
Abstract
Neurogenesis occurs continuously in two brain regions of adult mammals, underpinned by a pool of resident neural stem cells (NSCs) that can differentiate into all neural cell types. To advance our understanding of NSC function and to develop therapeutic and diagnostic approaches, it is important to accurately identify and enrich for NSCs. There are no definitive markers for the identification and enrichment of NSCs present in the mouse brain. Recently, a fluorescent rosamine dye, CDy1, has been identified as a label for pluripotency in cultured human embryonic and induced pluripotent stem cells. As similar cellular characteristics may enable the uptake and retention of CDy1 by other stem cell populations, we hypothesized that this dye may also enrich for primary NSCs from the mouse brain. Because the subventricular zone (SVZ) and the hippocampus represent brain regions that are highly enriched for NSCs in adult mammals, we sampled cells from these areas to test this hypothesis. These experiments revealed that CDy1 staining indeed allows for enrichment and selection of all neurosphere-forming cells from both the SVZ and the hippocampus. We next examined the effectiveness of CDy1 to select for NSCs derived from the SVZ of aged animals, where the total pool of NSCs present is significantly lower than in young animals. We found that CDy1 effectively labels the NSCs in adult and aged animals as assessed by the neurosphere assay and reflects the numbers of NSCs present in aged animals. CDy1, therefore, appears to be a novel marker for enrichment of NSCs in primary brain tissue preparations.
Introduction
N
Despite a large body of studies showing various methods for enrichment of NSCs, there is no definitive marker that incorporates all NSCs in the neurosphere-forming assays. Many [1,2] utilize Prominin-1 (CD133), with reports [3] of very high enrichment for SVZ NSCs, while other surface-labeling approaches [4 –7] also show efficacy. Indeed, in addition to CD133, CD15 used either alone [8] or in combination with CD184 [9] leads to enrichment. However, CD133-negative cell fractions can also give rise to neurospheres, which is indicative of NSC presence [10]. As antibody selections do not capture the complete population of NSCs present, other protocols have been developed. Two intracellular labeling protocols also enrich for NSCs, the first based on variations in the aldehyde dehydrogenase (ALDH) levels detected using the fluorescent label Aldefluor™ [11]. The second [12] relies on the differential uptake of the fluorescent dye Hoechst 33342 resulting in a discrete side population of cells (SP cells) when analyzed by flow cytometry. Sorted SP cells have been shown to enrich for NSCs [13]. However, separation of SP cells again does not specifically select for NSCs [6]. Both approaches have been combined with surface antibody binding to improve NSC-selective pressure. Given the variability reported in the literature, additional probes for defining the NSC population in primary tissue are critically important.
In a recent study, Kang and colleagues [14] systematically screened 280 modified fluorescent rosamine compounds and discovered a pluripotency-identifying fluorescent molecule, CDy1. In tissue culture, cell lines of either mouse or human embryonic stem cells (ESCs), as well as human induced pluripotent stem (iPS) cells, CDy1 specifically selected for pluripotency [14]. As similar cellular characteristics may enable the uptake and retention of CDy1 by other stem cell populations in culture, we hypothesized that this dye may also enrich for primary NSCs from the mouse brain. Because the SVZ represents the zone most enriched for NSCs in the adult mammalian brain, we primarily sampled the cells from this area to test our hypothesis. We also examined the effectiveness of CDy1 to select for NSCs derived from the aged SVZ of aged animals, where the total pool of NSCs present is significantly lower than in younger animals, along with its ability to label hippocampal NSCs.
Materials and Methods
Animals
Mice used in this study were adult C57BL6J females, 6 weeks, 3 months, or 12 months of age. All experiments were conducted in accordance with the Australian Code of Practice for the Care and Use of Animals for Scientific Purposes, with approval from the University of Queensland Animal Ethics Committee. Animals were maintained on a 12-h light/dark cycle with food and water provided ad libitum.
