
Mouse cytokine ELISA kits (R&D Systems) were used to detect levels of IL-4 (M4000B), IL-10 (M1000), IL-17 (M1700), TNF-α (MTA00), and IFN-γ (MIF00) in the supernatants of splenocytes in all groups (n=8). One hundred microliters of sample or standard was added to each well of 96-well plates coated with anti-mouse cytokines or hormone antibody. The plates were incubated at 37°C for 90 min and then washed 5 times. One hundred microliters of biotinylated cytokine or hormone-specific antibody was added into each well and incubated at 37°C for 60 min. Plates were then washed, treated with 100 μL of diluted streptavidin–horseradish peroxidase (HRP), and incubated at 37°C for 30 min. After washing, the color was produced by addition of 100 μL substrate solution for 10–15 min. Finally, 100 μL of stop solution was added to terminate the reaction. The optical density was measured at 450 nm within 10 min [16].
Real-time polymerase chain reaction
Real-time polymerase chain reaction (PCR) was performed as described previously [16]. Total RNA was extracted by using the Trizol reagent (Takara) and was reverse-transcribed into cDNA using a PrimeScript RT reagent kit (Takara) for Quantitative PCR (ABI 7500) in the presence of a fluorescent dye (SYBR Green I; Takara). The relative abundance of mRNA was calculated after normalization to glyceraldehyde-3-phosphate dehydrogenase ribosomal RNA. The primers (Invitrogen) were as follows:
LINGO-1: F: GCGCCACACCTGAGTTCGT, R: TGCACCGTGTGGCCCTCATCTA;
GAPDH: F: GCCAAGGCTGTGGGCAAGGT, R: TCTCCAGGCGGCACGTCAGA
Western blot
The procedure used was published previously [16]. Proteins from spinal cord were extracted and then quantified using a Coomassie Blue Fast Staining Solution according to the manufacturer's instructions. Equal amounts of total protein samples were separated by sodium dodecyl sulfate–PAGE and blotted onto polyvinylidene fluoride membranes. The membranes were probed with primary antibodies against LINGO-1 (1:2,000; Abcam), Fyn (1:500; Santa Cruz), phosphorylated-Fyn (p-Fyn, 1:500; Bioworld), and glyceraldehyde-3-phosphate dehydrogenase (1:2,000; Bioworld). HRP-conjugated anti-rabbit or anti-mouse secondary antibodies were then used, and the reaction was observed using chemiluminescence reagents provided with the ECL kit (Bioworld) and exposed to a film. The intensity of blots was quantified by densitometry. Glyceraldehyde-3-phosphate dehydrogenase was used as internal control.
Statistical analysis
Data were retrieved and processed using the Microcal Origin 7.5 software program. The group data were expressed as the mean±standard error. The intergroup difference was analyzed by t-test and one-way analysis of variance (ANOVA) with the Bonferroni's post hoc comparisons as the between-subject factor. The difference of behavioral-deficit scores between hUC-MSC-injected EAE and vehicle was analyzed by two-way ANOVA. SPSS 15 software was used for the statistical analysis. A probability of P<0.05 was considered to be significant.
Results
Properties of hUC-MSCs in vitro
As shown in Fig. 1B, hUC-MSC viability was >92%. The cells were negative for CD34, CD45, CD14, CD79, and HLA-DR and positive for CD105, CD73, and CD90 (Fig. 1C). To ensure that these cells were safe for treatment, common microorganisms and viruses such as bacterial, hepatic viruses, and human immunodeficiency virus were tested negative (Table 1).
hUC-MSCs ameliorate EAE symptoms
To investigate the hUC-MSC effect on EAE, the behavioral test was employed. The results indicated that hUC-MSCs remarkably decreased the behavioral-deficit scores at day 22 (the peak of the acute phase) post-EAE induction (2.70 vs. 3.18, P<0.05) and day 50 (the end of the follow-up, representing the chronic phase) (2.78 vs. 3.25; P<0.05, Fig. 2A), compared to the vehicle. Moreover, the area under curve, which approximated the cumulative severity of disease [12], showed 11.50% (at day 22) and 14.73% (at day 50) reduction in hUC-MSC-treated EAE mice compared to vehicle-treated EAE, respectively.
