
Abstract
Misexpression of the double homeodomain protein DUX4 in muscle is believed to cause facioscapulohumeral muscular dystrophy (FSHD). Although strategies are being devised to inhibit DUX4 activity in FSHD, there is little known about the normal function of this protein. Expression of DUX4 has been reported in pluripotent cells and testis. To test the idea that DUX4 may be involved in initiating a germ lineage program in pluripotent cells, we interrogated the effect of expressing the human DUX4 gene at different stages during in vitro differentiation of murine embryonic stem (ES) cells. We find that expression of even low levels of DUX4 is incompatible with pluripotency: DUX4-expressing ES cells downregulate pluripotency markers and rapidly differentiate even in the presence of leukemia inhibitory factor (LIF) and bone morphogenetic protein 4 (BMP4). Transcriptional profiling revealed unexpectedly that DUX4 induced a neurectodermal program. Embryoid bodies exposed to a pulse of DUX4 expression displayed severely inhibited mesodermal differentiation, but acquired neurogenic potential. In a serum-containing medium in which neurogenic differentiation is minimal, DUX4 expression served as a neural-inducing factor, enabling the differentiation of Tuj1+ neurites. These data suggest that besides effects in muscle and germ cells, the involvement of DUX4 in neurogenesis should be considered as anti-DUX4 therapies are developed.
Introduction
D
On the other hand, the normal function of DUX4 remains mysterious. The mouse has a homologue named Dux, which is also arrayed in tandem repeats (although fewer in number than those of the human) and expressed in various tissues, but most prominently in neurogenic tissues [13]. In humans, expression has been reported in testis [6] and a role in the expression of germ cell cancer/testis antigens demonstrated [14]. DUX4 message has also been detected at low levels in human embryonic stem (ES) cells [15].
To investigate the role of DUX4 in early development, we created a mouse ES (mES) cell line that would inducibly express DUX4. The early stages of embryo development are well recapitulated in mES cell growth and embryoid body (EB) differentiation; thus, these cells allowed the interrogation of the activity of this transcription factor on the development of all of major lineages. Although we had anticipated that DUX4 might drive germ line differentiation, we found only a very modest effect on germ line-specific genes and no evidence of directing pluripotent cells into a germ cell fate. Rather, DUX4 induced a neuroectodermal gene expression program at the expense of other major lineage-specific programs, and was able to drive neurogenesis in vitro even under nonoptimized conditions in which minimal neurogenesis normally occurs.
Materials and Methods
iDUX4 ES cell generation and culture
The DUX4 ORF was subcloned into p2Lox, the targeting vector for the inducible cassette exchange (ICE) recombination system [16]. DUX4-inducible ES cells were generated by cassette-inducible exchange recombination (ICE) into the doxycycline-inducible locus upstream of HPRT in ZX1 mES cells, an improved version of A2Lox.cre [17]. ZX1 cells were used as the parent control cell line in experiments. iDUX4 ES cells were cultured on mouse embryonic fibroblasts (MEFs) in Dulbecco's modified Eagle's medium (DMEM) (Invitrogen 10829-018) supplemented with 15% fetal bovine serum (FBS), 0.1 mM nonessential amino acids (Gibco), 2 mM glutamax (Invitrogen), penicillin/streptomycin (Gibco), 0.1 mM β-mercaptoethanol (Sigma), and 1,000 U/mL leukemia inhibitory factor (LIF) (Millipore), at 37°C in 5% CO2.
Embryoid body differentiation
ES cells were differentiated as EBs by preplating for 30 min to remove MEFs by attachment, followed by suspension culture in hanging drops (100 cells per 10 μL drop) in the EBD medium: Iscove's modifed Dulbecco's medium (IMDM, Invitrogen) supplemented with 15% FBS, 200 μg/mL iron-saturated transferrin (Sigma), 4.5 mM monothiolglycerol (Sigma), 50 μg/mL ascorbic acid (Sigma), penicillin/streptomycin (Invitrogen), and 2 mM glutamax (Invitrogen) at 37°C in 5% CO2, 5% O2. After 48 h, EBs were harvested from hanging drops by collecting and settling in IMDM. They were then resuspended in 10 mL of EBD and plated in nonadherent 10-cm dishes on a swirling rotator (70 rpm). EBs were fed after 48 h by exchanging 50% of a spent medium for fresh EBD medium. Doxycycline induction was started at day 2 with the concentrations indicated, and EBs were harvested either at day 4 or 6.
Growth curve
ES cells on MEFs were disassociated with trypsin (Invitrogen) and the MEFs were removed by preplating for 30 min. The 1×105 cells were plated on 35-mm tissue culture dishes in the knockout serum replacement (KOSR) medium (Gibco) supplemented with 1,000 U/mL LIF and 10 ng/mL bone morphogenetic protein 4 (BMP4) (Peprotech) along with 0, 100, 200, 300, 400, or 500 ng/mL doxycycline. Every 2 days, cells were passaged, cell numbers were counted from triplicate wells of each passage, and 1×105 cells were replated in the same conditions.