SVZ and hippocampal cell preparations
For each preparation, four mice were sacrificed by cervical dislocation; their brains immediately removed; and the SVZ and hippocampus dissected [15]. The adult SVZ and hippocampal tissue were digested by incubation in a mixture containing 0.1% papain (Worthington Biochemical Corporation) and 0.1% DNaseI (Roche) in Hank's buffered salt solution (Thermo Scientific) for 6 min at 37°C. After brief trituration, 2 mL of neurosphere growth medium was added and the cell suspension centrifuged at 250g for 5 min. The neurosphere growth medium contained the mouse NeuroCult™ NSC basal medium containing mouse NeuroCult™ NSC proliferation supplements (Stem Cell Technologies), 2% bovine serum albumin (Gibco/Invitrogen) and 2 μg/mL heparin (Sigma), 20 ng/mL purified mouse receptor-grade epidermal-like growth factor (EGF; BD Biosciences), and 10 ng/mL recombinant bovine fibroblast growth factor-2 (FGF-2; Roche). The pellet was resuspended in another 2 mL neurosphere growth medium and further mechanically triturated to obtain a single-cell suspension.
CDy1 staining
CDy1 was provided as a kind gift from Associate Professor Chang Young-Tae (Department of Chemistry, National University of Singapore). To label cells, CDy1 dye (0.1 μM) was added to the suspension incubated at 37°C for 1 h. The preparation was washed 3 times with 2 mL of culturing medium and incubated for 2 h at 37°C.
Neurosphere passaging and differentiation