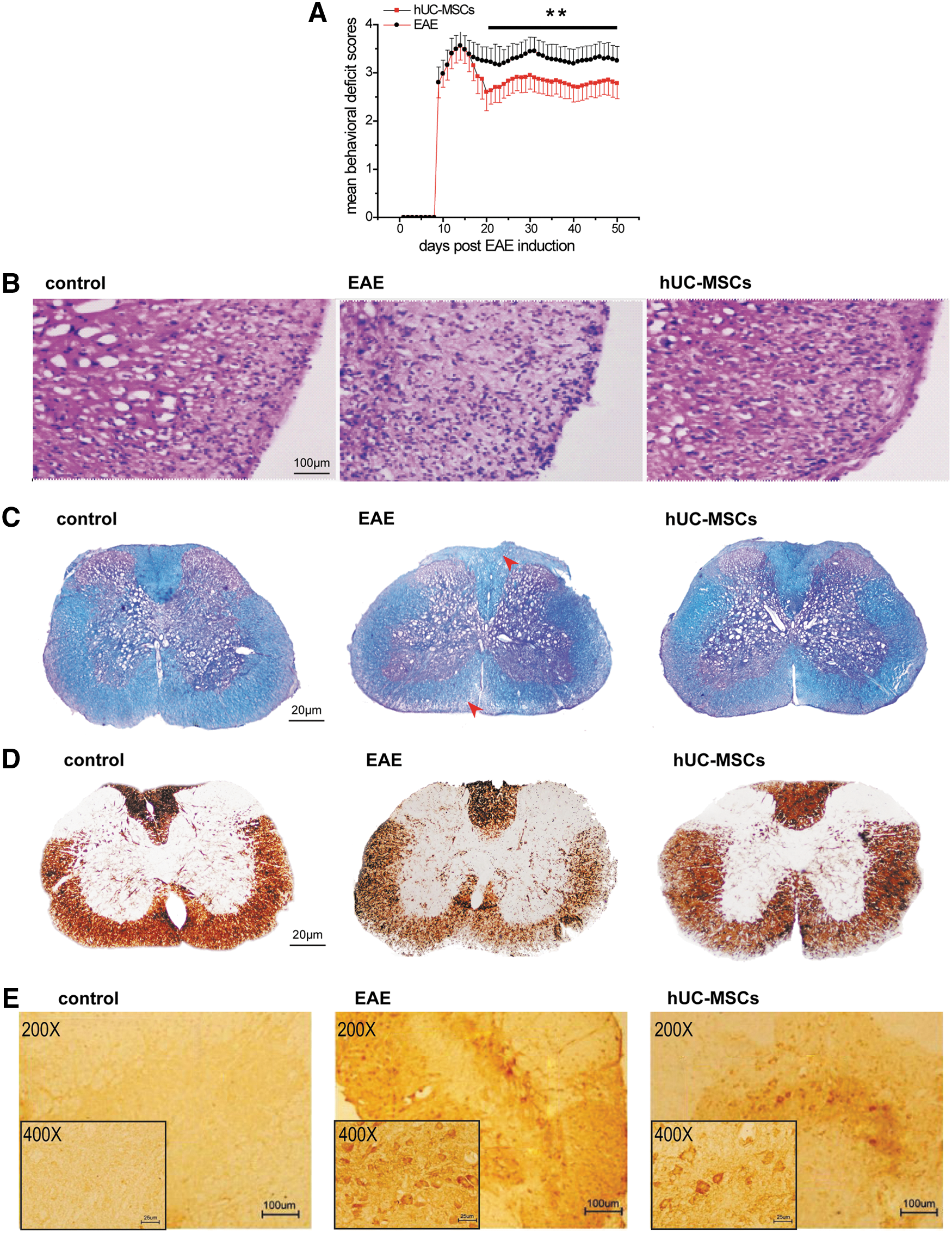
hUC-MSC-ameliorated EAE.