Fluorescence-activated cell sorting
Live cells were gated based on forward scatter/side scatter profile, and negativity for propidium iodine staining to eliminate debris and dead cells. The following antibodies were used: c-Kit-allophycocyanin (APC) (2B8), CD41-phycoerythrin (PE) (eBioMWREG30), CD31-Cy7-PE (390), CD45-PE (30-F11), Flk1-APC (Avas12a1), Flk1-PE (Avas12a1), platelet-derived growth factor receptor alpha (PDGFRα)-PE (APA5), SSEA1 (eBioMC-480), Tie-2 (TEK4), (all from eBiosciences) and polysialylated neuronal cell adhesion molecule (PSA-NCAM)-APC (130-093-272; Miltenyi Biotec). Cells were incubated with 1 ul of antibody per one million cells and analyzed and/or sorted on a fluorescence-activated cell sorting (FACS) Aria II (BD Biosciences).
Cell staining
Live cells or cells fixed first with 4% paraformaldehyde were imaged using phase-contrast microscopy (Evos; Advanced Microscopy Group). For immunostaining, cells were grown as described and fixed with 4% paraformaldehyde in the same plates as they were grown in. Cells were permeabilized, blocked, and immunostained according to the manufacturers' directions for each antibody: DUX4 (E5-5; Epitomics) or Nestin (AB0968100; Abcam) both used with goat anti-rabbit Alexa Fluor-555 conjugated secondary antibody (Invitrogen A21429), or Tuj1 conjugated with Alexa Fluor 488 (A488-435; Covance). Nuclei were imaged with DNA-staining 4′,6-diamidino-2-phenylindole (DAPI) dye. Images were taken with a Zeiss Observer Z1 using an Axiocam MRM camera and software.
Quantitative reverse transcription-polymerase chain reaction
RNA was extracted with Ambion RNAqueous® Kit (Life Technologies/Invitrogen); reverse transcription was performed with ThermoScript™ Reverse Transcriptase (RT) (Invitrogen). Quantitative polymerase chain reaction (qPCR) was performed using real-time PCR (7500 or 7900 Real-Time PCR Systems; Applied Biosystems). In brief, 40 cycles at 95°C for 12 s and 60°C for 30 s using TaqMan Gene Expression Assay primer–probe sets (Applied Biosystems). Quantitative analysis of gene expression was conducted using the comparative cycle threshold (Ct) method and means were normalized first to GAPDH, and then to noninduced controls and were compared by the t-test. All PCRs were done in triplicate with one control reaction containing cDNA that was reverse transcribed without the RT enzyme.
Microarray experiments
DUX4 was induced with 100 μg/mL doxycycline from EB day 2 and harvested after 48 h of doxycycline induction. The RNA from three independent replicates of both +doxycycline and control was used to generate the cRNA probe, which was hybridized to MouseWG-6 Bead Chip Arrays (Illumina). Raw data were processed using Beadstudio (Illumina) and analyzed using Genespring GX 7.3.1 (Agilent).
Results
Low-level expression of DUX4 in mES cells downregulates the pluripotent program and promotes neuroectodermal gene expression
