
Abstract
The immortal strand theory postulates stem cells protect themselves from DNA replication-associated mutations and subsequent cancer risk through selective segregation of template DNA strands. Stem cells self-renew by asymmetric cellular division. During asymmetric division, stem cells maintain their template DNA strands, while the newly synthesized DNA strands segregate to newly formed daughter cells. Previous studies have demonstrated that self-renewing mammary stem cells originate in the expanding mammary ducts during puberty-associated allometric growth. In this study, we labeled newly forming mammary stem cells with the thymidine analog 5-ethynl-2′-deoxyuridine for 2 weeks during allometric ductal expansion. Cells that incorporate and retain the nuclear label following extended chase periods are termed label-retaining cells (LRCs). A second nuclear label, 5-bromodeoxyuridine, was administered before euthanasia to identify cells traversing the cell cycle. Mammary cells collected following euthanasia were sorted based on nuclear label retention. Members of the Notch and Wnt signaling pathways were found differentially expressed by mammary LRCs. These pathways are involved in the regulation of stem cells in the mouse mammary gland. Upon further analysis, we found that in contrast to non-LRCs, Notch1 and Notch2 are expressed and localized in the nuclei of the LRCs. Expression of Notch-inducible genes, Hes1 and Hey2, was elevated in LRCs. Inhibition of Notch1 by shRNA reduced colony forming potential and label retention by mammary epithelial cells in vitro. These results indicate that genes are differentially regulated in the LRC population of mammary glands and Notch1 mediates asymmetric cell division of mammary progenitor cells.
Introduction
In the mouse mammary gland, label-retaining cells (LRCs) have been identified in the developing ducts during pubertal expansion, in the alveolar structures that form during pregnancy, and in the surrounding mammary stroma and blood vessels. The mammary LRCs are presumed to be mammary stem and progenitor cells [4,5,7]. A fraction of mouse mammary LRCs continue to cycle as demonstrated by their uptake of the second DNA label [4,5,7]. This process is not limited to mice, as LRCs have also been identified in mammary glands of immature bovine calves [9]. LRCs have been identified in premalignant mammary epithelial cell populations and gastrointestinal cancers [10,11] indicating that asymmetric division can be a characteristic of cancer stem cells.
The presence of resident stem cells in the mouse mammary gland has been confirmed. Studies have demonstrated clearly that the mammary gland has regenerative capacity when transplanted and that single cells isolated from a gland can regenerate an entire gland [Reviewed in Refs. 12,13]. Undifferentiated mammary stem cells have been identified in suprabasal positions between the luminal and myoepithelial cell layers based on morphological characteristics in rats and mice [14] and cells with similar characteristics have been found in human breasts [15,16].
We have demonstrated previously that LRCs are maintained in the mouse mammary gland through pubertal development and the population expands during pregnancy [5,7]. The mechanism employed by LRCs to differentiate between symmetric and asymmetric division is currently unknown. Here we investigated the gene expression profiles of LRCs and other non-LRC epithelial cells of the mouse mammary gland after a chase period following initial exposure to nucleic acid label.
Materials and Methods
Experimental plan
We carried out a procedure designed to label the DNA of proliferating cells, including symmetrically dividing stem cells, during the period of ductal elongation and development associated with pubertal expansion of the mouse mammary gland. Twelve Balb/c female mice were used for each experiment. Cycling mammary cells were labeled by the administration of EdU (250 μg; Invitrogen, Grand Island, NY) by intraperitoneal (i.p.) injection for 14 days starting on the first day of the 4th week of postnatal life (Fig. 1). This dose has been reported to be effective in a labeling dividing cell in vivo [17]. The label was chased from the glands for 2- or 3-month periods following the final injection to allow completion of the ductal development. At 2 months of age, 6 mice received i.p. injections of 5BrdU (100 mg/mL per g of body wt) 4 h before euthanasia. The inguinal and thoracic mammary glands were removed and dissociated for cell sorting. At 3 months of age, the remaining 6 mice received i.p. injections of 5BrdU. The 5BrdU recipients were sacrificed 4 h after the 5BrdU injection. The inguinal and thoracic mammary glands were removed and bisected with half of the inguinal gland fixed for paraffin embedding and the remaining half dissociated for cell sorting. The entire experiment was repeated twice.
