
Abstract
Approximately 12.5% of all 9,920 extant bird species in the world are threatened with extinction, and yet conservation efforts through natural breeding of captive species continue to encounter difficulties. However, sperm cryopreservation and artificial insemination offer potential benefits over natural breeding, but their applicability is still limited in nondomestic species. In this study, we aimed to exploit the potential of germ cell xenotransplantation as an alternative tool for preserving germplasm of endangered birds. The study was designed to investigate whether transfer of either spermatogonia-enriched cell fraction (SEF) or crude testicular cell fraction (CTF) from adult Japanese quails (as a model for wild species) would result in recolonization of gamma-irradiated gonads of adult recipient chickens. One month after transplantation, 75% of recipients injected with SEF and 25% of recipients injected with CTF resumed spermatogenesis. However, it took more than 3 months for 33% of the negative controls to resume marginal production of sperm. Some SEF recipients produced more spermatozoa bearing head morphology compared with donor controls. DNA analysis using quail-specific primers did not detect donor's DNA in these recipients' semen. However, 6 months after xenotransplantation, presence of quail germ cells was demonstrated by polymerase chain reaction and by immunohistochemistry in 1 rooster injected with SEF. These findings indicate that spermatogonia from adult quails were capable of colonizing immunocompetent testis of adult chickens but failed to produce sufficient sperm. Despite this limitation, the present approach represents a potential conservation tool that may be used to rescue germ cells of endangered adult male birds.
Introduction
In 1994, Brinster and Avarbock [10] demonstrated the first transplantation of male germ line stem cells in mice; the infertile recipient males used successfully produced donor-derived sperm. Since then, allogeneic germ cell transplantation has been examined in rodents [5,11 –15], larger mammals [16 –21], fish [22 –24], and birds [25 –29], proving its applicability in the investigation of basic features of spermatogonial physiology and male fertility, maintenance of genetic stock of superior males indefinitely in surrogate recipients, and production of transgenic spermatozoa through genetic manipulation of isolated germline stem cells [30,31]. In addition, some studies involving interspecies germ cell transplantation in mammals have achieved complete donor-derived spermatogenesis [32 –35], while others have only observed spermatogonial proliferation in recipient species [36 –39]. However, according to Dobrinski [40], even in cases where spermatogonial cells from donor animals did not result in complete spermatogenesis, the method offers a bioassay procedure for isolation and propagation of germ cells from other species. Consequently, many researchers have examined the applications of xenotransplantation of either germ cells or testicular tissue in the conservation of endangered mammals because it can provide an option to recover germ cells from individuals that die prematurely or from adult males with impaired spermatogenesis [41 –44].
Although restoration of spermatogenesis and male fertility by allotransplantation of dispersed testicular cells in adult recipients has been already reported in chickens [29], there are no studies addressing the significance of this methodology in the conservation of wild birds. Recently, some authors demonstrated that interspecies blastodermal (BC) or primordial germ cell (PGC) transfer into developing chicken embryos resulted in progenies with the donor's phenotype [8,45 –47]. However, the use of BCs or PGCs presents several limitations for their use in wild birds. It requires acquisition of massive amounts of cells, the efficiency to produce germline chimeras after long-term culture is low, and in addition, it requires manipulation of avian embryos from endangered or rare species [48,49]. Conversely, transplantation of spermatogonia from adult birds might have a tremendous impact on future conservation efforts, especially if combined with other technologies such as testicular biopsy and cultivation of germline stem cells. Herein, we explore the transfer of testicular cells into gonads of sterilized recipient chickens that offer several advantages: (1) ease of handling animals during semen collection; (2) year-round reproduction instead of seasonal breeding; (3) possibility to rescue germ cells postmortem; and (4) greater exchange of genetic material among institutions, since importation of domestic species is less prone to legal issues regarding animal health than wild birds. The current study was designed to examine the in vivo testicular colonization and regenerative capacity of testicular germ cells obtained from Japanese quail, a different but closely related bird species.
Materials and Methods
Animals and experimental groups
A schematic diagram displayed in Fig. 1 depicts the general procedure for interspecies germ cell transfer, from quail into adult chicken testes. Fifteen adult (6–8 weeks) male Japanese quail (Coturnix japonica) were used as donors of testicular germ cells, and 15 adult (25–32 week-old) chickens (Gallus gallus domesticus) were used as sterilized recipients. An additional group of roosters (n=3) was retained as positive controls (nonsterilized males), whereas 3 other males were irradiated but not transplanted and kept as negative controls. Quails were maintained in collective cages (10′′×20′′×9′′), whereas chickens were maintained in individual cages (12′′×18′′×18′′) under standard husbandry conditions with food and water provided ad libitum.
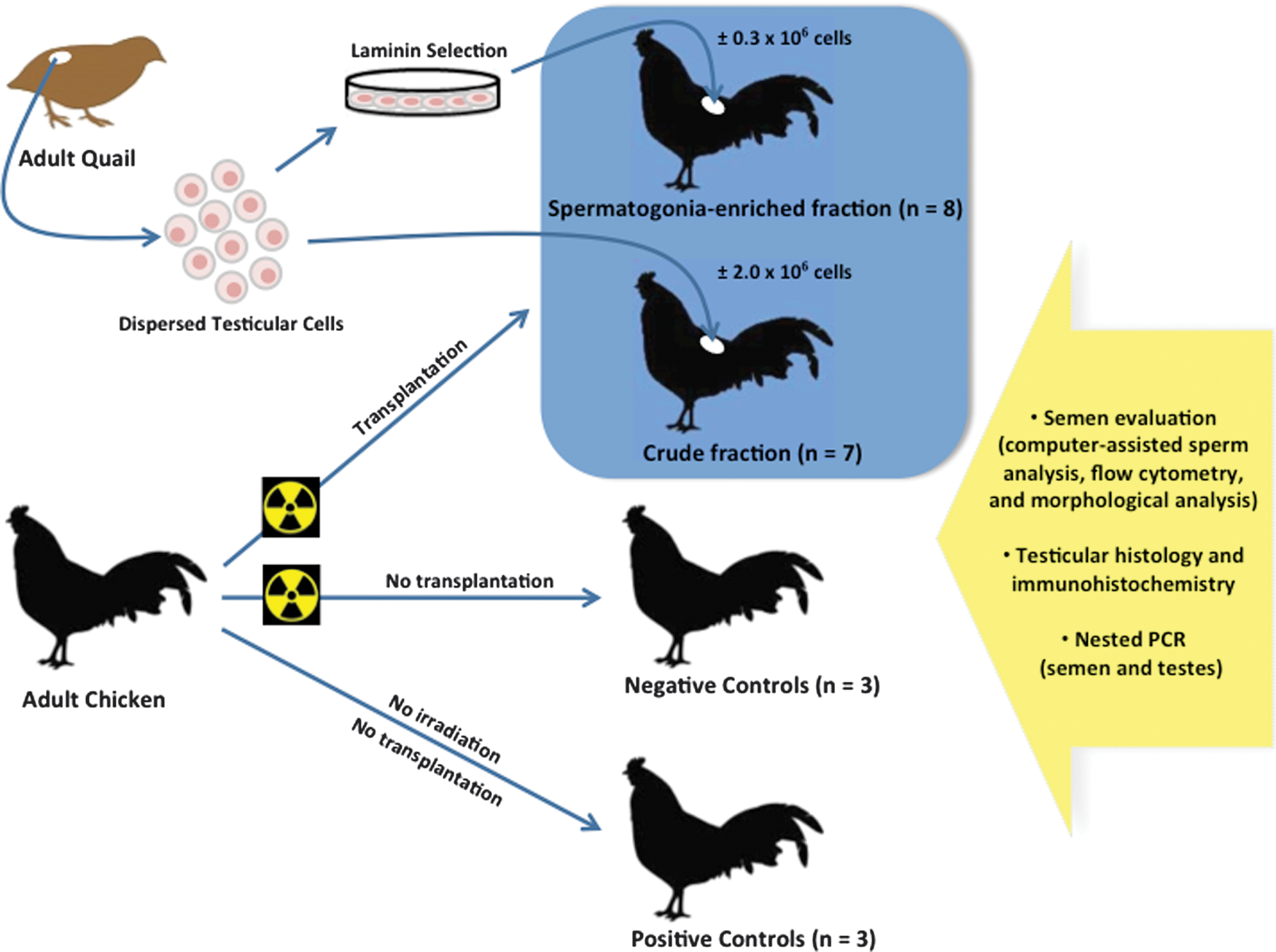
Flow diagram of the xenogeneic germ cell transplantation. Spermatogonia-enriched or crude testicular cell fractions from adult Japanese quails were transplanted into the testes of adult chicken previously sterilized by gamma irradiation. Nonirradiated and irradiated roosters were retained as positive and negative controls, respectively. Semen samples were submitted to morphological and computer-assisted sperm analysis, imaging flow cytometry, and DNA analysis by polymerase chain reaction (PCR) using quail-specific primers. Testes were collected about 6 months later for histology, immunohistochemistry, and DNA analysis. Color images available online at
