
Abstract
Recently, a cohort of miRNAs, including miR-31, was reported to be downregulated during osteogenic induction by miR microarray analysis. It remains unclear how changes in miR-31 expression collaborate with bone transcription factors to activate the biological pathways that regulate the differentiation of bone mesenchymal stem cells (BMSCs). Here the effects of miR-31, Runx2, and Satb2 on the osteogenic differentiation of BMSCs were investigated using mimics and inhibitors of miR-31, small interfering RNA for knockdown of Runx2 and plasmids for overexpression of Runx2. Our results showed that miR-31 expression decreased progressively in BMSC cultures during differentiation. Inhibition of miR-31 dramatically increased the alkaline phosphatase activity and mineralization in BMSC cultures. Additionally, miR-31 diminished the levels of the Satb2 protein without significantly affecting Satb2 mRNA levels, and Runx2 directly repressed miR-31 expression. Overexpression of miR-31 significantly reduced expression of the osteogenic transcription factors OPN, BSP, OSX, and OCN, but not Runx2. Furthermore, the high expression of miR-31 in BMSCs cultured in the proliferation medium repressed Satb2 protein levels, which may contribute to the maintenance of BMSCs in an undifferentiated state. In conclusion, our results suggest that a Runx2, Satb2, and miR-31 regulatory mechanism may play an important role in inducing BMSC osteogenic differentiation. The results of this study provide us with a better understanding of the molecular mechanisms that govern the BMSC fate.
Introduction
M
Bone mesenchymal stem cells (BMSCs) have the potential to undergo multilineage differentiation into multiple tissue cell types, such as osteoblasts, chondrocytes, adipocytes, and myoblasts [5 –8]. Runx2 has been identified as an early key transcription factor regulating osteogenesis, whereas Satb2, regulated by Runx2, has also been demonstrated to be an important transcription factor in the later stage of osteogenesis [9 –11]. The low expression of these regulatory factors in undifferentiated BMSCs maintains the differentiation potential. The regulation of key transcription factors, such as Runx2 or Satb2, may affect osteogenic differentiation [12,13].
Recent miR microarray analysis had revealed a cohort of miRs that function in regulating osteoblast differentiation and are downregulated during osteogenic induction [14 –18]. One of these miRs, endogenous miR-31, is involved in tumor cell migration and invasion and is expressed at a low expression level during the induction of osteogenic differentiation [19]. However, the mechanism by which miR-31 expression is negatively regulated during osteogenesis remains unclear. A previous study has confirmed that Runx2 regulates the expression of osteogenesis-associated miRNAs [15,20]. Moreover, the moderation of tumor cell migration and invasion by Satb2 was shown to be impaired by the overexpression of miR-31 in cancer-associated fibroblasts [21]. However, the role of miR-31 and its target genes in the regulation of BMSC differentiation has not been well characterized, and the mechanism governing miR-31 regulation of Satb2 is poorly understood. Furthermore, knockout of miR-31 may lead to upregulation of Satb2 and thereby promote osteogenesis.
Here, to define the regulatory mechanisms of miR-31 in the osteogenic differentiation of BMSCs, we compared the expression of miR-31 between undifferentiated and osteogenically differentiated BMSC cultures. The effect of miR-31 on the differentiation of BMSCs was explored by using miR-31 mimics as well as inhibitors of miR-31. To determine whether Runx2 and Satb2 regulated miR-31 expression, BMSCs were treated with small-interfering RNA (siRunx2) as well as an overexpression plasmid vector (p-Runx2-wt). Our data demonstrated that a regulatory loop consisting of Runx2, Satb2, and miR-31 regulates the osteogenic differentiation of BMSCs. This study provides us with a better understanding of the molecular mechanisms that govern the proliferation and differentiation of BMSCs.
