
Abstract
Neuronal stem cells residing in a niche on the surface of the adult crayfish (Procambarus clarkii) brain are not self-renewing. However, the neuronal precursors in the niche are not depleted despite continued neurogenesis and the exit of precursor cells from the niche throughout the organism's life. The neurogenic niche is therefore not a closed system, and we have previously proposed that the stem cell pool is replenished from the hematopoietic system. Noonin et al. (2012) demonstrated that the hematopoietic system in the crayfish Pacifastacus leniusculus includes an anterior proliferation center (APC) lying near the brain; they suggest that multipotent stem cells are concentrated in this region, and that the APC may provide neuronal stem cells for adult neurogenesis. The present study extends this work by describing the location and cellular organization of hematopoietic tissues in P. clarkii. We find that the APC lies within the cor frontale, or auxiliary heart, which pumps hemolymph to the brain and eyes through the cerebral and ophthalmic arteries, respectively. Vascular extensions of the cerebral artery converge on the neurogenic niche. APC cells lie in layered sheets within the cor frontale and form rosette-like structures reminiscent of stem cells in other developing tissues. We confirm here that APC cells in P. clarkii have characteristics of multipotent stem cells, and that their location within the cor frontale allows direct access to regions in the central nervous system in which adult neurogenesis occurs.
Introduction
Adult neurogenesis in crayfish
Studies in our laboratory have shown that cells circulating in the hemolymph, and not cells from other tissues (e.g., hepatopancreas and green gland), are attracted to the neural stem cell niche in vitro [6]. These findings have led to the hypothesis that cells associated with the hematopoietic system and that circulate in the hemolymph may replenish the neuronal precursors in the niche. A close association between the niche and the vasculature is established early, at the time the anlage of the niche appears at the end of embryonic life and in concert with angiogenesis in that region [7]. Morphological and ultrastructural observations confirm that the neurogenic niche in the mature brain is nearly encompassed by blood vessels that are embedded in connective tissues, suggesting a retia-like complex of fine channels between the niche and the vasculature emerging from the underlying accessory lobe [8]. The close association between the niche and vascular system also has been confirmed by experiments showing that dye-labeled dextran injected into the dorsal artery [3] or into the pericardial sinus [6] infiltrates a cavity located centrally in the niche, which has thus been named the vascular cavity. These known relationships between the vasculature and the niche are consistent with the possibility that stem cells circulating in the hemolymph could readily gain access to the niche.
The anterior proliferation center
The recent work of Noonin et al. [9] in the crayfish Pacifastacus leniusculus demonstrates that the hematopoietic system in crustaceans is far more extensive than previously recognized. In addition to well-known hematopoietic tissues located dorsally just beneath the carapace and covering the stomatogastric mill, Noonin and collaborators [9] have identified a specialized region of the hematopoietic system that is located near the brain and which they propose constitutes a multipotent stem cell center. This anterior proliferation center (APC) is distinct from the rest of the hematopoietic tissue (HPT) in several ways: The highest rate of BrdU incorporation is in the anterior part of the HPT and the APC. In addition, the majority of cells in the APC contain nuclei with loose chromatin (euchromatin), whereas most cells in the posterior HPT contain cells with condensed chromatin (heterochromatin). It is suggested that this difference in the chromatin structure is related to the degree of differentiation of the cells, with DNA becoming more condensed as the cells differentiate [9,10]. Further, reactive oxygen species (ROS), which induce the differentiation of hematopoietic stem cells in Drosophila [11] and in the mammalian myeloid lineage [12], are produced in high levels only in the most anterior part of the HPT, between the APC and the brain [9]. ROS labeling increased throughout the APC at 30 min following laminarin injection into crayfish, a treatment that mimics fungal infection. This timing corresponds with the decrease in circulating hemocytes and subsequent recruitment of new hemocytes from the HPT associated with laminarin treatment, suggesting that increased metabolic activity may be related to the differentiation of cells in the APC. Based on these and additional data, Noonin et al. [9] propose that cells in the APC are the multipotent stem cells of the crustacean HPT. Because of their properties and their location, these cells are of special interest in our efforts to identify the source of first-generation neuronal precursors that produce adult-born neurons.
This article extends the work of Noonin et al. [9] by localizing the APC in the closely related crayfish P. clarkii, and by characterizing the cellular organization and behavior of cells in the APC compared with more posterior hematopoietic tissues. Our studies show that the APC is localized within the cor frontale or auxiliary heart (Fig. 1A, B), which provides direct vascular access to the brain and eyes via the cerebral and ophthalmic arteries, respectively. Our data also show that secondary branches from the cerebral artery contact the neurogenic niche and streams, infiltrating these areas. Further, we show that unlike the more posterior HPTs that extend over the gastric mill, regions within the APC are characterized by proliferating cells that form rosette-like structures. In vertebrates, morphologically similar structures called neural rosettes are the developmental signature of neuroprogenitors in cultures of differentiating embryonic stem cells [13,14] and in neurogenic regions in the adult brain, resembling patterns seen in other developing epithelial tissues [15]. Based on these findings, we hypothesize that rosette cells composing the APC are indeed multipotent stem cells and that these are prime candidates to replenish the pool of first-generation neuronal precursors in the neurogenic niche as these divide and their daughters migrate away.
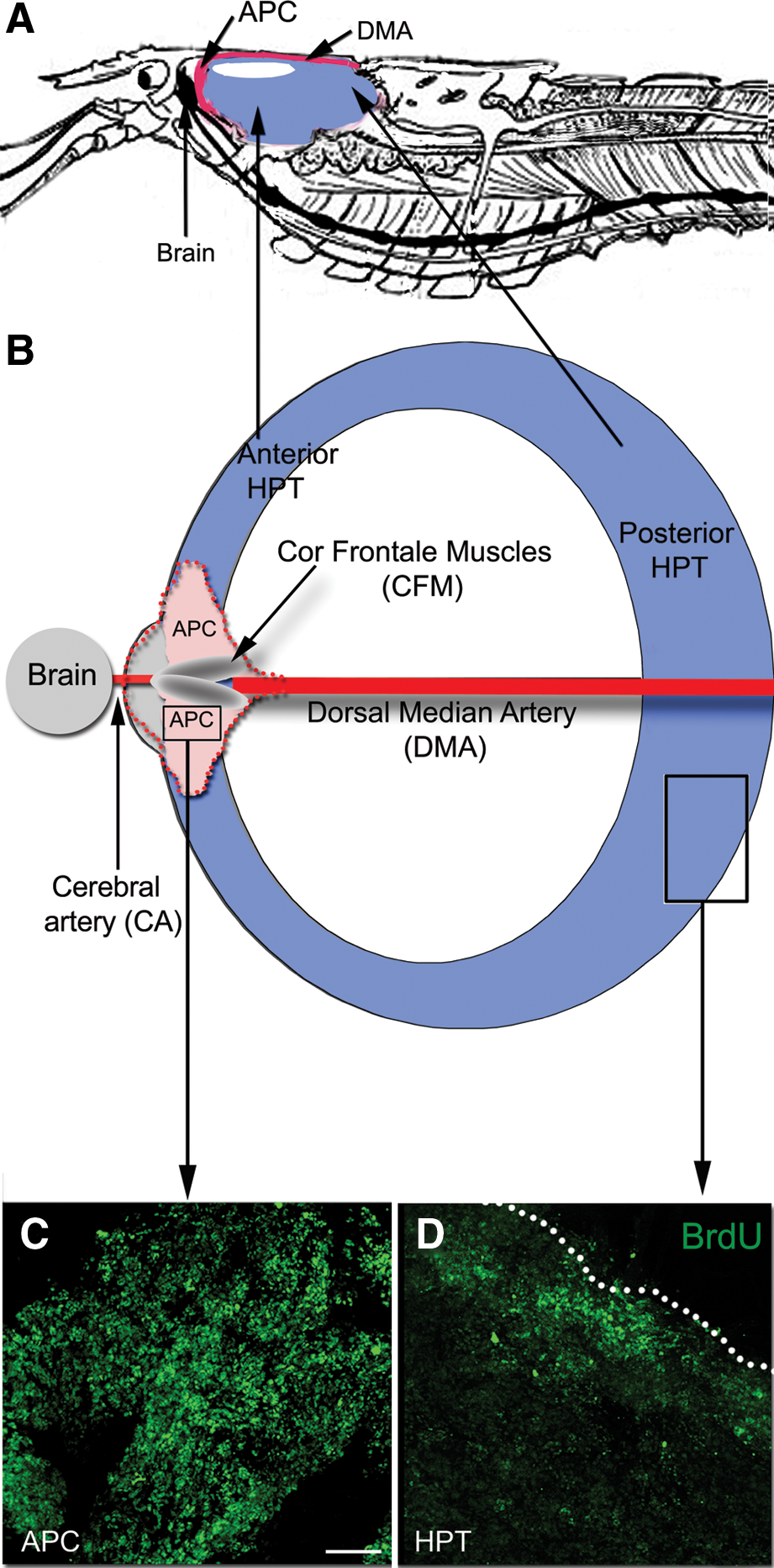
Location and proliferative characteristics of P. clarkii hematopoietic system.