The testicular cells isolated from quails were either transplanted immediately after collection [crude testicular cellular fraction (CTF); n=7] or subjected through a step of laminin selection for the enrichment of spermatogonial cells before transplantation [spermatogonia-enriched fraction (SEF); n=8]. Chicken semen samples were collected once a week for∼5 months (starting 3 weeks before radiation), and subsequently examined by computer-assisted sperm analysis (CASA) system, imaging flow cytometry, morphological analysis, and DNA analysis by polymerase chain reaction (PCR) with quail-specific primers. Roosters were euthanized 3 weeks after the last semen collection with the aim of collecting testes for histology, immunohistochemistry, and DNA analysis. All animal procedures were performed with approval and under the guidance of the Institutional Animal Care and Use Committee at the Michigan State University (AUF No. 03/09-039-00).
Irradiation treatment
A computed tomography (CT) scan (Brightspeed™; GE Healthcare®) of an anesthetized standard size adult rooster was performed to localize abdominopelvic organs, to accurately direct irradiation rays to the testes and plan irradiation sequences (Supplementary Fig. S1; Supplementary Data are available online at
Preparation of dispersed testicular cells
Immediately after euthanasia, quail testes were removed and washed in phosphate-buffered saline (PBS). Dispersal of testicular cells was performed as described by Guan et al. [49]. Briefly, the cells were prepared from∼2–3 g of testicular tissue finely minced into 1–2 mm pieces using sterile scissors. The minced tissue was incubated (37°C, 5% CO2) with 1 mg/mL collagenase IV (Sigma) in 4 mL of Waymouth medium (Invitrogen) for 20 min. Tubule fragments were dispersed by pipetting up and down several times, and the resulting suspension was centrifuged (200 g, 5 min) to get a pellet. The tissue pellet was reconstituted and washed with PBS, and then incubated in 4 mL of trypsin/ethylenediaminetetraacetic acid (EDTA) solution for 5 min (37°C, 5% CO2). After inactivation of trypsin, the cell suspension was filtered through a 70 μm nylon mesh (BD Biosciences), centrifuged (450 g, 5 min), and the pellet resuspended in 2 mL of M-2 mouse embryo culture medium (Millipore). Cell viability was then assessed by trypan blue exclusion.
Enrichment of donor spermatogonia
Enrichment of spermatogonia was done by laminin selection as described by Guan et al. [50]. Briefly, cell culture dishes were coated with 3 mL of laminin solution (20 μg/mL, overnight, 37°C, 5% CO2). Next day, laminin-coated dishes were washed with PBS and incubated for 1 h with bovine serum albumin (BSA) 0.5 mg/mL. Subsequently, dishes were washed with PBS and 2 mL of testicular cell suspension (10×106 cells) was added and incubated for 15 min (37°C, 5% CO2). Unattached cells were removed by 3 washings with PBS and discarded. Attached cells were removed (with trypsin/EDTA), washed thrice and resuspended in 2 mL of fresh M-2 mouse embryo culture medium. Next, viability and concentration of the enriched germ cell preparation were determined. To estimate the degree of spermatogonia enrichment provided by laminin plating, smears were made from pre- and postselection cell samples and stained with hematoxylin–eosin. Different cell types were identified on the basis of detailed morphological descriptions made by Lin and Jones [51] and Jones and Lin [52], and divided into the following groups: erythrocytes; spermatozoa; spermatids; spermatocytes; spermatogonia (dark type A and pale type A spermatogonia); other cells (eg, cells in division or with dubious morphology); and clumps of cells (eg, cells attached to Sertoli cells or other cell aggregates).
Transplantation of donor germline cells
Transfer of cells was performed 1 month after irradiation as described by Trefil et al. [29]. Roosters were anesthetized (15 mg/kg ketamine; 4 mg/kg xylazine, i.m.), and transfer of quail testicular cells into chicken gonads was performed via laparoscopy. The group injected with the CTF (no laminin selection) received 100 μL of cell suspension containing a total of∼2×106 cells per testis, while the group injected with the SEF received 100 μL containing a total of∼0.3×106 cells per testis (Fig. 1). Injections were made into each testis through the tunica albuginea at 2 or 3 different, randomly selected locations. Analgesics were given after surgery (0.3 mg/kg meloxican, oral).
Semen collection and evaluation
Semen was collected once a week for almost 5 months by the dorso-abdominal massage method [53], and care was taken to reproduce procedures (ie, same time of collection and staff members) so as to avoid introduction of additional variables to the semen analysis. Clean ejaculates (without fecal, uric, or blood contamination) were individually assessed within 30–40 min of collection, after dilution (1:100) in M-2 medium at room temperature. Sperm concentration and motility were examined using a computer-assisted sperm analyzer (CASA-system: HTM-IVOS; Hamilton-Thorne Biosciences) with software settings adjusted to obtain clear identification of both chicken and quail spermatozoa (Table 1). Diluted semen samples were loaded at room temperature into a 20-μm 2X-CEL chamber (Hamilton-Thorne Biosciences).
VSL, straight line velocity.
To assess morphological differences (mainly in head and midpiece length) between chicken and quail spermatozoa, sperm samples were co-stained using mitochondrial and nuclear fluorescent dyes. Briefly, 1 mL of diluted semen (1:100) containing live cells was incubated with 0.25 μL of MitoTracker Red CMXRos (Invitrogen; 250 nM final concentration) at 37°C for 30 min. Subsequently, the mixture was centrifuged (500 g, 1 min) and the supernatant was discarded. After fixation in 10% formalin (30 min at room temperature), 10 μL of each sample was placed on a microscope slide, air dried, and mounted in Prolong Gold antifade reagent with 4′6-diamidino-2-phenylindole (DAPI) (Invitrogen). One month after transplantations, spermatozoa from individuals that resumed spermatogenesis were categorized as chicken spermatozoon (shorter head and midpiece), quail spermatozoon (longer head and midpiece), abnormal spermatozoon (shorter head with unusual red staining of the midpiece/tail), and long head spermatozoon (longer head with shorter midpiece). In every slide, 500 cells were counted and classified within these categories. At least 5 different ejaculates from quail controls, positive and negative chicken controls, and transplanted chickens producing sperm were analyzed for morphology.
