
Abstract
Skin-derived precursors (SKPs) are an attractive stem cell model for cell-based therapies. SKPs can be readily generated from embryonic and adult mice and adult humans, exhibit a high degree of multipotency, and have the potential to serve as a patient autologous stem cell. The advancement of these cells toward therapeutic use depends on the ability to control precisely the self-renewal and differentiation of SKPs. Here we show that two well-known stem cell factors, Foxd3 and Sox2, are critical regulators of the stem cell properties of SKPs. Deletion of Foxd3 completely abolishes the sphere-forming potential of these cells. In the absence of Sox2, SKP spheres can be formed, but with reduced size and frequency. Our results provide entry points into the gene regulatory networks dictating SKP behavior, and pave the way for future studies on a therapeutically relevant stem cell.
Introduction
Neural crest cells (NCCs) are a highly multipotent cell type with broad differentiation potential. The NC is specified in the neurula-stage embryo; these cells undergo an epithelial to mesenchymal transition and migrate along defined paths through the embryo. Their fate depends on their rostral-caudal position of origin in the neural tube, their route of migration, and final destination (reviewed in [1]). NC defects result in a number of developmental disorders, including CHARGE syndrome, Hirschsprung disease, Waardenburg syndrome, DiGeorge syndrome, congenital heart defects, and craniofacial abnormalities [2 –4]. Cell-based therapies may be appropriate for some of these syndromes.
Neural crest stem cells (NCSCs) persist through development, retaining their multipotency in adult organisms. NCSCs can be isolated from a number of embryonic and postnatal derivatives of the NC: dermis of the skin, fetal peripheral nerves, and the fetal and adult enteric nervous system [5 –9]. Of these, skin-derived precursors (SKPs) are of particular interest. SKPs are derived from the dermis of rodents and humans, and display a characteristic NC-like gene signature [9]. SKPs generated from the whisker pads of mice are NC-derived and can be lineage-labeled using Wnt1-Cre [10], a transgene expressed throughout the majority of the NC [11]. In vitro, SKPs exhibit a highly multipotent phenotype; they can differentiate into neurons, glia, smooth muscle cells, adipocytes, osteoblasts, and chondrocytes [9,10,12 –16]. The therapeutic efficacy of these cells has been suggested by rodent transplant studies; undifferentiated SKPs contribute to newly formed bone in a fracture model, “predifferentiated” SKPs assist in myelination of nerves in a sciatic nerve injury model, and these cells may also serve as an alternative source for cutaneous nerve regeneration [12,13,15,17 –19]. Importantly, these cells are readily accessible from adult humans and have the potential to serve as a patient-autologous stem cell source for a diverse array of cell-based therapies.
Despite these preclinical advances and the vast knowledge of transcription factor function in the NC [20], little is known about the molecules dictating NCSC self-renewal and multipotency; the “ground state” of these multipotent stem cells has not been widely explored. Foxd3 and Sox2 are logical entry points into the genetic regulatory networks governing SKP behavior. Sox2 expression can be used to prospectively isolate SKPs in addition to other progenitor cells [6,14,21], while loss of Foxd3 in the NC causes NC-progenitors to lose multipotency and self-renewal ability [22,23]. Null mouse embryos for either Foxd3 or Sox2 have virtually indistinguishable phenotypes, with loss of epiblast and an expansion of extra embryonic tissue, and both proteins are required for the establishment of ESCs and trophoblast stem cells (TSCs) [24 –26]. Finally, Foxd3 and Sox2 are known to antagonistically regulate shared loci in ESCs [27]. Given the prominent role of Sox2 and Foxd3 as key regulators of pluripotency in a number of stem cell populations, we examined the consequences of a genetic deletion of Sox2 in the NC and in NC-derived SKPs, and the role of Foxd3 in SKPs.
Materials and Methods
Mouse lines
The Wnt1-Cre transgenic line [28] was used to conditionally modify either the Foxd3 conditional allele Foxd3tm3.Lby
(called Foxd3flox
here) [23] or the Sox2 conditional allele Sox2tm2Lpev
(called Sox2flox
) [29] in combination with the reporter allele Gt(ROSA)26Sortm1
Cell culture
ESCs were cultured as described elsewhere [32]. SKPs were generated and cultured as previously described [9,12]. Briefly, appropriate mice were paired and monitored daily for the presence of a vaginal plug. Noon on the day of an observed plug was considered 0.5 days postcoitum (dpc). In order to generate SKP cultures we used standard methods [12]. Whisker pads or dorsal back skin were dissected from 16.5 to 18.5 dpc embryos. Tissues were digested in 0.1% trypsin/ethylenediaminetetraaceticacid for 30–45 min, mechanically dissociated, and passed through a 40-μM cell strainer. Single-cell suspensions were plated in Dulbecco's modified Eagle's medium/Ham's nutrient mixture F12 3:1 in the presence of 2% B-27 (Invitrogen), 40 ng/mL basic fibroblast growth factor (BD Biosciences), and 20 ng/mL epidermal growth factor (BD Biosciences). For experiments using the CreER transgene, a single dose of 0.2 μM 4-hydroxytamoxifen (TM) was added to the culture on day 0. The TM powder was dissolved in 100% ethanol to make a 1 mM TM stock solution and stored at −20°C. To quantify the number of SKP spheres, images of cultures were systematically captured as follows: at least 15 individual 40× frames of each culture were taken and the number of spheres per frame was determined. Data are presented as the average number of spheres per frame. N=3–5 for each genotype. For quantification of SKP sphere size, floating spheres were collected, pooled, and imaged. Spheres were outlined in ImageJ and the area was calculated in arbitrary units. At least 65 spheres were counted for each culture. Two-tailed Student's t tests were used to determine statistical significance.