We then confirmed the effect of hUC-MSCs on the pathology of EAE mice. To characterize the influence of hUC-MSCs on inflammatory cell influx, sections of spinal cord from EAE mice were stained with hematoxylin–eosin to identify mononuclear cell and leukocyte infiltration at day 22. The number of perivascular immune cell infiltrations was significantly decreased by 35.50% in the spinal cord of hUC-MSC-transplanted EAE mice, compared with those in the vehicle-treated EAE mice (see Fig. 2B and Table 2, P<0.05). The severity of demyelination and axonal pathology in EAE mice was then assessed. Luxol fast blue staining demonstrated that there was a significant decrease of 64.80% (P<0.01) area of demyelination in hUC-MSC-treated mice, compared to vehicle (Fig. 2C and Table 2). Through Bielschowsky silver and APP staining, there was a 75.04% (P<0.014) and 59.53% (P<0.001) reduction in acute axonal injury in the spinal cord of hUC-MSC-treated EAE mice, compared to vehicle, respectively (Fig. 2D, E and Table 2).
A total of 3–4 mice per group, and 12 spinal cord sections per mouse that covered the whole length of the spinal cord were examined and quantified.
Inflammatory infiltrates: a blood vessel surrounded by 10 nuclei or more was considered as a positive pervascular infiltrate. The total average number of infiltrations per mm2 in spinal cord.
Demyelination and axonal loss were quantified as percentages of areas of demyelination and axonal loss against total white matter area examined by luxol fast blue and silver immunostaining, respectively.
APP positivity was quantified by counting APP-positive axons in a defined quarter of each section and calculated as positive axons per mm2.
P<0.05, significantly different from vehicle-treated control group.
hUC-MSCs, mesenchymal stem cells from human umbilical cord; EAE, encephalomyelitis; APP, amyloid precursor protein.
hUC-MSCs modulate immunoinflammatory response and promote spinal cord remyelination in EAE
Next, we evaluated the immunoregulatory effects of hUC-MSC transplantation on EAE. Because MS involves the T-lymphocyte reaction, Th17CD4+ T cells, CD4+CD25+FOXP3+T (Treg) cells, and IFN-γ/IL-4 ratio, which represents the Th1/Th2 T cell ratio in the spleens, were determined at day 22 by flow cytometry. A substantial decrease of Th17CD4+T cells and IFN-γ/IL-4 ratio was in hUC-MSC-treated EAE mice compared to control-treated EAE mice (4.63%±0.43% vs. 5.6%±0.84%, P<0.05; 1.33±0.02 vs. 1.83±0.04, P<0.05), respectively (Fig. 3A, B). Treg cells in the spleen were markedly enhanced by hUC-MSC treatment compared to the EAE group (15.17%±0.41% vs. 10.9%±0.35%, P<0.05) (Fig. 3A, B).