Previously, we found that high levels of DUX4 expression (from a construct that contained the ORF+1.5 kb of 3′ UTR) are toxic to myoblast cells and that low levels of DUX4 inhibited myogenic differentiation, suggesting a mechanism for the involvement of misexpressed FSHD in muscle pathology [12]. We generated a doxycycline-inducible DUX4 (iDUX4) mES line by recombining the DUX4 ORF into mES cells with an ICE locus upstream of HPRT [16]. In these ES cells, DUX4 protein expression was visible in the nuclei of cells at 24 h postinduction in iDUX4 cells, but not in uninduced cells (Fig. 1A) and there was a direct relationship between the amount of doxycycline added and the amount of DUX4 RNA (Fig. 1B and Supplementary Fig. S1A; Supplementary Data are available online at
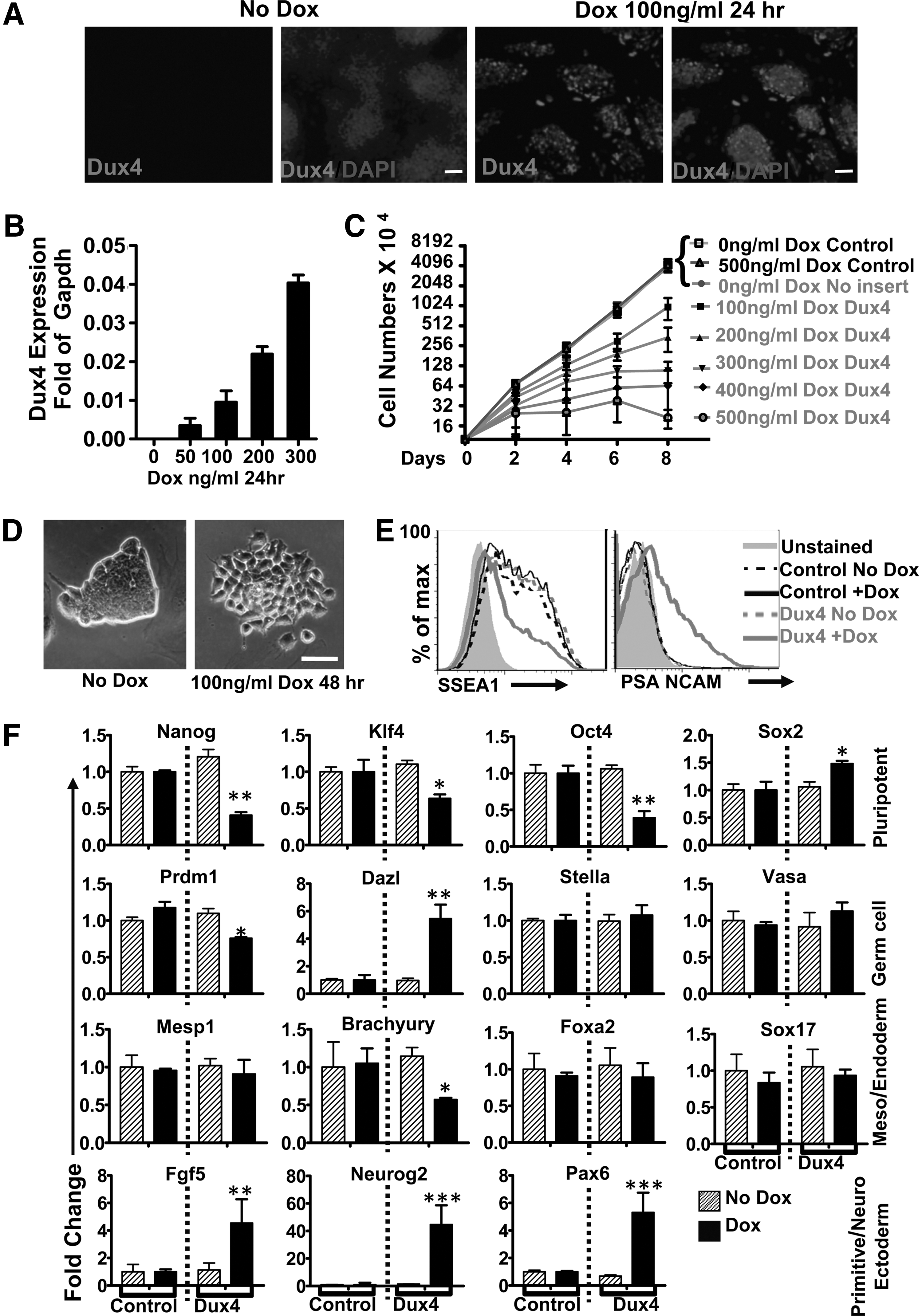
DUX4 perturbs pluripotency and induces neural gene expression.
To determine whether this morphological change indicated a change in pluripotency, we induced DUX4 at low levels for 48 h and evaluated a panel of pluripotent markers and differentiation, both by FACs as well as qRT-PCR (Fig.1E, F). With the exception of Sox2, which was upregulated, all markers of pluripotency, SSEA1, Nanog, Klf4, and Oct4 were downregulated with low-dose DUX4 induction. Since DUX4 was reported to be expressed in testis and to induce germ cell antigens (among many other changes) when expressed in human myoblasts [6,14], we examined germ cell lineage markers. These results were not consistent with the activation of a germ cell program: the master regulator, Prdm, was significantly downregulated; the germ cell marker, Dazl, was upregulated, while Stella and Vasa showed no change. Since a primodial germ cell linage was not being induced, we evaluated markers of meso-, endo-, and ectoderm. Early mesodermal cell fate markers showed no change (Mesp1) or were downregulated (Brachyury), while endoderm markers Foxa2 and Sox17 showed no change. Although Sox2 is expressed in pluripotent cells, it is also a key element of the early neurogenic program [18,19], so its upregulation might be interpreted as the induction of a neurogenic program. Supporting this idea, all the neuroectoderm markers evaluated (PSA-NCAM, Neurog2, and Pax6) were upregulated. In addition, it should be noted that the epiblast marker Fgf5 was upregulated, suggestive of a transition through epiblast to neuroectoderm.
DUX4 promotes neuroectodermal differentiation during EB differentiation
Although the monolayer differentiation study above clearly shows that DUX4 interferes with the pluripotent program, monolayer differentiation is not an efficient way to generate the mesoderm or endoderm. To test the effect of DUX4 expression on the generation of the three germ layers, we subjected iDUX4 ES cells to EB differentiation. During the first 2 days of EB differentiation, cells progress from the ES stage to epiblast. After this time point, the three germ layers are specified, with the earliest germ layer-specific markers appearing shortly afterward. We therefore applied 100 ng/mL of doxycycline at 48 h of differentiation and evaluated EBs 48 h later (after 4 days of differentiation). Under these conditions, doxycycline induction of DUX4 caused EBs to be somewhat more compact (Fig. 2B). We then performed microarray profiling on total EB RNA. We found no significant differences between induced and uninduced samples for most germline-specific or endodermal markers (Fig. 2C).