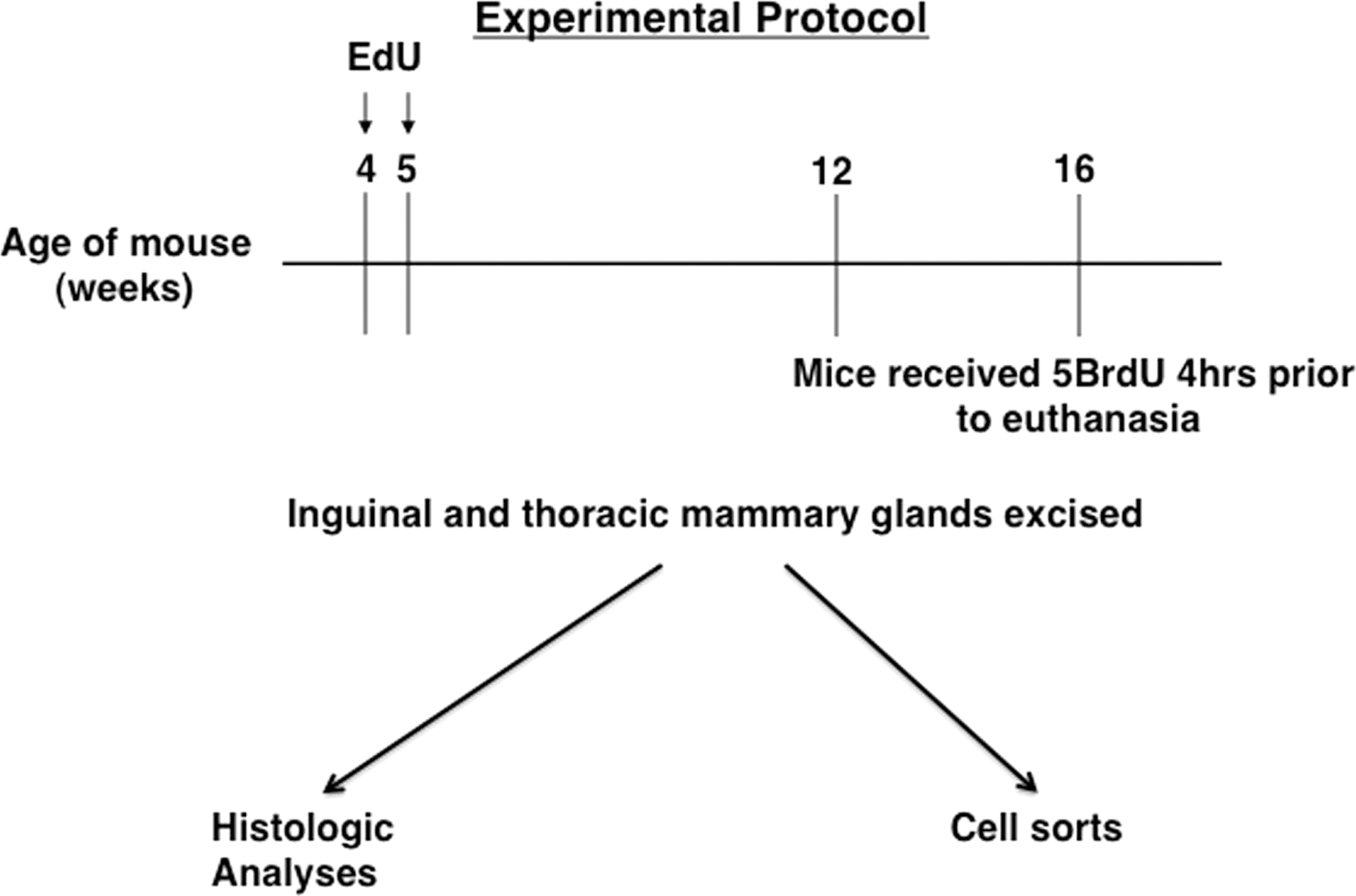
Schematic illustrating the experimental protocol. Mice received 5-ethynl-2′-deoxyuridine (EdU) intraperitoneal during the 4th and 5th weeks of life. Chase periods followed until either the 12th or 16th week when half of the mice received 5-bromodeoxyuridine (5BrdU) 4 h before euthanasia. The thoracic and inguinal mammary glands were excised, bisected with half of the gland fixed for histology, and the remaining half dissociated for cell sorting.
The protocols and procedures used to perform the experiments upon the animals were reviewed and approved by the Animal Care and Use Committee of Clemson University. Housing and care during the experimental period conformed to the guidelines provided by the National Institutes of Health.
Mammary dissociation and cell sorting
Mature mammary glands were excised from mice following treatment outlined above. One half of each gland was processed to produce single-cell suspensions for cell sorting. The cell sorting procedure used is based on sorting protocols of the mouse mammary gland published previously [17 –21]. Briefly, the glands were minced and then dissociated in 1× collagenase/hyaluronidase (1 mg/mL; StemCell Technologies, Vancouver, BC) in growth media containing 5% fetal bovine serum (FBS) and mammary epithelial growth medium (MEGM) Singlequots (Lonza, Walkersville, MD) at 37°C for 7 h. The gland/collagenase suspension was collected, resuspended, and incubated in 0.25% trypsin for 2 min at 37°C. The trypsin was neutralized and cells pelleted. The pellet was then gently resuspended in dispase (StemCell Technologies) and DNase (StemCell Technologies) and incubated in 37°C for 5 min. Hank's balanced solution with 5% FBS was added to the suspension and filtered through a 0.4-μm cell strainer creating single cells in suspension. These single cells were pelleted again and then resuspended in the RBS lysis solution to remove red blood cells. The cell suspension was pelleted and washed in phosphate-buffered saline (PBS). The cells were resuspended and fixed in cold 70% ethanol overnight.
EdU incorporation and retention was detected using Click-iT EdU AlexaFluor488 Imaging Kits (Invitrogen) according to included instructions. Cells that incorporated 5BrdU were identified using anti-5BrdU (Invitrogen; 1:50) conjugated with Alexa-594 for 1 h at room temperature (RT).
The cells were sorted using a MoFlo cell sorter (BeckmanCoulter, Brea, CA). The cells were sorted into 4 populations: EdU+5BrdU+, EdU+5BrdU−, EdU−5BrdU+, and EdU−5BrdU−.
RNA isolation and PCR array
Before RNA isolation, the sorted cell populations were pooled based on expression of nucleic acid labeling. The cells were pooled to increase the RNA yield. Total RNA was isolated from the sorted samples using an RNAaqueous Micro kit (Ambion, Austin, TX) that utilizes glass fiber filter column purification. Isolated total RNA was then treated with DNase (Qiagen, Valencia, CA) to remove any DNA contamination. The concentration of RNA was determined using a NanoDrop and the RNA integrity was analyzed with a RNA nanochip on a Bioanalyzer (Agilent Technologies, Santa Clara, CA). cDNA synthesis and amplification were performed using a RT2 PreAMP cDNA synthesis kit (Qiagen) and RT2 PreAMP Pathway Primer Mix (Qiagen) specific to the RT2 Profiler PCR Array Stem Cell Kit or the Mouse Stem Cell Signaling RT2 Profiler PCR Array Kit that was used (Qiagen). Once the cDNA was synthesized and amplified, it was mixed with RT2 SYBER Green mastermix and loaded on to a 96-well plate provided in the array kits. Each 96-well plate contained 84 genes associated with stem cells and 12 controls (housekeeping genes, genomic DNA contamination, reverse transcription, and positive PCR). The plate was run on an Eppendorf Realplex real-time machine. The data obtained were analyzed by uploading the data on to the Qiagen's data analysis website.
Cell culture
The normal mouse mammary cell line, HC-11, was maintained using Dulbecco's modified Eagle's medium (DMEM) plus 10% FBS, 1% antibiotic/antimycotic supplemented with MEGM SingleQuots (Lonza, Allendale, NJ). All cell cultures were maintained at 37°C with 5% CO2.
shRNA transfections
HC-11 mouse mammary cells were transfected and selected using commercially available shRNA that specifically targets Notch1 or scrambled control shRNA according to the manufacturer's protocol (Santa Cruz Biotechnology, Santa Cruz, CA).