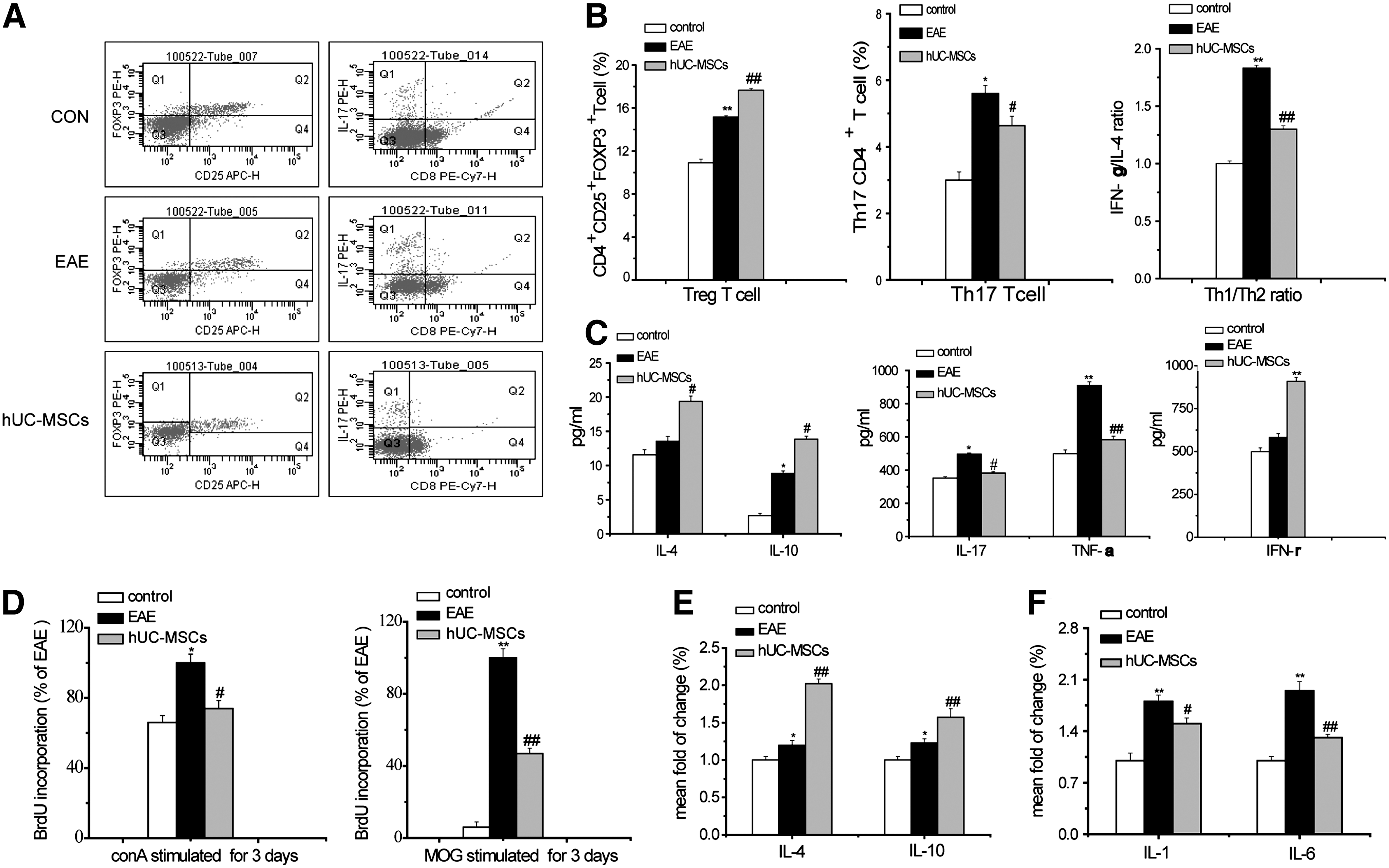
Peripheral immunoregulation effect on EAE of hUC-MSCs. The Th1/Th2 ratio, the proportions of Th17 and Treg cell of spleen cells, were measured by flow cytometric analysis at 22 days after MOG induction
Then, we tested the effect of hUC-MSCs on cytokine production in the supernatant of splenocytes in vitro. The level of anti-inflammatory cytokines IL-4 and IL-10 in hUC-MSC-treated EAE mice reached 142.87% (P<0.05) and 157.37% (P<0.05) of vehicle EAE mice, and the level of proinflammatory cytokines IL-17 and TNF-α was decreased to 64.31% (P<0.05) and 75.70% (P<0.01), compared to control. The results suggested that hUC-MSCs induced a shift from Th1 toward a Th2 phenotype contributing to disease amelioration. However, in agreement with Constantin [18], the basal secretion of IFN-γ by T cells was increased in hUC-MSC-treated EAE mice (Fig. 3C).
Furthermore, we tested whether the immune-modulating properties of hUC-MSCs refer to the generation of effector T cells. FCA of BrdU kit was used in the detection of splenocyte-proliferating cells at day 22 post-EAE induction (acute phase after hUC-MSC transplantation) in vitro. The results indicated that the level of BrdU+ cell was decreased to 46.5% in MOG35-55-stimulated hUC-MSC-treated-EAE mice and to 78.16% in ConA-stimulated hUC-MSC-treated-EAE mice, respectively, compared with that in vehicle-treated EAE mice (Fig. 3D).