Histology
Embryos were fixed in 4% paraformaldehyde overnight and standard histological procedures used [33]. Antibodies for immunofluorescence were rabbit-anti-Foxd3 (1:1,000) [26], goat-anti-Sox2 (1:500; Santa Cruz), and chicken-anti-GFP (1:500; Abcam). Secondary antibodies were from Jackson ImmunoResearch and 4′,6-diamidino-2-phenylindole (1:2,000; Molecular Probes) was used to visualize cell nuclei. Images were captured on either a Nikon Eclipse E600 or a Zeiss AxioCam MRc, both using Qcapture software. For quantification of dorsal root ganglia (DRG) size, serial sections of three pairs of DRGs at the level of the cardiac NC were examined for each genotype. DRGs were outlined using ImageJ software and the area calculated in arbitrary units. N=2 embryos for each genotype.
Quantitative real-time polymerase chain reaction
Total RNA was harvested from established SKP lines using the RNeasy Mini Kit (Qiagen) and genomic DNA removed using Turbo DNase (Ambion). cDNA was prepared using the GoScript™ Reverse Transcription System (Promega) and quantitative real-time polymerase chain reaction (qRT-PCR) performed using the GoTaq® qPCR Master Mix (Promega) and analyzed on an Applied Biosystems 7900 machine. Primer sequences were as follows: Foxd3, 5′-GTCC GCTGGGAATAACTTTCCGTA-3′ and 5′-ATGTACAAAGA ATGTCCCTCCCACCC-3′. Gapdh, 5′-TGAGGCCGGTGCTGA GTATGT-3′ and 5′-CCACAGTCTTCTGGGTGGCAGTG-3′. Sox2, 5′-GAGGAAAGGGTTCTTGCTGGGTTT-3′ and 5′-GGT CTTGCCAGTACTTGCTCTCAT-3′. Expression is presented as the difference in threshold cycle number of the transcript of interest relative to Gapdh.
Results
Sox2 and Foxd3 expression in NCCs and NCSCs/SKPs
The function of Sox2 in the NC has been largely overlooked. Early reports using in situ hybridization failed to detect Sox2 mRNA in NCCs [34]. However, several recent reports described roles for Sox2 in controlling the behavior of NC stem and progenitor cells: Sox2 maintains the progenitor status of NCCs fated to become melanocytes [35], and expression of Sox2 in the NC is essential for peripheral neurogenesis [36]. Given these recent findings, we re-examined the expression of Sox2 using more sensitive immunofluorescence techniques in combination with an NC-specific lineage label.
Sox2 protein is detected as early as 8.5 days dpc in pre-migratory and migratory cranial NCCs (Fig. 1A, B). This NC expression pattern is nearly indistinguishable from that of Foxd3 (Fig. 1C), a factor with established roles in the NC in multiple model systems [22,23,37 –40]. At 9.5 dpc, Sox2 is expressed at a low level in trunk NCCs as they emigrate from the neural tube (Fig. 1D, E). At 12.5 dpc, Foxd3, and Sox2 are co-expressed in the DRG, an NC-derived structure [41]. Our observations of Sox2 expression in the DRG are consistent with other reports [36,42].
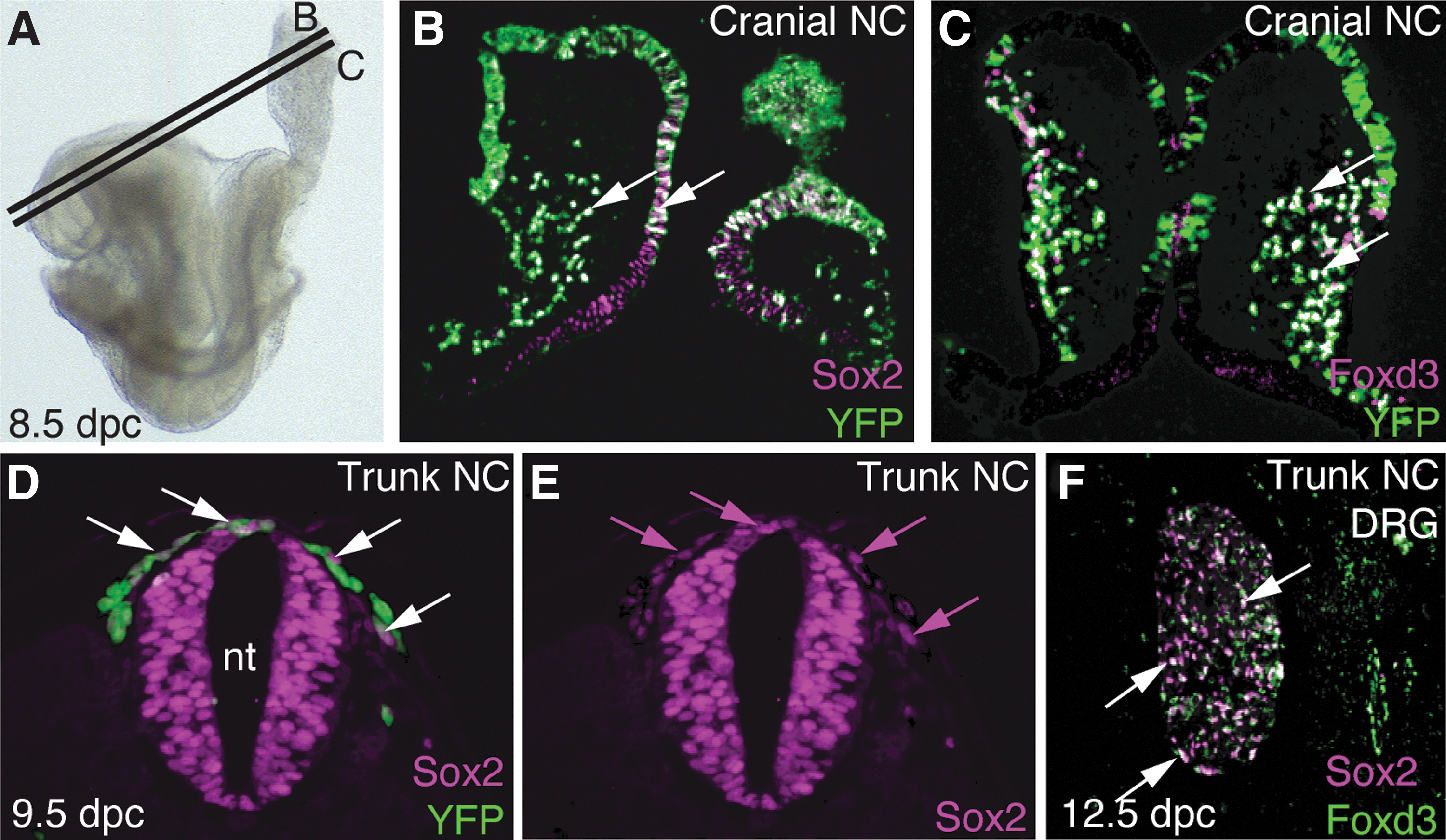
Sox2 and Foxd3 are expressed in NCCs.