Imaging flow cytometry
Semen samples from transplanted chicken recipients were also analyzed by a multispectral quantitative imaging flow cytometer (ImageStreamX Imaging Flow Cytometer; Amnis Corporation), which enabled us to objectively evaluate a larger number of spermatozoa (ranging from 2,000 to 18,000) per sample. After collection, semen was diluted in M-2 medium with 1% polyvinylpyrrolidone (PVP), and spermatozoa were stained using MitoTracker Red CMXRos (37°C, 30 min). Next, samples were washed twice with M-2 medium with 1% PVP, and fixed in 2% paraformaldehyde with 7% PVP. Following fixation, sperm was washed twice in PBS with 7% PVP and co-stained with DAPI. For analysis, samples were diluted using PBS with 7% PVP to a final concentration of 20×106 spermatozoa per mL. Spectral compensation was digitally calculated by analysis software (IDEAS; Amnis Corporation) on the basis of unlabeled cells and cells labeled with a single-color positive control for each fluorochrome employed. Chicken and quail sperm controls plus mixed samples containing both cell types (50%/50%) were used to develop an image based classifier for each type of spermatozoon. Single cells were separated from debris and multicellular aggregates in the brightfield channel using features aspect ratio and area, and only cells positive for both fluorochromes were used for all subsequent analysis. Because quail spermatozoa have a very large midpiece, this feature was used to distinguish them from chicken spermatozoa (which have a smaller midpiece and much brighter MitoTracker staining).
Tissue collection and histological analysis
At necropsy, recipient testes were rapidly collected and split; one half was prepared for light microscopy and one half was snap frozen and prepared for immunohistochemistry. The halves used for light microscopy were fixed in 10% formalin (4°C, 48 h), embedded in paraffin, and cut into 4–5 μm sections for hematoxylin–eosin staining. The unfixed halves were immediately placed in dry ice for 10 min, and cut into 3−7 fragments (depending on the testis size) of ∼10×5 mm each. Next, fragments were immersed in Tissue-Tek compound (Sakura Finetechnical) and immediately frozen using liquid nitrogen. Tissue molds were stored at −80°C until processing. Frozen sections of 4–5 μm thickness were cut by a cryostat (Cryotome FSE; Thermo Fisher) and attached to poly-
For immunostaining, slides were removed from −80°C freezer (left inside the box at room temperature for 15 min), and subsequently fixed with precooled methanol for 5 min at room temperature. Sections were then rinsed thrice in PBS, incubated twice with 0.1% BSA in PBS (2 min, each), permeabilized twice with 0.5% Triton X-100 in PBS (5 min each), and treated with normal goat serum (1:100 dilution, 30 min, room temperature) to block nonspecific background staining. Incubation with quail-specific antibody (QCPN concentrate; Developmental Studies Hybridoma Bank, University of Iowa) at a dilution of 1:25 with 0.5% Triton X-100 in PBS was carried out overnight at 4°C in a humidity chamber. After 3 washes with 0.5% Triton X-100 in PBS (5 min each), the sections were incubated with secondary antibody (Alexa Fluor 594 goat anti-mouse; Invitrogen) at 1:500 dilution for 90 min. Lastly, sections were washed for 5 min with 0.5% Triton X-100, counterstained with hematoxylin, and mounted with ProLong Gold antifade reagent (Invitrogen).
DNA analysis
To ascertain the presence of quail cells in recipient semen and testes, genomic DNA was extracted from freshly collected ejaculates and from testicular frozen fragments (from halves prepared for immunohistochemistry) using a High Pure PCR Template Preparation Kit (Roche). Genomic DNA from semen and testis of adult quails and nontransplanted chickens served as positive and negative controls, respectively. Purity and concentration of DNA recovered were determined with a NanoDrop® spectrophotometer (Thermo Scientific). The detection of quail cells was performed using qualitative nested PCR with the following primers for Japanese quail ovalbumin cDNA (GenBank No. X53964): 5′-outer primer, 5′-CAGAGGCTGGAGTGGATGCTA-3′; 3′-outer primer, 5′-TATTACTCTGTGTAAGGGAAGGGTGAAGT-3′; 5′-inner primer, 5′-GGGCTGACCATCCATTCCTC-3′; 3′ inner primer, 5′-GTCTGGATGCAGCAGAGAACATGC-3′ (designed by Soh et al. [54] and our laboratory, respectively) [54]. Ten microliter of the DNA solution (100 ng) was added to a reaction mixture (10 μL) consisting of 10×PCR buffer, 0.8 mM dNTP mix, 4.5 mM MgCl2, 1.5 μM each of the outer primers, and 2.5 U Taq polymerase (all reagents are from Invitrogen). The first amplification step was as follows: initial denaturation 95°C for 10 min; 15 cycles of 95°C for 30 s, 60°C for 30 s, 72°C for 30 s; and final extension at 72°C for 10 min. Then, 2 μL of the amplified products were used for a second amplification of 22 cycles (equal amounts of reagents and cycling conditions as before), using 1.5 μM each of the inner primers. PCR products (predicted size of 181 bp) were separated by electrophoresis on 1.5% agarose gels (150 V for∼90 min) and analyzed using a GelDoc 1000 UV Fluorescent Gel Documentation System-PC (Bio-Rad Laboratories). To confirm that the PCR products were in fact from quail, amplified products from quail controls and positive chicken recipients were sequenced using the primers described above. PCR products were extracted and purified using Wizard SV Gel and PCR Clean-Up kits (Promega). Sequencing was performed in an ABI PRISM 3730 Genetic Analyzer (Applied Biosystems). The assembled sequence was submitted to BLAST (
Statistical analysis
All data are presented as mean±standard error. Differences between means having of P<0.05 (*) were taken as significant for all statistical tests, which were performed using GraphPad Prism 5 for Windows (GraphPad Software, Inc.). Laminin selection data were analyzed using a Student's t-test, whereas values from the sperm morphology were subjected to one-way analysis of variance followed by Tukey test.
Results
Laminin plating effectively enriched the proportion of quail spermatogonia
After laminin selection, 3.1% of cells from enzyme digestion were recovered in the adherent fraction (an average of 0.31±0.06×106 cells per plate). Morphological analysis of the CTF revealed that only 8.9% of cells were spermatogonia, while the adherent fraction from laminin-coated plates contained 29.0% spermatogonia (Fig. 2). This represented 3.2-fold enrichment in the proportion of quail spermatogonia. Further, most cells (75.9%) in the CTF were categorized as spermatozoa, spermatocytes, and other cell types, whereas following differential plating most cells (81.4%) consisted of erythrocytes and spermatogonia. Cell viability averaged 96.70%±0.30% (n=4) for CTF and 96.62%±1.77% (n=4) for SEF.

Preparation of quail spermatogonia-enriched cell fraction by laminin plating.
