
Abstract
Mesenchymal stromal cells (MSC) are an important component of the bone marrow microenvironment. Notch ligands expressed by MSC are known to play a regulatory role for hematopoietic stem and progenitor cells (HSPC) and in support of bone marrow homeostasis. While the role of Notch signaling in HSPC, their progeny, and MSC has been relatively well studied, little is known about the Notch-independent regulatory impact of Notch ligands on MSC themselves. In the present study, we used genetically engineered bone marrow-derived human MSC to study the function role of Jagged-1 and the Jagged-1 intracellular domain (JICD) with regard to the interaction with HSPC. We demonstrate that Jagged-1 in human MSC undergoes cleavage to produce an intracellular domain that translocates into the nucleus. JICD but not Jagged-1 overexpression was associated with an increased expression of stromal cell-derived factor-1. Short-term co-culture (7 days) of HSPC with JICD-overexpressing MSC, but not with Jagged-1-overexpressing MSC, led to increased proliferation of CD34+ progenitors. In contrast, long-term co-culture of HSPC with Jagged-1-overexpressing MSC (up to 6 weeks) led to a significantly better support of cobblestone area-forming cells and long-term culture-initiating cells (LTC-ICs) compared with JICD-overexpressing MSC. Taken together, results of this study indicate that full-length Jagged-1 and JICD have differential effects on MSC and on their interaction with HSPC ex-vivo. JICD-overexpressing MSC induce proliferation of HSPCs in short-term culture at the expense of immature precursors (LTC-ICs), whereas Jagged-1 overexpressing MSC favor LTC-IC formation.
Introduction
M
MSC and osteoblasts expressing Notch ligands are able to activate Notch signaling in HSPC [6]. In-vivo and ex-vivo studies have reported that HSPC with activated Notch signaling maintain an increased capacity for self-renewal [1,7 –9]. In addition, MSC and osteoblasts express Notch receptors that make them sensitive to activation of Notch signaling [10]. Notch has been shown to be involved in the differentiation of embryonic cells into mesenchymal progenitors [11]. Furthermore, Notch signaling participates in the regulation of osteoblastogenesis. While some studies have shown that Notch activation stimulates osteogenic differentiation potential [12,13], other reports have found the opposite effect, suggesting Notch-mediated suppression of osteogenic potential [14,15]. Thus, although the precise mechanisms remain to be elucidated, Notch signaling plays an essential role in bone marrow homeostasis.
While the roles of Notch signaling in HSPC and their progeny, as well as in MSC and osteoblasts, have been relatively well studied, little is known about the signaling cascades specifically activated by Notch ligands independent of classical Notch signaling. It has been reported that Notch ligands undergo cleavage in some cell lines, with the formation of intracellular domains (ICDs) that translocate into the nucleus and possess the potential to mediate nuclear signaling events [16 –18]. Based on these previous reports, we hypothesized that Jagged-1 may play an independent regulatory role in MSC and can influence HSPC/MSC interactions, in addition to its function as Notch ligand. Therefore, we overexpressed full-length Jagged-1 and the Jagged-1 intracellular domain (JICD) in human bone marrow-derived MSC to investigate whether Jagged-1 or JICD alone influence MSC fate independent of Notch and whether overexpression of full-length Jagged-1 or JICD has the same effect on HSPC/MSC interactions ex-vivo. In addition, we knocked down Jagged-1 using shRNA to further elucidate its role in human primary MSC.
Materials and Methods
Isolation of CD34+ cells from leukapheresis samples
CD34+ cells were isolated from leukapheresis samples of healthy donors treated with 7.5 μg/kg granulocyte-stimulating factor subcutaneously per day for 5 subsequent days. Informed consent was obtained in accordance with research protocol approved by the local institutional review board. HSPC were isolated using CD34 antibody-conjugated magnetic beads, according to the manufacturer's instructions (Miltenyi Biotec, Bergisch Gladbach, Germany). The purity of the isolated CD34+ population was confirmed to be >99% by flow cytometry.
Isolation and culture of primary human MSC
Primary human bone marrow-derived MSC were collected from bone marrow aspirates from healthy donors after informed consent and approval by the local ethics committee. Mononuclear cells were collected using a Ficoll gradient and cultured at 37°C and 5% CO2 in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal calf serum (FCS; Biochrom, Cambridge, UK). After 24 h, nonadherent cells were removed, and adherent cells were expanded. The immunophenotype of all primary MSC was analyzed by flow cytometry (CD 166+, CD105+, CD90+, CD73+, CD34−, and CD45−). Primary MSC from passages 2–5 were used for the experiments.
Culturing of Scp-1 (immortalized human bone marrow-derived MSC), HeLa, HEK293, and HEK293T cells
Single-cell-picked clone 1 immortalized human bone marrow-derived MSC overexpressing the human telomerase reverse transcriptase (Scp-1 cells) were kindly provided by Matthias Schieker, München, Germany [19] and HeLa, HEK293, and HEK293T cells were cultured in DMEM supplemented with 10% FCS at 37°C and 5% CO2.
Short-term co-culture of CD34+ cells with primary MSC
Transduced primary MSC were plated at a density 1–2×104/cm2 and cultured in DMEM with 10% FCS until confluence. Freshly isolated CD34+ cells were plated on the confluent layer of primary MSC in CellGro stem cell growth medium (CellGenix, Freiberg, Germany) supplemented with 10% FCS, 10 ng/mL stem cell factor (Biosource, Camarillo, CA), 10 ng/mL FLT3-L (Miltenyi Biotec), and 10 ng/mL IL-3 (Biosource) at a density of 1×104/cm2. CD34+ cells under stroma-free conditions (plasma-treated polystyrene tissue plastic, PTP) were plated at the same density in the same medium. After 1 week, the co-culture supernatant was gently mixed, and the nonadherent HSPC fraction was collected. The MSC layers were washed twice with phosphate-buffered saline (PBS) to collect the remaining nonadherent cells. The MSC layers with attached HSPC, including HSPC beneath MSC layers (adherent HSPC fraction), were trypsinized. Nonadherent cells were treated with trypsin in the same manner. Cells were counted using a hemocytometer. The proportions of HSPC were quantified by flow cytometry (CD45+).
Clonogenic progenitor assay
After 1 week of short-term co-culture, mixtures of HSPC and primary MSC were harvested, counted using a hemocytometer, and plated (1×103 cells per mL) in triplicate in methylcellulose medium containing recombinant cytokines (MethoCult H4435; Stem Cell Technology, Vancouver, BC, Canada). Mixtures of HSPC and primary MSC were stained with CD45-APC antibody for flow cytometry analysis to determine the proportions of HSPC. After 2 weeks, colony-forming cells (CFC) were counted using an inverted microscope with dark-field illumination, and CFC frequencies were calculated according to the number of CD45+ cells.
Cobblestone area-forming cell and long-term culture-initiating cell assays
Freshly isolated CD34+ cells were seeded on confluent layers of transduced primary MSC and maintained for 6 weeks in MyeloCult H5100 medium (Stem Cell Technology) supplemented with 10−6 M hydrocortisone (Sigma-Aldrich, St. Louis, MO). Half-media exchanges and cobblestone area scoring were performed weekly. Cobblestone areas were counted using an inverted microscope (Axiovert 25; Carl Zeiss, Jena, Germany) and counted as colonies when at least 10 cells formed a colony beneath the stromal layer. After 6 weeks, mixtures of HSPC and primary MSC were harvested, and subsets were counted using a hemocytometer and stained with CD45-APC antibody for flow cytometry analysis to determine the proportions of HSPC. Cells (1×104) were plated and cultured for 2 weeks in enriched methylcellulose medium with recombinant cytokines (MethoCult H4435; Stem Cell Technology). After 2 weeks, secondary CFC were counted using an inverted microscope with dark-field illumination, and CFC frequencies were calculated according to the number of CD45+ cells.