Quantitative PCR
Quantitative PCR (qPCR) was performed as previously described [22]. Briefly, total RNA was isolated and concentration and integrity was determined as described above. cDNA synthesis was performed using Superscript II reverse transcriptase (Invitrogen). The cDNA was subjected to PCR amplification using TaqMan Universal PCR Master Mix reagents (Applied Biosystems, Foster City, CA). qPCR primers were from Applied Biosystems. For each gene, a standard curve was generated using a specific cDNA clone and qPCR efficiency was calculated from the slope of the standard curve. The standard curve slope and amplification plots were analyzed using MxPro PCR Software (Stratagene, Santa Clara, CA) and the relative abundance of target mRNA was calculated as the ratio of the copy number normalized to the copy number of Gapdh mRNA. Reactions were performed in a MX3000 multiplex qPCR system (Stratagene).
Matrigel colony-forming assay
Following shRNA transfections, untreated HC-11, HC-11 cells transfected with Notch1-specific shRNA, and HC-11 cells that received control shRNA were seeded onto growth factor-reduced Matrigel (BD Biosciences, Franklin Lakes, NJ) layers in 12-well plates at clonal density. The cultures received EdU for 7 days. The EdU was removed and the cultures were allowed to grow for 4 weeks.
Immunodetection studies
For immunodetection studies, 5–6-μm sections were cut of paraffin-embedded mammary glands and the sections were deparaffinized in xylenes and rehydrated through a graded series of ethanols ending in deionized water (diH2O). EdU was detected using a Click-iT EdU AlexaFluor488 Imaging Kit according to enclosed instructions (Invitrogen). Sections were blocked with 10% goat serum for 30 min at RT and then washed with PBS. Primary antibodies [anti-Notch1 (1:100), anti-Notch2 (1:400); Santa Cruz Biotechnology, or anti-5BrdU (1:50), Invitrogen] were applied overnight at 4°C. Sections were washed with PBS and Alexa-594-conjugated secondary antibodies (Invitrogen; 1:200) were applied for 30 min at RT. Sections were washed with PBS, rinsed with diH2O, and coverslipped using ProLong mounting media with DAPI (Invitrogen).
Staining of cells grown on Matrigel consisted of washing gels with PBS, and then fixing cultures with cold 70% EtOH. EdU was detected using a Click-iT EdU AlexaFluor488 Imaging Kit. The colonies were then blocked with 10% goat serum for 30 min at RT and then washed with PBS. Anti-Notch1 was applied followed by the Alexa-594-conjugated secondary antibody. Nuclei were stained with Hoechst 33342 (Invitrogen).
All images were captured using a Zeiss Axiovert 40, and the colony size was determined using the AxioVision Software (Zeiss, Thornwood, NY). Images were processed using Adobe Photoshop.
Western blotting
Protein lysates were collected using the M-PER Mammalian Protein Extraction Reagent with Halt Protease Inhibitor Cocktail added (Thermo Scientific, Rockford, IL). Protein concentrations were determined using a BCA Protein Assay Kit (Thermo Scientific). Collected lysates were boiled with a sample buffer for 5 min, and then separated via SDS-PAGE using a 4%–15% precast HCl-Tris gel (BioRad, Hercules, CA). The proteins were transferred to the nitrocellulose membrane that was blocked with 5% casein in PBS for 30 min at RT. Membranes were incubated overnight at 4°C with primary antibodies [anti-Notch1 (1:1,000) (Santa Cruz Biotechnology) or anti-β-actin (1:1,000) (Cell Signaling Technology, Danvers, MA)]. The membranes were washed twice with Tris-buffered saline with 0.05% Tween-20 (TBST), and then incubated with the HRP- conjugated secondary antibody (1:2,500; Cell Signaling Technology) for 2 h at RT. Blots were washed again with TBST, rinsed with diH2O, and developed with the LumiGLO Reagent (Cell Signaling Technology). Images were captured and analyzed using a FluorChemM (Cell Biosciences, Santa Clara, CA) and accompanying software.
Reverse transcriptase-PCR
Reverse transcriptase-PCR was done with Superscript III One Step RT-PCR System with Platinum Taq DNA Polymerase (Life Technologies, Grand Island, NY) according to the manufacturer's protocol except for the cycle number of 25 instead of 40. Primers used were from the shRNA kit for Notch1 (Santa Cruz Biotechnology). The bands were visualized on a 1.2% agarose gel with ethidium bromide.
Statistical analyses
Colony forming data were analyzed using the Student's t-test. Data were considered significant at P<0.05. Representative data are presented as means±standard deviation.
Results
Identification of LRCs
Following the labeling protocol outlined above, mammary sections were probed for EdU and 5BrdU. Cells that contained EdU are considered LRCs, while those that contain 5BrdU are considered to have been in cycle at the time of euthanasia. Figure 2 shows cells within mammary glands containing EdU (Fig. 2A), 5BrdU (Fig. 2B), or both nuclear labels (Fig. 2C). This confirms the presence LRCs (EdU+ cells), cycling cells (5BrdU+ cells), and cycling LRCs (EdU+5BrdU+ cells) within mammary glands treated with our labeling protocol. Figure 2B shows a cell undergoing symmetric division as the incorporated 5BrdU is being segregated evenly between the 2 cells. A fraction of the mammary LRC population has been demonstrated to be traversing the cell cycle [4]. Previous studies have also demonstrated that populations of luminal LRCs express the estrogen receptor (ER)α, progesterone receptor, and cytokeratins, while basal LRCs express smooth muscle actin [4,5,7].