Transplantation of SEF improved restoration of spermatogenesis
Our results showed that chicken testes were susceptible to repeated gamma-radiation, since sperm concentrations in all irradiated animals dropped from 4.52±2.05 billion spermatozoa/mL to 0 within 5 weeks after the first 8-Gy fraction (Fig. 3). Additionally, CT scans of chicken before and after irradiation demonstrated a significant decrease in testes size indicating that gonads were efficiently targeted by radiation (Supplementary Fig. S2). No adverse effects or behavioral changes were detected in these irradiated animals. Two out of three negative controls remained azoospermic until euthanasia (24 weeks after first radiation treatment), whereas 1 individual showed marginal recovery of sperm production (5.32±1.34×106 spermatozoa/mL) after 13 weeks (Fig. 3). In contrast, injection of SEFs restored spermatogenesis in 6 out of 8 recipients (75%) within 5 weeks, with 2 males yielding about half of their normal sperm concentration after transplantation (1494.46±169.07×106 spermatozoa/mL) (Fig. 3). At the same time, only 2 out of 7 recipients from the group transferred with the CTF ejaculated spermatozoa (starting 5 weeks after transplantation), but sperm concentrations ranged from 12.29 to 401.03×106 spermatozoa/mL (Fig. 3).
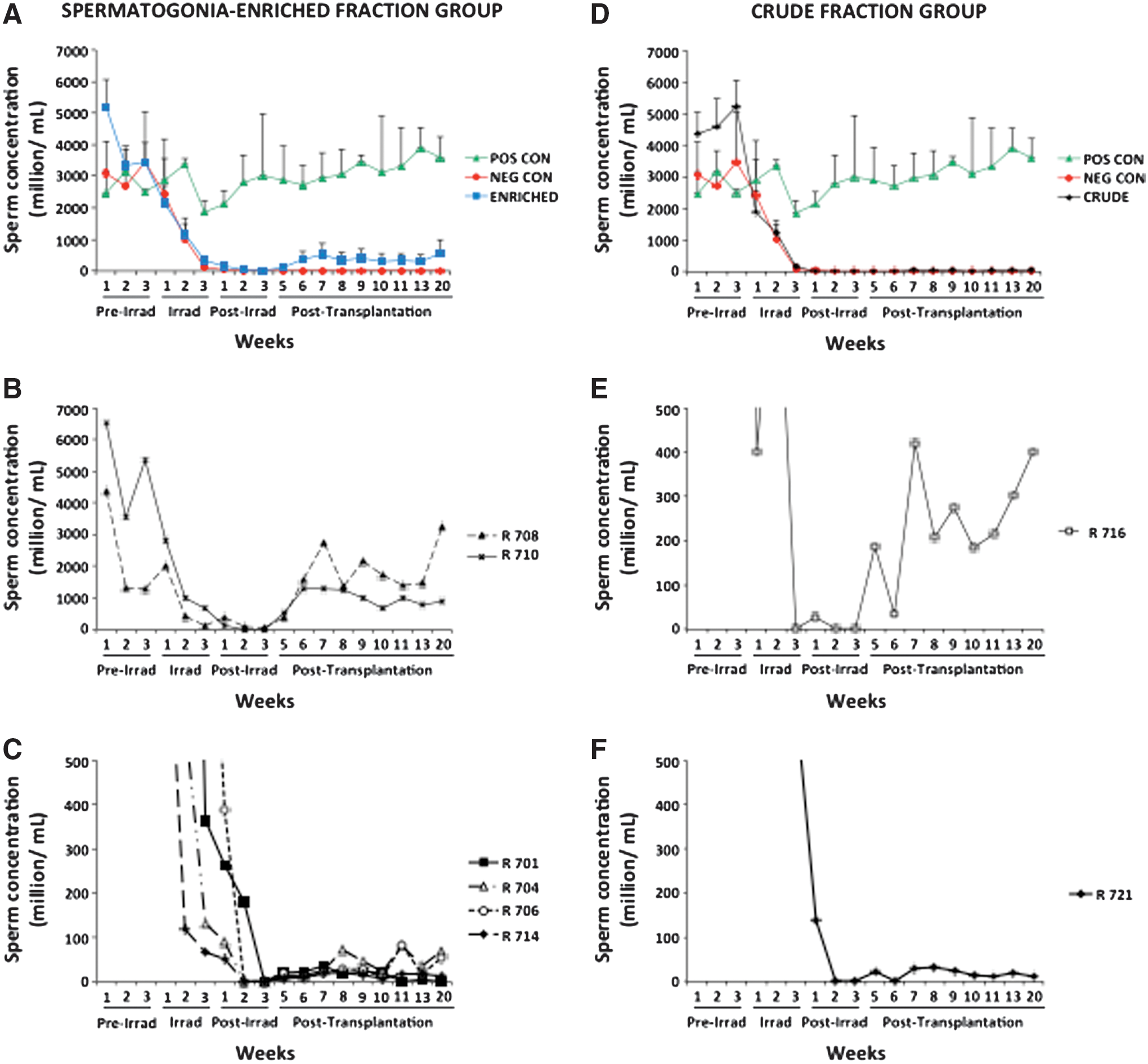
Time-course of sperm concentration in the various experimental groups. Weekly mean (±SEM) concentration (million/mL) of sperm cells in the spermatogonia-enriched fraction
Transplanted quail germ cells colonize chicken testis but were unable to generate sufficient donor spermatozoa
Five weeks after transplantation, morphological analysis of sperm from transplanted chickens revealed cells that resemble quail spermatozoa to some extent (Fig. 4). Several spermatozoa (ranging from 4.8% to 15.3%) showed an abnormal Mitotracker Red staining of midpiece/tail, whereas few cells (ranging from 0.1% to 0.9%) exhibited lengths of heads comparable to those observed in quail spermatozoa. However, despite being present in smaller percentages than most cells in transplanted recipients, both morphologies were also found in the positive controls (Fig. 4). Later on, assessment of 3 ejaculates from a negative control revealed the presence of sperm with long head (0.6%) and other abnormalities (13.6%), again these percentages were equivalent to those found in the transplanted chickens.