Osteogenic and adipogenic differentiation
Transduced primary MSC were plated at a density of 1×104/cm2 and cultured in DMEM with 10% FCS until cells reached 70% confluence. To induce osteogenic differentiation, the growth media were changed to DMEM 10% FCS supplemented with 200 μM ascorbic acid, 10 mM β-glycerophosphate, and 100 nM dexamethason. Alkaline phosphatase (ALP) activity was measured after 14 days in culture with osteogenic medium. Briefly, cells were washed with PBS and lysed in 1.5 M Tris, pH 10, supplemented with 1 mM ZnCl2, 1 mM MgCl2, and 1% Triton X-100. Lysates were centrifuged, and supernatants were incubated with 3.7 mM 4-nitrophenylphosphate for 30 min at 37°C. The reaction was stopped with 100 mM NaOH, and released 4-nitrophenolate was measured photometrically at 405 nm. Catalytic activity of ALP was calculated using a p-nitrophenolate calibration curve and protein concentration of the lysates. Mineralization was evaluated with von Kossa staining after 21 days in culture with osteogenic medium. Briefly, cells were fixed with 10% formalin, stained with 2% silver nitrate solution, and developed using 0.2% pyrogallol. To induce adipogenic differentiation, the growth media were changed to DMEM 10% FCS supplemented with 1 μM dexamethason, 500 μM IBMX, 1 μg/mL insulin, and 100 μM indometacin. Adipocyte formation was evaluated with Oil Red staining after 21 days in culture with adipogenic medium. To evaluate the expression of late markers of adipocyte differentiation (aP2 and LPL) after 21 days in culture with adipogenic medium, cells were washed and proceeded to mRNA extraction as described next.
Proliferation assays
Transduced primary MSC were plated at a density of 2×104/cm2 and cultured in DMEM with 10% FCS. After 72 h, EdU (Life Technologies, Carlsbad, CA) was added to cell culture medium at a final concentration of 0.1 μM. After 72 h, cells were trypsinized, fixed, permeabilized, and stained with Alexa Fluor azid antibody (Life Technologies) according to the manufacturer's instructions to quantify EdU incorporation by flow cytometry. For 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) proliferation assay, transduced primary MSC were plated at a density of 2×104/cm2 in 96-well plates in quadruplicates. After 3 days, MTT (Roche, Mannheim, Germany) was added to cell culture medium at a final concentration of 0.5 mg/mL. After 4 h MTT, solubilization solution (Roche) was added, left overnight, and absorbance was measured at 570 nm with a reference filter of 650 nm.
Lentiviral plasmid generation, virus particle production, and transduction
Jagged-1, JICD, and dnMAML lentiviral transfer vectors were generated as previously described [12]. Briefly, vectors were created with open reading frames of human Jagged-1, JICD (from aa1094 to aa1218), and dominant negative mastermind 1 (dnMAML1). The transgenes were inserted into the pRRL.SIN.cPPT.SFFV.GFP.WPRE vector [20] (kindly provided by Christopher Baum, Hannover, Germany) to replace green fluorescent protein. For the Jagged-1 and JICD vectors, an IRES-GFP sequence was added downstream of the transgenes. dnMAML1 was cloned as a C-terminal fusion with the mCherry reporter protein. pRRL.SIN.cPPT.SFFV.IRES.GFP.WPRE vector was used for control transduction (for schematics of the vectors, refer to Fig. 1A).
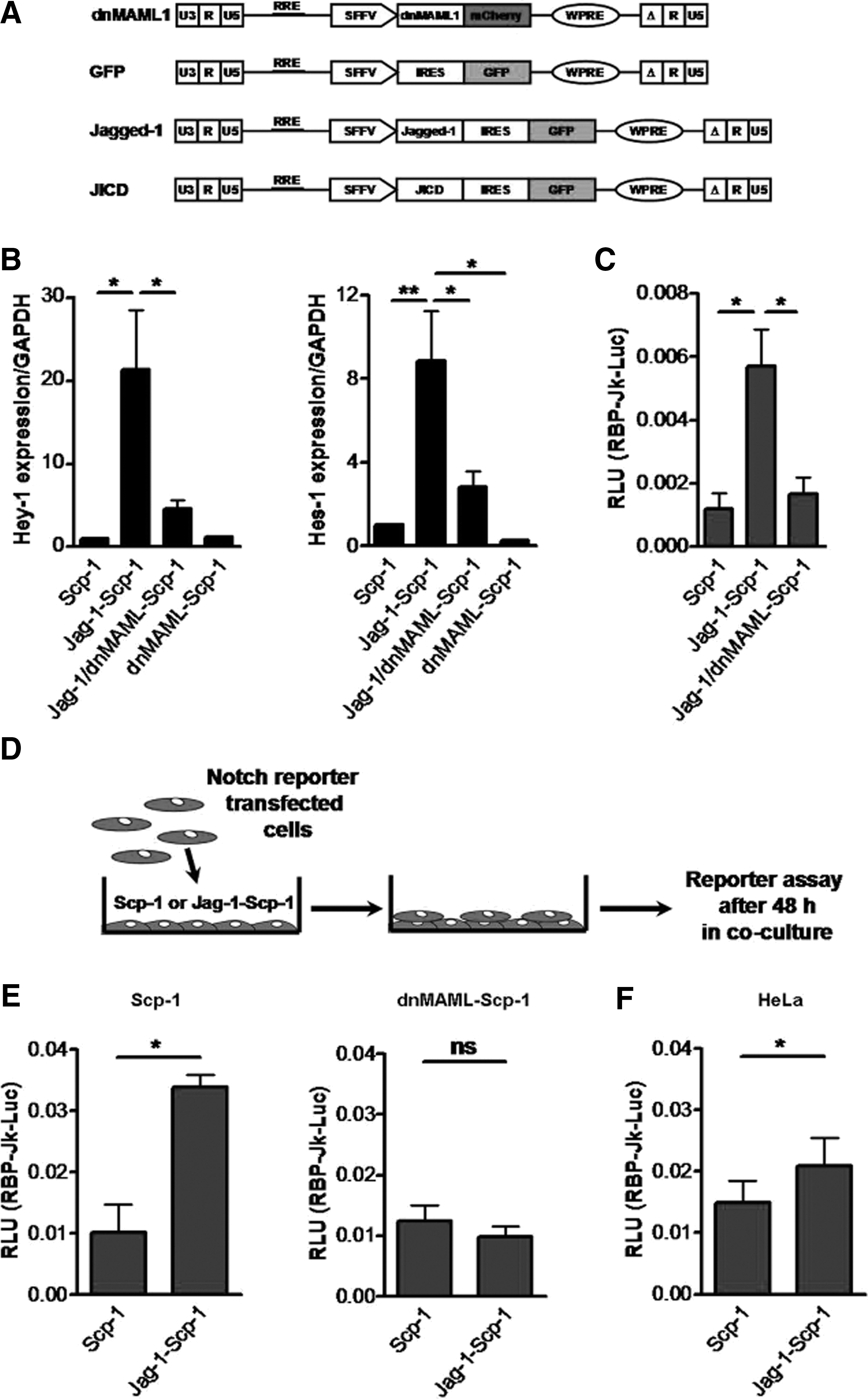
Overexpression of Jagged-1 in MSC is followed by activation of the Notch pathway in neighboring cells.
Not-target shRNA control pLKO.1-puro lentiviral transfer vector (scramble) and Jagged-1 small hairpin RNA (CCCGAATGTAACAGAGCTATT) pLKO.1-puro lentiviral transfer vector (Jag-1-shRNA) were kindly provided by Frank Buchholz, Dresden, Germany.
To produce lentiviral vector particles, HEK293T cells were transfected with lentiviral vectors in combination with the packaging plasmids psPAX and pVSVg using polyethylenimine [20,21]. Virus titers were determined for each batch of virus vector particles in order to use comparable titers for the transductions. Transductions were performed overnight at 37°C with virus supernatants supplemented with 1 μg/mL protamine. Transduction efficiency of GFP, Jagged-1, JICD, and dnMAML lentiviral vectors was evaluated by flow cytometry analysis using FACSCalibur or FACSAria II (BD, Franklin Lakes, NJ). For nomenclature of cells transduced with different lentiviral vectors, refer to Table 1. Cells transduced with scramble and Jag-1-shRNA lentiviral vectors were selected using 0.75 μg/mL puromycin for 1 week.