Materials and Methods
Animals
Adult male and female crayfish P. clarkii (carapace length [CL] 15–30 mm) were obtained from Carolina Biological Supply Company and maintained at room temperature in aquaria containing aerated artificial pond water and exposed to a light:dark cycle of 12:12 h. Crayfish of this size are sexually mature adults.
BrdU timed series: cell proliferation studies of the APC
The BrdU (Sigma-Aldrich) labeling reagent (5 mg/mL in saline) was injected (200 μL) into the base of the third walking leg of 12 crayfish (CL 28–30 mm). At four intervals following injection (3, 6, 24, and 72 h), three crayfish were killed and the brain and cor frontale (auxiliary heart) were dissected and fixed overnight in 4% paraformaldehyde in 0.1 M phosphate buffer (PB; pH 7.4) at 4°C. Tissues were rinsed several times in PB with 0.3% Triton X-100 (PBTx) followed by a 30-min treatment in 2 N hydrochloric acid. After rinsing again in PBTx, tissues were incubated for 18 h at 4°C in mouse monoclonal (Clone MoBU-1) anti-BrdU conjugated to Alexa Fluor® 488 (Invitrogen; B35130). After rinsing in PBTx, tissues were treated with the nucleic acid stain propidium iodide (PI; Sigma-Aldrich; 10 μg/mL PB) for 10 min at 18°C–20°C. After rinses in PB, the preparations were mounted in Fluoro-Gel (Electron Microscopy Sciences) and examined and imaged using a Leica TCS SP5 confocal microscope equipped with argon 488-nm and 561-nm and 633-nm diode lasers. Serial optical sections were taken at 0.5–1-μm intervals and saved as both three-dimensional stacks and two-dimensional projections.
Detection of ROS and localization of mitochondria
ROS production in APC tissue and posterior HPT was monitored in live tissues using the protocol of Noonin et al. [9]. Saline (200 μL) was injected into the pericardial sinus of crayfish (∼18 mm CL) 1 h before dissection to stimulate a brief hematopoietic response [9]. Posterior HPT and APC tissue were then dissected in oxygenated crayfish saline. Freshly prepared 2′,7′-dichlorofluorescein diacetate (D6883; Sigma) in dimethyl sulfoxide (DMSO; 5 mg/mL; Sigma) was diluted with 0.1 M PB (1:1,000; 10 μM concentration). Whole, dissected HPTs were incubated with the detector for 10 min with gentle shaking in a dark chamber at room temperature. Three 5-min rinses in PB followed by mounting in 30% glycerol in PB allowed for immediate observation of the tissues with a Nikon 80i fluorescent stereomicroscope.
While there are several nonmitochondrial sources of ROS, mitochondria are major contributors, permanently producing ROS as a by-product of oxidative phosphorylation [16]. MitoTracker® Deep Red (Molecular Probes; FM M22426), a cell-permeant probe, contains a mildly thiol-reactive chloromethyl moiety for labeling mitochondria and is retained in the mitochondria after fixation. We therefore used this agent to localize mitochondria in cells in the APC and posterior HPT, as one indicator of metabolic activity and the potential for higher ROS production. One hour before dissection, saline (200 μL) was injected into the pericardial sinus of crayfish (∼18 mm CL) to stimulate a hematopoietic response. A labeling solution of MitoTracker® at a working concentration of 100 μM in crayfish saline was prepared. Freshly dissected tissues (n=8) were immersed in the labeling medium for 30 min at 37°C, rinsed in PB, and fixed with 4% paraformaldehyde in PB (pH 7.4), followed by staining with the nucleic acid marker 4′,6-diamidino-2-phenylindole (DAPI; Molecular Probes).
Morphological examination of fixed tissue
The posterior and anterior regions of HPT were carefully dissected from crayfish (CL 20 mm) in cold saline (205 mM sodium chloride, 5.4 mM potassium chloride, 34.4 mM calcium chloride, 1.2 mM magnesium chloride, and 2.4 mM sodium bicarbonate) and fixed overnight in 4% paraformaldehyde (pH 7.4). To maintain the structural integrity of the APC, fix was injected medially into the base of each eyestalk a few minutes before dissection. Tissues were visualized and imaged using an Insight-spot 2 camera coupled to a Nikon SMZ 1500 stereomicroscope.
Immunohistochemical localization of tubulins
Fixed APC tissue and posterior HPT were rinsed multiple times with 0.3% PBTx and incubated overnight at 4°C in either mouse monoclonal anti-β tubulin (1:10; Developmental Studies Hybridoma Bank, No. E7) or mouse monoclonal anti-tyrosinated tubulin (1:1,000; Sigma; T-6557), which reacts with tubulin's carboxy-terminal tyrosine. Tissues were rinsed with 0.3% PBTx for 2 h and incubated in goat anti-mouse IgG Cy2 (1:100; Jackson Immunoresearch) at room temperature for 4 h. After rinsing with 0.3% PBTx, they were counterstained with PI (10 mg/mL; 10 min at 18°C–20°C; Sigma). The samples were then rinsed with PB and mounted with Fluoro-Gel (Electron Microscopy Sciences). Preparations were viewed and images captured with a Leica TCS SP5 confocal microscope.
BrdU labeling of cells in the dorsal artery and APC
Crayfish (CL 15–20 mm) were maintained in pond water containing BrdU (2 mg/mL) for 24 h before sacrifice. Tissues were fixed and processed immunocytochemically as described under the “BrdU timed series: cell proliferation studies of the APC” heading except that the nucleic acid marker DAPI was used instead of PI.
Dextran fills of the vasculature
Labeling of the brain vasculature in crayfish was achieved by injecting fluorescein isothiocyanate (FITC)-conjugated dextran (3,000 MW; Invitrogen; D-3306; 500 μL of a 1 mM solution in crayfish saline) into the pericardial sinus. Animals (15–20 mm CL; n=5) were placed in artificial pond water for 5 min following injection. The brain, cor frontale and associated APC were dissected in saline and fixed in 4% paraformaldehyde overnight at 4°C. Using standard immunohistochemical techniques, tissues were labeled with mouse anti-glutamine synthetase (1:100; Becton Dickinson; 610517) followed by overnight incubation in goat anti-mouse CY5 (1:100; Jackson Immunoresearch), and then stained with the nuclear marker PI.
Data analyses and statistical treatment
The numbers of PI- and BrdU-labeled cells were quantified from stacked images obtained with a Leica TCS SP confocal microscope. A single optical section was projected onto the monitor and the labeled cells traced onto a transparent sheet to count the numbers of labeled cells. The area of each APC tissue imaged was standardized so that each region (i.e., a stack) was a 130-μm2 region (one optical section at 120×magnification) with a depth of 4 μm. Percentages of BrdU-positive cells and mitotic profiles were calculated relative to the total number of PI-labeled cells and mitotic profiles, respectively. The number of BrdU-labeled rosettes was counted in three different areas bilaterally of the APC (see diagram Fig. 6A). The cor frontale muscles (CFMs), medially located in the APC, were used as a reference point to select these regions. Rosette labeling was classified into three different types: Center—only the center cell within the rosette was labeled with BrdU; Partial—BrdU labeling in the center cell plus two or three cells around the center cell; and Full—all of the cells composing the rosette were labeled. Counts of the rosette types and mitotic profiles were taken at four different time points after BrdU injection (3, 6, 24, and 72 h) and tested for differences using a one-way analysis of variance (ANOVA) followed by Tukey's multiple comparisons (GraphPad Software).
Results
Because of the potential importance of the hematopoietic system for adult neurogenesis in crustaceans, in the present study we have localized these tissues in the crayfish P. clarkii, a species in which mechanisms underlying the production of adult-born neurons have been carefully examined [17]. The posterior HPT in P. clarkii, like that described in P. leniusculus [9], spreads over and around the stomatogastric mill. The HPT also extends forward toward the brain and part of it can be found lying between the anterior part of the gastric mill and the cor frontale, or auxiliary heart [18,19]; this is the anterior HPT described by Noonin et al. [9] in P. leniusculus. Dissections of the cor frontale and confocal examination of fixed tissues in P. clarkii also revealed highly proliferative tissue lying within the cor frontale. The location, extent of BrdU incorporation, tests for ROS and other morphological features of this tissue in P. clarkii lead us to believe that this is equivalent to the APC discovered in P. leniusculus [9]. The studies presented here also extend previous findings by characterizing the regional and cellular organization of the APC and examining the vascular route by which APC cells travel to and access brain tissues.