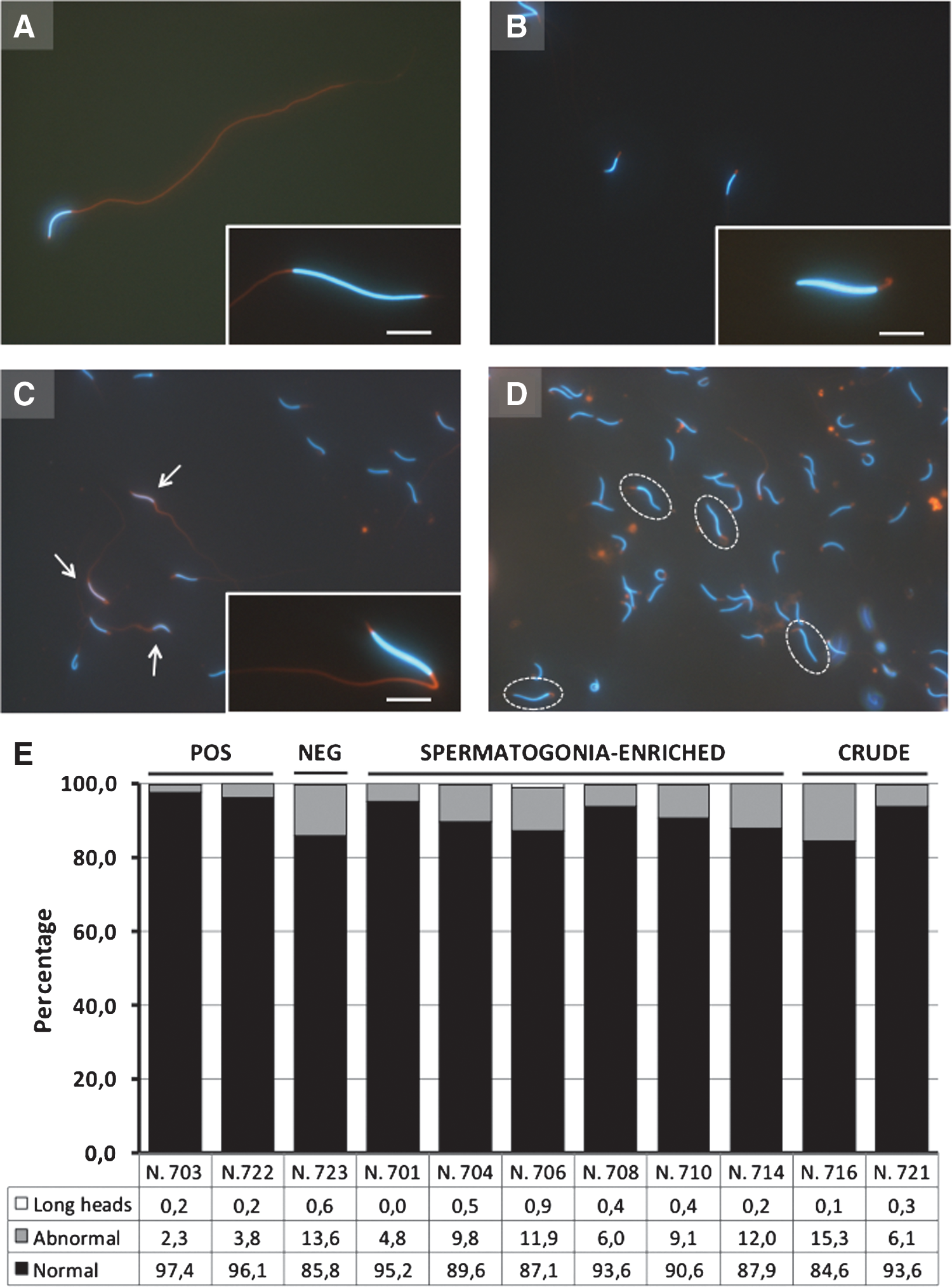
Morphological analysis of spermatozoa from transplanted and control chickens and quails.
Next, imaging flow cytometry was performed to objectively examine larger quantities of spermatozoa per ejaculate (Fig. 5). Based on the pattern of Mitotracker red staining, we were able to accurately detect cells in manually prepared samples containing a 50% mixture of quail/chicken spermatozoa (Fig. 5F). Adjustments of the R4 and R5 regions in the image graphs (using quail and chicken controls) allowed less than 15% false positive for both species, but also created a small area of unclassified events between R5 and R4 regions (Fig. 5E). Only 2 transplanted animals (males R704 and R710) exhibited percentages of quail cells greater than that observed in the chicken control. Subsequently, images from all events considered positive for quail morphology were visually reviewed so as to confirm that cells were correctly classified by the flow cytometer. Such evaluation demonstrated that most spermatozoa (95%) were misclassified as quail sperm due to abnormal red staining of the midpiece/tail in chicken cells, while only a small number of images were correctly classified as quail sperm. Nevertheless, quail genomic DNA was not detected in sperm from any of the recipients transplanted with quail germ cells (total of 4 ejaculates examined per animal).
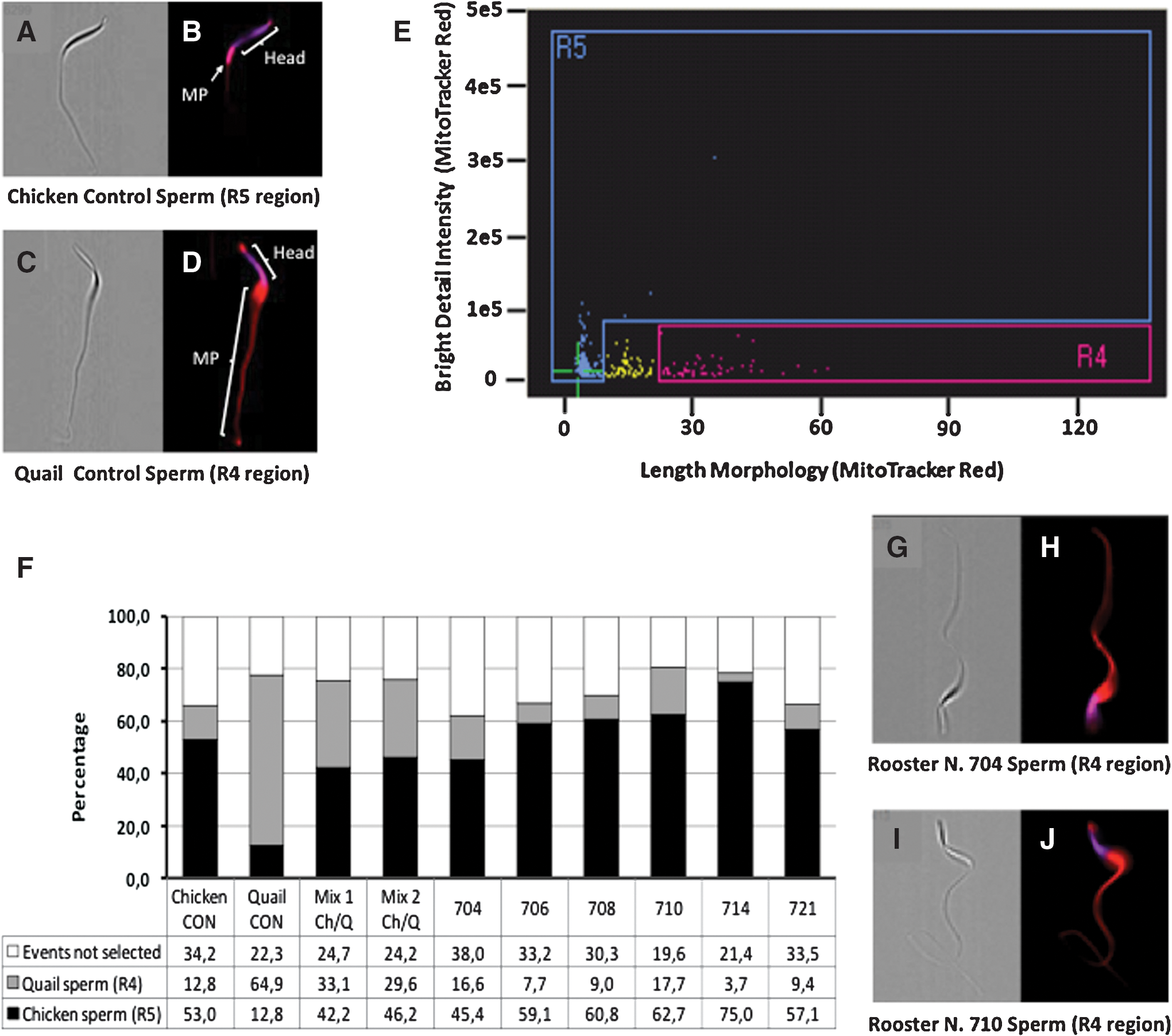
Identification of quail and chicken spermatozoa by imaging flow cytometry. ImageStreamX images of chicken
At necropsy, we observed that testes size varied among groups, with positive controls presenting the highest dimensions, whereas negative controls and transplanted chickens without sperm production had the smallest sizes (Fig. 6). Males exhibiting post-transplantation sperm output showed intermediate testes sizes, although 2 males injected with the SEF (R708 and R710) presented testicular dimensions close to those of positive controls. Unlike positive controls, which showed full spermatogenesis, seminiferous epithelium from 2 negative controls comprised only of Sertoli cells (ie, complete absence of germ cells). In 1 negative control recovery of spermatogenesis was only noticed in a few seminiferous tubules of the left testis (less than 1% of the parenchymal area). No signs of cellular damage were detected in the interstitial tissue of all irradiated males (Fig. 6). During analysis of the transplanted recipients producing semen, we observed variable ratios of seminiferous tubules showing spermatogenesis, in which individuals yielding higher sperm concentrations had greater ratios of functional seminiferous epithelium.