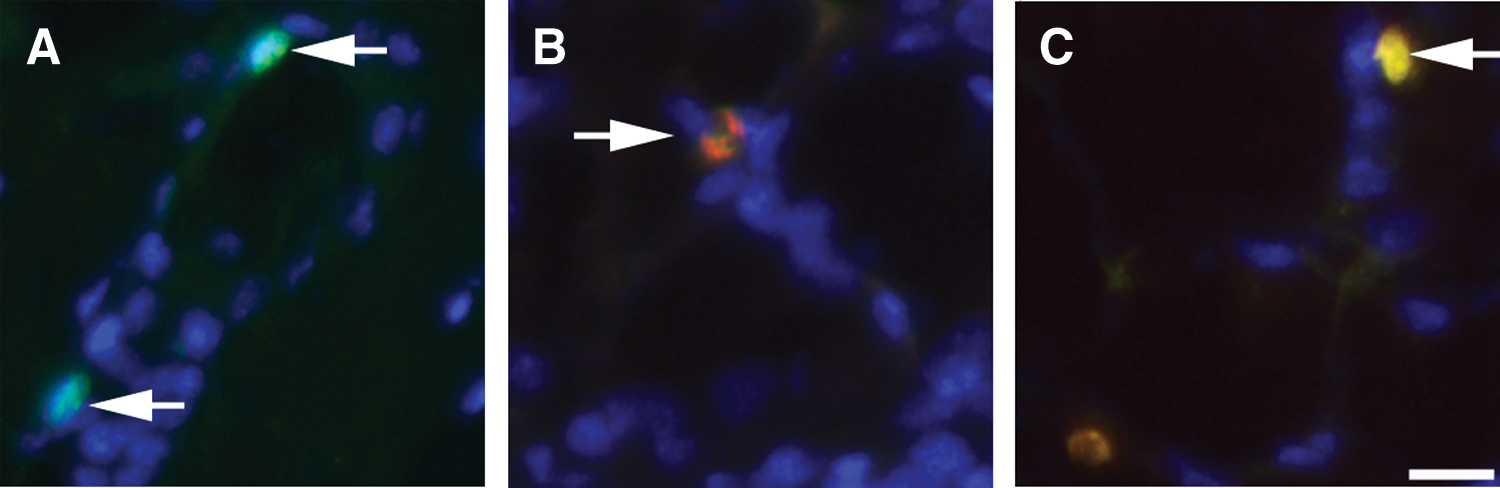
Presence of nuclear label-retaining cells (LRCs). Female mice received EdU for 2 weeks before a 3-month chase period at which time they received 5BrdU 4 h before euthanasia.
Isolation and gene expression of LRCs
Single-cell suspensions of pooled mammary glands were sorted into 4 populations, EdU+5BrdU+, EdU+5BrdU−, EdU−5BrdU+, and EdU−5BrdU−. The populations that are EdU+ are LRCs, while the EdU+5BrdU+ population is made up of cycling LRCs. The results of the sorts are displayed in Supplementary Fig. S1 (Supplementary Data are available online at
Four mammary glands from each mouse in the group for a total of 24 glands were harvested, dissociated, and sorted into 4 populations. % Sorted indicates the percent of the total sorted population±standard deviation.
EdU, 5-ethynl-2′-deoxyuridine; 5BrdU, 5-bromodeoxyuridine.
Gene expression profiling was then run on the 4 sorted samples as outlined in the Materials and Methods section. A total of 168 genes were investigated. The results are listed in Supplementary Tables S1–S3. Genes from 2 known stem cell signaling associated pathways, Notch and Wnt, were differentially expressed between the populations of sorted cells compared to the EdU−5BrdU− population. In the quiescent LRC EdU+5BrdU− population, we found expression of Notch1 and Notch2 elevated, while the Notch ligands Jagged1 and Delta1 were downregulated (Table 2). The genes of the Wnt signaling pathway we found differentially regulated in the EdU+5BrdU− population are Wnt1 and disheveled-1 (Dvl1). The TGFβ family members BMP1 and BMP3 were also downregulated. Other genes of note differentially expressed between the EdU+5BrdU− population and the EdU−5BrdU− population include the gap junction protein alpha 1 (Gja1), Cyclin D1 (Ccnd1), histone deacetylase 1 (Hdac1), and myelocytomatosis oncogene (Myc). All 4 were found downregulated in the EdU+5BrdU− population. Each of these 4 genes is involved in cell division.
Fold change compared to gene expression of EdU−5BrdU− sorted population.
LRCs express Notch proteins and have elevated levels of Notch-inducible gene mRNA
To confirm that LRCs were expressing the proteins of the differentially expressed genes identified in Table 2, we performed immunodetection for selected proteins from the list. Figure 3 demonstrates that LRCs, as determined by positive staining for EdU, express Notch1 (Fig. 3A) and Notch2 (Fig. 3B). The Notch expressing epithelial LRCs were in basal and suprabasal positions, not luminal, along mammary ducts, and in primitive acinar structures (white arrows in Fig. 3). Mammary stem cells are positioned in such locations [14,23]. Interestingly the Notch staining in the LRCs was nuclear in contrast to non-LRCs where Notch was detected on the periphery of cells. Nuclear localization of Notch indicates activation and translocation of the intracellular domains of the Notch receptors [24]. Nonepithelial LRCs express Notch2 on the cell surface, but not in the nucleus (green arrow Fig. 3B). LRCs have been identified previously in mammary-associated blood vessels, nerves, and stroma [7].