SKPs are derived from the dermal papillae of mouse hair and whisker follicles [10], and can be prospectively isolated based on Sox2 expression [14]. Although Foxd3 expression has been reported in SKPs [43], it was unknown whether Foxd3 is expressed in the endogenous stem cell niche. Using immunofluorescence, at 12.5 dpc Sox2 was expressed in the NC-derived dermal condensates preceding whisker follicle morphogenesis (Fig. 2A–C). Foxd3 protein was localized to the periphery of the dermal condensates and was co-expressed with Sox2 (Fig. 2A, B, and D). At 17.5 dpc, an age when SKPs are readily generated, Sox2 is expressed in two distinct populations: (1) the NC-derived dermal papilla and dermal sheath, and (2) a non-NC lineage that was previously described as diagnostic of Merkel cells [44]. In contrast, Foxd3 is expressed exclusively in the NC lineage (Fig. 2E). It is not present in the dermal papilla, but was detected more distally in the whisker follicle, where it was co-expressed with Sox2. In cranial NC mesenchyme Foxd3 is expressed independently of Sox2 (Fig. 2F).
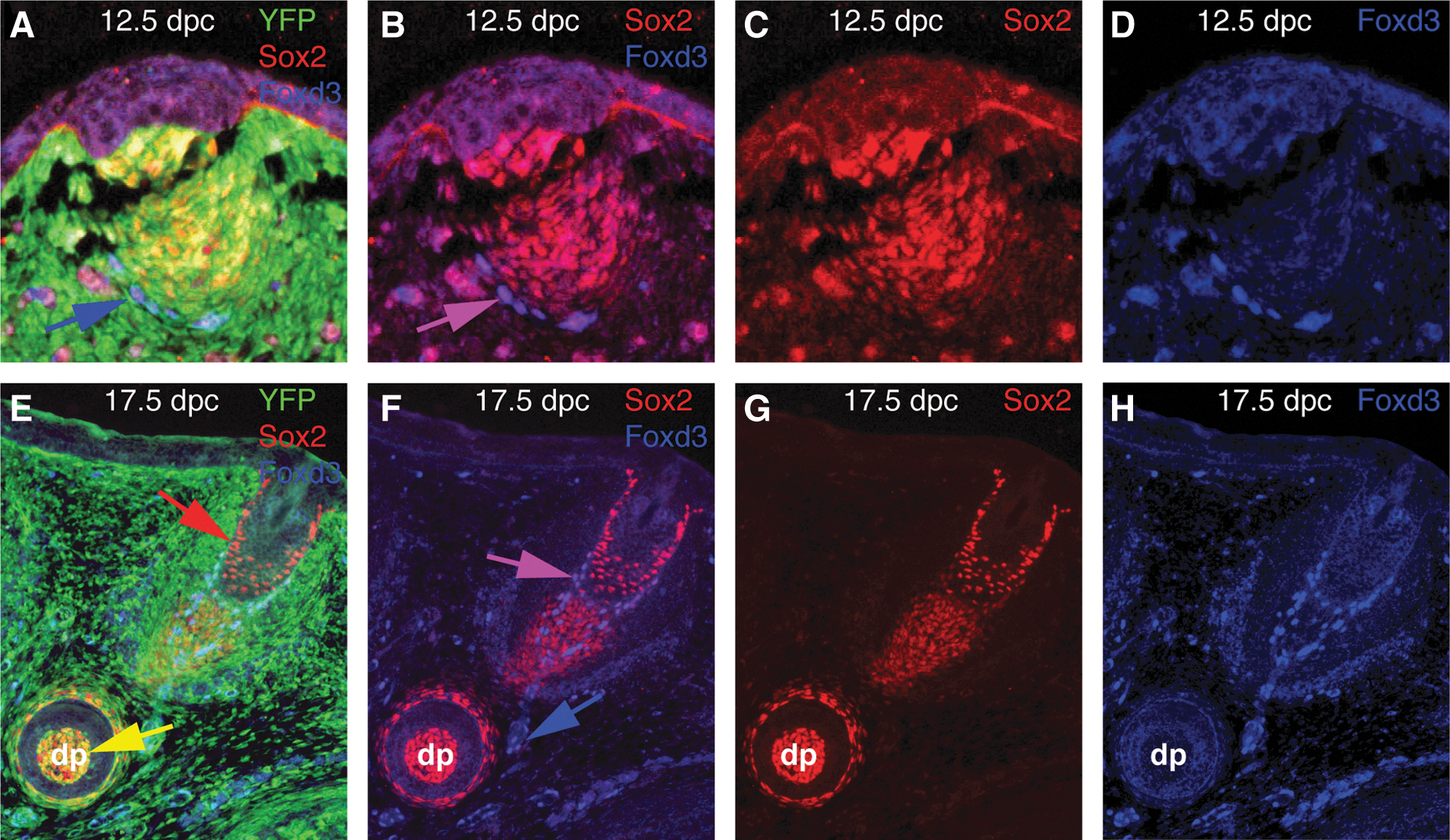
Sox2, but not Foxd3, is expressed in the endogenous SKP niche.