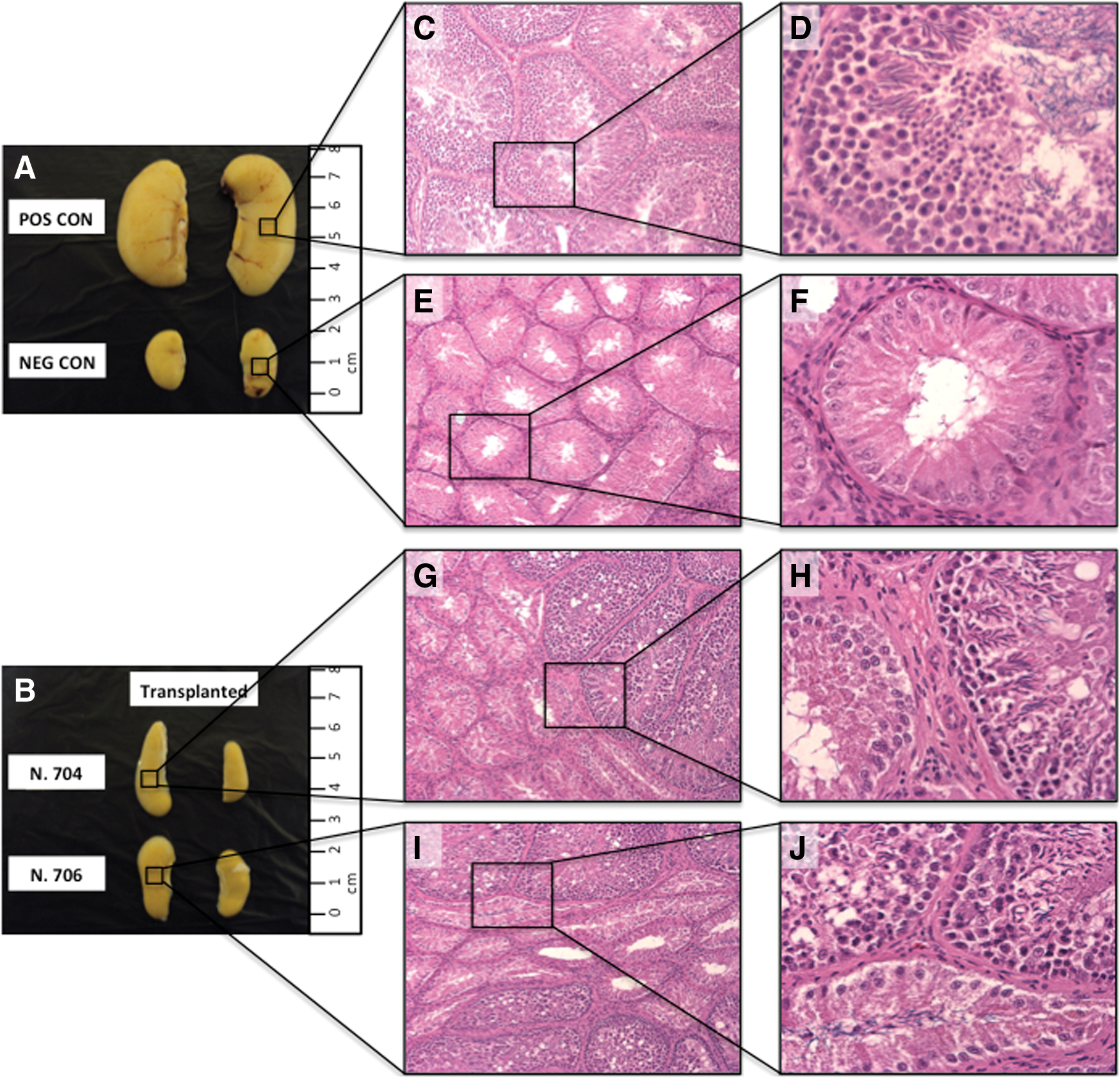
Macroscopic and histological comparison of chicken testes from nonirradiated controls and irradiated controls and transplanted groups. Testes of nonirradiated (positive) controls were larger in size
QCPN successfully detected testis cells in quail controls, by labeling nuclei of both germ cells in the seminiferous tubules and interstitial cells (Fig. 7). Notice that this antibody cross-reacted with interstitial cells in chicken controls, but instead of nuclear localization, it presented a cytoplasmic staining pattern. Since, nuclear localization appears to be the specific signal, we used this antibody to demonstrate the presence of quail cells in the seminiferous tubules of all recipient chickens. Six months after transplantation, donor germ cells were found in a small number of seminiferous tubules of 1 recipient of the SEF (Fig. 7). Quail germ cells proliferated to form chains of connected cells that in most positive tubules were located near the basement membrane. None of these seminiferous tubules showed the typical signs of spermatogenic differentiation toward the lumen, indicating an arrest during the horizontal expansion.