Materials and Methods
Cell culture
Bone marrow was isolated from the femur and tibia of 4-week-old male Sprague-Dawley rats as previously described [22]. Primary BMSCs were incubated in a humidified 5% CO2 atmosphere at 37°C. For proliferation assays, the proliferation medium (PM) consisted of the α-minimum essential medium (Gibco) supplemented with 10% fetal bovine serum (FBS, Gibco), 100 units/mL penicillin, and 100 μg/mL streptomycin. For osteogenic induction, the differentiation medium (DM) consisted of the α-MEM supplemented with 10% FBS, 10−8 M dexamethasone, 10 mM β-glycerol phosphate, and 50 mg/mL L-ascorbic acid (Sigma).
The murine preosteoblast cell line MC3T3-E1 clone 4 and premyogenic C2C12 cells were obtained from the Chinese Academy of Sciences Cell Bank and were maintained in the regular growth medium (DMEM, including 10% FBS and 1% antibiotics).
Transfection of miRNA mimics and small-interfering RNAs
The miR-31 oligonucleotides (mimics, inhibitors, and negative control) and small-interfering RNAs (siRunx2, siSatb2, and negative control) were synthesized by GenePharma Co., Ltd. For transient transfections, 40 nm miR-31 mimics, inhibitors, miR-negative control (miR-NC), or siRNA were mixed with the X-treme GENE HP DNA transfection reagent (Roche) in the Opti-MEM medium, and the mixture was directly added to cells in six-well plates at a density of 3×105 cells/well. The cells were harvested 48 h after transfection. For long-term detection, miR mimics or inhibitors were repeatedly transfected every 3 days. The oligonucleotide sequences of siRunx2 and siSatb2 were as follows: siRunx2 si1 5′-UGCCUCUGCUGUUAUGAAA-3′, si2 5′-GGUUCAACGAUCUGAGAU-3′; and siSatb2 si1 5′-GAACCUUUCAGACUAUUGU-3′, si2 5′-CCUCUAUCCUGACCAAGAA-3′.
Construction of Runx2 expression vectors
Total RNA extracted from MC3T3-E1 cells was used to generate cDNA by reverse transcript polymerase chain reaction (PCR) using the PrimeScript™ PT reagent kit (TaKaRa). cDNA of PCR products was confirmed by DNA sequencing. After digestion with BglII and NotI (Promega) and agarose gel purification, PCR products were subcloned into pCMV-myc. The primer sequences and vector construction procedure have been described previously [18]. The Runx2 plasmid DNA (p-Runx2-wt) was transfected into cells using the X-treme GENE HP DNA transfection reagent (Roche).
Quantitative real-time PCR analysis
Total RNA was extracted from the cultured cells with Trizol (Invitrogen), and first-strand complementary cDNA was synthesized using the PrimeScript™ RT reagent kit (TaKaRa). qPCR was performed as previously described [23]. Briefly, qPCR was performed using Power SYBR Green PCR Master Mix (Applied Biosystems) in combination with a 7500 Real-Time PCR Detection System. The gene-specific primers are shown in Table 1. The mature miR-31 expression levels were examined using a miRcute miRNA qPCR detection kit (Tiangen Biotech). The relative mRNA or miRNA level was expressed as fold change relative to untreated controls after normalization to the expression of GAPDH or U6, respectively.
Northern blot analysis
Total RNA was extracted from BMSCs with the Trizol reagent (Invitrogen). Northern blotting was performed as previously described [2, 20]. About 30 μg of total RNA samples were run on 12% acrylamide denaturing (urea) gels, and then transferred to a Hybond-N+ nylon membrane (Amersham Biosciences) by electrophoresis using a semidry transfer cell (Bio-Rad). Hybridization was performed according to a standard protocol. Digoxin-labeled oligonucleotide probes complementary to the mature miR-31 were used in the hybridization. The probes were 5′-CAGCTATGCCAGCATCTTGCCT-3′ for miR-31 and 5′-ATATGGAACGCTTCACGAATT-3′ for U6.
