
Abstract
Mesenchymal stromal cells (MSCs) have been isolated from many tissues, including gestational tissue. To date, a study comparing the properties and suitability of these cells in cell-based therapies is lacking. In this study, we compared the phenotype, proliferation rate, migration, immunogenicity, and immunomodulatory capabilities of human MSCs derived from umbilical cord lining (CL-MSCs), umbilical cord blood (CB-MSCs), placenta (P-MSCs), and Wharton's jelly (WJ-MSCs). Differences were noted in differentiation, proliferation, and migration, with CL-MSCs showing the highest proliferation and migration rates resulting in prolonged survival in immunodeficient mice. Moreover, CL-MSCs showed a prolongation in survival in xenogeneic BALB/c mice, which was attributed to their ability to dampen TH1 and TH2 responses. Weaker human cellular immune responses were detected against CL-MSCs and P-MSCs, which were correlated with their lower HLA I expression. Furthermore, HLA II was upregulated less substantially by CL-MSCs and CB-MSCs after IFN-γ stimulation. MSC types did not differ in indolamine 2,3-dioxygenase (IDO) expression after IFN-γ stimulation. Despite their lower IDO, HLA-G, and TGF-β1 expression, only CL-MSCs were able to reduce the release of IFN-γ by lymphocytes in a mixed lymphocyte reaction. In summary, CL-MSCs showed the best characteristics for cell-based strategies, as they are hypo-immunogenic and show high proliferation and migration rates. In addition, these studies show for the first time that although immunomodulatory molecules HLA-G, HLA-E, and TGF-β play an important role in MSC immune evasion, basal and induced HLA expression seems to be decisive in determining the immunogenicity of MSCs.
Introduction
M
To date, most research on MSCs has concentrated on bone marrow-derived cells (BM-MSCs); however, they are not an ideal source for cellular therapy, as their isolation is associated with donor morbidity and, even more importantly, the number of MSCs found in the bone marrow and their regenerative potential significantly declines with age [18,19]. On the other hand, MSCs have also been isolated from extraembryonic gestational tissues such as the placenta [20] (P-MSC), umbilical cord lining [15] (CL-MSC), cord blood [21] (CB-MSC), and Wharton's jelly [22] (WJ-MSC). Due to the young chronological age of the donor, these cells are devoid of mutations and their isolation is noninvasive and so safe to both the infant and mother [23]. Furthermore, in our previous study, we showed that CL-MSCs were superior to BM-MSCs in their immunomodulatory capabilities and their hypo-immunogenicity [15]. In our present study, we compared the proliferation rates, migration capabilities, immunogenicity, and multiple immunomodulatory mechanisms exhibited by MSCs from four extraembryonic gestational tissues: CL-MSCs, CB-MSCs, P-MSCs, and WJ-MSCs. The results of this study have given insight into the mechanisms involved in MSC-mediated immunomodulation and more importantly, have allowed to identify the MSC type most suitable for cell-based therapies.
Materials and Methods
Extraembryonic gestational tissue-derived MSC isolation and morphology
CL-MSCs were isolated as previously described [24] and cultured in the PTT4 medium (CellResearch Corporation). CB-MSCs were isolated and cultured in the low-glucose DMEM+GlutaMAX-1 and Pyruvate supplemented with MSCGM SingleQuots (Lonza) [21]. P-MSCs were isolated as described by Barlow et al. and cultured in the low-glucose DMEM+GlutaMAX-1 and Pyruvate supplemented with 20% heat inactivated fetal calf serum and gentamicin [20]. WJ-MSCs were obtained from Thermo Scientific and cultured in the AdvanceSTEM Mesenchymal Stem Cell Basal Medium supplemented with AdvanceSTEM Mesenchymal Stem Cell Growth Supplement (Thermo Scientific) and 1% penicillin–streptomycin or the Human Mesenchymal Stem Cell Expansion Medium (CET) supplemented with 10 ng/mL recombinant human FGF (PeproTech). The low-glucose DMEM, fetal calf serum, and penicillin–streptomycin were all obtained from Gibco, Invitrogen. About 0.1% gelatin (Millipore)-coated culture flasks (SARSTEDT AG & Co.) were used for P-MSCs and WJ-MSCs.