For passaging, neurospheres were collected by centrifugation (100 rcf for 5 min) and incubated with 1 mL of 0.1% trypsin–EDTA for 2 min at room temperature, followed by washing with 1 mL of the trypsin inhibitor in HEM. Next, neurospheres were mechanically triturated until dissociated and replated in six-well plates at a density of 1×104 cells/cm2 in 3 mL of neurosphere growth medium as detailed above. Neurospheres were passaged every 7 days.
For differentiation, neurospheres were plated onto poly-
Flow cytometry
All experiments were performed using a BD Influx™ fluorescence-activated cell sorter using Sortware 1.0.1.6 software. Instrument settings were as follows: sheath pressure of 27.7 PSI, 100-μm nozzle, 49.50 KHz drop drive frequency, and sorting with a 1.0 drop pure sort mode resulting in a sort efficiency of always ≥85%. We used a 488 nm laser excitation at 145 mW power to excite CDy1-positive cells and a 528-38BP filter to collect emitted light. Dead cells were excluded using the viability indicator DRAQ7 (Biostatus) excited at 633 nm and 156 mW power with 750LP filter to collect emitted light. CDy1pos and CDy1neg cells were directly sorted into 24-well tissue culture plates (50,000 cells/well) based on the gates illustrated in Fig. 1C. SVZ neurospheres were cultured for 7 days; hippocampal neurospheres were cultured for 14 days at 37
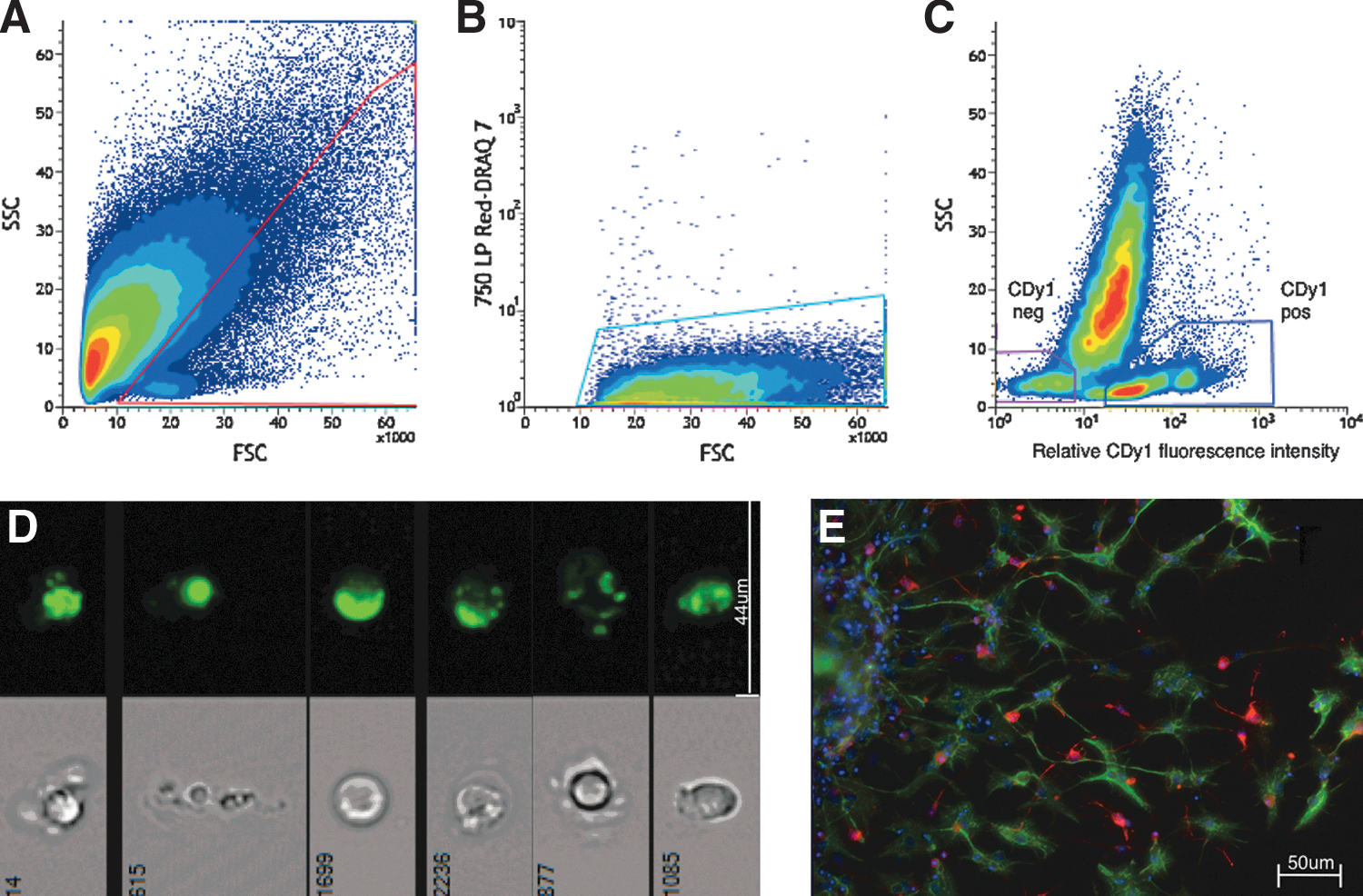
Flow cytometry plots illustrating the gating strategy employed and the resultant cell populations.
Statistical analysis
Statistical analysis was performed using GraphPad Prism (GraphPad Software, Version 5.0c). Data were analyzed using an unpaired two-tailed Student's t-test. Values are expressed as mean±standard error of the mean with significance determined at P<0.05.