Western blot analysis
Protein lysates were generated using the radioimmunoprecipitation assay lysis buffer (Thermo Fisher Scientific), and the protein concentration was determined using a BCA protein assay (Thermo Fisher Scientific). Proteins were separated on a 10% SDS-PAGE gel and transferred to polyvinylidene fluoride membranes (Millipore) as previously described [24]. After blocking with 5% nonfat milk, the membranes was incubated with primary antibodies for Runx2, Satb2, OPN, OCN, and Actin (all from Abcam), and then they were incubated with the HRP-conjugated anti-rabbit secondary antibody (Sigma). Immunoblots were visualized by Odyssey V3.0 image scanning. The densitometric intensities of the individual protein bands were quantified using Bandscan 5.0 software, and values were normalized to the β-actin values for each sample.
Chromatin immunoprecipitation
BMSCs were crosslinked with 1% formaldehyde (Sigma) at 37°C for 15 min once it reached to 90% confluence. The cell lysis, immunoprecipitation of DNA-protein complexes, and the reverse crosslink of formaldehyde crosslinked DNA were performed with a chromatin immunoprecipitation (ChIP) assay kit (Upstate) according to the manufacturer's protocol. The DNA-chromatin complexes were immunoprecipitated with anti-Runx2 (Santa Cruz Bio-technology, Inc.), no antibody, or normal mouse IgG (Santa Cruz Bio-technology, Inc.) as an internal control. The precipitated DNA was analyzed by PCR. Based on the sequence, ACCACA was the core binding site for Runx2 as recently published [20], 2 kb of the promoter of the miR-31 was examined, and the ACCACA sequence was found and located 156/151 nucleotides upstream from the transcription start sites (TSS) of the miR-31 (Fig. 5A). The primers used for this analysis were as follows: primer-A: forward (5′-TGCTATCTGCAGTACTGTGCTGAGG-3′), reverse (5′-AGAGAGGCTGTGGTTAGTTCCTGCT-3′), and primer-B: forward (5′-ACTGCCTTGACTTCCTGCCTTGGTG-3′), reverse (5′-TAGCTAGGTCTGTACCCTGTCTCAG-3′).
Alkaline phosphatase and Alizarin Red staining
Osteoblast differentiation was identified by alkaline phosphatase (ALP) staining and Alizarin Red staining (ARS). BMSCs were cultured in six-well plates at 3×105 cells/well. The cells were fixed with 95% ethanol (v/v) after 7 days, and ALP staining was performed according to the manufacturer's instructions (Rainbow). For the determination of the ALP activity, BMSCs were seeded in 24-well plates and maintained in the differentiation culture medium for 3 days after transfection. The ALP activity was determined at 405 nm using p-nitrophenylphosphate (p-NPP; Sigma) as the substrate, as described previously [25]. All experiments were conducted in triplicate. For the detection of calcium nodule formation during differentiation, BMSCs were transfected, and after 14 days, cells were fixed in cold 70% ethanol for 1 h, and then incubated with a staining solution (Sigma) for 20 min at room temperature, The stain was desorbed with 10% cetylpyridinium chloride (Sigma) for 1 h [26]. The solution was collected and distributed at 100 μL per well on 96-well plates for absorbance reading at 590 nm with a microplate reader (Bio-Tek). Finally, ALP and ARS levels were normalized to the total protein content. Phosphate-buffered saline was used for washing and as the negative control.
Bioinformatics analysis
To identify the target genes of miR-31 in osteogenesis, we searched for candidate genes using three miRNA target prediction databases: TargetScan (
Statistical analysis
The experimental statistics presented in this study are expressed as the mean±standard derivation (SD). All experiments were performed at least three times unless otherwise specified. Data were analyzed using the Student's two-tailed t test to compare the means of two groups or a one-way ANOVA for comparison of the means of more than two groups using SPSS Statistics 17.0 software. P<0.05 was considered statistically significant.
Results
Expression levels of miR-31 during osteogenic differentiation
The qPCR analysis results showed that endogenous miR-31 expression decreased gradually during BMSC osteogenic induction (Fig. 1A). After being cultured in the DM for 12 h, the expression of miR-31 in BMSCs was significantly decreased (P<0.05), and it decreased further when the duration of the differentiation was prolonged (P<0.05). The decreasing miR-31 levels during osteogenic differentiation were further confirmed by Northern blotting analysis (Fig. 1B). However, no significant change in miR-31 expression was detected in BMSC cultures maintained in proliferation conditions (P>0.05).