A major downstream signaling pathway of immunological reaction is the release cytokines. This study showed that hUC-MSCs significantly increased IL-4 and IL-10 by 65.57% and 27.87%, and decreased the IL-1 and IL-6 by 17.59% and 33.68% of control in the spinal cord, respectively (Fig. 3E, F).
An electron microscope was used to examine whether hUC-MSCs could regenerate the myelin sheath. As shown in Fig. 4A, the normal myelin sheaths were thick and dense in control mice (triangles); in vehicle-treated EAE mice, damaged myelin sheaths often had loose and separated layers or degraded sheath structures (arrows in Fig. 4A). In contrast, hUC-MSC-treated EAE mice showed an abundance of newly formed myelin sheaths that were notably thinner (asterisks in Fig. 4A).
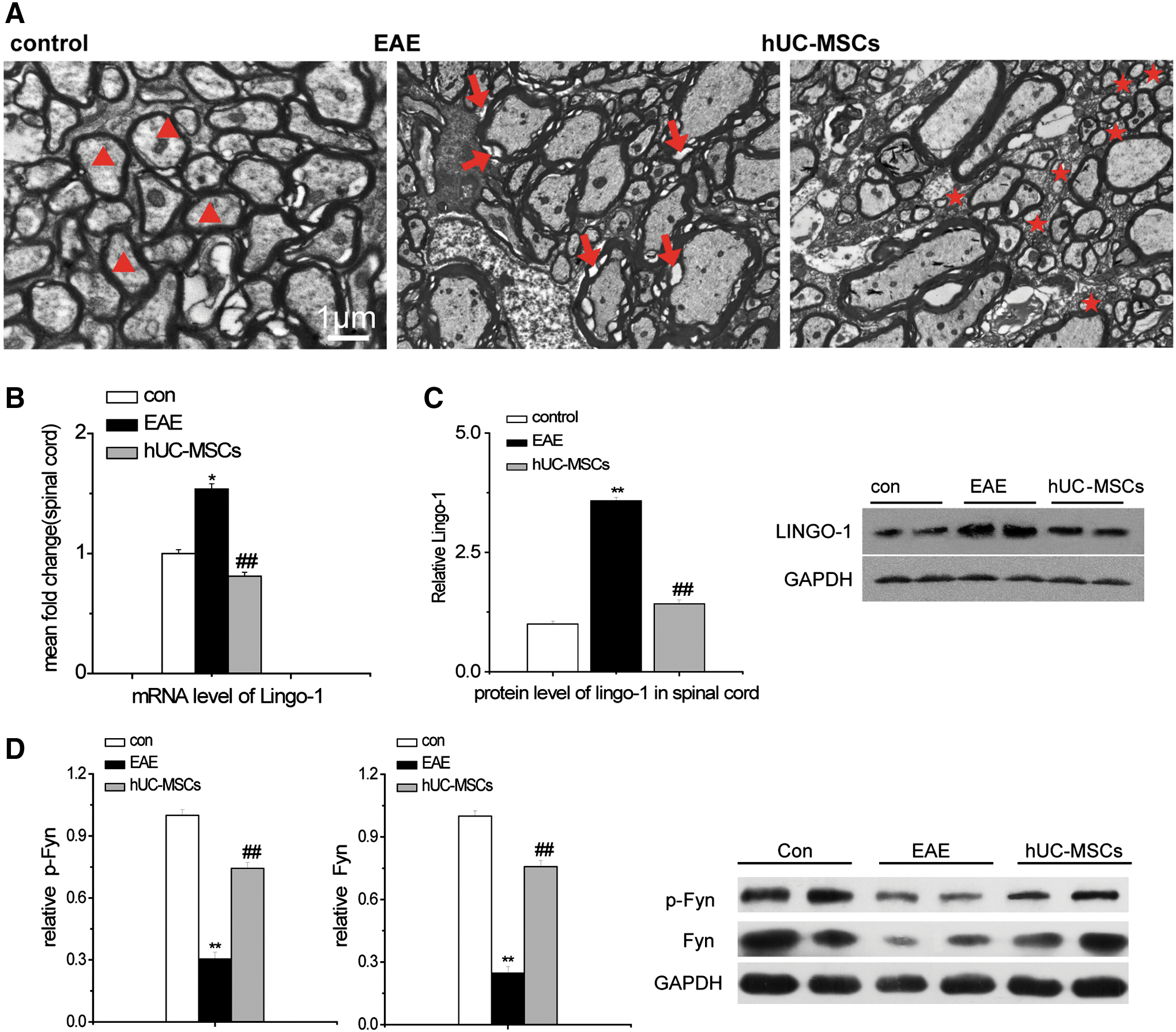
hUC-MSCs promoted remyelination in the spinal cord of EAE mice.