Scp-1, single-cell-picked clone 1 immortalized human bone marrow-derived MSC overexpressing the human telomerase reverse transcriptase; MSC, mesenchymal stromal cells; dnMAML, dominant-negative Mastermind 1; GFP, green fluorescent protein.
Stromal cell-derived factor-1 reporter plasmid generation and Gaussia luciferase and secreted ALP reporter assays
For construction of the stromal cell-derived factor-1 (SDF-1) reporter plasmid, a bacterial artificial chromosome containing the human SDF-1 promoter (RP11-691N23) was purchased from BACPAC Resources (Oakland, CA), and a cassette containing the Gaussia luciferase gene was inserted directly after the transcription start site of the SDF-1 promoter region. Next, the fragment containing 1.4 kb of the SDF-1 promoter region and the Gaussia luciferase cassette was subcloned into the XhoI/SalI cloning sites of the lentiviral transfer vector pRRL.SIN.cPPT.SFFV.GFP.WPRE. As an internal control and to normalize SDF-1 promoter activity, a fragment containing secreted ALP (SEAP) under control of the SV40 promoter was amplified by PCR and subcloned into the SDF-1 reporter plasmid (for a schematic of SDF-1 reporter vector, refer to Fig. 4B).
For SDF-1 reporter experiments, HEK293 cells were transiently transfected with the lentiviral dnMAML1-encoding vector. After 24 h, these cells were transiently co-transfected with GFP-, Jagged-1-, or JICD-encoding lentiviral vector (GFP/dnMAML-, Jag-1/dnMAML-, and JICD/dnMAML-HEK293 cells, respectively) along with the SDF-1 reporter plasmid. After 48 h, the supernatants were collected and assayed for Gaussia luciferase and SEAP activities. Gaussia luciferase activity was measured using a BioLux Gaussia Luciferase Assay Kit (New England Biolabs, Ipswich, MA) according to the manufacturer's instructions. SEAP activity was measured using a NovaBright™ Phospha-Light™ EXP Assay Kit (Life Technologies) according to the manufacturer's instructions. Gaussia luciferase activity was normalized to SEAP activity. Each experiment was performed in triplicate. For nomenclature of HEK293 cells transiently transfected with different lentiviral vectors, refer to Table 1.
Notch-signaling reporter assays
RBP-Jκ is a key mediator of Notch signaling [4]. Therefore, to quantify Notch activity, we transiently co-transfected Scp-1, dnMAML-Scp-1, HeLa, and HEK293 cells with a plasmid containing RBP-Jκ transcriptional response elements upstream of the firefly luciferase gene and a plasmid constitutively expressing the Renilla luciferase (SABiosciences, Hilden, Germany), using Lipofectamine 2000 (Life Technologies).
Scp-1 cells were lysed 48 h after transfection, and luciferase activity was determined using the Dual-Luciferase Reporter Assay System (Promega, Madison, WI) according to the manufacturer's instructions.
For co-culture experiments, Scp-1, dnMAML-Scp-1, or HeLa cells were trypsinized 24 h after transfection and plated on confluent layers of untransduced or Jag-1-Scp-1. After 48 h, cells were lysed, and luciferase activity was assayed as described earlier.
HEK293 cells were initially transfected with the dnMAML-encoding plasmid using Lipofectamine 2000 and were then co-transfected with GFP-, Jagged-1-, or JICD-encoding plasmids (GFP/dnMAML-, Jag-1/dnMAML-, and JICD/dnMAML-HEK293 cells, respectively) along with the Notch reporter plasmid. After 48 h, the co-transfected cells were lysed, and luciferase activity was measured as described earlier. Each experiment was performed in triplicate.
Quantitative real-time PCR
Total RNA was isolated using TRIzol reagent (Life Technologies). Single-stranded cDNA was synthesized with oligo-dT primers using a RevertAid First-Strand cDNA Synthesis Kit (Thermo Scientific, Franklin, MA) according to the manufacturer's instructions. Quantitative real-time PCR (qRT-PCR) was performed using a SYBR Green PCR Master Mix (Thermo Scientific) on an ABI7500 Fast Real-Time PCR System (Life Technologies). The housekeeping gene GAPDH was used as a reference gene. The gene-specific primer sets are listed in Supplementary Table S1; Supplementary Data are available online at
Western blot analysis
Whole-cell lysates were obtained using M-PER Mammalian Protein Extraction Reagent supplemented with protease inhibitor (Thermo Scientific) according to the manufacturer's instructions. Cytoplasmic and nuclear protein fractions were prepared using NE-PER nuclear and cytoplasmic extraction reagents (Thermo Scientific) according to the manufacturer's instructions. Proteins were subjected to sodium dodecyl sulfate polyacrylamide gel electrophoresis and separated proteins were transferred to nitrocellulose membranes. After transfer, the membranes were blocked and incubated overnight at 4°C with antibodies against the cytoplasmic domain of human Jagged-1 (1:4000; Cell Signaling Technology), human β-actin (1:2000; Sigma-Aldrich), or human GAPDH (1:4000; Cell Signaling Technology). Goat anti-rabbit-HRP antibody (1:5000; Thermo Scientific) was used as the secondary antibody. Western blots were visualized with the ESC Plus western blotting detection system (GE Healthcare, Chalfont St. Giles, UK) on Amersham Hyperfilm (GE Healthcare). GAPDH, as a cytoplasmically localized protein, served as the control for the purity of the cytoplasmic and nuclear fractions.
Enzyme-linked immunosorbent assay
SDF-1α concentrations in GFP/dnMAML-, Jag-1/dnMAML-, JICD/dnMAML-MSC, scramble/dnMAML-, and Jag-1KD/dnMAML-MSC cultures medium were detected using human SDF-1α DuoSet Enzyme-linked immunosorbent assay (ELISA) Development kit (R&D Systems, Minneapolis, MN) according to the manufacturer's instructions.
Immunofluorescence
Transduced primary MSC seeded on cover slips were fixed in 4% paraformaldehyde and then permeabilized with 0.1% Triton X-100. After blocking, the cells were incubated overnight at 4°C with rabbit antibody against the Jagged-1 cytoplasmic domain (1:100; Cell Signaling Technology). Anti-rabbit-Cy3 secondary antibody (1:200; Sigma-Aldrich) was added for 1 h at room temperature, and then, the DNA dye 4',6-diamidino-2-phenylindole (Sigma-Aldrich) was used for nuclear staining. Cell evaluation was performed by confocal microscopy (Zeiss LSM 510; Carl Zeiss, Jena, Germany). 3D reconstruction was done using Velocity×64 software (Perkin Elmer, Waltham, MA).
Flow cytometry analysis and fluorescent-activated cell sorting
Primary MSC transduced with different virus vector particles were analyzed using FACSAria II to evaluate the percentage of cells positive for both GFP and mCherry. HSPC after co-culture experiments were analyzed using FACSCalibur. HSPC harvested after 1 week of co-culture were incubated with CD45-APC and CD34-PE or CD133-PE (1:25; Miltenyi Biotec) at 4°C for 45 min. HSPC were gated according to their CD45 expression. Appropriate murine isotypes served as negative controls.
After 1 week of co-culture, HSPC were harvested as described earlier. The adherent HSPC fraction containing MSC was incubated with CD45-APC and CD166-PE (1:25; Miltenyi Biotec) at 4°C for 45 min. The cells were sorted using FACSAria II, and HSPC were collected based on their CD45+ CD166− GFP− phenotype. Appropriate murine isotypes served as negative controls. Transduced Scp-1 and one of four primary MSC donors were sorted using FACSAria II to obtain the GFP/mCherry double-positive population.
Statistics
All results are reported as means±standard error of the mean. Data were analyzed using a two-sided paired Student's t-test, one-way ANOVA with a Tukey multiple comparison test, or two-way ANOVA with a Bonferoni multiple comparison test, using GraphPad Prism 4.03. Statistical significance was established as *P<0.05, **P<0.01, or ***P<0.001.