miR-31 expression and its effect on Satb2 levels in the proliferation and differentiation of bone mesenchymal stem cells (BMSCs).
Effects of miR-31 on Satb2 expression in BMSCs
The levels of the Satb2 protein were negatively regulated by miR-31 without any significant effect on Satb2 mRNA levels. Target gene prediction showed that there were two binding sites for miR-31 in the 3′-UTR region of Satb2 mRNA (Fig. 1C). BMSCs cultured in the PM were transfected with miR-31 mimics or inhibitors. After 48 h, western blot analysis (Fig. 1D) demonstrated that the protein level of Satb2 was repressed in cells transfected with miR-31 mimics. In contrast, Satb2 protein expression was dramatically upregulated by miR-31 inhibitors (P<0.05, Fig. 1E). Most importantly, in cells transfected with miR-31 inhibitors, the Satb2 protein level was increased nearly four-fold in comparison to the control cells, indicating that miR-31 is an important negative regulator of Satb2. Similarly, Satb2 protein levels also showed a reciprocal relationship with miR-31 levels during osteogenic induction of BMSCs maintained in the DM (Fig. 1D, E). However, the mRNA levels of Satb2 were not significantly affected by either miR-31 knockdown or overexpression during the proliferation and differentiation of BMSCs (Fig. 1F). Therefore, these results suggest that miR-31 negatively regulates Satb2 by targeting translational control.
Inhibition of miR-31 enhances osteogenic differentiation
The miR-31 inhibitors enhanced osteogenic differentiation of BMSCs. The protein levels of the osteogenic transcription factors bone sialoprotein (BSP) and osteopontin (OPN) were examined in BMSCs cultured in the DM for 2 days. Our results indicated that the protein levels of BSP and OPN dramatically decreased in BMSC cultures transfected with miR-31 mimics (P<0.05) (Fig. 2A). The levels of these two proteins exhibited an approximately three-fold increase in the miR-31 knockout cells compared to control cells (Fig. 2B). In transfected cells, the levels of BSP and OPN mRNA displayed a similar pattern to that of these two proteins in four groups (Fig. 2C), indicating that the BSP and OPN expression were not directly regulated by miR-31 at translational control. Thus, the promotion of osteogenic differentiation via miR-31 inhibition may depend on upregulation of bone transcription factors, and the high expression of miR-31 in BMSCs may attenuate osteogenic differentiation and help to sustain the cells' undifferentiated state.

Inhibition of miR-31 enhances osteoinductive differentiation.
ALP mRNA expression was reduced by transfection with miR-31 mimics and was promoted by miR-31 inhibitors (P<0.05, Fig. 2C). As shown in Fig. 2D, The semiquantitative analysis of the ALP activity showed that the absorbance index values in miR-31 overexpressing cells were significantly suppressed compared to non-miR-31 transfected cells (P<0.05), but increased by miR-31 inhibitors (P<0.05) at day 7. The semiquantitative analysis of the ARS results showed that the absorbance index values from miR-31 inhibitor transduced groups were 1.7-fold higher than the control group at day 14 (Fig. 2D). At day 7, cells transfected with miR-31 inhibitors showed robust positive ALP staining. In addition, the mineralization in BMSC cultures at day 14, as exhibited by ARS, was compatible with the ALP staining results (Fig. 3). These results suggest that miR-31 acts as an important negative regulator of BMSC osteogenesis.

ALP and ARS staining. ALP expression was clearly enhanced by the inhibition of miR-31 at day 7. ARS staining on day 14 revealed a significant increase in calcium deposition in BMSCs transfected with miR-31 inhibitors.