Vasculature associated with the hematopoietic system: nomenclature
The following account uses the terminology of Steinacker [19] in referring to the blood vessels associated with HPT and the cor frontale. Accordingly, the dorsal median artery (DMA) is the primary median vessel from the heart to the brain, extending anteriorly just beneath the carapace to a point close to the rostrum where it turns ventrally and descends toward the brain (Fig. 1A). Just dorsal to the brain, the DMA distends and forms a relatively large sac (the cor frontale) that encloses the longitudinally-oriented CFMs (Fig. 1B). Three blood vessels emerge from the cor frontale: two lateral vessels (the ophthalmic arteries) go to the optic ganglia in the eyestalks, and a medial branch (the cerebral artery, CA) projects to the brain. Historically, a variety of terms have been used by various authors in referring to these vessels, with no general consensus as to their assignments. Thus, the DMA also has been called the median artery [18,20,21] or the ophthalmic arteries [22 –24], the latter of which is easily confused with the true ophthalmic arteries servicing the eyes and which emerge from the cor frontale. Because HPTs are intimately associated with these vessels, the designations of Steinacker [19] will be used in this article, as these terms are unambiguous and permit a precise assignment of location.
Organization of HPTs in P. clarkii
Most crustacean species have a distinct hematopoietic organ that is composed of a series of ovoid lobules that collectively form a thin sheath of connective tissue, found in close proximity to the DMA as it extends on the dorsal surface of the foregut just beneath the carapace (hereafter referred to as the posterior HPT; Fig. 1A, B). In P. clarkii, HPT is located in this typical position, as in P. leniusculus [24] and other species [22]. Proliferating hematopoietic cells (HPCs) tend to be located at the edges of the HPT (Fig. 1D), from which they are released into the surrounding sinus. In P. clarkii, cells composing these lobules range in size from 10 to 15 μm.
Two other distinct regions of HPT, the anterior HPT and the APC (Fig. 1A, B), recently described in P. leniusculus [9], are readily identifiable in P. clarkii on the basis of location and relative levels of proliferative activity. At roughly the position where the dorsal artery distends to form the cor frontale, the HPT forms extensions, defined by Noonin et al. [9] as the anterior HPT (Fig. 1A, B). Cells in the anterior HPT in P. clarkii are organized similarly and are of the same size as cells in the posterior HPT.
The APC, which Noonin et al. [9] report as being connected to the brain, is readily identifiable in P. clarkii. Timed studies were conducted where the S-phase marker BrdU was injected into crayfish, followed by sacrifice of groups of crayfish at 3, 6, 24, and 72 h post-injection (Fig. 2). These time points were chosen to occur after any transient evoked hematopoietic response, which peaks by ∼1 h after injection of fluid [25,26]. It should be noted that the BrdU clearing time in P. clarkii is relatively long, between 30 and 48 h [6], and therefore the increasing times between 3 and 24 h represent longer exposures to BrdU, while the 72-h time point would have occurred after the available BrdU had cleared. More than 10% of cells in the P. clarkii APC label 3 h after BrdU injection (Fig. 2A, F), and 40%–45% of cells label with BrdU at 24 h post-injection (Fig. 2C, F). The highest percentages of mitotic figures were labeled at the 6- and 24-h time points (Fig. 3), which suggests that the time from S- to M-phase is at least 6 h long. Alternatively, the maximal BrdU labeling of mitotic figures at the 24-h time point could reflect multiple divisions of cells with a shorter cell cycle, and therefore may represent accumulated, multiple generations. The vast majority of cells in the APC range in size from 8 to 12 μm, with occasional cells reaching up to 20 μm; these unusually large cells are often labeled with BrdU, and probably represent cells that have duplicated their DNA and grown in size in preparation for mitosis.
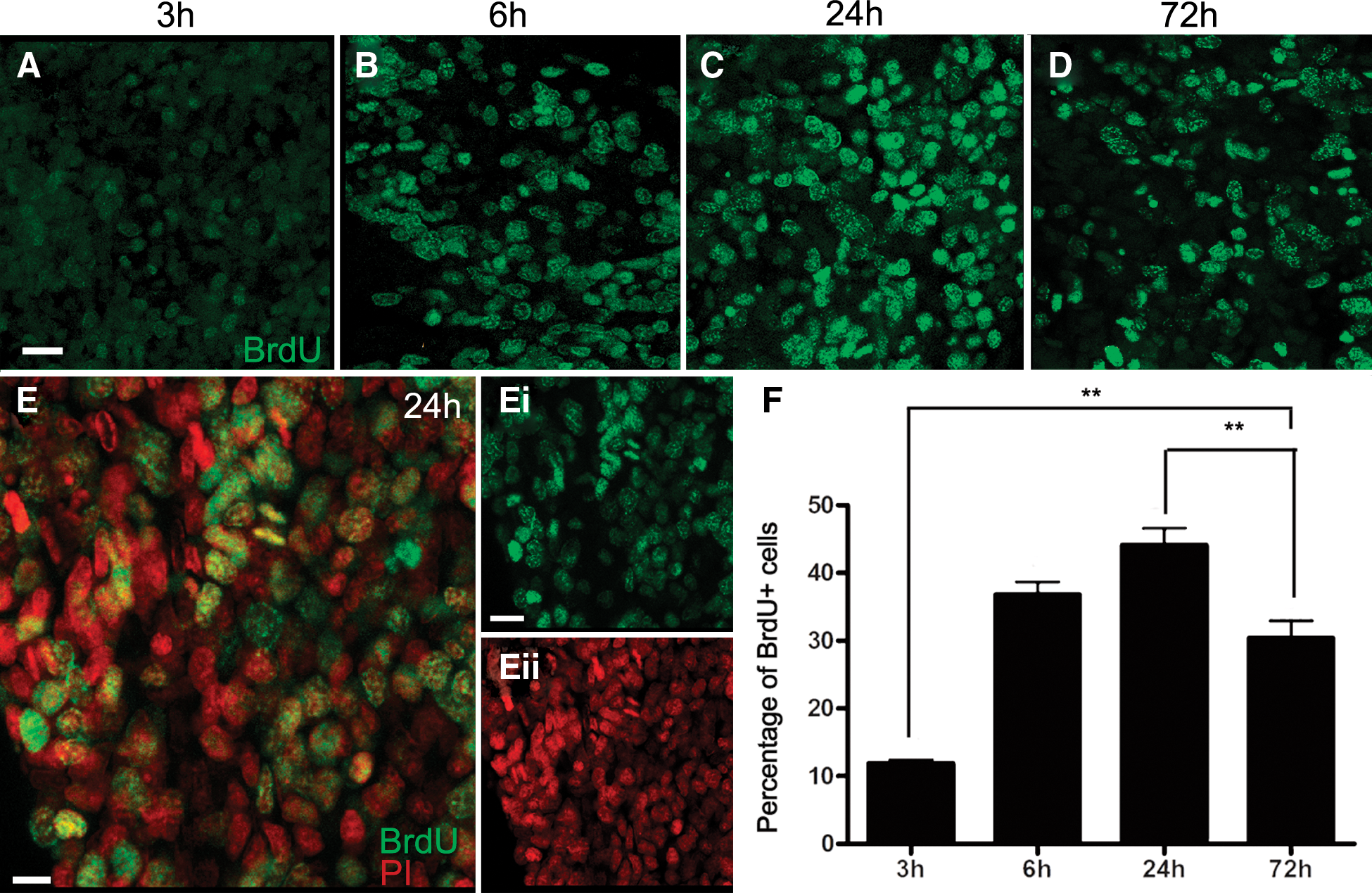
BrdU labeling of cells at four time points following injection of BrdU into adult crayfish.