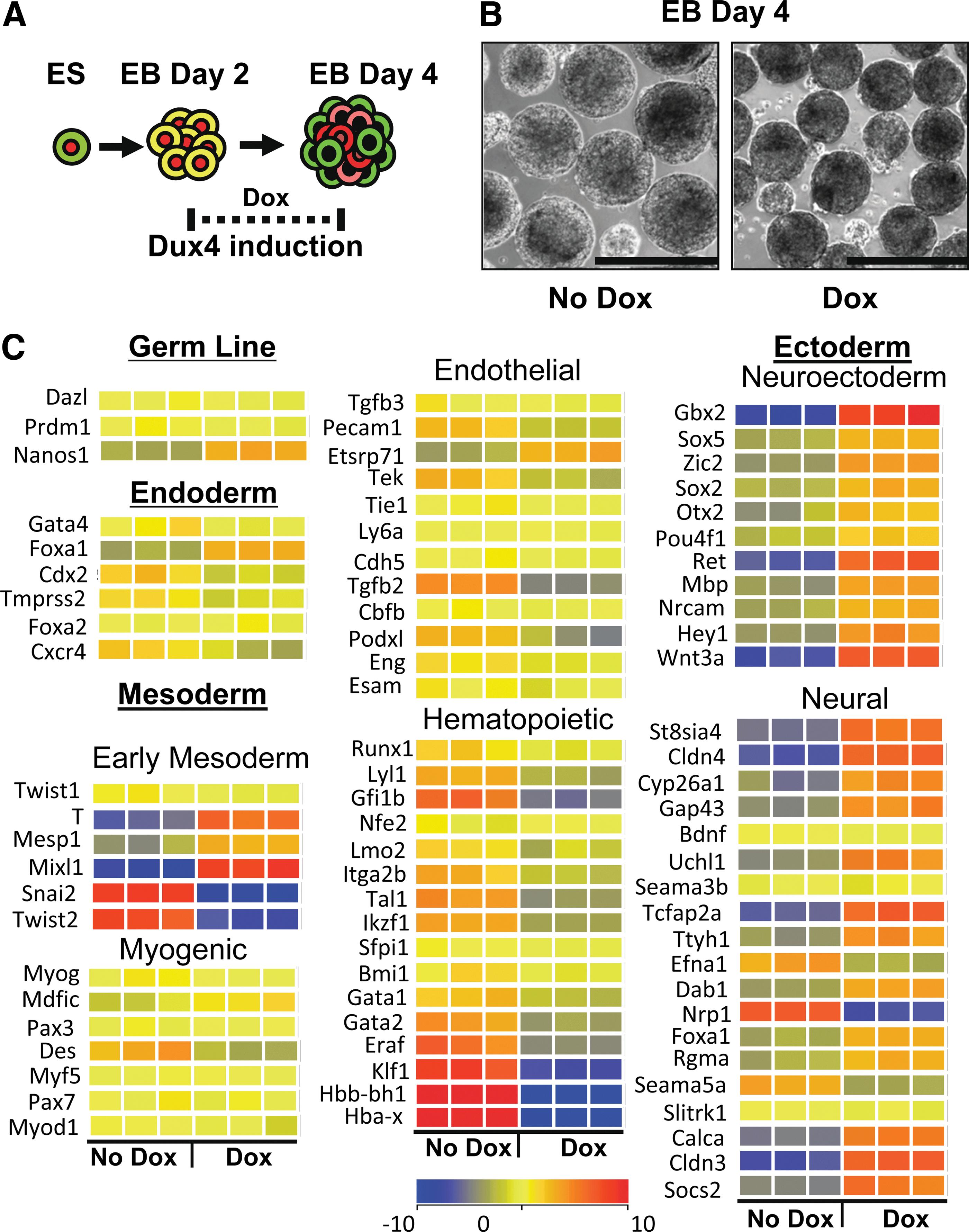
Microarray profiling of day 4 embryoid bodies (EBs) demonstrates neurogenesis.
For genes associated with early mesodermal lineages, results were mixed with some early markers upregulated, however, this likely reflects lack of appropriate downregulation due to impaired differentiation of early specified mesoderm lineage founders as these markers normally peak at day 3 and are much lower at day 4. By this time point, the first wave of mesodermal differentiation is well underway, and this first wave prominently produces hematopoietic lineage founders, while other mesodermal lineages arise slightly later [20]. Looking at the signature for this first wave, we observed clear downregulation of both hematopoietic transcription factors and globins. Coordinate with this was a decrease in most endothelial markers. However, the most striking signature change was that of the neurectoderm and neural genes. This lineage is typically quite underrepresented in EBs grown in serum, however, when low levels of DUX4 are induced, almost every marker is upregulated. These results are consistent with DUX4 expression in early development inhibiting various mesodermal lineages particularly the lateral plate and promoting neuroectoderm/neurogenesis.
To support this interpretation, we repeated low-dose DUX4 inductions starting at 48 h, and interrogated various markers specifically, including surface markers that have been associated with specific lineages, and RNA markers by qRT-PCR (Fig. 3A). Day 4 EBs were examined for the early mesoderm markers Flk1 for lateral plate [21], and PDGFRα for paraxial mesoderm [22] by FACS analysis (Fig. 3B). In uninduced cultures, differentiating EBs developed into Flk1−PDGFRα+ (10.6%), Flk1+PDGFRα+ (34.3%), and Flk1+PDGFRα− (9.6%) populations on day 4 of differentiation. Doxycycline induction increased the PDGFRα single-positive population to 44.4%, and decreased Flk1 expression to negligible levels. While there was a similar proportion of PDGFRα+ cells in treated and untreated samples (45% and 46%), most of these were single-positive with doxycycline treatment. PSA-NCAM is expressed in early neuronal progenitor cells [23 –25] and functions in cell migration [26]. Interestingly, this protein was expressed with DUX4 induction, but almost absent in uninduced EBs and also absent in control induced and uninduced cells from the parent cell line of iDUX4 (Supplementary Fig. S1B). Although PDGFRα marks the paraxial mesoderm, it also marks cells that will become oligodendrocyte progenitors [27]. The presence of double-positive PSA-NCAM/PDGFRα cells suggests DUX4 is promoting a neuronal cell fate (Fig. 3C). To examine the effects of DUX4 on the lateral plate mesoderm, we performed FACs with PSA-NCAM/Flk1 and found that DUX4 inhibits the lateral plate mesoderm, while at the same time promoting neuroectoderm (Fig. 3D). We further explored the induction of neurogenesis by quantitative reverse transcription PCR (qRT-PCR) analyses for expression of selected neuroectodermal genes. Pax6, Neurog2, Gbx2, Otx2, Sox2, and Nestin were all significantly elevated in DUX4-induced EBs (Fig. 3E and Supplementary Fig. S1C). These results support the interpretation that DUX4 promotes early neural progenitors, but inhibits mesoderm, particularly the lateral plate mesoderm.