Results
Cdy1 labels a subpopulation of cells in primary tissue from the adult mouse SVZ and hippocampus
Recently published findings have shown that CDy1 selectively stains mouse and human ESCs and human iPSCs grown in culture. As dissociated primary brain tissue is characterized by large amounts of cellular debris that impedes identification of NSCs, we sought to establish a protocol to effectively select for putative NSCs using scattered-light characteristics and CDy1 fluorescence. To that end, we designed a flow cytometry gating strategy that allowed clear separation of CDy1-positive (CDy1pos) and CDy1-negative (CDy1neg) events from single-cell preparations of the adult mouse SVZ and hippocampus (Fig. 1A–C). Dead cells and cellular debris were excluded by modifying a previously published gating strategy for neural tissue [16] and identifying dead cells using DRAQ7 dye. Of all the live cells (i.e., DRAQ7neg cells), CDy1pos cells constitute 10.56%±0.81% (n=8) of SVZ cells and 7.26%±1.10% (n=4) of hippocampal cells; labeled cells exhibited punctate fluorescence throughout the cytoplasm when visualized using AMNIS imaging cytometry (Fig. 1D) and fluorescence microscopy (data not shown).
CDy1 enriches for neurosphere-forming cells from the adult mouse brain
Having established that CDy1 labeled a population of brain cells, to assess the suitability of CDy1 for identification of putative NSCs, we next investigated the performance of CDy1pos cells obtained from either the SVZ or hippocampus in the neurosphere assay. This assay allows for retrospective identification of NSCs based on their ability to form neurospheres (along with progenitor cells that have a more limited proliferative ability); NSCs make up ∼3% of neurosphere-forming cells in primary cultures [17]. NSCs can also differentiate into neurons, astrocytes, and oligodendrocytes in culture [18,19]. Cultured CDy1pos cells gave rise to a significantly greater number of neurospheres than CDy1neg cells (Fig. 2A). In fact, virtually no neurospheres were generated from the CDy1neg cell population. SVZ CDy1pos cells generated 6.50±0.83 neurospheres/1,000 events versus 0.04±0.02 neurospheres/1,000 events generated from CDy1neg cell population (n=8, P<0.001). As such, there were ∼162 times more neurospheres generated from the CDy1pos compared to CDy1neg population. For the CDy1pos fraction, 1 in ∼154 cells derived from the SVZ was a neurosphere-forming cell compared to only 1 in 25,000 of all CDy1neg cells, thereby demonstrating significant NSC enrichment by employing CDy1 as a selection marker for SVZ NSCs. Sustained proliferative ability of the primary SVZ neurospheres was confirmed through passaging (passage 1: 0.52×104±0.14×104 total cells; passage 2: 2.03×104±0.37×104 total cells; n=3, P<0.05). In addition, some of the differentiated neurospheres gave rise to cells of the neuronal and glial phenotype (Fig. 1E).

Comparison of the neurosphere-forming frequency of CDy1-selected cells from the subventricular zone of the mice brains.
CDy1 staining also allowed for enrichment of neurosphere-forming cells from the hippocampus. CDy1pos cells derived from the hippocampus generated 0.45±0.07 neurospheres per 1,000 events (n=3), whereas no neurospheres were obtained from the Cdy1neg population.
In summary, CDy1 staining allowed for enrichment and selection of all neurosphere-forming cells from the SVZ and the hippocampus.
CDy1 selects for neurosphere-forming cells from the SVZ of young and aged animals
Having established that labeling-dissociated cell suspensions from the adult mouse brain with CDy1 allows the enrichment of neurosphere-forming cells, we then examined whether the same approach could be applied to the young and aged brains. This approach allowed us to test the robustness of CDy1 as a marker to isolate the cells of interest, that is, NSCs, as previous reports indicated that effectiveness of some previously used markers can be affected by the physiological status [20].