Runx2 represses endogenous miR-31 expression
Expression of miR-31 was negatively correlated with Runx2 expression levels. As shown in Fig. 4A, the expression of the Runx2 protein in the BMSC cultures was decreased by transfection with siRunx2, but was upregulated nearly four-fold by transfection with a Runx2 overexpression vector (P<0.05) (Fig. 4B). The expression of miR-31 in BMSCs can be significantly increased by siRunx2 (P<0.05) or prominently repressed by Runx2 overexpression (P<0.05), but no obvious changes were observed in cotransfected cells relative to nontransfected cells (P>0.05) (Fig. 4C). In summary, during osteoblast differentiation of BMSCs, there was a reciprocal relationship between Runx2 protein levels and miR-31 levels, with the highest expression of the Runx2 protein correlating with the lowest level of miR-31. As stated before, Runx2 is an upstream regulatory factor of Satb2, and the levels of Satb2 mRNA matched the pattern of the Runx2 protein in cells, where Runx2 was knocked down or overexpressed (Fig. 4B, D). The levels of miR-31 were upregulated in MC3T3 and C2C12 cells when Runx2 protein levels were decreased by siRunx2 transfection (P<0.05, Fig. 4E, F).

Runx2 represses miR-31 expression.
To verify whether Runx2 could physically bind with the promoter of miR-31 in BMSCs, ChIP assays were performed. Primer A (−323/−143) that spans Runx2 potential binding sites (−156/151) in the promoter was used for PCR, and primer B (−1434/−1300) was used as negative controls (Fig. 5A). The antibodies against Runx2 could specifically immunoprecipitate the DNA fragment containing primer A (Fig. 5B, lane 6). These findings confirmed that Runx2 directly bound to the miR-31 promoter. Collectively, a Runx2 regulatory element directly negatively regulates the expression of miR-31 during osteoblast differentiation.

Analysis of Runx2 binding to the promoter of miR-31.
miR-31 attenuates BMSC differentiation without affecting Runx2 levels
The expression level of miR-31 dramatically affected the expression of osteogenic transcription factors, with the exception of Runx2. Runx2 protein expression was not suppressed by miR-31 mimic transfection or increased by miR-31 inhibitor transfection (P>0.05, Fig. 6A, 6B), and these results were supported by an analysis of Runx2 mRNA levels (P>0.05, Fig. 6C). Conversely, the miR-31 mimics significantly reduced the mRNA and