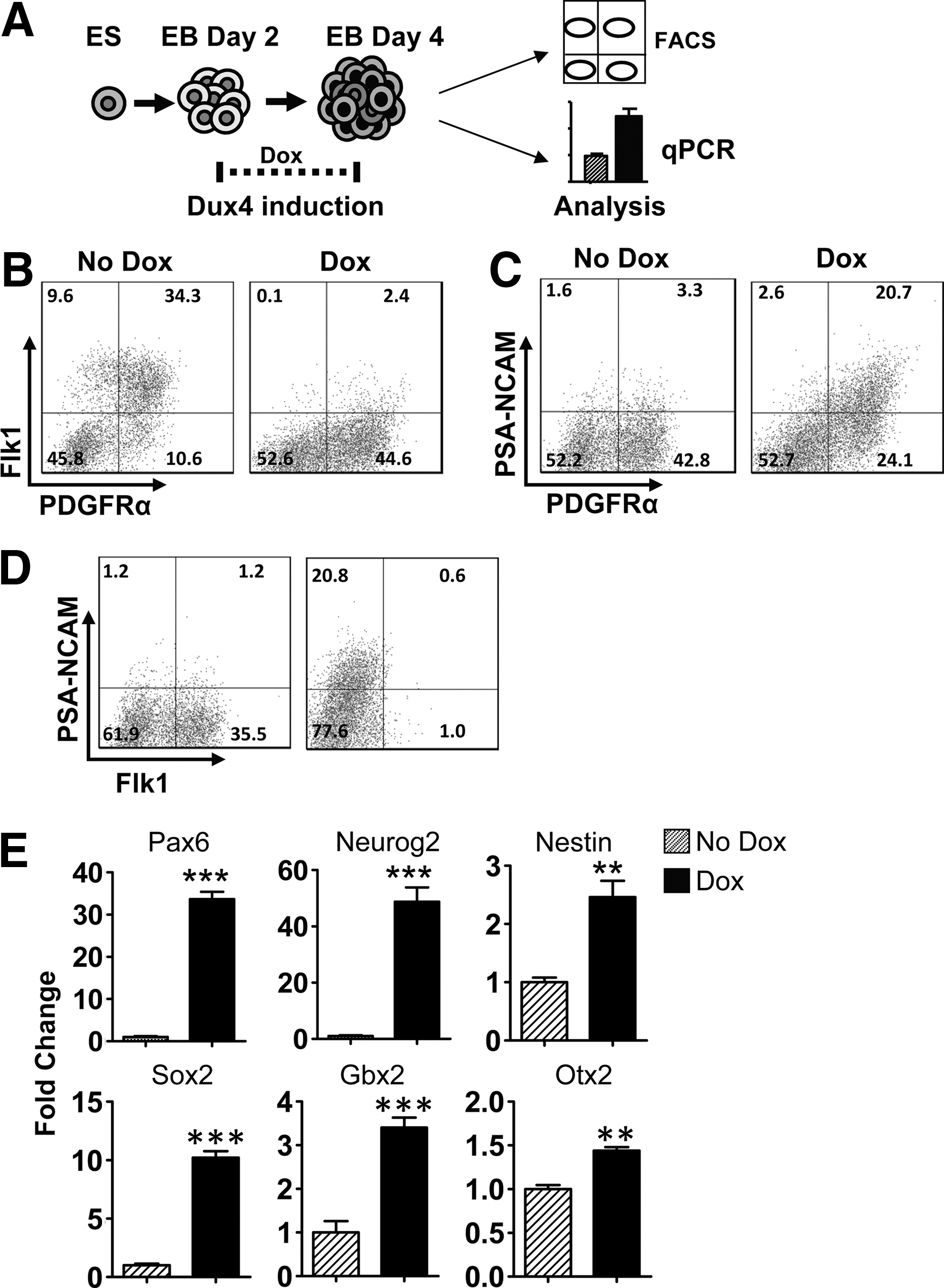
DUX4 inhibits lateral plate mesoderm and promotes neurectoderm in day 4 EBs.
We performed additional experiments at a later stage of EB development to examine the effects of DUX4 on hematopoietic, endothelial, and cardiac progenitors. DUX4 was induced by applying doxycycline at 100 ng/mL from days 2–6 and cells were assayed by FACS and qRT-PCR (Fig. 4A). In accordance with the previous results, we obtained fewer undifferentiated hematopoietic progenitors (c-Kit/CD41, c-Kit/CD45), and fewer Flk1/Tie2 or Flk1/CD31 endothelial cells (Fig. 4B). To evaluate cardiogenesis, we analyzed two early cardiac markers Nkx2.5, Tbx5 [28] and the terminal marker cardiac troponinT2 (cTnnt2) by qRT-PCR and found the expression of all of these markers significantly inhibited by DUX4 induction in comparison to uninduced samples (Fig. 4C). These results demonstrate that DUX4 is incompatible with the differentiation of the major mesodermal lineages assayable in EBs.