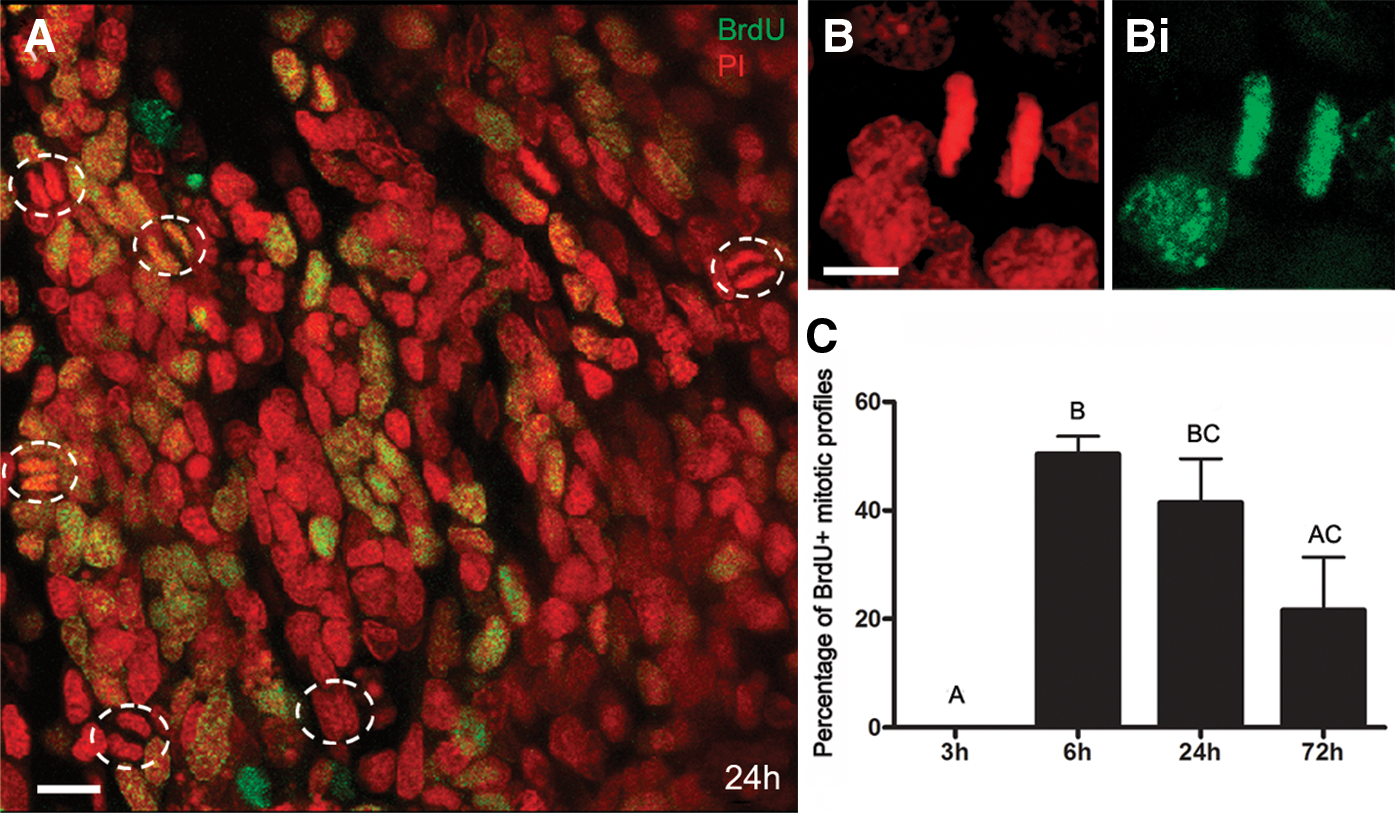
The percentage of mitotic profiles increases to a maximum at 24 h.
ROS labeling in the APC and posterior HPT
The production of ROS, which is indicative of the metabolic state of cells, was monitored in live APC tissue and posterior HPT treated with 2′,7′-dichlorofluorescein diacetate. High levels of ROS production were detected in the APC, but ROS activity was not detected in the brain (Fig. 4A). A weak ROS signal was limited to the peripheral margins of the posterior HPT.
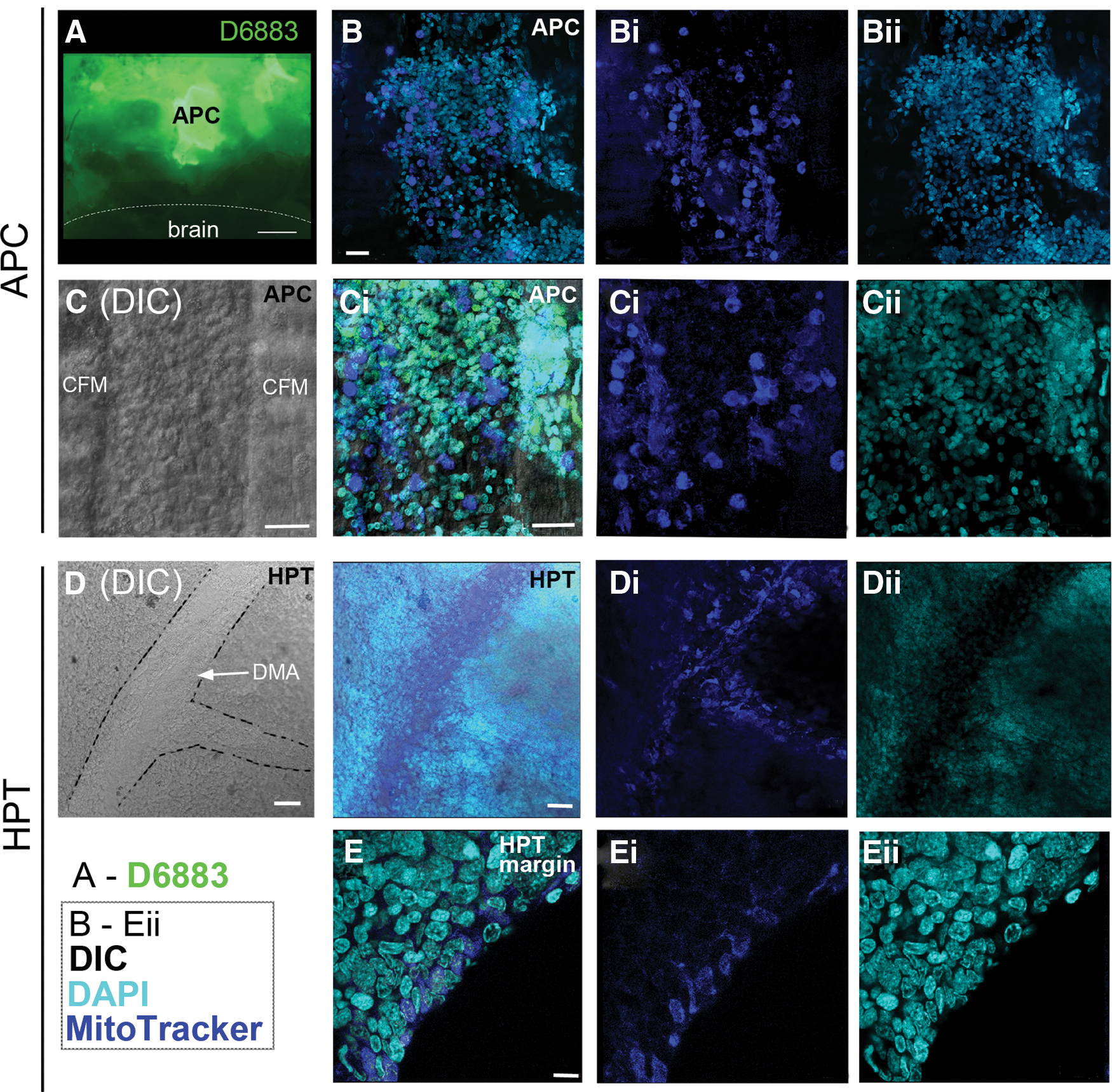
Labeling of reactive oxygen species (ROS) and mitochondria in APC and HPT cells.
MitoTracker® was used to label mitochondria in live tissues followed by fixation, as another indicator of metabolic activity and the potential for higher ROS production. APC cells contained intense MitoTracker® labeling (Fig. 4B, C), a finding that parallels the detection of high ROS levels in this tissue (Fig. 4A) [9]. Cells were particularly intensely labeled in the region on top of and between the CFMs (Fig. 4C).
In parallel with high levels of BrdU labeling along the lateral margins of the posterior HPT (Fig. 1D), a few cells in this same location contained moderate levels of MitoTracker® labeling (Fig. 4E). More central regions of the posterior HPT were not labeled, suggesting that metabolism and cell cycle activity are maximal at the margins of this tissue, as previously reported by Noonin et al. [9]. MitoTracker® also labels cells in the DMA (Fig. 4D, Di) and in small vessels connected to the DMA, suggesting that cells circulating in the hemolymph maintain high metabolic activity, perhaps because they are undergoing their final differentiation [24] or a few cells may still be in the cell cycle.
The high degree of cell cycle activity in cells near and surrounding the CFMs, their location between the anterior HPT and the brain, and high levels of ROS and mitochondrial labeling in P. clarkii are consistent with characteristics of the APC in P. leniusculus, thus confirming the identity of this tissue as equivalent to the APC described by Noonin et al. [9].