All cells were cultured at 37°C in a humidified atmosphere of 95% air and 5% CO2 until 70%–80% confluency before trypsinization (TrypLE Express, Gibco, Invitrogen) or being removed by scraping with a cell scraper (Falcon BD Biosciences) (for FACS, MTS, and western blot analysis).
Scratch assay
MSCs were plated in 0.1% gelatin-coated (Millipore) six-well plates (Falcon; BD Biosciences) (1×106 cells/well). A scratch in the confluent cell monolayer was made with a pipette tip (P200) and after 6 h, cells were fixed with 10% PFA (paraformaldehyde) (Electron Microscopy Sciences) and washed with PBS (Gibco). Phase-contrast images were acquired using a Leica microscope and QWin acquisition software (Leica Microsysteme Vertrieb GmbH) (magnification 100× ). Migrated cells in 3–4 segments of the scratch area (300×500 μm each) were counted. Each analysis was performed 10 times.
Cell proliferation assay
Cells were seeded in a 96-well flat-bottom plate (Nunc) (4,000 cells/well) and incubated at 37°C in 5% humidified CO2. The medium was changed every second day and cell counts were quantified every day for 4 consecutive days using the 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium, (MTS) assay (CellTiter 96 Aqueous One Solution Cell Proliferation Assay; Promega). Absorbance at 490 nm was measured with the Magellan ELISA Reader and Software (Tecan Systems, Inc.).
Differentiation assays
For adipocyte and osteocyte differentiation, cells were plated in 24-well plates (6×104 cells/well) (Falcon, BD Biosciences) in a growth medium and incubated at 37°C in 5% humidified CO2. After 24 h, the culture medium was changed to the appropriate differentiation medium (PromoCell GmbH). For chondrocyte differentiation, cells were plated in 96-well suspension plates (SARSTEDT AG & Co) (1×105 cells/well in 200 μL). The differentiation medium was changed three times per week for 21 days or 14 days (adipocytes). Cells were stained for adipocyte (Oil Red O and hematoxylin counterstaining), chondrocyte (Toluidine Blue), and osteocyte (Alizarin Red S) differentiation as previously described [15]. Images were acquired using a Leica microscope and QWin acquisition software (Leica Microsysteme Vertrieb GmbH).
Flow cytometry
Cells were labeled with phycoerythrin (PE)-conjugated antibodies against human: CD31 (clone WM59), CD34 (clone 563), CD44 (clone G44-26C26), CD45 (clone H130), CD90 (clone 5E10), CD117/c-Kit (clone 104D2), HLA-ABC (clone DX17), TRA-1–60 (clone TRA-1–60) (all from BD Biosciences), HLA-DR+DP+DQ (clone WR18; Abcam), SSEA-4 (clone MCB13–70), and CD105 (clone 166707) (all R&D Systems). Unconjugated antibodies: HLA-E (clone MEM-E/06; Santa Cruz Biotechnologies) and HLA-G (clone MEM-G/9; Santa Cruz Biotechnologies) were visualized using PE-conjugated anti-mouse IgG1a or IgG2a (both Santa Cruz Biotechnologies). Mouse isotype matched antibodies IgG1, IgG2a, IgM (clones: MOPC-21, MOPC-173, and G155–228, respectively; all BD Biosciences), and IgG3 (clone 133316; R&D Systems) were used as controls. Data were acquired on a FACSCalibur flow cytometer using CellQuest Pro software (all BD Biosciences) and analyzed using FlowJo version 7.2.5 software (Tree Star). The results were expressed as folds of the isotype control. The data are shown as the mean from three independent experiments. Recombinant human IFN-γ (25 ng/ml; PeproTech) was added to the cell culture medium for 48 h before flow cytometry, where indicated.