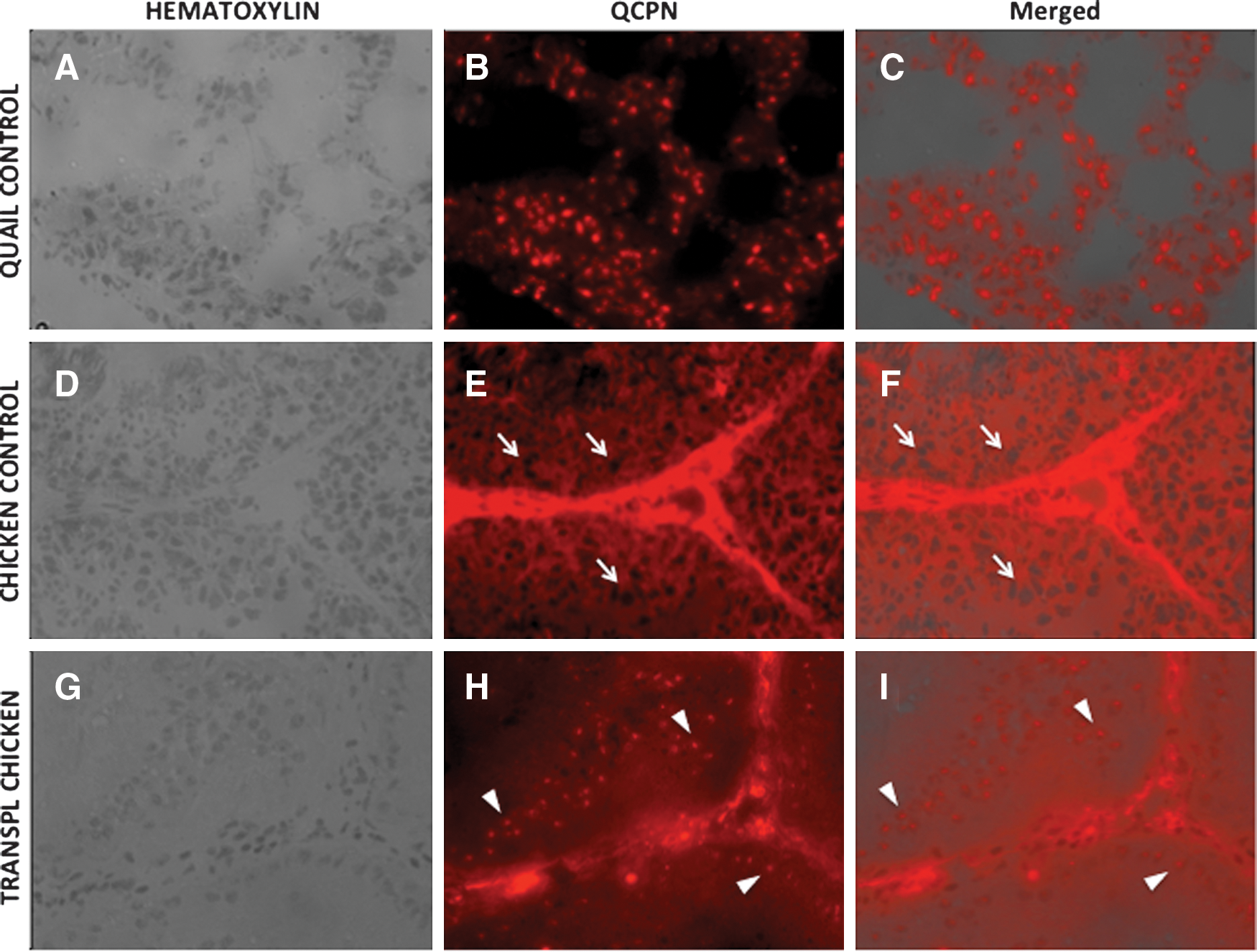
Immunodetection of quail germ cells using quail-specific antibody (QCPN, a nuclear marker) in testes of transplanted chickens. Seminiferous tubules of adult quail [positive control
DNA profiling of the testicular tissue using quail-specific primers revealed the presence of quail DNA in 1 recipient transplanted with the SEF (Fig. 8). Subsequent sequencing of PCR products from this individual demonstrated 100% identity with the published Japanese quail ovalbumin cDNA sequence.
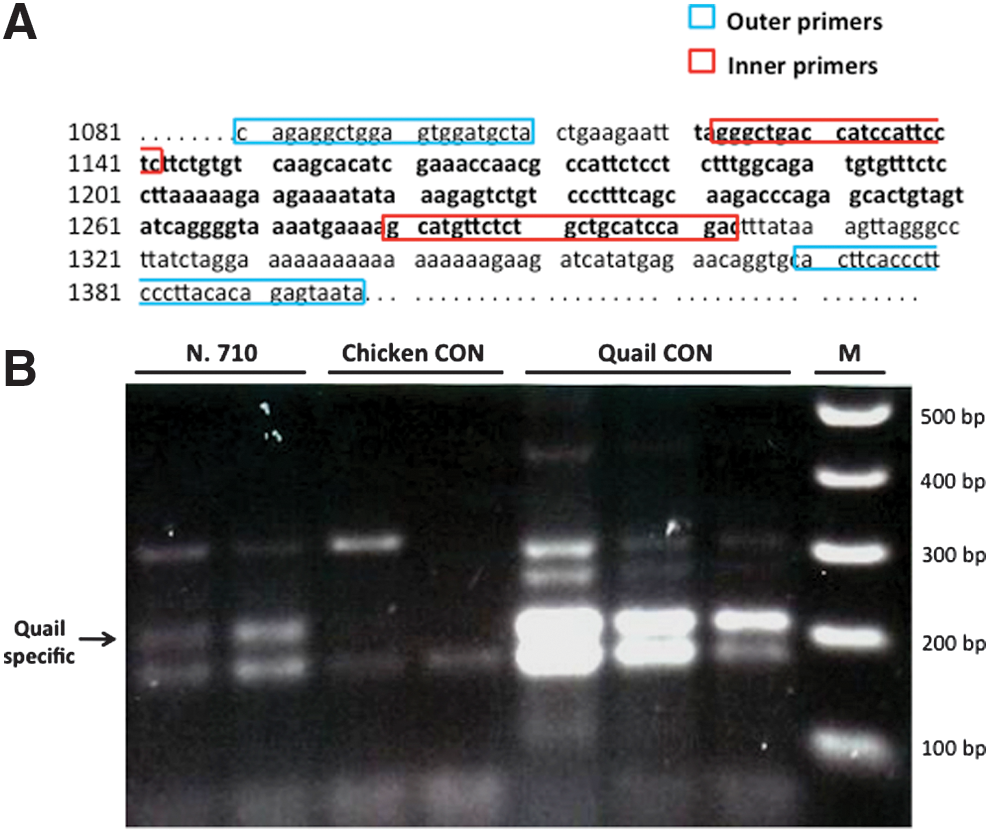
Detection of Japanese quail ovalbumin DNA by nested PCR in the testes of a transplanted chicken.
Discussion
Presently, conservation of wild bird germplasm faces technical challenges mostly emanating from interspecies variability in semen cryopreservation and technical difficulties in manipulating large-yolk eggs [8,28]. Hence, there has been considerable interest over the past decade in the propagation of endangered birds species through interspecies germline chimeras produced by transfer of BCs or PGCs into developing embryos [8,45 –47,54 –56]. Although these studies have confirmed the feasibility of the technique for preserving avian species, there is still a limiting factor associated with germ cell transplantation using embryos as donors, as the procedures entail manipulation of fertile eggs, which in many instances are not available or are too valuable to undergo surgery. Besides, application of PGCs offers additional disadvantages associated with the high number of embryos necessary to collect enough number of cells and the maintenance of long-term cultures [48,49,57,58]. Thus, a practical alternative that can overcome the limitations of embryo-mediated systems is to perform transplantations using adult testicular germ cells.
In this study, germ cells harvested from sexually mature quail testes successfully colonized the seminiferous tubules of adult immunocompetent recipient chicken and survived for at least 6 months after transplantation. However, despite their proliferation in the chicken testicular environment quail progenitor germ cells were unable to fully restore quail spermatogenesis. Similar findings have been previously described in interspecies germ cell transplantations involving transfer from nonrodent mammals into mouse seminiferous tubules [36,37,39,59]. According to Dobrisnki [30], this arrest in spermatogenesis is possibly a consequence of the increasing phylogenetic distance between donor and recipient species, leading to an impaired interaction between testis niche and spermatogonial stem cells (SSCs). Nevertheless, based on former achievements of BC and PGC transplantations using the quail–chicken model, we consider such incompatibility unlikely because testicular environment has proven to be capable of supporting spermatogenic differentiation and meiosis [54,55]. Moreover, although rejection of xenogeneic tissue cannot be completely discarded, recent reports on the immunoprotective properties of the testis suggest that the rejection phenomenon might not be responsible for the failure of quail cells to achieve spermatogonia differentiation in chicken testis [16,17,60 –64].