The APC in P. clarkii is located within the cor frontale
A striking feature of the APC in P. clarkii is its location inside the cor frontale (Fig. 5). In preparations where the brain, cor frontale, and all three regions of the P. clarkii hematopoietic system (posterior HPT, anterior HPT, and APC) were dissected and laid flat for observation (Fig. 5B–D), the spatial relationships between these tissues could be examined. APC tissue wraps around and between the CFMs (Fig. 5B, D) and sheets of cells (Fig. 5A, purple; B, C, dotted outline) form lateral extensions attached to the dorsal and ventral walls of the cor frontale. The CFMs originate within the rostrum on the dorsal carapace and are anchored caudally to a fibrous strap lying across the dorsoanterior edge of the brain. Two fine ligaments extend posteriolaterally from the caudal ends of the muscles. The cor frontale itself extends sideways, ventral to the large muscle blocks holding the gastric mill anteriorly in the animal, and attaches laterally to the exoskeleton (see Fig. 5E). In studies where saline was injected into the cerebral artery to inflate the cor frontale, it was very clear that the sheer membrane to which APC cells are attached forms the walls of the cor frontale.
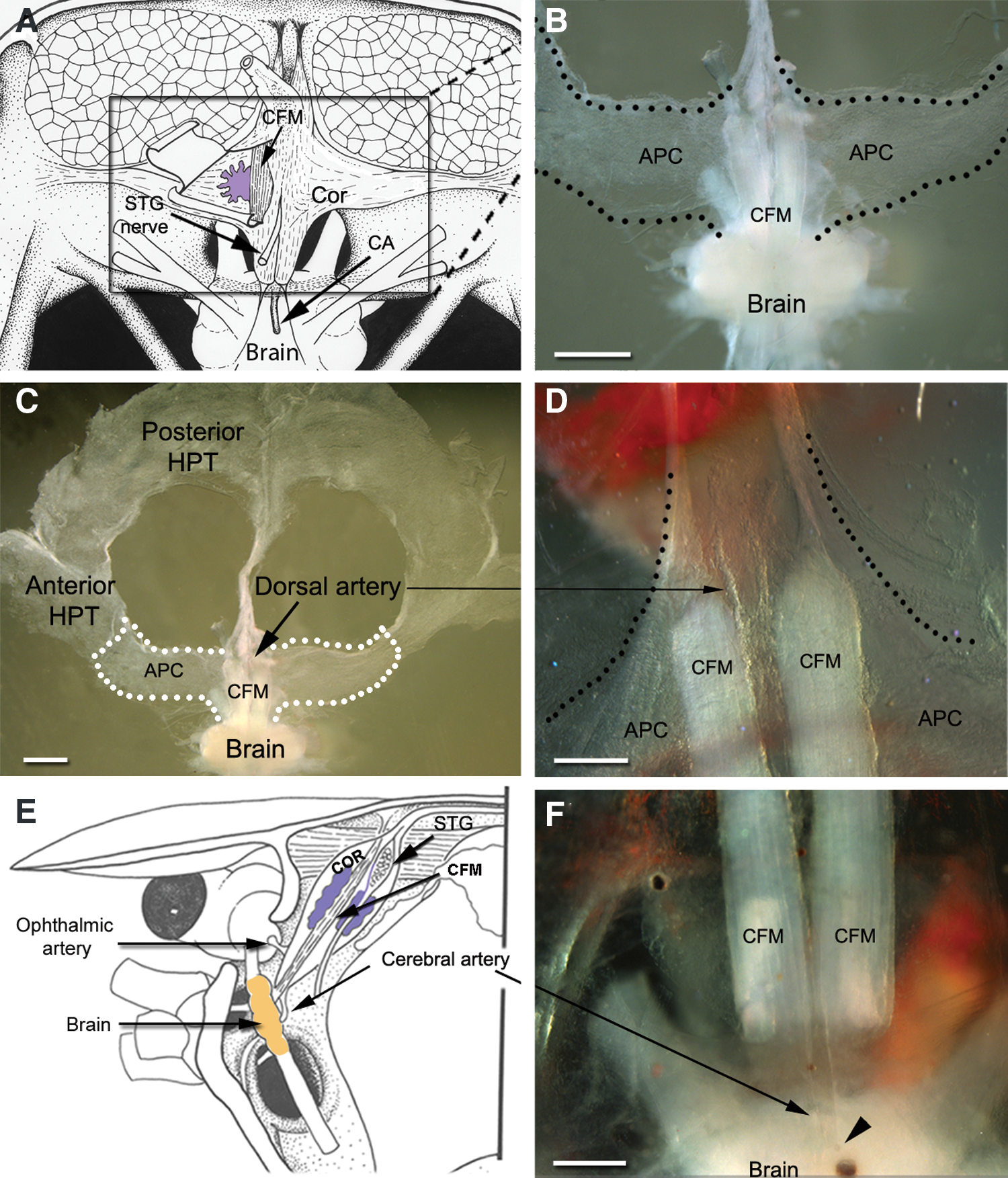
Localization of APC tissues.
The localization of the APC inside the cor frontale is of major interest because there are only three exits from this auxiliary heart: two lateral exits, the ophthalmic arteries, which supply blood to the optic neuropils contained in the eyestalks (Fig. 5E); caudally, a single median vessel, the cerebral artery, supplies blood to the brain (Fig. 5E, F) [19]. Therefore, all of the blood that reaches the cor frontale, which is most of that pumped from the heart into the DMA, will pass through the neural tissues in the optic and brain neuropils; moreover, it is important to recognize that the cerebral artery provides the entire blood supply to the brain. Significantly, the only regions identified in the crustacean nervous system where neurogenesis continues throughout adult life, are in cell clusters 9 and 10 in the brain [3,17] and in the eyestalks [27]. These relationships will be addressed further in the Discussion section.
Cellular organization of the APC
The APC is different from the rest of the HPT not only due to its distinctive location, but also in its cellular composition and organization. Cells composing the anterior and posterior regions of the HPT are arranged in lobules that contribute to large sheets of tissue, with no obvious cellular pattern. In contrast, APC cells located near the CFMs (regions A and B, Fig. 6A) are organized into rosette-like configurations; very few rosettes are found in region C, where most cells are organized in sheets without any obvious pattern. However, cells in all three regions are equally labeled with BrdU. Our observations suggest that the majority of cells in regions A and B participate in rosettes, although these are in different orientations in the tissue and therefore all cannot be visualized in confocal stacks viewed in a single plane. All members of the rosettes can be of similar size (8–12 μm), although sometimes, the central cell within the rosette is noticeably larger than those forming the outer circle. BrdU labeling suggests that the cells composing the rosettes may have a lineage relationship, because short-term (6 h) exposure to BrdU generally labels only the central cell in the rosette (Fig. 7A, B) while a 72-h exposure tends to label all members of the rosettes (Fig. 7E, F), even though BrdU would have cleared from the system by 30–48 h. This temporal pattern of labeling suggests that the surrounding cells could be progeny, labeled during the S-phase of the central mother cell. However, it also is clear that the outer cells can undergo unrelated division, as they are occasionally labeled with BrdU, while the central cell is not.
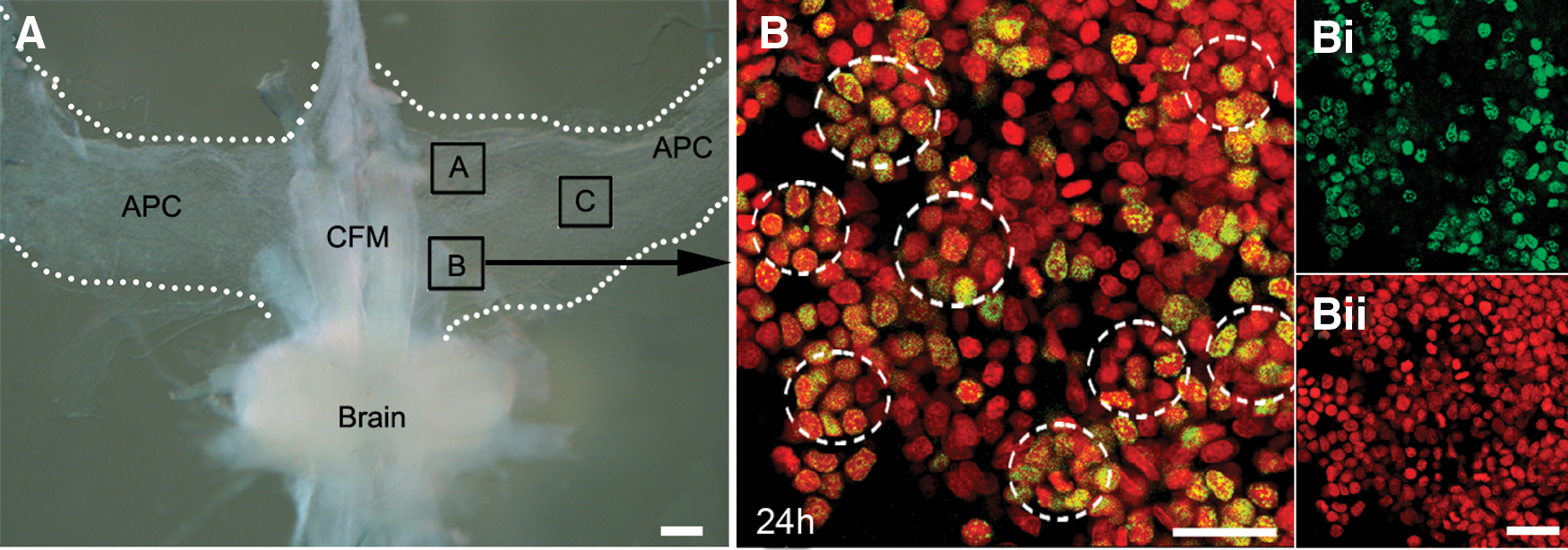
APC cells are arranged in rosettes near the midline and CFMs.