Results
Overexpression of Jagged-1 in MSC is followed by activation of the Notch pathway in neighboring cells
Initially, we examined basal expression of all Notch receptors and all Notch ligands in both immortalized human MSC overexpressing the human telomerase reverse transcriptase (Scp-1 cells) and primary human MSC. A similar expression pattern was observed for Scp-1 and primary MSC with predominant expression of Notch-1, Notch-2, and Notch-3 among Notch receptors and Jagged-1 among Notch ligands (Supplementary Fig. S1A, B). To examine whether overexpression of Jagged-1 is followed by stimulation of Notch signaling activation in an autonomous manner, we transduced Scp-1 with Jagged-1 lentivirus-vector particles (Fig. 1A) (Jag-1-Scp-1). As a control, Jag-1-Scp-1 was co-transduced with dnMAML1 lentivirus-vector particles (Fig. 1A) to specifically block Jagged-1-mediated Notch signaling (Jag-1/dnMAML1-Scp-1) [22]. Gene expression analysis of Notch target genes Hey-1 and Hes-1 (Fig. 1B) and the RPB-Jκ-responsive reporter assay (Fig. 1C) showed that overexpression of Jagged-1 was followed by activation of the Notch pathway with a partial block in Jag-1/dnMAML1-Scp-1 [For Hey-1, Jag-1-Scp-1 vs. untransduced Scp-1 showed a 21.3±7.2-fold increase, P<0.05, and Jag-1/dnMAML1-Scp-1 vs. untransduced Scp-1 showed a 4.6±1.0-fold increase, which was not significant (ns). For Hes-1, Jag-1-Scp-1 vs. untransduced Scp-1 showed an 8.8±3.0-fold increase, P<0.05, and Jag-1/dnMAML1-Scp-1 vs. untransduced Scp-1 showed a 2.8±1.0-fold increase, P=ns. An increased activity of Notch reporter could be detected in Jag-1-Scp-1 [5.7±1.0×10−3 relative luciferase units (RLU)] compared with untransduced Scp-1 (1.0±0.5×10−3 RLU, P<0.05) and Jag-1/dnMAML1-Scp-1 (1.7±0.5×10−3 RLU, P<0.05)].
To determine whether dnMAML alone induces Notch signaling, we analyzed expression of Notch target genes in Scp-1 cells overexpressing dnMAML (dnMAML-Scp-1) only. No increase in expression of Hey-1 and Hes-1 was observed in dnMAML-overexpressing Scp-1 (Fig. 1B). To further confirm that dnMAML prevents Jagged-1-mediated Notch signaling activation, Scp-1 and dnMAML-Scp-1 were transiently transfected with the Notch-reporter RBP-Jκ-responsive luciferase constructs and cultured with untransduced or Jagged-1-overexpressing Scp-1 (Fig. 1D). Notch signaling was activated in Scp-1 cultured with Jagged-1-overexpressing cells (Fig. 1E; 3.4±0.2×10−2 RLU with Jag-1-Scp-1 vs. 1.0±0.4×10−2 RLU with untransduced Scp-1, P<0.05), but not in dnMAML-Scp-1 (Fig. 1E; 1.0±0.2×10−2 RLU with Jag-1-Scp-1 vs. 1.2±0.3×10−2 RLU with untransduced Scp-1, P=ns). Thus, these results indicate that dnMAML specifically prevents Notch signaling activation.
To determine whether Jagged-1 can mediate Notch signaling in other cells, we cultured HeLa cells previously transfected with the Notch-reporter RBP-Jκ-responsive luciferase constructs with untransduced or Jag-1-Scp-1. Notch signaling was activated in HeLa cells that were cultured with Jag-1-Scp-1 as compared with those cultured with untransduced Scp-1 (Fig. 1F; 2.0±0.5×10−2 RLU vs. 1.0±0.4×10−2 RLU, respectively, P<0.05).
Jagged-1 is cleaved, and JICD translocates into the nucleus
On binding, Notch ligands can undergo cleavage into several protein fragments [16,17,23,24]. To assess whether full-length Jagged-1 also undergoes cleavage in human MSC, we initially used whole-cell lysates from untransduced Scp-1 and Jag-1-Scp-1 for western blot analysis using an antibody against the cytoplasmic tail of Jagged-1. Three specific fragments were identified in cell lysates from Jag-1-Scp-1, as predicted by previous reports [16,17], to be full-length Jagged-1, the C-terminal fragment (CTF), and JICD (data not shown). We next used primary MSC for lentiviral transduction to overexpress Jagged-1 (Jag-1-MSC). In addition, we transduced primary MSC with a construct encoding the ICD of Jagged-1 only (JICD-MSC; aa: 1094–1218; Fig. 1A). By western blot analysis of the whole-cell lysate from Jag-1-MSC, we identified full-length Jagged-1 (∼150 kDa) and its derivatives, presumably CTF (∼19 kDa) and JICD (∼15 kDa). In JICD-MSC, only one fragment was detected at the same level as the JICD from Jag-1-MSC (Fig. 2A).

Jagged-1 in human MSC is cleaved with formation of Jagged-1 ICD, which translocates into the nucleus.
To evaluate the subcellular localization of full-length Jagged-1 and its derivatives, we stained transduced primary MSC for fluorescent microscopy with an antibody binding to the cytoplasmic domain of Jagged-1. In Jag-1-MSC, we detected Jagged-1 not only in cytoplasmic and membrane fractions, but also in the nuclei, suggesting that this fragment corresponds to cleaved JICD (Fig. 2C and Supplementary Movie S1). JICD-MSC showed predominantly intranuclear localization. These results indicate that JICD translocates into the nucleus of human MSC (Fig. 2C).
Blocking of Jagged-1-mediated Notch activation by co-overexpression of dnMAML1 does not influence Jagged-1 cleavage and JICD translocation
Similar to Scp-1, overexpression of Jagged-1 in primary MSC resulted in Notch-signaling activation. The Hey-1 expression level was increased approximately seven fold in Jag-1-MSC as compared with control-transduced MSC (Fig. 3A; Jag-1-MSC vs. GFP-MSC, 7.4±0.4-fold increase, P<0.001). Since Jagged-1-mediated Notch activation makes it difficult to distinguish between Jagged-1/JICD-associated and Notch-associated effects, we next generated cells co-expressing dnMAML1 along with GFP only, Jagged-1, or JICD (GFP/dnMAML-, Jag-1/dnMAML-, and JICD/dnMAML-MSC, respectively; for schematics of the vectors, refer to Fig. 1A). Transduction efficiency was approximately 70% of co-expressing cells as quantified by mCherry (dnMAML) and GFP (GFP only, Jagged-1, and JICD) expression (Fig. 3B). In co-expressing cells, dnMAML1 blocked the increased expression of Hey-1 to a level similar to that in GFP-MSC (Fig. 3A; Jag-1/dnMAML-MSC vs. GFP-MSC, 1.1±0.2-fold change, P=ns) and down-regulated Hes-1 in all co-expressing cells (P<0.001; Fig. 3A). Only cells co-expressing dnMAML1 were used in subsequent experiments.

Blocking of Jagged-1-mediated Notch activity by co-expression of dnMAMl1 does not influence Jagged-1 cleavage and JICD translocation.
To determine whether co-expression of dnMAML1 protein influences Jagged-1 cleavage and translocation of JICD into the nucleus, we prepared subcellular protein fractions from transduced Scp-1 co-expressing dnMAML along with GFP only, Jagged-1, or JICD (GFP/dnMAML-, Jag-1/dnMAML-, and JICD/dnMAML-Scp-1, respectively) for western blot analysis. Full-length Jagged-1 was identified in the whole-cell lysate and cytoplasmic protein fraction of Jag-1/dnMAML1-Scp-1, but not in the nuclear fraction (Fig. 3C). Both cleaved and recombinant JICD were found in the whole-protein lysate, the cytoplasmic fraction, and the nuclear fraction. These results indicate that both physiologically cleaved and overexpressed JICD translocate into the nucleus independently of dnMAML1 co-expression (Fig. 3C).