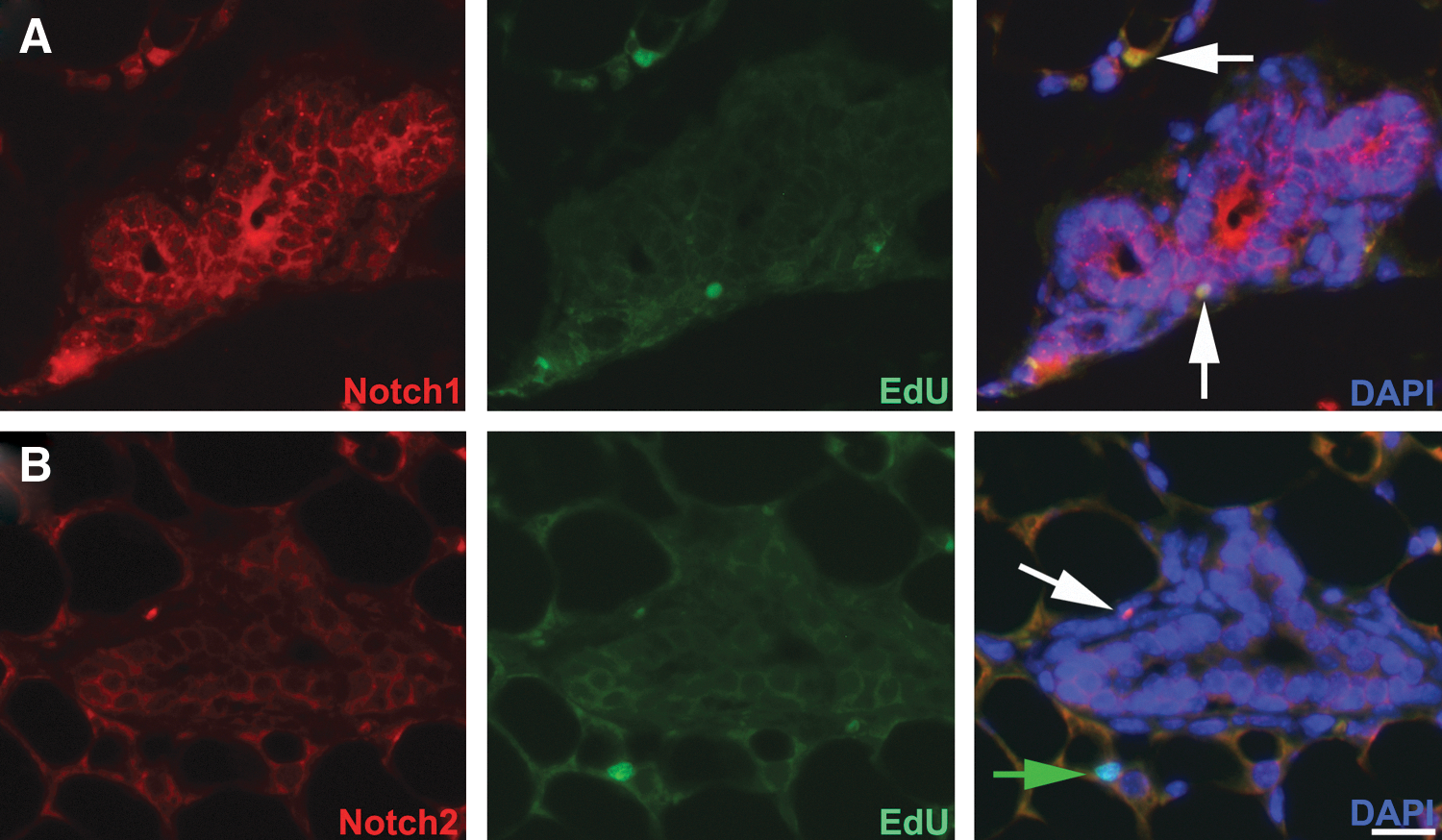
Mammary LRCs express Notch isoforms. After a 3-month chase period following EdU administration, mammary sections were probed for the presence of LRCs and Notch isoforms. Mammary epithelial cells express both
We next measured the gene expression levels of 2 Notch-inducible genes, Hes1 and Hey2, in LRCs. qPCR was performed on samples obtained from the sorted cell populations. Expression of both Hes1 and Hey2 was significantly elevated in the EdU+5BrdU+ population compared to the other 3 populations of cells (Fig. 4). This result indicates that within the EdU+5BrdU+ population of LRCs the Notch signaling pathway is active
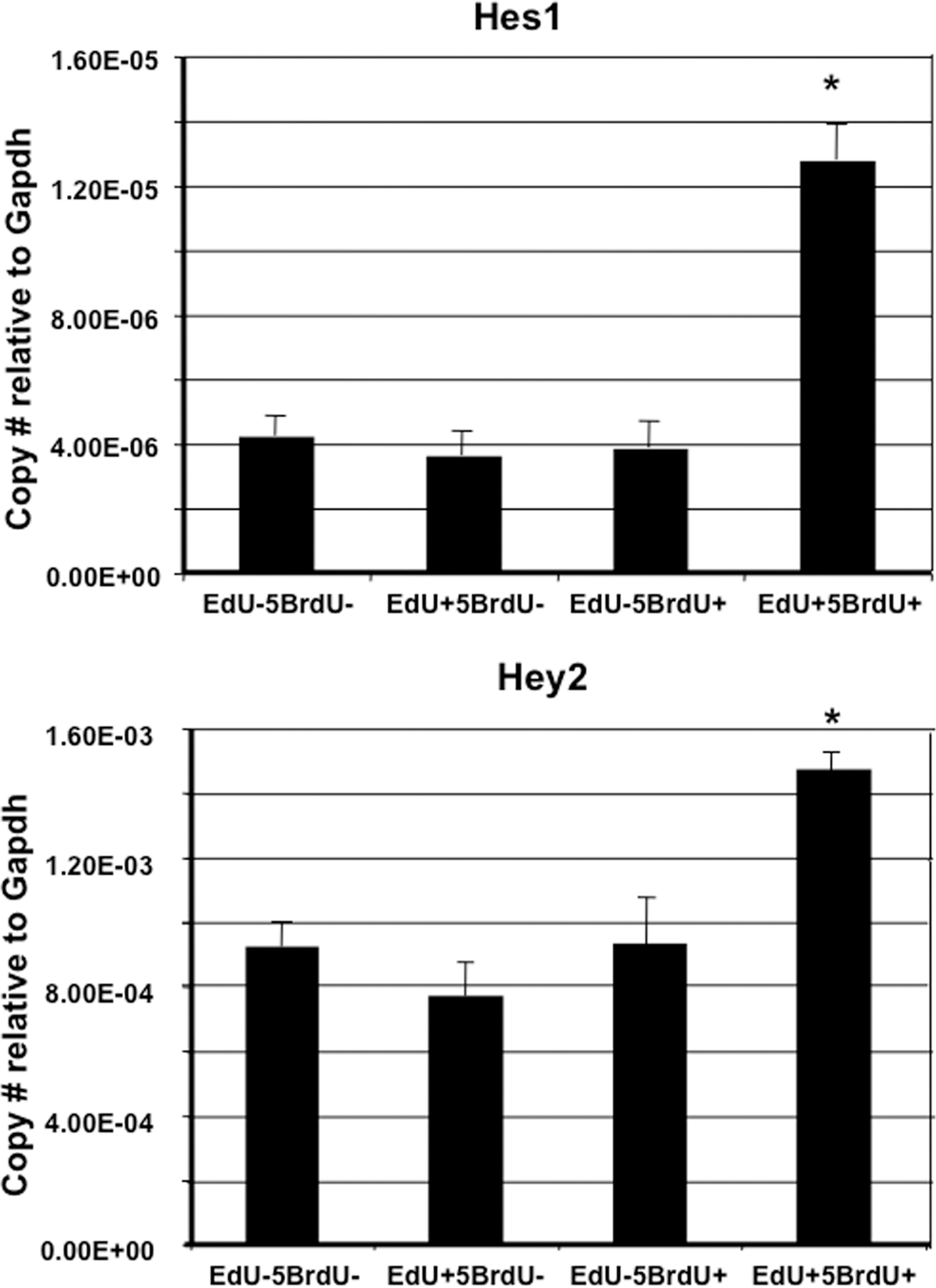
Activation of Notch-inducible genes in LRCs. Real-time PCR results demonstrating that cells within the sorted EdU+5BrdU+ population express significantly elevated levels of Hes1 and Hey2 compared to the other 3 populations. *P<0.05 versus each population.