We found the highest percentage of CDy1pos cells in the young brains and lowest in the aged brains, which is consistent with previous studies that have reported an age-related decrease in the total number of neurospheres [21]. Specifically, in comparison to the adult SVZ (3-month-old: 10.56%±0.81%, n=8), the percentage of CDy1pos cells was significantly greater in the young SVZ (6 weeks: 24.53%±5.13%, n=3, P<0.01), while lower in the aged SVZ (12-month-old: 4.85%±0.25%, n=3, P<0.01) (Fig. 2B
Discussion
NSC enrichment
NSCs are retrospectively identified by their ability to form passagable neurospheres in culture that can generate large numbers of progeny and differentiate into neurons, astrocytes, and oligodendrocytes. While a recent study [22] has focused on antibody panels to identify NSCs, intracellular labeling approaches such as Aldefluor that are not reliant on antibody recognition of cell surface binding sites have received far less attention. As there is no a priori single surface marker combination, or intracellular marker that defines NSCs in primary tissue, any additional markers that enrich for NSCs are beneficial. In this context, CDy1 staining represents a useful marker for the selection of NSCs. We show that CDy1 can be used to selectively stain a subpopulation of cells dissociated from primary tissue removed from the SVZ and the hippocampus of the mouse brain. The CDy1 staining approach selectively targets all NSCs within the SVZ as assessed by their ability to form neurospheres, a defining characteristic of NSCs.
To the best of our knowledge, the fluorescent ALDH substrate Aldefluor, SP staining, and now CDy1 are the only nontransgenic, nonsurface labeling methods that enrich for NSCs from young, adult, and aged primary tissue. Interestingly, both the Aldefluor and CDy1 methods rely on the identification of a low side-scattered light and intracellular stained positive population [11]. However, CDy1 appears not to target the ALDH present in stem cells, at least based on evidence from other cell types [23].
While our data indicate that CDy1 selects for all neurosphere-forming cells present in the SVZ, the ratio of these putative stem cells relative to the total CDy1-stained positive cell population remained low at ∼1 in 200 cells. This indicates that CDy1 loads into both stem and non-stem cells. A significant population of cells, characterized by relatively high side-scattered light, (Fig. 1C), showed variable uptake and retention of CDy1. Sorting and subsequent analysis of these cells revealed that this population did not contain any neurosphere-forming cells (data not shown).
Microscopy investigation at 1 week after sorting of CDy1pos cells from the SVZ showed no CDy1 labeling in neurospheres, indicating that the dye is not retained or rapidly photobleaches under culture conditions. Directly poststaining, investigation of CDy1pos cells showed that the dye binds to the yet-unidentified locations with a punctate distribution in the cytoplasm, suggesting its localization to the organelles. Rhodamine dyes have a propensity to label mitochondria [24], and as rosamine dyes are rhodamines without 2′-carboxylic acids, they may also localize to similar locations and exhibit the cytoplasmic fluorescence distributions that were observed in the present study. Further studies are planned to investigate organelle labeling, specifically the role mitochondria play in the uptake and retention of CDy1, and whether the latter may be an NSC characteristic as shown in brain tumors [25].
Having identified a putative NSC population using CDy1, we sought to confirm whether the dye effectively selected for NSCs in both young and aged mice. Our results indicate that this was indeed the case with an approximate reduction of 50% of total SVZ cells positive for CDy1 in aged mice compared to younger specimens, which is consistent with previously published data. Specifically, Blackmore and colleagues [21] also reported a 50% reduction in the number of neurospheres obtained from the SVZ of 12-month-old mice compared to the 6 to 8-week old mice. The frequency of neurosphere formation from CDy1pos cells did not change between young and old mice, and therefore a reduction of the total number of neurospheres formed in aged mice would similarly be ∼50%.
In light of these results, CDy1 can be considered a useful marker for enrichment of putative NSCs. While this study has focused on the use of CDy1 in the absence of the other markers known to enrich for NSCs, further studies combining cell surface markers and CDy1 could lead to isolation of pure NSC populations and lead to novel insights into the biology of these cells.
Footnotes
Acknowledgments
This study was partially funded by a National Health and Medical Research Council program grant (PFB) and the estate of Dr Clem Jones AO. JV was supported by a Smart Futures Fellowship from the Queensland Government. The authors thank the staff of the University of Queensland Biological Resources facility for breeding and maintaining the animals used in this study. We also thank Gregory Robinson for technical assistance.
Author Disclosure Statement
The authors state that no competing financial interests exist.