JICD promotes SDF-1 expression
It has been reported that ICDs of Notch ligands activate gene reporters and regulate the expression of several genes, especially p21 [25] and connective tissue growth factor [18], and possess the potential to bind to activator protein 1 (AP-1) response elements [17]. Hypothesizing that Jagged-1 and JICD could be involved in the interaction between MSC and HSPC, we screened transduced primary MSC for the expression levels of a range of HSPC-supportive genes containing binding sites for AP-1 in their promoter regions, in particular Angiopoietin-1 [26], Cyclooxygenase-2 (COX-2) [27], and SDF-1 [28]. Angiopoietin-1 and COX-2 showed no change in expression (data not shown). Both SDF-1α and SDF-1β, two isoforms of the chemokine with relevance in the bone marrow niche [29], were up-regulated in JICD/dnMAML1-MSC, but not in Jag-1/dnMAML1-MSC (Fig. 4A). Furthermore, we detected increased secretion of SDF-1α by JICD/dnMAML-MSC in comparison with GFP/dnMAML-MSC as detected by ELISA (Supplementary Fig. S2A), although the protein level of SDF-1α in supernatant from Jag-1/dnMAML-MSC was also higher than from GFP/dnMAML-MSC (Supplementary Fig. S2A).
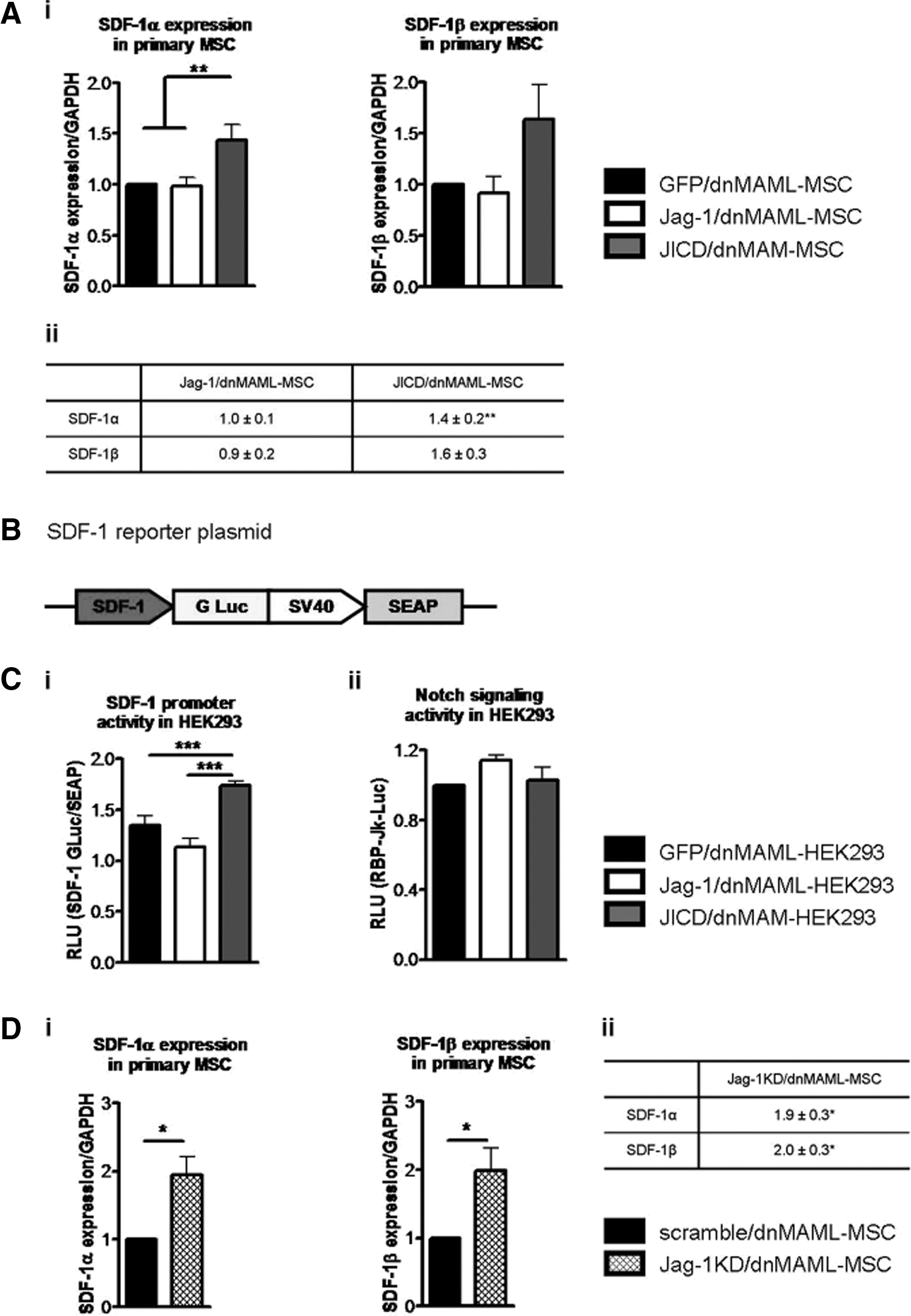
Effects of Jagged-1/JICD overexpression and Jagged-1 KD on SDF-1 expression.
To further address the role of Jagged-1 and JICD in the regulation of SDF-1 expression, we used HEK293 cells for transient co-transfection with GFP only, Jagged-1-, or JICD-encoding constructs and the SDF-1 reporter construct (Fig. 4B) to assess the influence of Jagged-1 and JICD on SDF-1 promoter activity. At basal conditions, HEK293 cells also express both Notch receptors and Notch ligands (Supplementary Fig. S1C). To prevent any Notch-associated effects, HEK293 cells were initially transiently transfected with the plasmid encoding dnMAML1 to prevent Jagged-1-mediated Notch signaling activity and, after 48 h, these cells were co-transfected with GFP-, Jagged-1-, or JICD-encoding constructs (GFP/dnMAML-, Jag-1/dnMAML, and JICD/dnMAML-HEK293 cells, respectively) along with the SDF-1 reporter plasmid. Again, JICD, but not Jagged-1, activated the SDF-1 promoter, as shown by the reporter assay (Fig. 4Ci; P<0.001). Cleavage of transiently overexpressed Jagged-1 with formation of CTF, and JICD was confirmed by western blot analysis of whole-cell lysate from Jag-1/dnMAML-HEK293 cells (data not shown). Notch signaling was monitored by transient transfection of GFP/dnMAML1-, Jag-1/dnMAML-, and JICD/dnMAML-HEK293 cells with the Notch reporter construct to prove that co-expression of dnMAML in HEK293 cells prevents Jagged-1-mediated Notch activation (Fig. 4Cii; P=ns). These observations provide evidence that JICD alone can induce SDF-1 expression, although the activity of physiologically cleaved JICD in Jagged-1-overexpressing cells is suppressed.
Down-regulation of Jagged-1 in human MSC promotes SDF-1 expression
To study the effect of Jagged-1 down-regulation on SDF-1 expression, primary MSC overexpressing dnMAML1 were transduced with scramble or Jagged-1-shRNA lentivirus-vector particles. The efficacy of knockdown (KD) for Jagged-1-shRNA was evaluated by qRT-PCR. Jagged-1 mRNA was approximately 25% down-regulated in Jagged-1 KD primary dnMAML-MSC (Jag-1KD/dnMAML-MSC) compared with scramble-transduced MSC (scramble/dnMAML-MSC) (Supplementary Fig. S2B). Jagged-1 KD in primary MSC induced SDF-1α and SDF-1β expression compared with scramble-transduced cells (Fig. 4D). Furthermore, increased secretion of SDF-1α by Jag-1KD/dnMAML-MSC was confirmed by ELISA (Supplementary Fig. S2C). These findings suggest that full-length Jagged-1 partially inhibits SDF-1 expression under steady-state conditions.