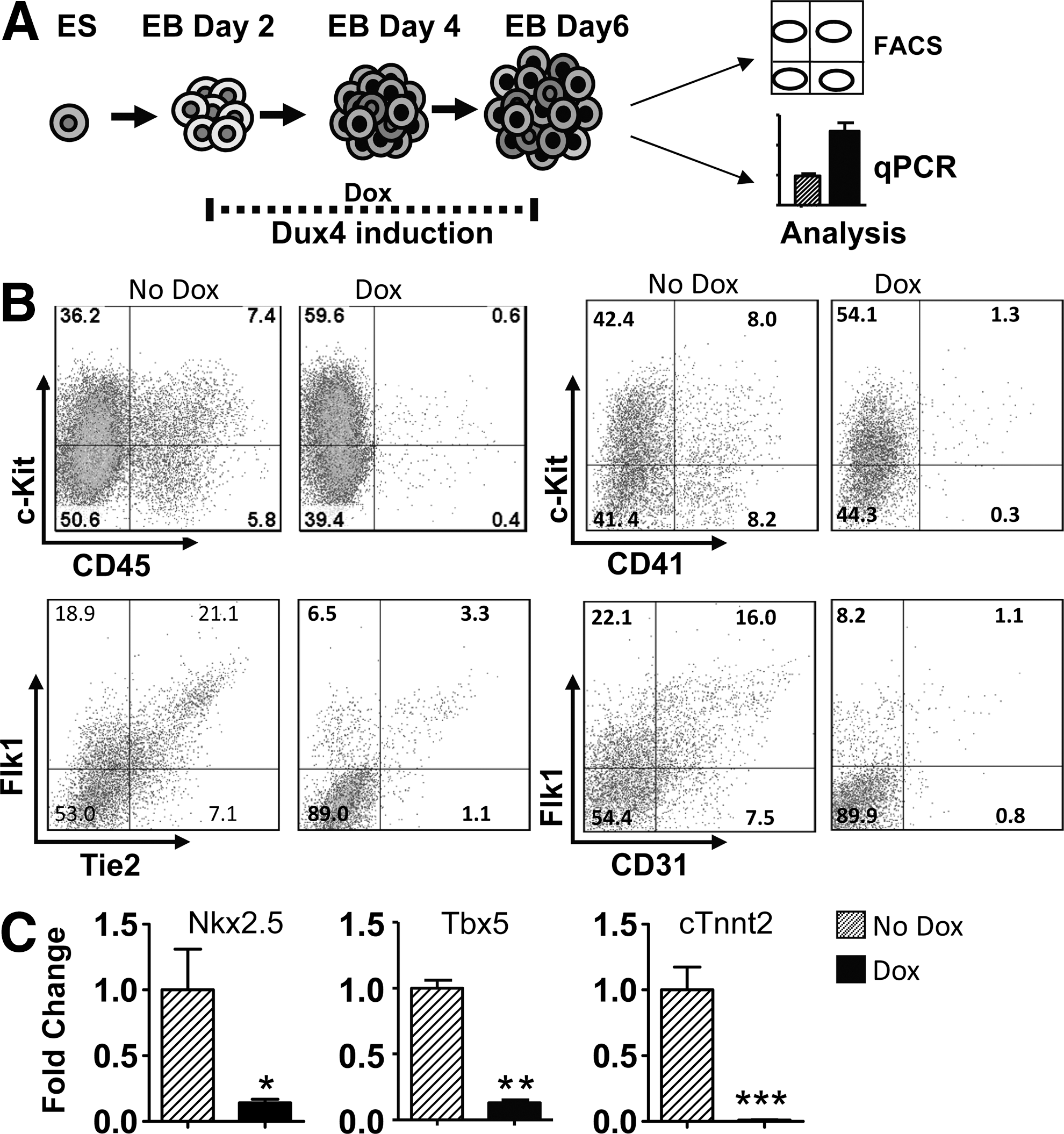
DUX4 inhibits cardiac and hemangiogenic progenitors in day 6 EBs.
DUX4 promotes neurogenesis
We wished to follow up on the observation that DUX4 promotes neurectoderm, to see whether it could generate differentiated neural cell types under conditions in which, few such cells are normally formed. EBs were formed by hanging drops for 2 days, and then moved to a swirling suspension with doxycycline being added from day 2 through day 6, when EBs were allowed to attach to a solid surface and grow for an additional 3 days to allow for terminal differentiation (Fig. 5A). In samples not treated with doxycycline, the attached EBs gave rise to minimal neurite-like sprouts, while doxycycline induced a tremendous number of sprouts. To quantify this observation, we counted the number of EBs displaying sprouts. The DUX4-induced EBs had significantly more neurite-like sprouting outgrowths compared to uninduced samples (Fig. 5B, C and Supplementary Fig. S2A). These cultures were fixed and stained for Tuj1, which identifies β-tublin III, a structural protein of mature neurons. There was minimal Tuj1 staining in control EBs, but in DUX4-induced EBs, Tuj1 was visible throughout the attached colonies, and particularly strongly detected in the sprouts, confirming their neuronal identity (Fig. 5D). This differentiation was in the absence of conventional neuronal-inducing or -supportive factors like retinoic acid, basic fibroblast growth factor, or a medium optimized for neuronal differentiation of ES cells. We also examined cultures in which, EBs had been trypsinized and cells plated directly on plastic for Tuj1 immunofluorescence 2 days later, which also demonstrated Tuj1+ cells (Fig. 5E and Supplementary Fig. S2B). Since Tuj1 is a marker associated with later stage development of neurons, we also tested for an early marker of neuronal development, Nestin. We dissociated cells from day 6 EBs and examined them 2 days after plating onto gelatin-coated dishes. We found that DUX4 potently induced Nestin+ cells, while there was very little staining in uninduced cultures (Fig. 5F and Supplementary Fig. S2C). Finally, we evaluated several neural markers over a time course from days 6 through 9. Three terminal neural differentiation markers were examined. Tuj1 was dramatically upregulated by DUX4 (Fig. 5G). There was also significant upregulation of expression of two glial genes for terminal differentiation: GFAP, a marker for astrocytes and Olig2, a marker for oligodendrocytes. These experiments show that DUX4 not only promotes neuroectodermal fates, it can induce the formation of neuronal progenitors that are capable of terminal differentiation.