Notch1 influences nuclear label retention in vitro
To examine the effects of Notch on mammary progenitor cells, we used shRNA to directly inhibit the Notch1 activity. The normal mouse mammary epithelial cell line, HC-11, was used for studies involving Notch shRNA [22,25]. The HC-11 cell line is a prolactin-responsive clone of COMMA-D (CD) cells [26]. We have previously demonstrated that the normal CD cell line, derived from a midpregnant Balb/c mouse [27], forms mammospheres, an in vitro function of stem cells, equivalent to primary mammary epithelial cells, and that only 1–2 cells/mammosphere are LRCs [28]. Specific shRNA that targets Notch1 diminished expression of the Notch1 protein and mRNA levels in the HC-11 cells (Supplementary Fig. S2). Both control HC-11 cells and Notch1 shRNA-treated HC-11 cells were grown on growth factor-reduced Matrigel (Fig. 5A, B). HC-11 cells transfected with Notch1-specific shRNA formed significantly fewer colonies than the untreated HC-11 cells after 7 days in culture (Fig. 5C). The transfected HC-11 colonies were significantly smaller than the colonies formed by untreated HC-11 cells (Fig. 5D).
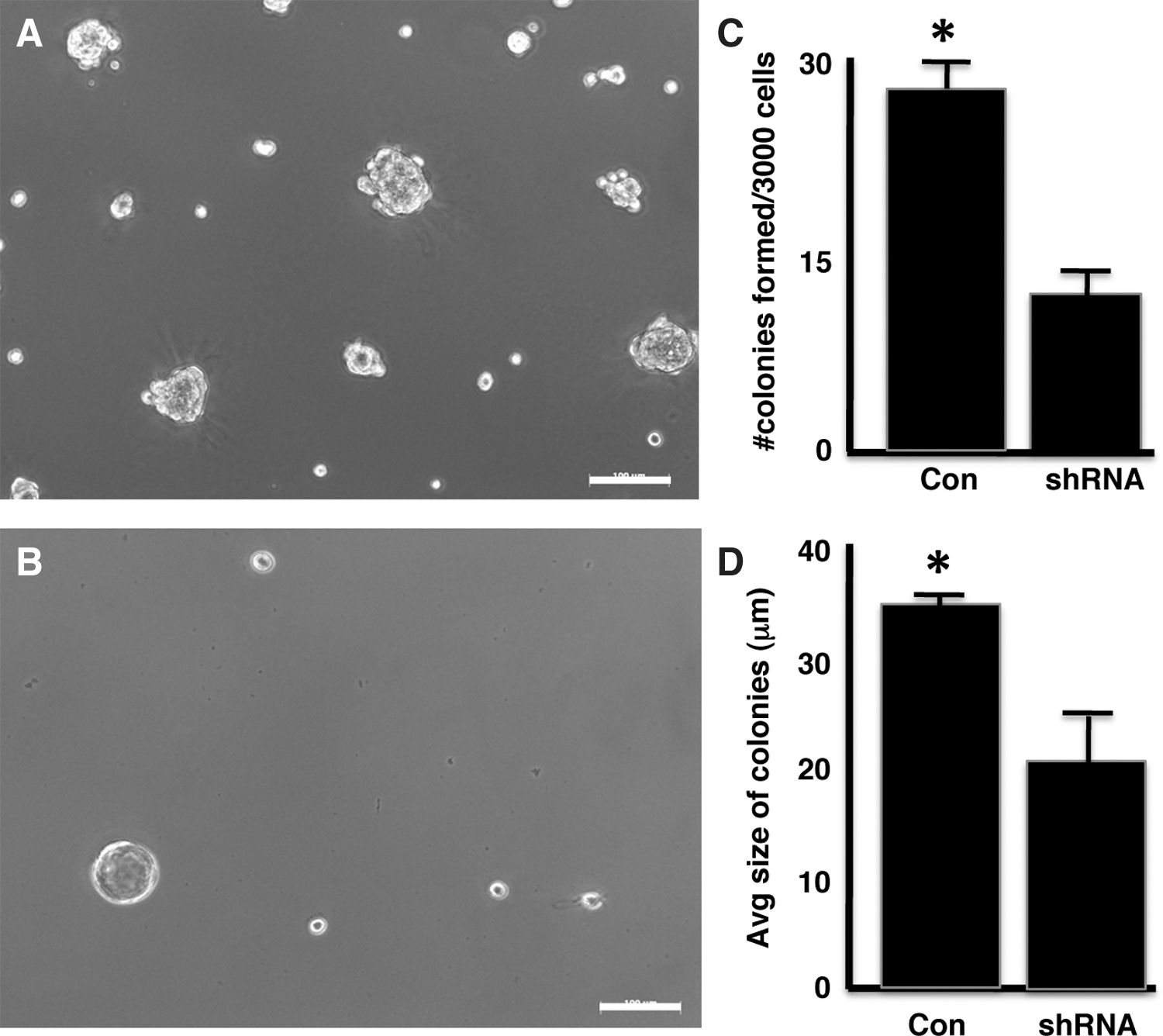
Reduction of Notch1 by shRNA reduces colony-forming capacity of HC-11 cells. HC-11 cells transfected with
We next investigated the role of Notch1 in asymmetric divisions of HC-11 mouse mammary epithelial cells. After a 7-day chase period following EdU exposure, the majority of cells in colonies formed on growth factor-reduced Matrigel by both transfected and control HC-11 cells contained EdU (Fig. 6A, B). Transfection with Notch1 shRNA reduced the total number of colonies formed by HC-11 cells, while simultaneously reducing the number of EdU+ colonies after a 28-day chase period (Table 3). Colonies formed by control HC-11 cells contained at least one cell that was EdU+ (Fig. 6C), while the colonies formed by Notch1 shRNA- transfected HC-11 cells did not contain EdU+ cells (Fig. 6D). The cells in the colonies that contained EdU after the 28-day chase period also expressed Notch1 (Fig. 6C). The few EdU+colonies formed by transfected HC-11 cells, 1.8% of the total number of colonies formed, also expressed Notch1 (Table 3). The knockdown efficiency of the shRNA was not 100% (Supplementary Fig. S2). The protein and gene expression is reduced, but not completely blocked by the shRNA. The EdU+Notch1+ colonies present are believed to arise from cells in which the shRNA was not 100% effective.