protein levels of the bone-specific markers osterix (OSX) and osteocalcin (OCN) (P<0.05, Fig. 6D, E). In addition, to determine whether Satb2 inhibited miR-31 expression, BMSCs were transfected with Satb2 small interfering RNA (siSatb2) (Fig. 6F, G). Unlike the effects of siRunx2 (Fig. 4A, C), no obvious changes in miR-31 expression were observed in Satb2 knockdown BMSCs (P>0.05, Fig. 6H). Taken together, these results suggest that miR-31 does not regulate Runx2 expression, and miR-31 expression is unaffected by the expression levels of Satb2.
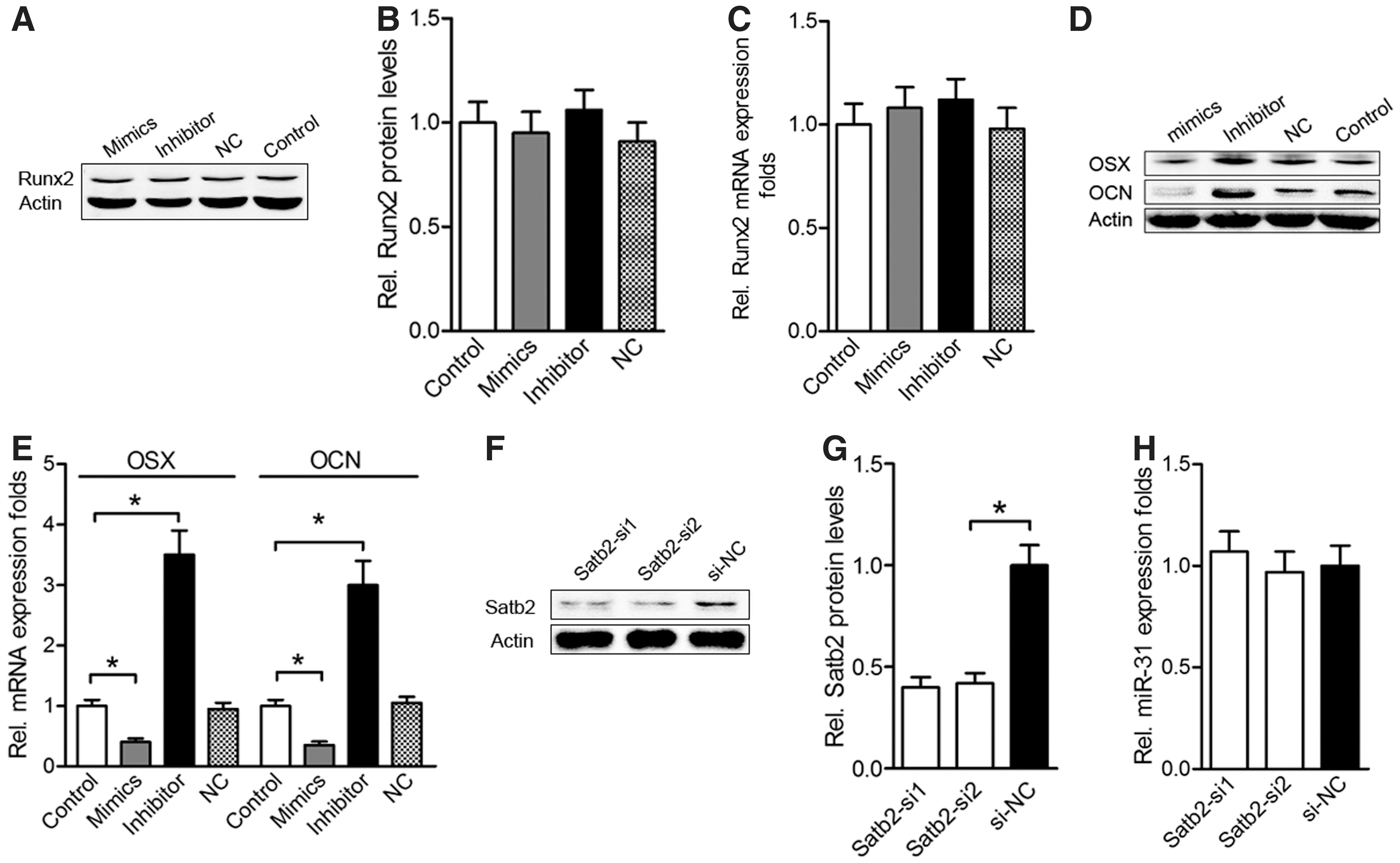
miR-31 attenuates BMSC differentiation without affecting Runx2 levels.
Discussion
Our data uncovered a regulatory loop in osteoblastic differentiation consisting of Runx2, Satb2, and miR-31. The miR-31 regulated translational control of Satb2 by directly targeting the 3'UTR region of Satb2 mRNA, and this was confirmed by a previous luciferase reporter assay [21]. The promotion on osteogenic differentiation was enhanced by low expression of miR-31 during BMSC osteoinduction, and miR-31 expression was repressed by Runx2 overexpression. The inhibition of miR-31 significantly increased the expression of bone-specific markers and mineralization in BMSCs.
miR microarray analysis revealed a cohort of miRs that were downregulated during osteoblastic induction. These miRs usually target the 3'-UTR region of master bone transcription factors, negative regulating of protein expression via translational control and inhibiting osteoblastic differentiation [14,16,17,27]. However, it was unclear how these miRs were repressed during BMSC differentiation. Several studies have previously demonstrated that the expression of osteogenesis-related miRs is directly regulated by Runx2 [15,20]. A potential binding site (′5-ACCACA-3′) in the promoter region for Runx2 was confirmed by formerly published data [20]. In our studies, ChIP experiments confirmed that Runx2 directly binds to the promoter regions of the miR-31 (Fig. 5). Furthermore, overexpression of Runx2 attenuated miR-31 transcription, and knockdown of Runx2 expression increased miR-31 expression (Fig. 4). Therefore, Runx2 may be the master and direct regulator of endogenous miR-31 expression. Considering the low expression of Runx2 in undifferentiated BMSCs, the consequent high expression of miR-31 likely maintains BMSCs in an undifferentiated state.