LINGO-1 may suppress oligodendrocyte differentiation and myelination, and the neurological function scores of EAE mice could be recovered with inhibition of LINGO-1 [19]. Here, we tested the effect of hUC-MSCs on the level of mRNA and protein expression of LINGO-1. Real-time PCR and western blot showed that hUC-MSCs significantly lowered the level of mRNA and protein of LINGO-1 in EAE mice (81.20%±4.3% vs. 153.84%±3.3%, P<0.01; 142.38%±8.34% vs. 357.92%±7.53%, P<0.01) than that in vehicle-treated EAE mice (Fig. 4B, C).
LINGO-1 increases the expression and activation of RhoA-GTP through decreasing the expression of Fyn and p-Fyn. RhoA-GTP regulates cytoskeletal reorganization, cell adhesion, and proliferation [20]. This study indicated that the expression of Fyn and p-Fyn was increased in the hUC-MSC-treated EAE group (75.73%±3.14%, 74.25%±2.87%), compared to the vehicle-treated EAE group (24.71%±2.94%, 30.43%±3.05%, P<0.01), respectively (Fig. 4D).
Discussion
This is the first report that hUC-MSCs improve clinical manifestations in the MS animal model of EAE, characterized by areas of inflammation and demyelination throughout the spinal cord. The EAE-induced behavioral impairments were attenuated in animals that received hUC-MSCs as revealed by significant reduction in disease severity at days 22 and 50 post-transplantation. Moreover, this functional recovery was associated by similar amelioration of EAE-induced histopathological deficits as revealed by significantly decreased perivascular immune cell infiltrations, demyelination, and axonal injury in the spinal cord of hUC-MSC-treated EAE mice.
Our previous study verified that hUC-MSCs can cross the blood–brain barrier via intravenous transplantation [21]. However, the efficacy of hUC-MSCs on ataxic mice was related to their release of neuronal growth factors, not differentiation into neuronal cells [21]. This study showed that hUC-MSCs exerted immune modulatory effects by inducing a shift of T cells from Th1 proinflammatory cells to Th2 anti-inflammatory cells. In fact, in acute EAE, like most autoimmune diseases, a shift from Th1-type cells to Th2-type cells does exist to function as self-modularized protection [22], and treatment of MSCs is found to enhance the shift. IL-10 and IL-4, typical cytokines secreted by Th2 cells, are increased in the hUC-MSC-treated group, in accordance with the earlier study of human derived BM-MSCs in chronic EAE [23]. Relevant to the present immunomodulatory effects of hUC-MSCs, the previous experiments demonstrated a decreased expression of IFN-γ, mostly produced by Th1 cells in stem cell-treated mice [22]; our experiments, as well as those of Constantin et al. [18], confirmed that the level of IFN-γ was increased in MSC-treated mice. This phenomenon sheds new light on the dual function of IFN-γ, as previous findings have shown that IFN-γ-deficient mice are more susceptible to MOG-induced EAE and have higher mortality rates [24]. Thus, IFN-γ may not only shape this autoimmune disease but also exert a protective role by reducing activation, proliferation, and infiltration of neutrophils. In addition, recent studies have described an important role of Th17 T cells in autoimmune disease [25]. In this study, the production of IL-17 was significantly decreased in the stem cell-treated group and almost reached the normal level, providing a profound evidence of reduced Th17 cells or/and suppressed Th17 cytokine secretion. Generally, hUC-MSCs decrease inflammatory response by reaching a balance of the immune system.