Temporal studies of BrdU labeling within rosettes (dashed lines) suggest that cells have a lineage relationship.
Immunocytochemical studies of the posterior HPT and the APC were conducted with antibodies against β-tubulin and tyrosinated tubulin, providing some points of morphological comparison between these tissues. Microtubules are primarily formed from α- and β-tubulin, the most common members of the tubulin family. Tubulins also are subjected to post-translational modifications, one of which involves the cyclic removal of the carboxy-terminal tyrosine of α-tubulin by a carboxypeptidase and the re-addition of a tyrosine residue by the tubulin–tyrosine ligase [28].
The most notable aspects of β-tubulin labeling are the observations that β-tubulin labels a scaffold or cage encircling rosette complexes, and that rosettes are frequently tethered to each other via β-tubulin immunoreactive fibrils (Fig. 8A, B). Dividing cells in the rosettes often have fibrous extracellular extensions connecting mitotic profiles in neighboring rosettes to one another (Fig. 8A, Ai). β-Tubulin immunoreactivity in the posterior HPT also reveals fibrous elements that form a web within which the HPCs are nestled (Fig. 8C, D). Cells in the process of dividing are intensely immunoreactive for β-tubulin (Fig. 8C).
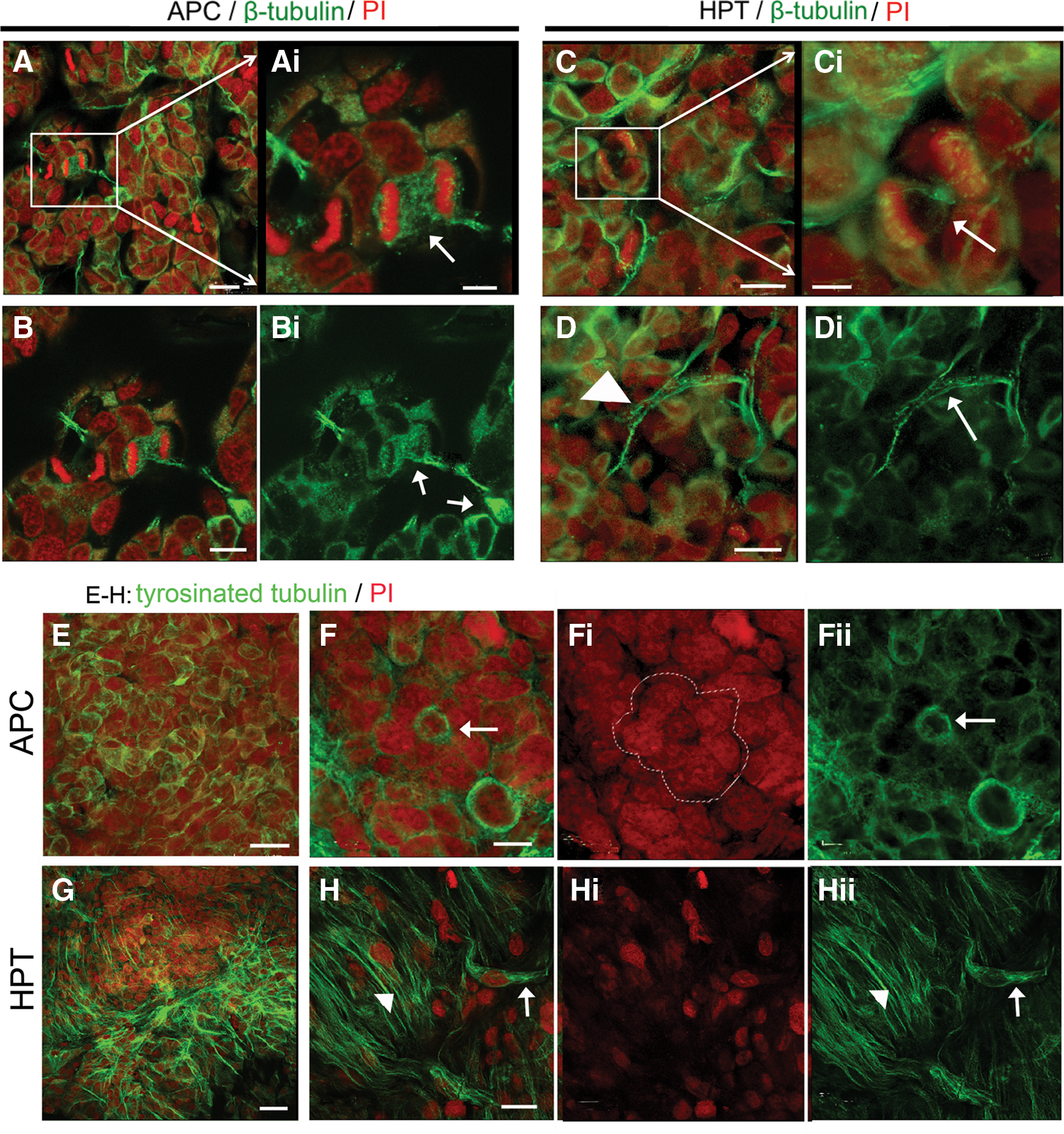
Comparison of the distributions of β-tubulin and tyrosinated-tubulin immunoreactivities in APC tissue and HPT.
Antibodies generated against tyrosinated tubulin label very different elements in the APC and posterior HPT than those immunoreactive for β-tubulin. In the APC, tyrosinated tubulin immunoreactivity is found in the cytoplasm of cells composing the rosettes (Fig. 8F). Other cells that are not obviously contributing to a rosette also are encircled by tyrosinated tubulin labeling (Fig. 8E). In contrast, tyrosinated tubulin immunoreactivity is found in fibrous elements in the posterior HPT (Fig. 8G, H), as well as in the cytoplasm of what appear to be bipolar cells (Fig. 8H, arrow). Fibers closely resembling these label the early protoniche during development, at the time of hatching [7].
Access to the neurogenic niche
The DMA posterior to the cor frontale and the cerebral artery emerging from the cor frontale were examined in BrdU- and DAPI-labeled preparations to observe the prevalence of S-phase cells found within these structures (Fig. 9). A few elongated, BrdU-labeled cells 8–15 μm in their largest dimension were found in the DMA in the region just posterior to where the artery expands to form the cor frontale (Fig. 9, regions 2 and 3). Because of their position and size, these are likely to be hemocytes that were labeled during their final division in the HPT, and which were then released into the circulation. Hemocytes do not divide after they are released from the HPT [29]. Many more BrdU-labeled cells of various sizes also are observed in the area between the CFMs, in the region where the cerebral artery that connects the cor frontale with the brain emerges (Fig. 9; regions 4a–d). This provides evidence that cells emerging from the APC may contribute to the cellular population that flows from the cor frontale into the brain.