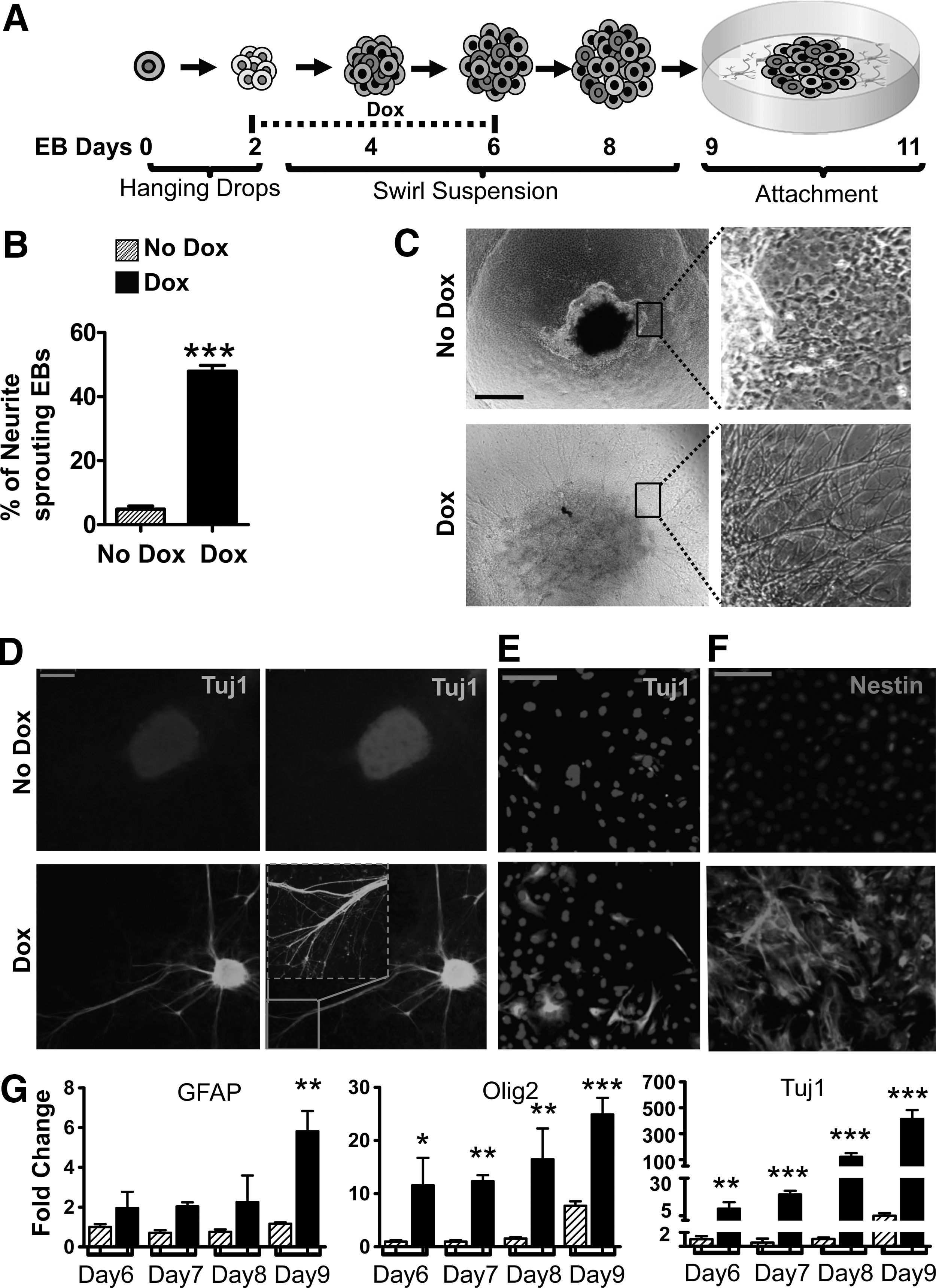
DUX4 promotes neuronal differentiation.
Discussion
The recent correlation of FSHD pathogenesis with the chromosome 4 permissive allele that contains a signal sequence allowing polyadenylation of DUX4 RNA from the last repeat [5], has encouraged researchers to consider using methods to reduce DUX4 RNA levels as a therapy for patients [29]. However, it is likely that DUX4 expression has a role in normal development because the open reading frame encoding DUX4 is clearly under positive selection, that is, if it were not functional and required for something essential, it would have acquired mutations that disrupt the ORF [13]. Therefore, it is important to investigate the normal function of DUX4 when therapies reducing the level of DUX4 in cells are considered. For this reason as well as to gain a further understanding of the effects of ectopic expression of DUX4 in cells, we have determined the effect of expressing the human DUX4 gene during mES cell in vitro differentiation, a powerful system for determining the function of genes involved in development.