Promoting remyelination remains a major challenge in the treatment of MS and is of critical importance in the perspective of future application. Cell-based therapies may boost remyelination in addition to the stem cells' anti-inflammatory and trophic factor secretory properties, which altogether may promote endogenous cell regeneration [12]. However, the direct role of stem cells in affording therapeutic effects in response to autoimmune injury and demyelination remains not well defined. Meanwhile, LINGO-1 selectively expressed on oligodendrocyte precursor cells, and neuron is a key negative factor regulating oligodendrocyte differentiation and myelination. Recent studies have shown that loss of LINGO-1 function by either LINGO-1 gene knockout or treatment with antagonist resulted in functional recovery of EAE [19,20,26]. In this study, LINGO-1 mRNA and protein expression was downregulated in the spinal cord of hUC-MSC-treated EAE mice compared to those treated with vehicle alone.
LINGO-1, a transmembrane glycoprotein, has a leucine-rich repeat (LRR) and an immunoglobulin domain that forms A kind of complexes, including NgR1/p75 and NgR1/Taj (Troy). The complexes inhibit axon growth by activating RhoA in the presence of myelin inhibitors in neurons [27,28]. Moreover, they block LINGO-1-associated oligodendrocyte differentiation and axon myelination in oligodendrocytes. The 2 downstream signal molecules of LINGO-1 are Fyn and RhoA [20]. As one of the best-known members of Rho GTPases, RhoA activates and recruits macrophages/microglia and directs the migration of these proinflammatory cells to the lesioned tissue, defining a critical cell death cascade mediating the deterioration of MS/EAE [29]. RhoA also exerts cytotoxic effects, such as aberrant accumulation of reactive oxygen species, active inflammatory response, and increased intracellular apoptosis [30]. Fyn promotes the differentiation of oligodendrocyte by activating p190 RhoAGAP, which decreases RhoA activity [20]. Consistent with these observations, our study showed that hUC-MSC treatment downregulated LINGO-1 levels from mRNA to protein and increased Fyn expression and phosphorylation, indicating that RhoA activity may be reduced by increased Fyn expression and phosphorylation. We advance the notion here that hUC-MSCs regulate the LINGO-1-signaling pathway and its downstream proteins Fyn and RhoA in facilitating remyelination.
Conclusion
In summary, this study provides the scientific basis for proceeding with the clinical application of hUC-MSC transplantation in patients with MS. The present laboratory evidence indicates long-term (ie, 50 days) recovery of behavioral functions and improvement of histopathological symptoms, including suppression of perivascular immune cell infiltrations and reduction in both demyelination and axonal injury in the spinal cord of hUC-MSC-treated EAE mice. The novel finding from this laboratory is the observation of multipronged, but convergent, regenerative pathways involving immunomodulation and remyelination that likely mediate the beneficial outcomes of hUC-MSCs. In particular, the pivotal role of LINGO-1 as the key signaling protein mediating both immunomodulatory and myelin-inducing mechanisms offers new venues on further optimizing the therapeutic effects of hUC-MSC transplantation in MS and other disorders characterized by aberrant immune and myelin dysfunctions.
Footnotes
Acknowledgments
This study was supported by the National Natural Science Foundation of China (81171085, 30971010, 81100863, and 81200876), the Medical Leading Talent and Innovation Team Project of Jiangsu Province (LJ201101), the Natural Science Foundation (BK2009037) of Jiangsu Province of China, Science and Technology Achievement transformation Foundation of Jiangsu Province (BA2009124) and Medical Technology Development Project of Nanjing (ZKX09022, ZKX08030).
Author Disclosure Statement
All authors declare that there are no conflicts of interest.