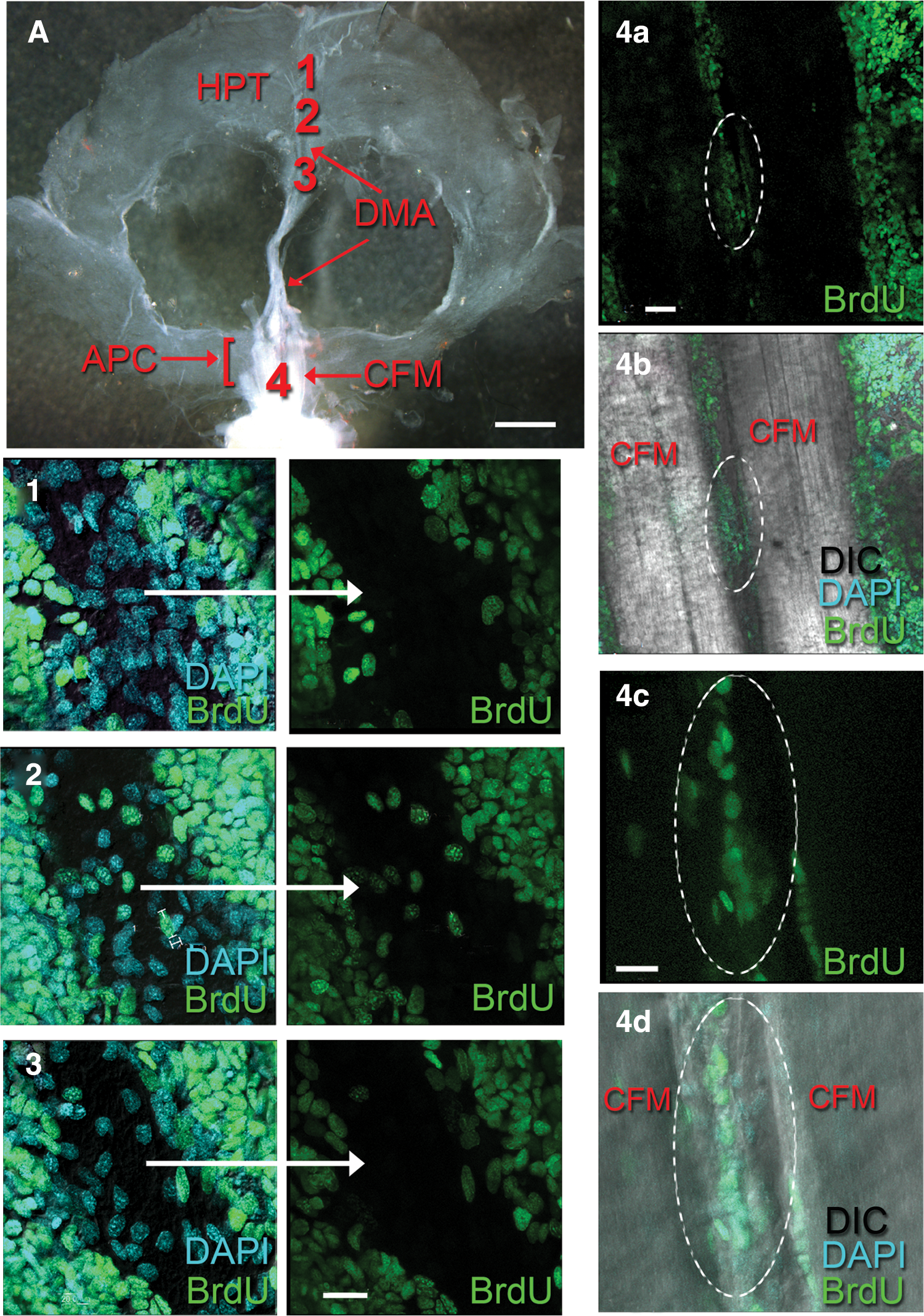
Regions along the DMA were examined following a 24-h BrdU incubation.
The cerebral artery emerges from the cor frontale and enters the brain through the dorsal surface (Fig. 5F), where it extends ventrally into two large lateral branches that ramify in the olfactory lobes, accessory lobes, antenna II neuropils, and other smaller neuropil areas (Fig. 10A–C). Afferent vessels project directly from the lateral branches and can be traced to the bilateral neurogenic niches and streams lying on the ventral surface of the accessory lobes (Fig. 10C, D). It is clear from confocal sections through the neurogenic niche that fine capillaries infiltrate the niche and several converge on the vascular cavity, where these appear to terminate (Fig. 10D). Semithin sections through the neurogenic niche also show a distinctive cell type with a prominent nucleolus and dispersed chromatin concentrated around the edge of the nucleus in high numbers in connective tissue lying between the niche and accessory lobe (Fig. 10E). Cells with these same characteristics are observed in blood vessels that penetrate APC tissues (Fig. 10F).
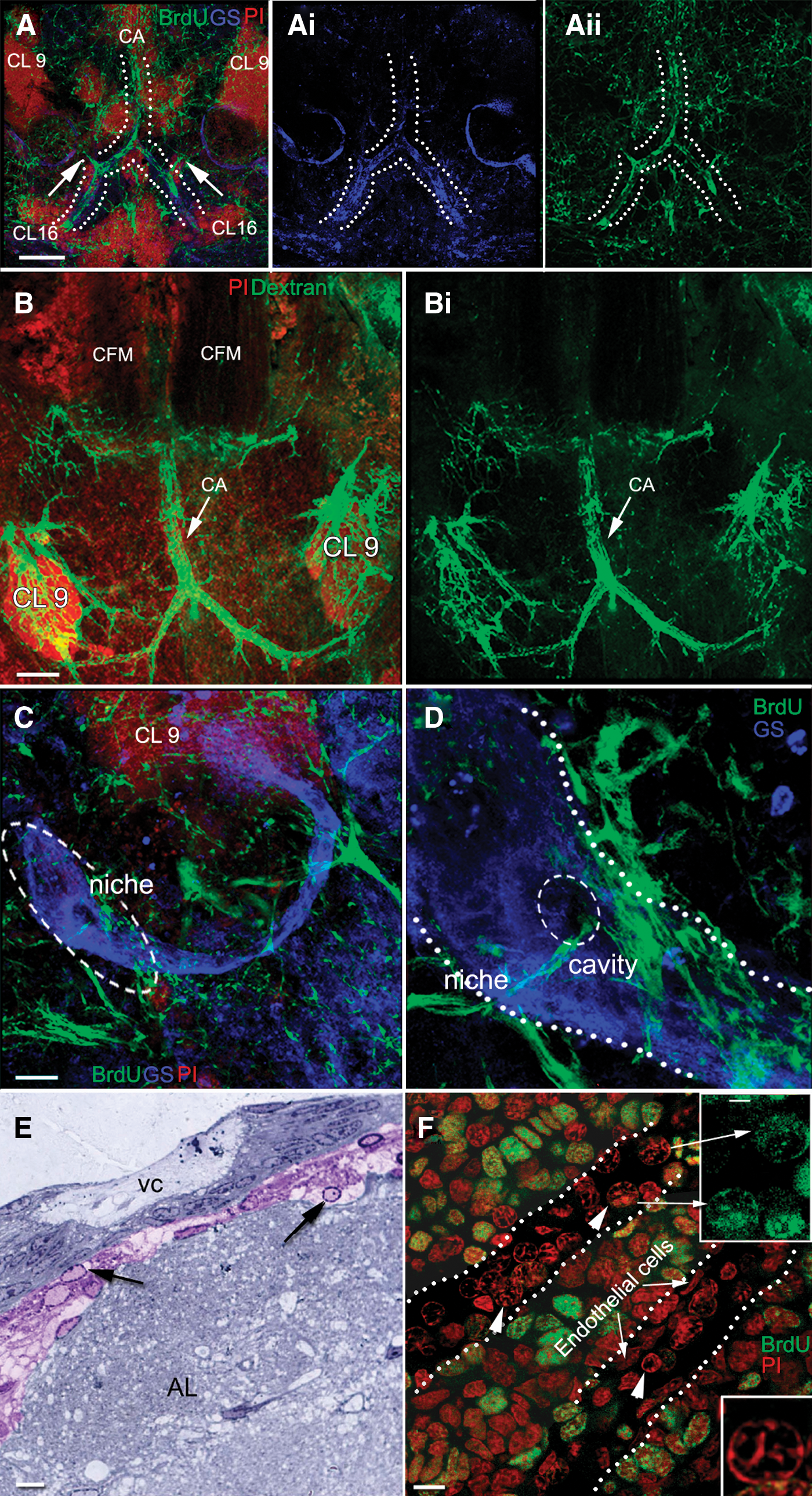
Dextran labeling of the brain vasculature. The brains used in this experiment were not desheathed to examine vascular connections that might be disrupted by removal of the outer membranes enclosing the brain.