Using a doxycycline-inducible promoter to express DUX4 allowed us to titrate down the level of expression, to a level at which DUX4 was no longer potently cytotoxic to ES cells growing in a medium with beta-mercaptoethanol. Achieving low levels of expression is important, as the physiological level of DUX4 expression in primary cells, even in FSHD muscle cells, is low [4,6,15,30]. Most previous studies on DUX4 action in cells have used overexpression from strong promoters [4,14,31], and there is clearly a high dose-dependent toxicity of DUX4. However, very low doses that do not cause overt cell death are able to inhibit MyoD expression and block myogenic differentiation of C2C12 cells [12], effects that may be more relevant to FSHD than is cell death.
We found that pluripotency was rapidly compromised by low levels of DUX4. Within 2 days, pluripotent markers were virtually eliminated (SSEA1, Nanog, Oct4, and Klf4). This strongly suggests that DUX4 does not play a role in maintaining cells in the pluripotent stage. However, there have been reports of DUX4 RNA expression in WT hES and induced pluripotent stem (iPS) cells [6,15]. We suspect that this may be due not to expression in pluripotent cells, but to expression in some type of early differentiated cells, of which, there are always a few in healthy karyotypically normal ES cell cultures. Because germ cells are closely related to ES cells, because DUX4 has been demonstrated to be expressed in human testis [6], and because overexpression of DUX4 in human myoblasts induced the expression of a selection of germ cell cancer antigens [14], we presumed initially that such rare DUX4-expressing cells in hES cultures might be early germ lineage progenitors, and therefore, we would find that DUX4 would drive ES cells into the germ lineage. However, we did find strong evidence for this. In ES monolayers, by qRT-PCR, the master regulator of primordial germ cell development, Prdm1, was downregulated. One germ cell marker, Dazl, was upregulated, while others, Vasa and Stella, were unchanged. By microarray in day 4 EB cells after a 48-h pulse of DUX4, Nanos was upregulated, but Prdm1 and Dazl were unchanged. On the other hand, a clear effect of DUX4 expression in both ES cells and in very early differentiating EBs was the induction of markers indicative of neurogenesis (PSA-NCAM, Sox2, Neurog2, Pax6). ES cells differentiated in vitro as EBs in a serum-containing medium give a preponderance of lateral plate mesoderm derivatives [32]. We found that ES cells differentiated under these conditions, but in the presence of DUX4 induction showed a severely inhibited lateral plate mesoderm and derivatives (blood and cardiac tissue), but had acquired neurogenic potential, even to the point of producing terminally differentiated neuronal cells. These were obtained when a pulse of low-level DUX4 expression was applied during EB differentiation, but then removed as EBs were attached to plastic and allowed to differentiate for several more days. Differentiated Tuj1+neurons were observed by day 11. It should be emphasized that in these experiments, the growth medium was devoid of exogenous retinoic acid, fibroblast growth factor 5 (FGF5), or other growth factors and cytokines that are usually required for neural induction of ES cells. In addition, they were done in the presence of serum, which is inhibitory to neurogenesis in EBs.
We have previously demonstrated a competitive interaction between DUX4 and Pax3/Pax7 for regulation of MyoD and other targets, and proposed that the similarity of the homeodomains of DUX4 to those of Pax3 and Pax7 may enable inappropriate interaction with Pax3/7 targets in satellite cells, which would perturb muscle regeneration [12]. It is interesting to note that both Pax3 and Pax7 are expressed in early neural development, and that the DUX4 homeodomains are equally similar to those of Pax6, an important neurogenic transcription factor. Based on the molecular similarities therefore, a role for DUX4 in neural differentiation is not unreasonable. We interrogated the neuroectodermal markers used in this study (Sox2, Pax6, and PSA-NCAM) for DUX4 consensus DNA-binding elements, and found no conserved sites for Sox2 or Pax6, and only a few nonconserved sites for NCAM, so these genes may not be direct targets. Most of the direct DUX4 targets in myoblast cells appeared to be within primate-specific MaLR repeat elements, possibly suggesting acquired human-specific activities [14]; however, we note that several without primate-specific MaLR sites, have functions in some aspect of neurogenesis or neural function, for example, RFPL1 and 2 [33], and PPP2R2B [34]. Studies of a related gene in mouse, Dux, found expression in brain tissues [13]. The particular expression pattern observed suggested that mouse Dux might have a role in embryonic development within granule neurons and when young mouse brains were examined, expression was observed throughout the cortex with localized areas of increased expression. Taken together, the experiments presented here suggest the importance of looking beyond muscle and germ line function to understand DUX4 and that we should consider the possibility that DUX4 has a role in neurogenesis.
Footnotes
Acknowledgments
We thank Sunny Chan for helpful suggestions with this study and Cynthia DeKay for help with illustrations. The project was supported by the Bob and Jean Smith Foundation and by NIH grant R01 AR055685. D.B. was supported by a Muscular Dystrophy Association Development Grant (MDA 4361) and a Marjorie Bronfman Research Fellowship from the FSH Society (FSHS-MGBF-016).
Author Disclosure Statement
The authors declare no competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.