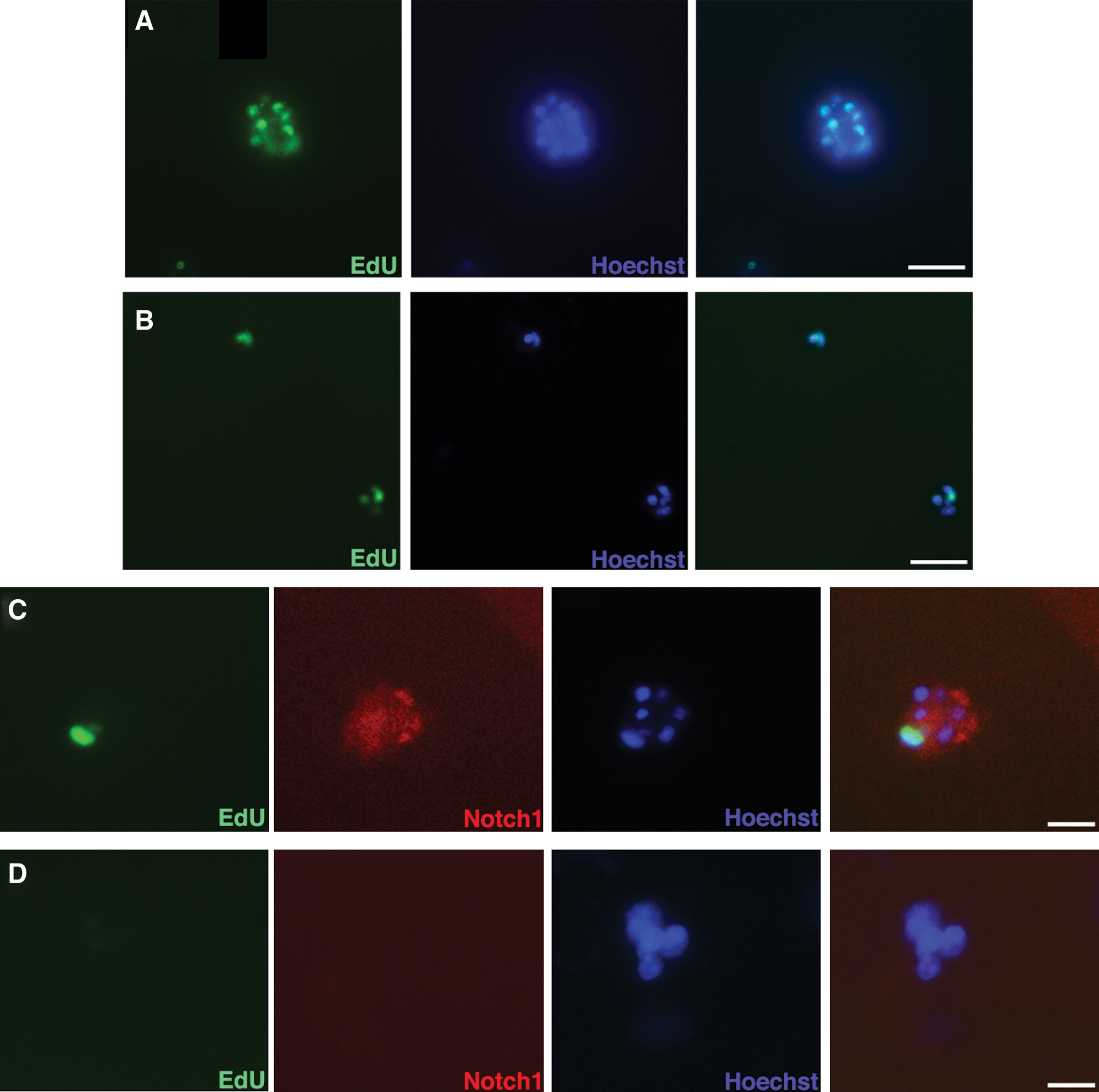
Reduction of Notch1 by shRNA reduces LRC population of HC-11 cells. Untreated and shRNA-transfected HC-11 cells were grown on GF-Matrigel with EdU added for the first 7 days of culture. After a 7-day chase period, both
Discussion
The fact that cells and tissue fragments from any portion of the mouse mammary gland can repopulate a mammary fat pad cleared of endogenous epithelium in juvenile female recipients regardless of age of the tissue transplanted is an evidence of the presence of mammary stem cells [12,13,29 –31]. Additional functional studies have gone on to demonstrate that a single mammary epithelial cell can self-renew and contribute to the regeneration of a functional mammary gland [18,19,32]. Currently, mammary epithelial stem cells are identified and collected based on cell surface markers.
Somatic stem cells are believed to protect themselves and the organism from DNA replication-associated mutations through selective segregation of template DNA strands during asymmetric cellular divisions [1]. Following DNA replication, newly formed DNA strands are segregated to the newly formed daughter cells thus passing any acquired mutation to the short-lived differentiating cells. Asymmetrically dividing cells have been identified and characterized through nuclear label incorporation and retention in developing and pregnant mouse mammary glands [4,5,7]. Functional demonstration that LRCs are in fact stem cells has not been accomplished due to technical limitations where viable sorts of LRCs are not possible as the detection of nuclear label kills the cells.