These results suggest the expression of Foxd3 is initiated during SKP establishment and is not present in the progenitors in vivo. To confirm this, Foxd3 expression was visualized in SKPs using a Foxd3GFP allele that has been used for this purpose in other contexts and faithfully represents the endogenous express of the locus in all contexts tested [45,46] (Fig. 3A, B). This was confirmed by analyzing Foxd3 transcripts in SKPs using qRT-PCR (Fig. 3C), consistent with previous reports [43]. As a positive control, qRT-PCR was used to show similar levels of Foxd3 transcripts in ESCs, a cell type that requires Foxd3 for self-renewal and inhibition of differentiation [32]. The expression of Sox2 and Foxd3 in NCCs is consistent with the notion that these two transcription factors function in NCSCs. Sox2 and Foxd3 are critical factors in multiple stem cell populations, but their role in NCSCs, and the role of Sox2 in the NC, had not been examined.
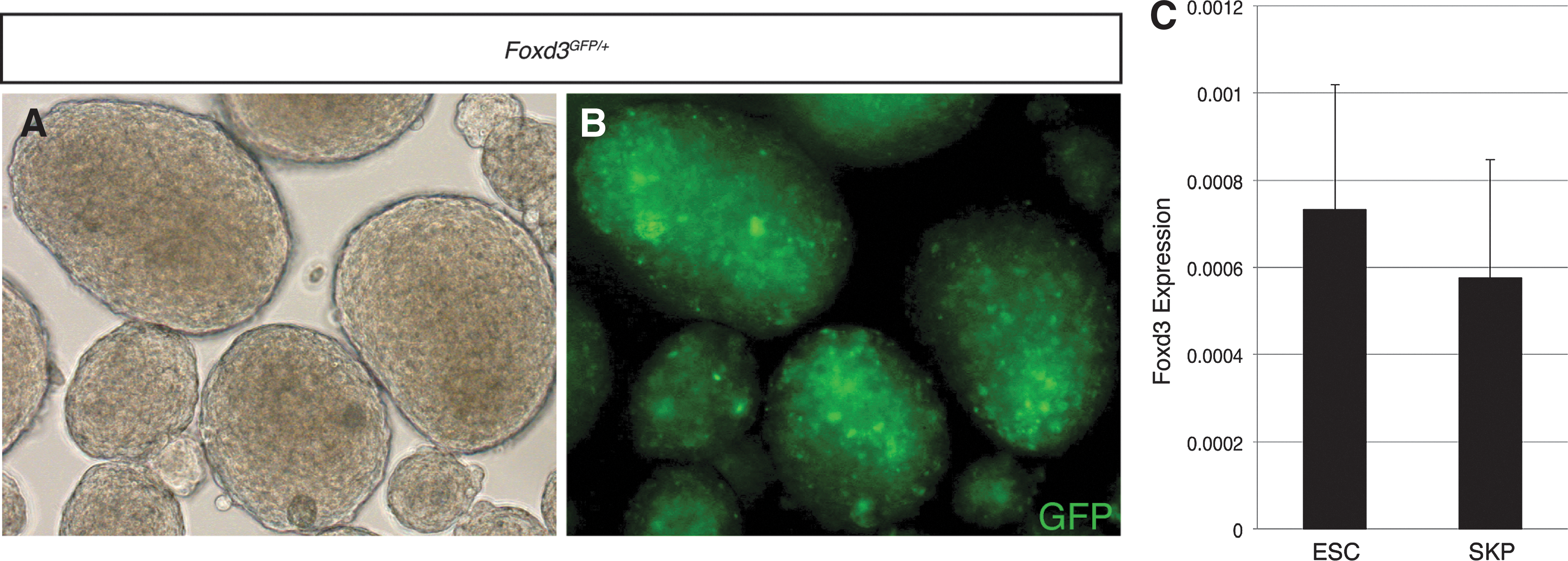
Foxd3 is expressed in SKPs.
A Wnt1-Cre-driven deletion of Sox2 results in perinatal death, but causes minimal morphological defects
The role of Foxd3 in the NC has been extensively studied; however, the function of Sox2 in the NC is largely unknown. We used Wnt1-Cre to drive deletion of Sox2 in the NC. In contrast to previous reports that showed embryonic lethality of these animals [36], mice derived from a Wnt1-Cre-driven deletion of Sox2 were born alive (n=4). However, these mice did not survive beyond 2 days. Sox2flox/flox ; Wnt1-Cre embryos had no detectable abnormalities in their craniofacial skeletons, nor did they exhibit any congenital heart defects (data not shown). As a potential cause of death, diaphragms were examined for proper innervation. At 16.5 dpc, no defects were seen with the innervation of Sox2flox/flox ; Wnt1-Cre diaphragms as visualized by PGP9.5 immunolocalization to mark neurons. In both Sox2flox/flox ; Wnt1-Cre embryos and Sox2wt/wt ; Wnt1-Cre littermate controls the phrenic nerves ranged the length of the diaphragms, extending to the proximal and distal tips, with numerous secondary branches (Fig. 4A, B). These data suggested that the cause of death is likely not due to respiratory defects. In addition to the NC, the Wnt1-Cre transgene is expressed in the developing midbrain and dorsal neural tube, both areas of robust Sox2 expression [28]. Thus, the cause of death may be due to neural tube defects, but at this time remains undetermined. Consistent with a previous report [36] Sox2 expression was absent in the DRGs of 16.5 dpc Sox2flox/flox ; Wnt1-Cre embryos (Fig. 4C, D). We also observed a 47% reduction in the area of the DRGs with a 2.36% standard error of the mean change, n=3 (Fig. 4E, F), again consistent with previous reports [36].