JICD-overexpressing MSC promote support of CD34+ cells proliferation in 7 day culture
To determine whether overexpression of Jagged-1 or JICD alone influences the supportive potential of MSC, we evaluated the ability of MSC to support the proliferation of HSPC in a short-term co-culture. In this study, we distinguished between two populations of cultured HSPC: adherent and nonadherent cells, including those beneath the MSC layers (Fig. 5A). Freshly isolated CD34+ cells were cultured for 1 week, and nonadherent and adherent cells were collected at the indicated time point. To exclude variations in proliferation and viability of transduced MSC as a cause of different HSPC-supportive potential, we performed MTT and EdU proliferation assays. In addition, we analyzed the absolute number of MSC after 1 week in co-culture with HSPC. No significant differences were found between GFP/dnMAML-, Jag-1/dnMAML-, and JICD/dnMAML1-MSC (Supplementary Fig. S3).
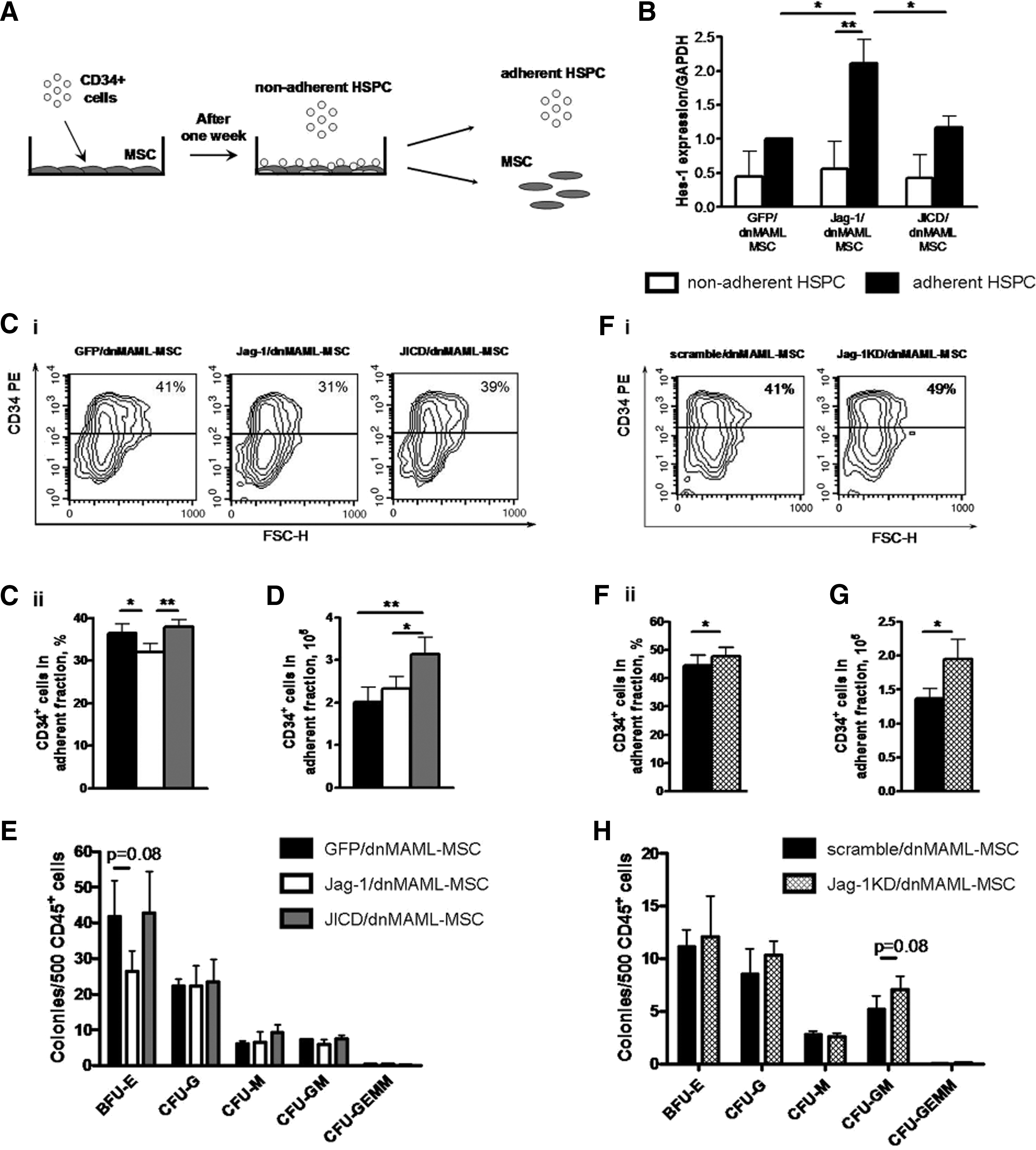
JICD-overexpressing MSC promote support of CD34+ cell proliferation in a short-term culture.
To evaluate the induction of Notch signaling in cultured HSPC, we assayed Hes-1 expression in adherent and nonadherent HSPC fractions after 1 week in co-culture. There was no variation in the Hes-1 expression level in nonadherent HSPC, whereas among adherent HSPC, endogenous Notch signaling was induced in cells cultured with Jagged-1-overexpressing MSC in comparison with other MSC layers (Fig. 5B; Jag-1/dnMAML-MSC vs. GFP/dnMAML-MSC, 2.1±0.6-fold change, P<0.05; JICD/dnMAML-MSC vs. GFP/dnMAML-MSC, and 1.2±0.2-fold change, P=ns). Moreover, Hes-1 expression in adherent HPSC cultured with Jag-1/dnMAML-MSC was higher than in nonadherent cells (Fig. 5B; adherent 2.1±0.6-fold vs. nonadherent 0.6±0.6-fold changes, P<0.01). These data indicate that co-culture with MSC overexpressing Jagged-1 resulted in the induction of Notch signaling in HSPC in a cell-to-cell contact-dependent manner.
After 1 week, all MSC layers showed better potential for expanding the total number of HSPC (CD45+ cells) as compared with stroma-free culture conditions (Supplementary Fig. S4A). To monitor the self-renewal capability of cultured HSPC, we analyzed the expression of CD34, which is an accepted marker of HSPC [30,31], in adherent and nonadherent fractions by flow cytometry. After 1 week, we observed differences in CD34 expression depending on the MSC feeder layer in both adherent and nonadherent fractions of HSPC, with more pronounced effects in adherent fractions. We observed a decrease of CD34+ in adherent fractions of HSPC when cultured with Jagged-1-overexpressing MSC (Fig. 5C) as compared with the control MSC (32.0%±2.0% with Jag-1/dnMAML1-MSC vs. 36.5%±2.1% with GFP/dnMAML-MSC, P<0.05) and with JICD-overexpressing MSC (32.4%±2.0% with Jag-1/dnMAML1-MSC vs. 38.0%±1.6% with JICD/dnMAML1-MSC, P<0.01). In the nonadherent HSPC fractions, JICD-overexpressing MSC maintained an increase of CD34+ cells in comparison with control MSC, partially recapitulating the effects observed in adherent HSPC (Supplementary Fig. S4Bi).
Furthermore, we analyzed the absolute number of CD34+ cells in adherent and non-adherent fractions to evaluate the potential of different stromal layers to expand HSPC. After 1 week, JICD/dnMAML1-MSC maintained a higher number of adherent CD34+ cells (Fig. 5D) than the control MSC (3.1±0.4×105 cells with JICD/dnMAML1-MSC vs. 2.0±0.3×105 cells with GFP/dnMAML-MSC, P<0.01) and than Jag-1/dnMAML-MSC (3.1±0.4×105 cells with JICD/dnMAML1-MSC vs. 2.3±0.3×105 cells with Jag-1/dnMAML1-MSC, P<0.05). In nonadherent fractions, JICD-overexpressing MSC also promoted enhanced expansion of CD34+ cells compared with Jagged-1-overexpressing MSC, although not compared with control MSC (Supplementary Fig. S4Bii). Besides CD34, we analyzed expression of CD133, another marker of primitive HSPC [31,32]. We observed a similar tendency for CD133+ cells from adherent and nonadherent HSPC fractions (data not shown). Thus, these results indicate that MSC overexpressing only JICD increases the proliferation of CD34+ progenitors during 7 days of co-culture compared with control MSC and Jagged-1-overexpressing MSC with a more pronounced influence on adherent HSPC.