Alternatively, the nonrestoration of quail spermatogenesis could be connected to recipient age since none of the individuals used was immature nor hatchling. Such assumption relies on previous observations that seminiferous tubule environment from mice pups provided more colonization foci and allowed greater colony expansion than the adult tubule [65]. For these authors, this difference is probably linked to an increased accessibility of the donor cells to the testicular stem cell niche soon after birth, given that Sertoli cells are still proliferating and do not form tight junctions. Recipient age led to a remarkable 39-fold increase in SSC number in the testis from newborn to adult mice, and allowed pup recipients to generate offspring faster and more efficiently than adult recipients [65]. Thus, it is possible that tubular compartmentalization into basal and apical regions by Sertoli cell tight junctions hampered migratory access of quail cells to the seminiferous tubule basement membrane. This reduced accessibility may have affected colonization by SSCs and subsequently may have had a detrimental effect on the differentiation of quail germ cells. A more hospitable microenvironment of the embryonic gonad could explain why xenogeneic injections of PGCs into developing avian embryos successfully resumed gametogenesis from different species [8,45 –47,54 –56]. Hence, we intend to conduct further studies to demonstrate the impact of recipient age in the transplantation of adult testicular germ cells in the quail–chicken model.
Depletion of endogenous recipient spermatogenesis prior to the time of transplantation is another aspect that probably improves the efficiency of colonization of seminiferous tubules by donor germ cells [42,66 –68]. In this regard, our data show that fractionated testicular irradiation effectively halted spermatogenesis in adult chickens, with negative controls showing complete absence of spermatozoa for∼13–24 weeks (depending on the individual) following the last radiation exposure. Further, as previously reported by Trefil et al. [29,69], we found that irradiation did not destroy Leydig and Sertoli cells (Fig. 6), providing a supportive environment for donor spermatogenesis without systemic effects. These findings suggest that irradiation can be employed as an effective method to deplete endogenous germ cells from testes of other avian species that could be possibly used in future as hosts for transplantation. However, it is important to emphasize that irradiation protocols for preparing bird recipients present certain drawbacks pertaining to the need for specialized radiotherapy equipment, what makes this system not feasible to be used in the field. We are currently exploring other more practical and cost efficient approaches to efficiently deplete germ cells in the recipients.
Enrichment of germline stem cell populations has also been implicated in the enhancement of donor cells to colonize recipient testes, and among the several procedures currently available, we chose laminin plating due to its efficiency and convenience for enriching large quantities of cells with minimum damage [50,67,70,71]. Such selection is made possible by the presence of α6- and β1-integrins, which are specific surface markers on SSCs that act as laminin receptors [50]. In our study, isolation of testicular cells by adhesion to laminin yielded a 3.2-fold enrichment in the number of quail spermatogonia. Laminin-coated plates have been previously used for enriching SSCs in mice, rats, pigs, and bulls, but results ranged from 2- to 8.5-fold enrichment depending on the species [70,72 –74]. For some authors, these variations represent species-specific affinities to laminin, and probably reflect the differential expression of integrins [72]. Nonetheless, to our knowledge, the only attempt to enrich avian testicular cells was performed in domestic chickens by sorting of c-kit-positive cells, but neither the degree of SSCs enrichment nor the degree of colonization in recipient testes were reported [75].
Surprisingly, although chicken recipients did not produce sufficient quail sperm, injections with the SEF seemed to favor the reestablishment of endogenous spermatogenesis. This is based on the observation that most recipients transplanted with the SEF resumed spermatogenesis, We cannot rule out that gamma irradiation resulted only in a partial elimination of germ cell population in these recipients, especially considering the technical difficulties of targeting abdominal testes as also noted by other researchers [69,75]. However, this possibility seems remote for the following reasons: (1) CT was used to accurately establish the targeting area during sterilization; (2) recipients were randomly assigned to transplantation and control groups after the last irradiation exposure to avoid allocation bias; and (3) none of the negative controls exhibited similar recovery of spermatogenesis in terms of time span or sperm output.
Another explanation for the difference in the restoration of endogenous spermatogenesis may be related to the potential therapeutic effects of transferred SSCs on the postirradiated testes. For example, several lines of evidence suggest that mesenchymal stem cells (MSCs) alleviate irradiation injuries by ameliorating the microenvironment in the bone marrow, thereby accelerating hematopoietic recovery in vivo [76,77]. The mechanisms by which MSCs promote the recovery of hematopoiesis include secretion of growth factors (eg, interleukins-6 and -11, leukemia inhibitor factor, stem cell factor, Flt3 ligand, and stromal derived factor), protection against apoptosis, and reestablishment of the stem cell niche [76 –78]. Further, a recent study assessing the testicular system has confirmed that the granulocyte colony-stimulating factor can protect germ cells from gamma ray-induced apoptosis [71]. Thus, we hypothesize that larger numbers of SSCs injected in the enriched group enhanced spermatogenesis recovery by secreting higher amounts of bioactive molecules (eg, growth factors), which in turn contributed to the survival, self-renewal, and differentiation of the endogenous germ cells. Besides, it is also possible that factors secreted by spermatogonia have affected Sertoli cell activity, thus creating a more favorable microenvironment. Previous studies have shown the regulatory influence of germ cells on Sertoli cell function, and those findings indicate that this control is germ cell stage dependent [79 –81]. In addition, Fujino et al. [82] demonstrated that spermatogonial cell-derived factors can induce expression of lipocalin-2 gene in Sertoli cells, sustaining the idea that spermatogonia are also involved in the regulation of Sertoli cells activity. However, more detailed analysis will be necessary to confirm the therapeutic effects of SSCs following testicular irradiation injuries, and to elucidate the possible mechanisms through which these cells counteract radiation-induced damage.
Herein, in our current study, neither the assessment of sperm morphology nor imaging flow cytometry provided a reliable discrimination between quail and chicken spermatozoa, despite their very distinct structural features. For instance, quail spermatozoon has a longer head and an exceptionally elongated midpiece; lengths of the head and midpiece are respectively twice and 40 times longer than in chicken [83]. One reason for the misclassification of cells may be associated with the existence of abnormalities in the head and midpiece of chicken spermatozoa, making them resemble quail spermatozoa. For example, macrocephalic chicken sperm showed head lengths similar to quail sperm, whereas the abnormal red staining of midpiece/tail observed in some chicken cells may be confused with long midpiece of the quail spermatozoa. This might explain why irradiated chickens both negative controls and transplanted males yielded sperm with higher percentages of long heads and unusual red staining of midpiece/tail, since radiation has been proven to induce morphological abnormalities in sperm cells [84 –87]. Likewise, results from flow cytometry seem to be influenced by artifacts such as, nonspecific Mitotracker staining in the tails of some chicken sperm (making them look like quail sperm), or quail cells with broken midpieces (making them look like chicken sperm, shorter midpiece).
Our results, overall, indicate that xenogeneic transplantation using germ cells from adult donor birds is feasible and could be used as a technological approach for conservation of progenitor germ cells from endangered wild birds. However, continued investigation of this approach is required to overcome problems pertaining to spermatogenic differentiation and meiosis of donor germ cells in the recipient testicular environment.
Footnotes
Acknowledgments
These studies were supported by the College of Veterinary Medicine at Michigan State University (endowed research grant), the Department of Physiology at Michigan State University, and the Andalusian Laboratory of Cellular Reprogramming (LARCEL; Seville, Spain). The authors would like to acknowledge Dr. Brian E. Hall (Amnis Corporation) for his technical assistance with the imaging flow cytometry, Dr. James Sikarskie (College of Veterinary Medicine, Michigan State University) for lending his laparoscopic equipment, and Dr. Tamao Ono (Laboratory of Animal Developmental Genetics, Shinshu University) for kindly providing us with the QCR1 and QB2 antibodies. Many thanks go to Dr. Loro Lo-LajaKujjo for editorial review of the article.
Author Disclosure Statement
The authors declare that there is no conflict of interest that would prejudice the impartiality of this scientific work.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.