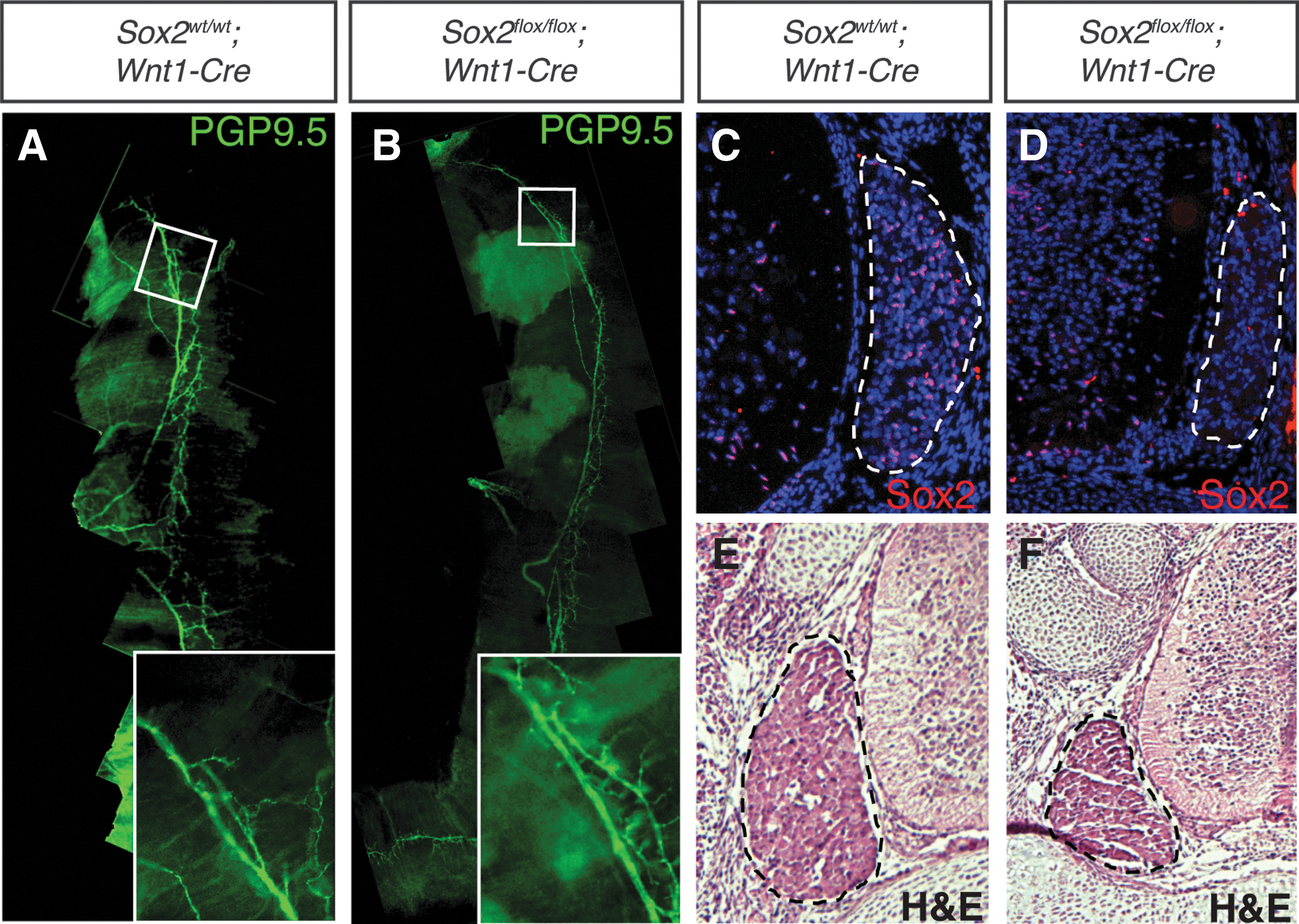
Sox2flox/flox
; Wnt1-Cre embryos showed normal innervation of the diaphragm, but reduced DRG size.
SKPs can be established without Sox2, but at a significantly reduced frequency
While SKPs can be prospectively isolated based on Sox2 expression [14], the requirement for Sox2 in SKPs had not been investigated. SKPs generated from whisker pads of mice are derived from NC and therefore are lineage labeled with the Wnt1-Cre transgene [10]. This Cre was used to determine the requirement for Sox2 in the establishment of SKPs.
First, to confirm Sox2 was deleted in the endogenous SKP niche, whisker pads were examined from 17.5 dpc Sox2flox/flox ; Wnt1-Cre embryos. Sox2 was no longer detected in the NC-derived portion of the whisker follicle (Fig. 5E), although Sox2 expression was unchanged in the non-NC portion. We observed no qualitative differences in the morphology of the follicles at a macroscopic or histological level. Co-expression of Sox2 and Foxd3 was completely lost in Sox2flox/flox ; Wnt1-Cre whisker pads (Fig. 5F). SKPs generated from whisker pads of Sox2flox/flox ; Wnt1-Cre embryos were modestly reduced in number compared to littermate controls; this difference was not statistically significant (Fig. 6A–D, quantified in Fig. 6E). Sox2 transcripts were reduced ∼14-fold in Sox2flox/flox ; Wnt1-Cre SKPs compared to Sox2wt/wt ; Wnt1-Cre littermate control SKPs (Fig. 6F). To extend this line of investigation, we used a TM inducible CreER to conditionally delete Sox2 from SKPs during establishment of the cultures. Cells from Sox2flox/flox ; CreER embryos treated with a single dose of 0.2 μM TM on day 0 of culture resulted in an 66% reduction in the number of spheres compared to untreated Sox2flox/flox ; CreER cells from the same embryo (Fig. 6G–K). TM treatment of cells from Sox2flox/flox embryos lacking the inducible Cre transgene showed no difference in the number of spheres generated compared to untreated cells from the same embryo. Additionally, spheres derived from TM-treated Sox2flox/flox ; CreER cells were reduced in size by 72% compared to untreated cells from the same embryo (Fig. 6L). These results demonstrate that SKPs are able to form in the absence of Sox2, but do so at a significantly reduced capacity.