Next, we investigated the effect of Jagged-1- and JICD-overexpressing MSC on the differentiation potential of adherent HSPC cultured for 1 week on MSC layers. HSPC cultured with Jagged-1-MSC produced fewer colonies in clonogenic assay compared with control MSC, which was mainly due to the reduction in erythroid colonies (Fig. 5E; 26±6 colonies with Jag-1/dnMAML-MSC vs. 42±10 colonies with GFP/dnMAML1-MSC, although difference did not reach the level of statistical significance, P=0.08). Taken together, our data suggest that Jagged-1-overexpressing MSC suppress proliferation of HSCP, consequently reducing the output of progenitor cells, in particular erythroid progenitors.
Down-regulation of Jagged-1 in human MSC promotes support of CD34+ cells proliferation in short-term culture
To further address the role of Jagged-1 for the maintenance of HSPC ex-vivo, we evaluated the HSPC-supportive potential of Jag-1KD/dnMAML-MSC. Freshly isolated CD34+ cells were cultured for 1 week, and adherent and nonadherent cells were collected at the indicated time point. The total number of CD45+ HSPC after 1 week in co-culture with scramble/dnMAML- and Jag-1KD/dnMAML-MSC was higher compared with stroma-free culture conditions. In addition, Jag-1KD/dnMAML-MSC showed a superior potential of HSPC expansion compared with scramble-transduced MSC (Supplementary Fig. 4C).
Next, we analyzed the expression of CD34 separately in adherent and nonadherent fractions of HSPC after 1 week in co-culture. We observed a slight, but significant, increase in the proportion of CD34+ cells in the adherent fraction when cultured with Jag-1KD MSC (Fig. 5F (47.7%±3.4% with Jag-1KD/dnMAML1-MSC vs. 44.6%±3.7% with scramble/dnMAML-MSC, P<0.05). In the nonadherent HSPC fraction, no difference in the proportion of CD34+ cells was observed (Supplementary Fig. S4Di).
In order to evaluate the potential of Jag-1KD/dnMAML-MSC to expand primitive HSPC, we analyzed the absolute number of CD34+ cells in adherent and nonadherent fractions of HSPC. After 1 week, the concentration of adherent CD34+ cells was higher when cultured with Jag-1KD/dnMAML-MSC (Fig. 5G) compared with scramble-transduced MSC (2.0±0.3×105 cells with Jag-1KD/dnMAML1-MSC vs. 1.4±0.1×105 cells with scramble/dnMAML-MSC, P<0.05). In nonadherent fractions, the absolute number of CD34+ was also higher in a culture with Jag-1KD/dnMAML-MSC than with control MSC (Supplementary Fig. S4Dii).
Clonogenic assays revealed that HSPC cultured with Jag-1KD/dnMAML-MSC for 1 week yielded an increase in total colony numbers, which was primarily due to an increase in the absolute number of granulocytes and granulocytes–monocytes colonies (Fig. 5H), although differences did not reach the level of statistical significance.
Jagged-1-overexpressing MSC promote HSPC support in long-term culture
To test HSPC-supportive potential of Jag-1/dnMAML-, JICD/dnMAML-, and Jag-1KD/dnMAML-MSC in long-term co-culture, we seeded freshly isolated CD34+ cells onto different MSC layers for a 6 week culture. Cobblestone areas were evaluated weekly, and at the indicated time point, co-cultured cells were plated for 2 weeks in methylcellulose medium to screen for secondary CFC. Both Jagged-1- and JICD-overexpressing MSC maintained a higher number of cobblestone areas than control MSC until week 3 (Fig. 6A). At later time points, Jag-1/dnMAML-MSC was superior in promoting the formation of cobblestone areas (Fig. 6A). Results of the cobblestone area-forming cell (CAFC) assays corresponded to those of the long-term culture-initiating cells assays. We found more secondary CFC in HSPC cultured with Jagged-1-overexpressing MSC (Fig. 6B), which was primarily due to an increase in the absolute number of colony-forming unit granulocytes (Fig. 6C). Furthermore, only Jagged-1-overexpressing MSC maintained secondary CFC that are capable of generating all subtypes of colonies (Fig. 6D). Jagged-1KD/dnMAML-MSC provided a comparable support of CAFCs and secondary colonies at week 4 to 6 as controls (Supplementary Fig. S5).
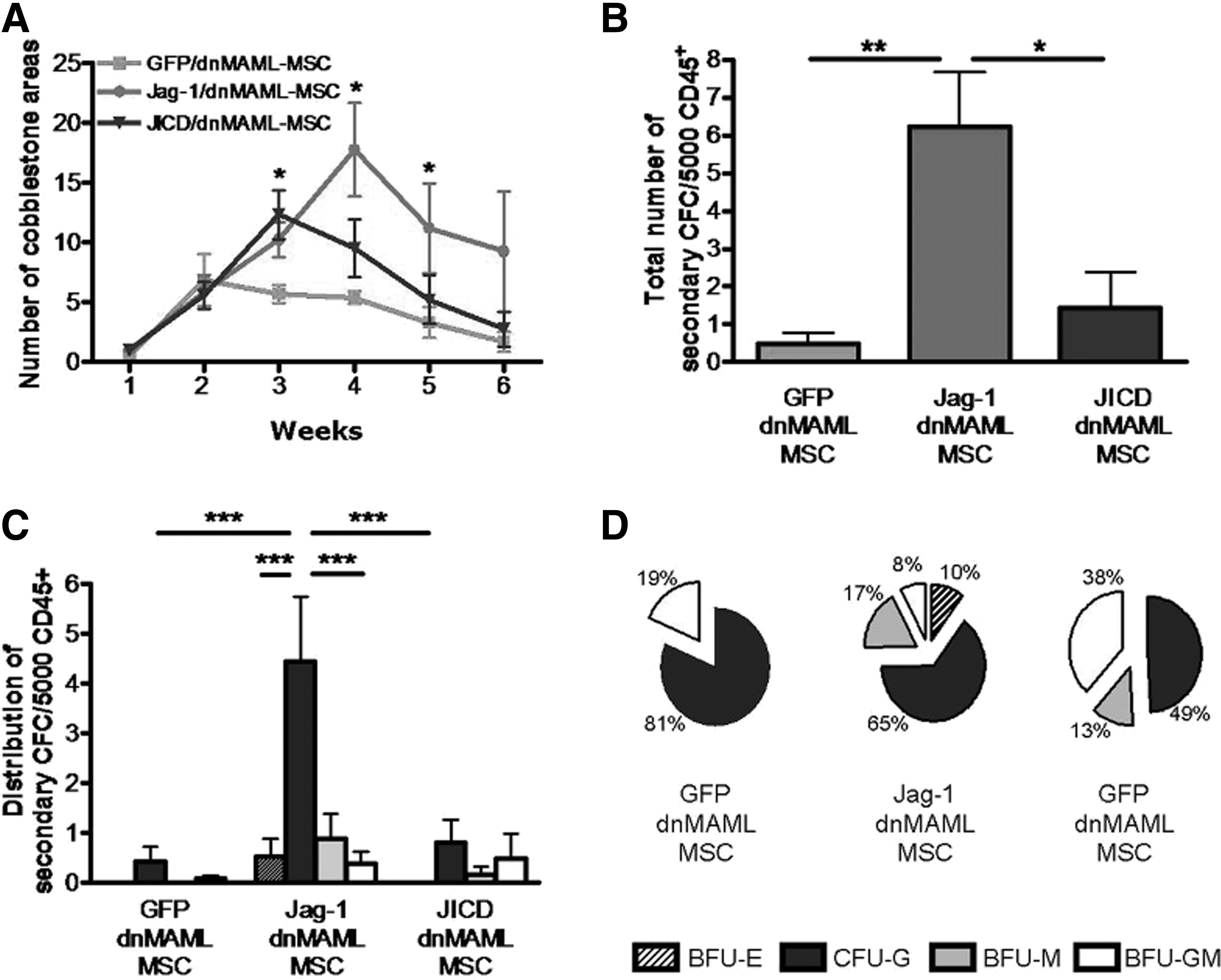
Jagged-1-overexpressing MSC promote HSPC support in long-term culture.