Discussion
The HPTs in the crayfish P. clarkii are organized similarly to other decapod crustaceans [22 –24], with the major hematopoietic organ located in lobules attached to a thin layer of connective tissue on the dorsal and dorsolateral surfaces of the stomach and surrounding the DMA. The recent work of Noonin et al. [9] in P. leniusculus shows, however, that this long-recognized hematopoietic organ is only one part of a much more extensive hematopoietic system. Their work confirms that the lobular tissues composing the posterior HPT are responsible for hemocyte production, as previously demonstrated in several crustacean species [23,24,30]. However, the APC, which they discovered lying near the brain, appears to be a region where multipotent stem cells of the HPT are concentrated. They propose this based on several lines of evidence, including the high level of proliferation in the APC compared with the posterior HPT and loose euchromatin in cells of the APC relative to the more highly condensed chromatin in cells of the anterior and posterior HPT. Unlike cells of the posterior HPT that form monolayers in culture, divide sporadically and differentiate into granular and semigranular precursor cells in vitro after induction [26,31], cells of the APC when cultured divide and form densely packed cell clusters after 3–7 days, some forming bowl-shaped formations. However, after about 2 weeks in culture, these APC-derived clusters stopped growing and did not differentiate, even when cytokines (astakine1 and 2) that influence posterior HPT cell differentiation were added. Noonin et al. [9] have proposed that the location of the APC near the brain may facilitate physiological communication between this tissue and the brain, for example, perhaps by providing a source of neuronal precursor cells that will generate adult-born neurons. Because our primary interest is in adult neurogenesis in the crustacean brain, we therefore pursued this idea and have localized and examined the components of the hematopoietic system in the model we utilize, the crayfish P. clarkii.
The studies presented here demonstrate that all three regions of HPT described by Noonin et al. [9] are readily identifiable in P. clarkii: the posterior HPT, anterior HPT, and the APC. As in P. leniusculus, APC cells in P. clarkii have far more proliferative activity relative to the posterior HPT. High ROS production and intense mitochondrial labeling are found in cells throughout the APC. MitoTracker® labeling was localized at the lateral borders of the posterior HPT, as was BrdU, and in cells within the DMA. We also have discovered that the APC in P. clarkii is located within the cor frontale, which vascularizes the brain via the cerebral artery and the eyes via the ophthalmic arteries. Further, we show that APC cells can gain access to the neurogenic niche via capillary branches from the cerebral artery. These vessels have not been noted in earlier studies because we generally remove the sheath covering the brain after fixation and before immunocytochemistry, to facilitate antibody penetration. However, when the ventral sheath is left intact, a series of superficial vessels that infiltrate the niche are revealed (e.g., Fig. 10C, D).
The presence of such proliferative tissues within the cor frontale is of considerable interest because precursor cells released from this tissue would pass immediately to the brain and eyestalk neuropils, both of which contain regions that have the capacity for life-long neurogenesis [17,27]. As suggested by Noonin et al. [9], the APC may indeed be a source for multipotent stem cells which, in the case of adult neurogenesis in the brain of P. clarkii, must come from a source extrinsic to the neurogenic niche.
The cor frontale in P. clarkii is similar in location and morphology to that found in related crustacean species (e.g., the spiny lobster [Panulirus interruptus] and crabs [Callinectes sapidus, Scylla serrata, Cancer productus, and Cancer antennarius]) [19]. The cor frontale also contains the stomatogastric ganglion, which is located between the two CFMs. These features of the auxiliary heart are highly uniform among decapod crustacean species, and thus, we suspect that the presence of the APC within this structure is also likely to be a consistent finding.
The cells of the APC are distinctive from cells in the rest of the HPT in that APC cells are distributed in multilayered sheets rather than in lobules, and these are attached to the muscles deep within the cor frontale as well as to the dorsal and ventral membranes that form the cor frontale. In addition, many APC cells are organized in rosette-like structures; these formations were not observed in either the anterior or posterior HPT. The generation of these structures in vivo may correspond to the behavior of APC cells in culture described above, where bowl-shaped cell clusters form.
The APC rosettes are of particular interest because these types of cellular structures have been associated with early stages of neural development and the formation of epithelial tissues in both vertebrates and invertebrates. During neural differentiation, human and mouse embryonic stem cells go through morphogenic events that result in the formation of radially organized columnar epithelial cells called neural rosettes. Cells in these structures label with a variety of neuroectodermal markers and are believed to represent early-stage neural stem cells that have the capacity to form many cell types in the developing brain. Rosettes also form in vitro from embryonic stem cells. The formation of rosettes during gastrulation in Drosophila has been related to the establishment of polarity in epithelial cells [32], while rosette structures in zebrafish give rise to neuromasts of the lateral line [33]. In these examples, rosettes are typically organized around zones of cell–cell contact where apical ends undergo a coordinated constriction, such as in the early formation of the neural tube. Mammalian embryonic neural stem cells in culture form rosettes [13], and a similar pinwheel structure is associated with the subventricular zone in mice that is responsible for adult neurogenesis of olfactory interneurons [15]. Elkabetz et al. [14] suggest that neural rosettes derived from human embryonic stem cells (R-NSCs) may represent the first neural cell type capable of recreating the full cellular diversity of the mammalian nervous system, thus implying that these assemblies of cells represent stem cells with a broader potential than traditional neural stem cells, whose lineage is generally restricted to specific types of neuronal cells. Others claim that the rosettes are reminiscent of early neural tubes, and indeed they do express markers consistent with neural plate identity, such as Pax6, nestin, NCAM, and Sox1 [34].
In the current study, β-tubulin labeling in the APC reveals a scaffold associated with the rosettes as well as tethers between the rosettes. Microtubules of the eukaryotic cytoskeleton are composed of a heterodimer of α- and β-tubulin, each in itself a family of several highly homologous isoforms [35]. In vertebrates, six types of β-tubulin (class I to class VI) have been identified and these are thought to have functional and cell-type specificities [36]. For example, class III β-tubulin (also known as TuJ-1), one of the most specialized tubulins, is specific for neurons and committed neuronal precursors in birds and mammals, while there is evidence that class I β-tubulin is associated with neuronal commitment and adult neurogenesis in fish [36]. In mammals, the expression of the class III β-tubulin isotype has also been associated with rosette-type cells [37 –39], suggesting that these cells are committed to a neural fate.
Among invertebrates, four β-tubulin isoforms have been identified in Drosophila [40,41], and three to five in crustaceans [42]. The Drosophila β3-tubulin gene has a structure extremely similar to that of the vertebrate β-tubulin genes [40]; however, the distribution patterns of the various isoforms have not linked a specific type with a neural fate [41].
The β-tubulin antibody used in the current experiments does not distinguish between different β-tubulin isotypes, and because so little is known about the distribution and function of various tubulins in crustacean tissues, defining the tubulin isotype will not clarify issues related to cellular commitment or fate. In the present study, therefore, β-tubulin labeling allowed an analysis and comparison of general morphological features of the posterior HPT and APC, but does not support functional conclusions. Nevertheless, the intriguing β-tubulin labeling associated with APC rosettes suggests a prominent role for these molecules in the organization of these assemblies of cells.
An antibody generated against tyrosinated tubulin also was used to label APC and posterior HPT. Tyrosination is a post-translational modification of the tubulin that leads to the presence of tyrosinated tubulin at the dynamic plus-ends of microtubules, while older and more stable microtubules are mainly composed of detyrosinated tubulin (glutamylated-tubulin) [43,44]. Antibodies against tyrosinated tubulin have therefore been particularly useful in the developing nervous system where cytoskeletal reorganization is rampant and, indeed, these antibodies have been employed to reveal the crayfish neurogenic niche and streams as these emerge around the time of hatching [7]. In the current work, tyrosinated tubulin immunoreactivity is found in the cytoplasm of APC cells, perhaps reflecting the dynamic cytoskeletal rearrangements associated with the high degree of mitotic activity in which these cells are engaged. In contrast, tyrosinated tubulin in the posterior HPT is found in fibrous elements resembling those associated with the early protoniche during development [7]. This labeling may suggest that these fibers, like the growth cone, are involved in dynamic shape changes or are active extensions of HPCs.
At this stage in our work, the significance of rosettes in the APC is not obvious. Nevertheless, the “bowl-shaped cell clusters” of Noonin et al. [9] that arise from APC cells in vitro are remarkably similar to the rosettes formed by cultured human embryonic stem cells (e.g., compare Fig. 10 in Ref. [9] and Fig. 4 in Ref. [13]), indeed suggesting that rosette formation in culture could be indicative of a multipotent stem cell type. The common feature in the many systems where rosettes have been identified is that these structures represent an early developmental stage that is generally associated with a neuroectodermal fate. A meaningful interpretation of the APC rosettes awaits immunocytochemical, molecular biological, and fate studies of these intriguing cell assemblies.
Footnotes
Acknowledgments
The authors thank P. Carey and V. LePage for care of the animals used in these studies, and Melanie Prasol for critical comments on the article. The studies reviewed in this article were supported by NSF IOS 1121345 (to B.S.B.) and Brazilian Financial Agency CAPES (to P.G.C.S.).
Author Disclosure Statement
No competing financial interests exist.