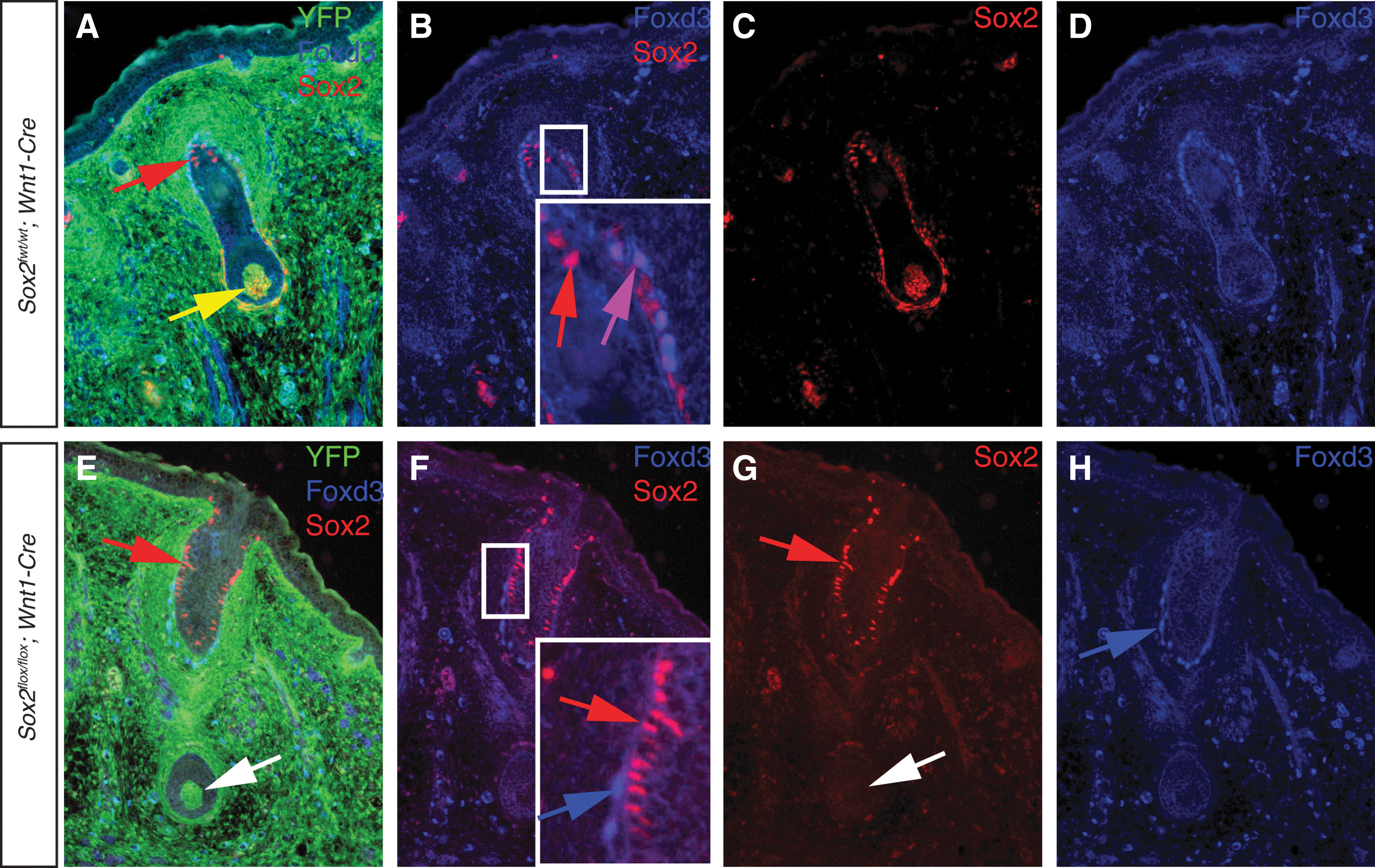
Sox2-expressing cells remain in whisker follicles of Sox2flox/flox
; Wnt1-Cre embryos.
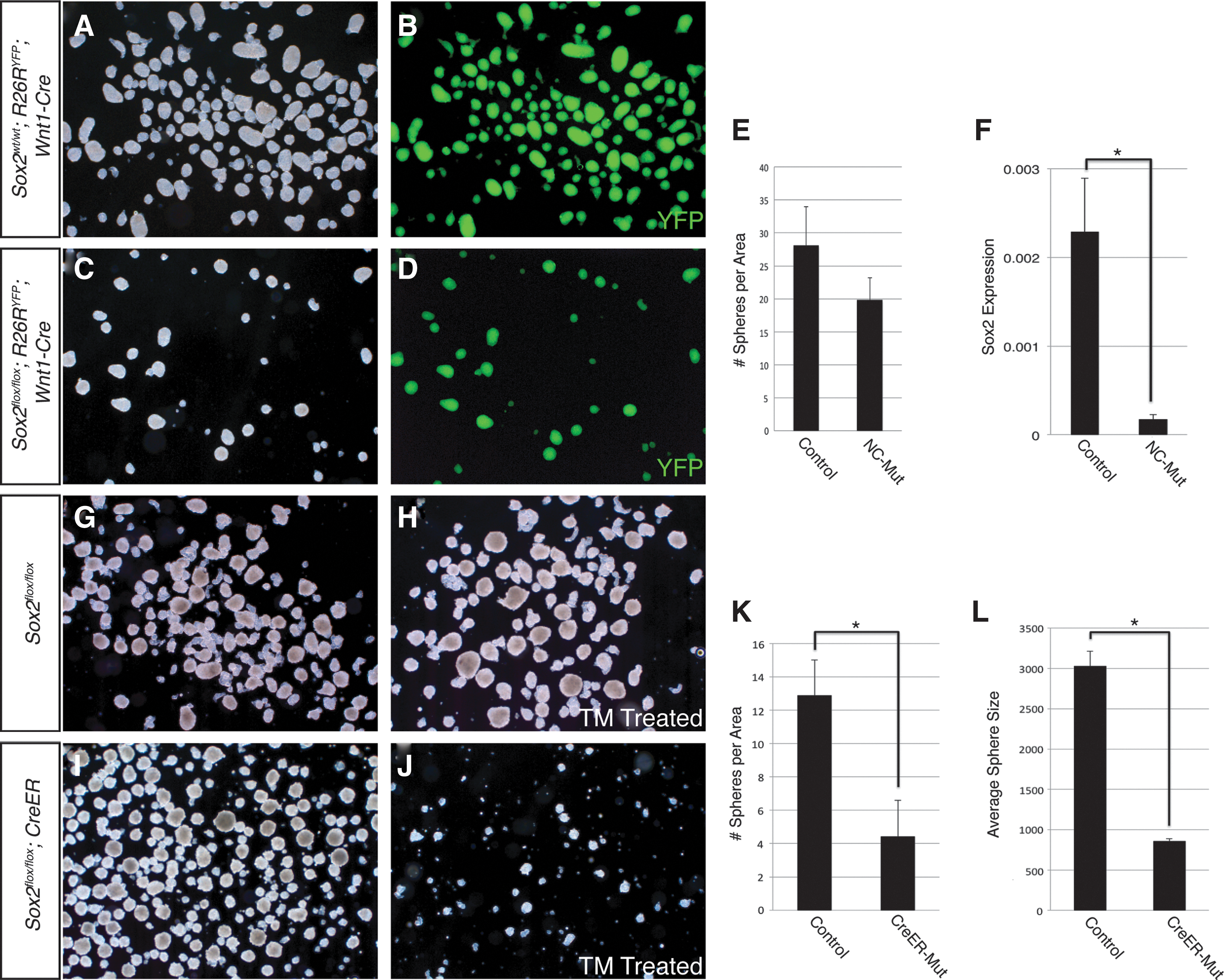
Deletion of Sox2 results in reduced sphere forming potential.
Foxd3 is required for the establishment of SKPs
Foxd3 is one of the earliest markers of NC [37,38,40,47 –50] and its role in NC maintenance has been well documented [22,23,38,51]; however, its role in NCSC/SKPs was unknown. As described above for Sox2, we tested the requirement for Foxd3 in establishment of SKPs. In control embryos heterozygous for Foxd3, all SKPs were derived from NC as