Jagged-1 and JICD do not affect osteogenic and adipogenic potential of MSC with blocked Notch signaling
To determine whether Jagged-1 or JICD independently of Notch signaling influences osteogenic or adipogenic potential of MSC, we investigated GFP/dnMAML-, Jag-1/dnMAML-, and JICD/dnMAML-MSC for their ex-vivo osteogenic and adipogenic potential. We observed no differences in osteogenic potential as evaluated by von Kossa staining and ALP activity measurement (Supplementary Fig. S6A) and no differences in adipogenic potential as evaluated by Oil Red staining and expression of late markers of adipocyte differentiation (Supplementary Fig. S6B).
Discussion
Notch signaling is one of the essential signaling pathways in HSPC [1]. Stromal cells expressing Notch ligands, in particular osteoblasts and MSC [33], induce Notch signaling in HSPC [6] maintaining HSPC self-renewal ability. In addition to Notch ligands, stromal cells themselves express Notch [33], which makes them susceptible to autocrine Notch signaling. While the biology of Notch receptors and the role of Notch signaling in the bone marrow are under active investigation, little is known about the independent role of Notch ligands in the bone marrow microenvironment.
Notch ligands themselves undergo cleavage, with the formation of ICDs that shuttle to the nucleus and regulate gene expression [16 –18,24]. In particular, it has been reported that ICDs of Notch ligands regulate the expression of p21 [25] and connective tissue growth factor [18] and possess the potential to bind to AP-1 response elements [17]. Moreover, recent reports have shown that the ICDs of Jagged-1 and Delta-1 interact with the NICD and negatively regulate Notch signaling [34,35]. These data demonstrate a previously unknown regulatory role for Notch ligands that could be realized either directly via regulation of gene promoters or via interaction with other signaling pathways.
In this study, we have investigated Notch-independent regulatory influence of Jagged-1, and, in particular, its ICD on human MSC and HSPC/MSC interactions. We used primary MSC overexpressing not only Jagged-1, but also the ICD of Jagged-1, to monitor the regulatory potential of physiologically cleaved and recombinant JICD. In agreement with previous reports, we have shown that Jagged-1 undergoes cleavage with formation of a CTF and an ICD in human MSC. We have shown that not only recombinant JICD, but also cleaved JICD translocates into the nucleus. Next, we speculated that JICD in MSC could realize its regulatory effects via regulation of HSPC-supportive genes containing AP-1 response elements in their promoter regions, as the potential of JICD to bind to AP-1 response elements has been reported by LaVoie and Selkoe [17]. Consequently, we assayed the expression levels of Angiopoietin-1, COX-2, and SDF-1 and found differences in the SDF-1 expression level. In the bone marrow, SDF-1 is expressed by stromal cells, and SDF-1/CXCR4 signaling has been shown to maintain the pool of HSPC [36,37] and to stimulate the homing of circulating HSPC to the bone marrow [38,39]. Interestingly, SDF-1 expression was up-regulated in JICD-overexpressing MSC, but not in Jagged-1-overexpressing cells. The results of reporter assays also clearly confirmed the transcriptional activation of SDF-1 mediated by recombinant JICD, but not physiologically cleaved JICD when JICD and Jagged-1, respectively, were transiently overexpressed in HEK293 cells. Our data suggest that the activity of endogenous JICD is suppressed in cells overexpressing Jagged-1. This observation raised the question of whether full-length Jagged-1 itself represses the expression of SDF-1 in human MSC.
To test whether Jagged-1 negatively regulates SDF-1 expression, we knocked down Jagged-1 in primary human MSC, again in the presence of dnMAML to exclude Notch-associated effects. Down-regulation of Jagged-1 resulted in up-regulation of SDF-1. Thus, these results confirm the suggestion that full-length Jagged-1 suppress SDF-1 gene expression.
To study the effects of Jagged-1/JICD overexpression and Jagged-1 KD within the MSC compartment on HSPC self-renewal, we used an ex-vivo co-culture system [40 –43]. Results of short-term culture (7 days) suggest that primary MSC overexpressing only JICD induce the expansion of primitive HSPC. Overexpression of Jagged-1 in primary MSC, in turn, does not favor early HSPC proliferation and reduces especially the erythroid clonogenic potential of co-cultured HSPC. In contrast to Jagged-1-overexpressing MSC, Jagged-1 KD MSC better support short-term HSPC expansion and colony formation. Superior supportive potential of both JICD-overexpressing MSC and Jagged-1 KD MSC toward HSPC proliferation may be explained by the higher levels of secreted SDF-1, as SDF-1/CXCR4 signaling has been shown to promote human CD34+ cell proliferation and colony formations during ex-vivo expansion [38,44].
Previous reports have described improved strategies for the expansion of HSPC that are capable of in-vivo reconstitution using ligand-mediated activation of Notch in HSPC. These systems are mostly based on immobilization of Notch ligands [8,45,46] or a co-culture system with murine stromal and fibroblast cell lines overexpressing human or mouse Notch ligands [7,47 –49]. Although shown to be effective, these approaches do not fully recapitulate the in-vivo environment. Human proteins overexpressed in murine cell lines may not maintain their signaling potential due to species differences. This could explain why our observations with human HSPC cultured with human Jagged-1-overexpressing MSC for a short term differ from previous reports describing effects of ligand-mediated Notch signaling activation on human HSPC expansion using mouse cell lines [7,47 –49]. Unlike the immobilization of Notch ligands to culture carriers, the overexpression of Jagged-1 in a more complex cellular system, as tested here, may affect the supportiveness of MSC toward HSPC by paracrine effects of the overexpressed proteins, which seems to be counterproductive in terms of rapid expansion of CD34+ HSPC within 1–2 weeks. Still, longer culture periods revealed the supportive effects of Jagged-1 overexpression.
Results of long-term co-cultures demonstrated that both Jagged-1- and JICD-overexpressing MSC favor the formation of “early” CAFCs compared with control MSC, while Jagged-1 overexpressing MSC favors the formation of “late” CAFCs. In addition, with regard to CAFC assay, Jagged-1-overexpressing MSC maintain more secondary CFC after long-term co-culture than JICD-overexpressing and control MSC.
Taken together, our results provide new insights into the signaling role of Notch ligands in MSC and their HSPC-supportive potential. Cleavage of Jagged-1 releases endogenous JICD, which, similar to recombinant JICD, translocates into the nucleus. In contrast to recombinant JICD, endogenously cleaved JICD in Jagged-1-overexpressing cells does not induce SDF-1 expression, supporting the hypothesis that full-length Jagged-1 itself suppresses SDF-1 expression; whereas down-regulation of Jagged-1 induces an up-regulation of SDF-1. Our results suggest that the reduced expression of SDF-1 in Jagged-1 overexpressing MSC is associated with a less pronounced proliferation of CD34+ cells in the first weeks of co-culture and a better maintenance of long-term (6 weeks) culture initiating cells. Further experimental work, including relevant in-vivo models, is required to fully elucidate the biological effects of full-length Notch ligands and their ICDs on the hematopoietic environment.
Footnotes
Acknowledgments
The authors thank Fernando Ugarte for his initial studies that enabled the experiments described here. They also acknowledge Katrin Müller for providing and handling MSC samples, Jun Fu for assistance with reporter-construct design, Matthias Schieker for providing them with the Scp-1 cell line, Frank Buchholz for providing them with scramble and Jagged-1-shRNA vectors, Aylin Camgoz for assistance with handling scramble and Jagged-1-shRNA vectors, and Ute Hempel and Carolin Preissler for performing ALP activity measurements. This work was supported by the DFG grant Collaborative Research Center 655 “Cells into tissues: Stem cell and progenitor commitment and interactions during tissue formation” (Projects B1 to K.A., B2 to M.B. and C.W., and B6 to S.B.).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.