In this study, we identified and isolated mature mouse mammary LRCs (EdU+) and compared the expression of a limited number of genes in LRCs that were in cycle (EdU+5BrdU+) to those LRCs not in cycle (EdU+5BrdU−) and to mammary non-LRCs (EdU−). The gene expression profile of the EdU−5BrdU− population was used as the baseline for comparisons. Genes involved in the Notch, Wnt, and TGFβ pathways were identified as being differentially expressed. All 3 pathways have been implicated in mammary stem cell function [Reviewed in Refs. 33 –37]. Our methodology ensured that those cells analyzed for expression were predominantly epithelial [17 –21] and, furthermore, the genes we found differentially expressed in our studies have been previously shown to be differentially expressed in stem cells of not only the mammary gland, but also in cells of nonmammary epithelial origin [38 –45].
Within the quiescent EdU+5BrdU− population, several genes involved in cell division and differentiation were found downregulated compared to the EdU−5BrdU− population. Gja1, also known as connexin43, was downregulated over 12-fold in the EdU+5BrdU− population. Gja1 is required for normal gland development and function as reduced Gja1 expression results in delayed development and reduced milk secretion and ejection [46,47]. Cyclin D1 and Myc were also found downregulated in the EdU+5BrdU− population. ERα binds both the Cyclin D1 and Myc promotors resulting in induction of the genes [48]. As mammary stem cells are believed to be ERα-negative, this finding of decreased expression of Cyclin D1 and Myc in LRCs fits into the emerging profile of mammary stem cells. Ablation of Cyclin D1 results in a marked reduction of mammary acinar development and inhibition of lactation [49].
CD24+CD29high mammary progenitor cells are expanded in transgenic mice overexpressing intracellular activated Notch1 and this expansion is regulated by cyclin D1 [50] lending further credence that LRCs may be stem cells. These mice eventually develop mammary tumors with the CD24+CD29high acting as cancer stem cells. When cyclin D1 is knocked down in the CD24+CD29high cancer stem cells by shRNA treatment, they revert to bipotential mammary progenitor cells [51].
Overexpression of Notch receptors and ligands has been associated with various developmental disorders and cancer initiation and progression. Antibody targeting of Notch1 depleted breast cancer cell proliferation, induced apoptosis, and reduced expression of the Notch target genes HES1, HES5, and HEY-L [52]. Inhibition of Notch1 also attenuates mammosphere formation by breast cancer cells and reduces the population of CD44
Notch1 and Notch2 are differentially expressed in the EdU+5BrdU− population, but not the EdU+5BrdU+ cell fraction relative to the double-negative population. We did not find commonly differentially expressed genes of the 168 examined between the 2 LRC populations. This difference reflects a difference in the cell cycles of the 2 LRC populations. It was within the EdU+5BrdU+ population that the Notch-inducible genes Hes1 and Hey2 were elevated.
Although the arrays run were limited in the number of genes arrayed (168 total), it is plausible that the 2 LRC populations reflect different classes of mammary progenitor cells. We know that the different classes of mammary progenitor cells react differently to various stimuli. In the developing mouse mammary gland, high levels of Notch3 and Notch4 are found in the terminal end buds (TEBs) that drive ductal morphogenesis, while Notch1 or Notch2 expression was absent [54]. The TEBs contain a population of duct-limited progenitor cells that give rise to the myoepithelial cell layer of the ducts. Notch receptors are temporally regulated in mammary epithelial cells [22]. Activation of Notch1 induces luminal progenitor cell expansion [55]. Notch4-induced mammary tumorigenesis does not target the lobule-limited progenitor cells [56]. We have previously demonstrated that knocking down the expression of the EGF family member amphiregulin inhibited the mammosphere formation by ductal-limited progenitor cells, but had no effect on the mammosphere formation by lobule-limited progenitor cells [28]. The different classes of mammary progenitor cells react differently to the same signals.
The Notch signaling pathway regulates stem cell functions in numerous tissues. Notch plays a pivotal role in neural and brain development by controlling stem cell function and regulating symmetric and asymmetric divisions. In dividing radial glial cells of the zebrafishbrain, Notch regulates self-renewal of glial stem cells. After asymmetric divisions, the daughter cells are in an apical position, while the parent stem cell lies in a basal position and the daughter cell is the source of Notch. This intralineage Notch signaling regulates self-renewal in these cells [57]. A similar mechanism is found in chick neural tube development where Notch signaling dynamics are regulated by cell division orientation [58]. Notch signaling also regulates asymmetric divisions in the Drosophila brain [59]. This is similar to what we observe in the mammary gland where the progenitor cells reside in basal positions, while the surrounding cells have increased expression of Notch ligands, whereas the basally positioned progenitor cells demonstrate increased Notch activation and nuclear translocation. Interestingly, both neural tissues and mammary tissues are of ectodermal origin.
The Notch pathway is not only important for normal mammary development, but also plays key roles in breast cancer development. Notch1-specific monoclonal antibodies reduced mammosphere formation by triple-negative breast cancer cells and reduced the CD44+CD24− cancer stem cell population [60] and the CD24+CD29high normal progenitor cell population [52]. In a transgenic model where Notch1 expression is doxycycline regulated, Notch1 inhibition resulted in rapid tumor regression and a decrease in mammosphere formation, while Notch1 activation stimulates luminal progenitor cells [61].
In summary, we have presented data that mouse mammary LRCs differentially express a number of genes and that these genes change depending on the proliferative state of the LRC. Inhibition of one of these differentially expressed genes, Notch1, inhibits in vitro stem cell characteristics of colony-forming capacity and nuclear label retention.
Footnotes
Acknowledgments
The authors would like to acknowledge Dr. Meredith Morris at the Clemson University Genomics Institute and Dr. Julie Nelson at the University of Georgia Flow Cytometry Core Facility for their assistance with cell sorting, Ms. Tina Parker and the Clemson University Animal Services at Godley-Snell Research Facility for their help with in vivo studies, Miss Hiral Shah for assistance with the colony-forming assays, and Miss Aurora Booth and Miss Anja Booth for help with immunohistochemistry. Funding for the project was provided by the Institute for Biological Interfaces of Engineering of Clemson University.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.