shown using the Wnt1-Cre; R26RYFP lineage-label (Fig. 7A–C). In contrast, although numerous spheres were formed from whisker pads of Foxd3flox/flox ; Wnt1-Cre embryos (Fig. 7D–F), none of the resulting spheres were derived from NC, evidenced by a lack of YFP-positive spheres (Fig. 7E). Additionally, Foxd3 transcripts were detected at levels comparable to littermate controls (Fig. 7G). These data suggest that when Foxd3 is deleted, NC-derived SKPs are not established, yet spheres were generated from an as-yet-unidentified non-NC source.
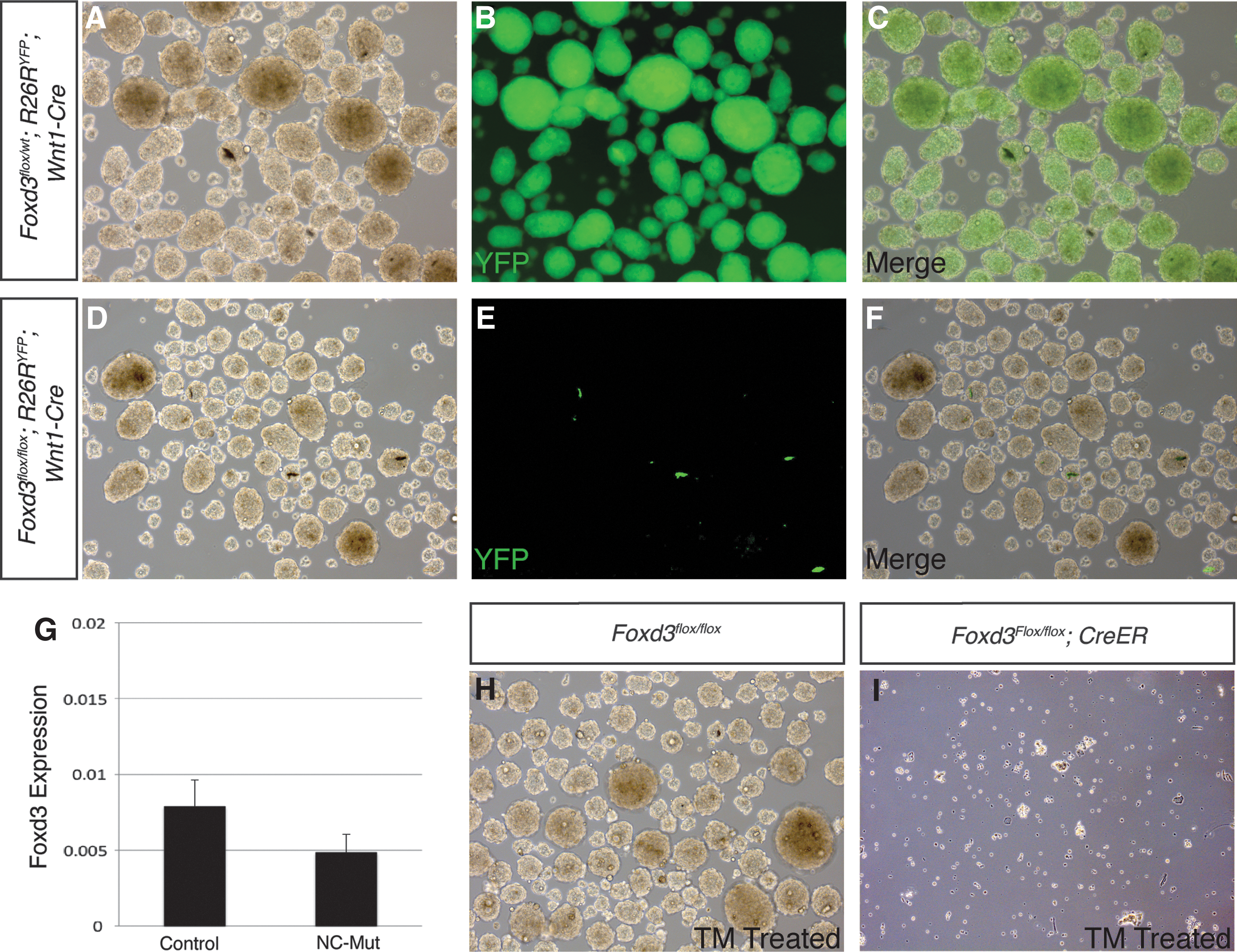
Foxd3 is required for establishment of SKPs.