Currently, Runx2 remains the earliest and most important transcriptional regulator of bone formation [28, 29]. The miR-133, miR-30, and miR-23∼24∼27 clusters were shown to directly target the 3′UTR region of Runx2 mRNA and inhibited or delayed osteocyte maturation in a mineralized matrix [13,15,18]. Previous study showed that Runx2 is the target for miR-31 in human BMSCs when it undergoes osteoblastic differentiation [19]. However, our results showed that the mRNA and protein levels of Runx2 were not affected by up- or downregulation of miR-31 levels in rat BMSCs, and this was supported by putative targets of miR-31 via TargetScan (
As a transcriptional regulator, Satb2 functions broadly and plays an important role in craniofacial patterning and bone development [10]. Other studies showed that overexpression of Satb2 can improve the expression of bone transcriptional factors (BSP, OPN, OSX, and OCN) and promote osteogenic differentiation [11,30]. Satb2 has been confirmed as a direct target of miR-31 by luciferase reporter assays [21], and the ability of miR-31 to modulate Satb2 protein levels without obviously influencing Satb2 mRNA levels has been demonstrated by our study. Previous study shows that Runx2 directly binds to the promoter of genes and regulates the expression of BSP, OPN, and OCN [31]. Our results showed that miR-31 regulated the mRNA and protein levels of BSP, OPN, OSX, and OCN, but Runx2, it may be noted that the expression of bone transcriptional factors (BSP, OPN, OSX, and OCN) were directly regulated by Satb2, but not directly by miR-31 or Runx2 in transfected cells. Importantly, the expression of transcriptional markers of bone differentiation and ALP activity were both increased by transfection of BMSCs with miR-31 inhibitors. Similarly to ALP staining, miR-31 knockout BMSCs showed robust positive staining for bone nodule formation during differentiation. The upregulation of Satb2 is one of the major mechanisms for promoting osteogenic differentiation in miR-31 knockout BMSCs.
In summary, these data allowed us to suggest a Runx2, Satb2, and miR-31 regulatory loop (Fig. 7). The role of miR-31 is to provide negative regulation of Satb2 at the post-transcriptional or translational level, and this attenuation is controlled precisely by Runx2 expression. The inhibition of miR-31 positively affected osteoblast differentiation by increasing Satb2 protein expression. This Runx2, Satb2, and miR-31 regulatory mechanism may have an important impact on the differentiation of BMSCs. Furthermore, the repression of Satb2 due to high expression of miR-31 may contribute to the maintenance of BMSCs in an undifferentiated state.

Graphic representation of the regulatory feedback loop between Runx2, Satb2, and miR-31. During osteoblast differentiation of BMSCs, the role of miR-31 is to provide negative post-transcriptional or translational regulation of Satb2. In turn, Runx2 plays an important role by repressing endogenous miR-31 expression and increasing the expression of Satb2.
Conclusion
In osteogenic differentiation, miR-31 negatively regulates BMSC differentiation by targeting translation of the master transcription factor Satb2. The inhibition of miR-31 showed a robust positive effect on osteoblast differentiation based on increased expression of bone-specific transcription factors. In contrast, Runx2 directly repressed endogenous miR-31 expression and encouraged the expression of Satb2, which may significantly induce BMSC differentiation. The results of this study provide us with a better understanding of the molecular mechanisms that govern the proliferation and differentiation of BMSCs.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (31271029, 81170876, 81000404, 81070737, and 81200720), the Education Commission of Shanghai (11YZ47), and the Outstanding Young Talents Training Plan of the Shanghai Health System (XYQ2011053).
Author Disclosure Statement
The authors report no conflict of interests. The authors alone are responsible for the content and writing of the article.