To test the requirement for Foxd3 in the establishment of SKPs independently, SKPs were generated from Foxd3flox/flox ; CreER embryos. A single dose of 0.2 μM TM completely abolished the sphere-forming potential of these cells (Fig. 7H, I), while untreated cells and Foxd3flox/flox cells lacking the inducible Cre transgene treated with 0.2 μM TM behaved identically to control cells (Fig. 7G). These results indicate that Foxd3 is required for establishment of SKP cultures.
Discussion
Sox2 and Foxd3 are key regulators of pluripotency and multipotency in several different stem cell populations [24 –26]. Our findings suggest these factors function in a similar manner in NCSCs. As with ESCs and TSCs there is an absolute requirement for Foxd3 in the establishment of SKP cell lines. We found that SKP spheres are formed in the absence of Sox2, but at a severely diminished frequency.
Given the robust expression of Sox2 in the endogenous SKP niche, and the apparent absence of Foxd3 in the dermal papilla, it is somewhat surprising that deletion of Foxd3 from SKPs exhibits a more robust phenotype. Foxd3 is one of the earliest markers of the NC. Expression of Foxd3 is highest in uncommitted NC stem and progenitor cells, and is diminished as cells lose multipotency and differentiate [47,49]. Our observation that Foxd3 expression must be re-acquired in order to establish SKP cultures further confirms its role as a key factor controlling stem cell behavior.
There are a number of possibilities to explain the differences we describe between deletion of Sox2 with the Wnt1-Cre versus the CreER transgene. Sox2 is a prospective marker of SKP progenitor cells [14], yet in Sox2flox/flox ; Wnt1-Cre embryos, a population of Sox2-expressing cells remains in the whisker follicle. It is currently unknown whether these cells outside the NC are able to contribute to SKP formation. Additionally, SKP spheres are isolated by serial transfer of the floating spheres from the underlying adherent population of cells. However, these adherent cells are always re-established at each passage. The relationship between these support cells and SKPs is not understood and there could be a requirement for Sox2 in the support cells. These cells might not be targeted using the Wnt1-Cre and Sox2 might retain functionality in these cultures. The more dramatic phenotype seen with Sox2flox/flox ; CreER SKPs could account for both these possibilities. Alternatively, the difference could be due to the timing of deletion of Sox2. Sox2 is deleted much earlier in the embryo using the Wnt1-Cre as opposed to the CreER, and it is possible the SKP precursor cells have sufficient time to compensate for the loss of Sox2 while still in the embryo.
A unique feature of SKPs is that these transcriptionally similar stem cells can be generated from divergent embryonic origins: NC in the facial dermis versus mesoderm in the dorsal skin [43,52]. Recent work demonstrated that mesodermally derived SKPs are able to generate Schwann cells in vitro, a cell type thought to be exclusively derived from the NC [53]. Our findings are consistent with the possibility that cells of mesodermal origin may compensate for the lack of NC precursors in Foxd3flox/flox ; Wnt1-Cre embryos in vivo. Given the size, importance, and highly regenerative nature of the skin, it is perhaps not surprising that multiple mechanisms exist to generate the multipotent progenitor cell types in the skin.
In ESCs, Sox2 and Foxd3 regulate several shared loci, with Sox2 maintaining DNA in an open chromatin conformation, while Foxd3 actively represses transcription of these shared target genes [27]. This antagonistic relationship is hypothesized to keep target genes in a poised state, able to respond to differentiation cues in a rapid manner. It is currently unknown whether this relationship is conserved in NC progenitors or in SKPs. In fact, little direct regulatory information exists regarding the gene regulatory networks of NC and NCSCs. SKPs provide an abundant cell source for addressing these questions.
Conclusion
Being able to effectively control the self-renewal, proliferation, and differentiation of SKPs is key to advancing these cells toward clinical use. Toward this goal, we have demonstrated here that two well-known stem cell proteins, Foxd3 and Sox2, are important regulators of the stem cell properties of SKPs. This study provides the framework to move forward our understanding of the molecular controls of self-renewal and multipotency in a potentially therapeutic stem cell.
Footnotes
Acknowledgments
We thank Alison LeGrone for technical assistance, Dr. Scott Baldwin for cardiac analysis of the Sox2 mutant embryos and newborn mice, and Dr. Thomas Andl for reading of the manuscript. We also thank Dr. Mark Magnuson for use of his Applied Biosystems 7900 machine for qRT-PCR and his Zeiss AxioCam MRc microscope. This work was supported by grants from the NIH (HD36720, HD036720-11S109 and a pilot grant from 5P01GM085354) to P.A.L., NIH support for M.T.S. (T32HD007043) and E.R.P. (T32HD007502), predoctoral support from the American Heart Association (12PRE10950005) to E.R.P., predoctoral fellowships from the AHA (0615209B) and NIH (NS065604) to N.A.M., and Vanderbilt University Medical Center Academic Program Support. All the work carried out here was performed at Vanderbilt University. Dr. Larysa Pevny passed away during the time this manuscript was being revised—she will be missed.
Author Disclosure Statement
We declare no potential conflicts of interest.