Indolamine 2,3-dioxygenase detection
Cellular proteins were extracted using the RIPA buffer supplemented with Protease Inhibitor Cocktail according to the manufacturer's instructions (both Sigma-Aldrich). Protein concentrations were measured using the Pierce BCA Protein Assay Kit (Thermo Scientific). Ten microgram protein per well was loaded on a NuPAGE Novex 4%–12% Bis-Tris Gel and separated using the NuPAGE Bis-Tris Electrophoresis system (Invitrogen). Proteins were transferred onto a 0.2 μm PVDF membrane (Invitrogen), where indolamine 2,3-dioxygenase (IDO) (clone: H-110; Santa Cruz Biotechnologies) or GAPDH (clone: 14C10; Cell Signaling Technology) were detected using unconjugated antibodies. After an overnight incubation step, horseradish peroxidase-conjugated antibodies (Amersham Biosciences) were used to detect bound rabbit monoclonal antibodies. Antigens were identified with the ECL plus a western blotting detection kit (Amersham Biosciences) and imaged with the IVIS 200 system (Xenogen, Caliper Lifesystems). Blots were stripped with the restore western blot stripping buffer (Pierce, Thermo Scientific) before being reprobed for the housekeeping GAPDH. For IDO induction, 500 ng/mL recombinant human IFN-γ (PeproTech) was added to the cell culture medium for 48 h.
ELISA
Human IL-2, IL-10, and TGF-β1 were detected in cell culture supernatants using the BD OptEIA ELISA sets and BD OptEIA Reagent SET B according to the manufacturer's instructions (BD Biosciences). Cells were cultured at a density of 2.7×104 cells/cm2 for 48 h before the cell culture supernatant was collected and stored at −20°C. The data are shown as mean from three independent experiments, after normalization to their cell counts using the MTS assay and presented as (pg/mL)/OD.
Animals
Six- to 8-week-old male BALB/c mice were purchased from Charles River Laboratories and housed under specific pathogen-free conditions in the animal care facilities of the University Hamburg, and received humane care in compliance with University guidelines. Animals were randomly assigned to one of the four study groups and received either CL-MSC, CB-MSC, P-MSC, or WJ-MSC transplantations (n=6 per group) by intramuscular injection of 1.0×106 firefly luciferase (fLuc)-positive MSCs.
MSC lentiviral transduction
Cells were made to express fLuc under the CMV promoter by transducing with pLenti CMV Puro fLuc (w168-1) [25] (Addgene plasmid 17477). Transduction efficiency was enhanced by adding 8 μg/mL polybrene (Sigma-Aldrich Corporation) into the cell culture medium. Stable transfectants were enriched by puromycin (Santa Cruz Biotechnologies) selection at 0.08 μg/mL (CL-MSC, P-MSC, and WJ-MSC) or 0.5 μg/mL (CB-MSC). To estimate transduction efficiency, the cells were tested for the fLuc activity in a cell number titration assay. For bioluminescence imaging (BLI), 9.1 mg D-luciferin was dissolved in 10 mL PBS pH 7.4 (Gibco, Invitrogen) and added into each confluent cell flask.
Bioluminescence in vivo imaging
For BLI, D-luciferin firefly, potassium salt (375 mg/kg) (Biosynth AG) dissolved in PBS pH 7.4 (Gibco, Invitrogen) was injected intraperitonealy (200 μL per mouse) into anesthetized mice (2% isoflorane; Forene, Abbot GmbH & Co. KG). Animals were imaged using the IVIS 200 system (Xenogen). Region of interest (ROI) bioluminescence was quantified in units of maximum photons per second per centimeter square per steridian (p/s/cm2/sr). The maximum signal from an ROI was measured using Living Image 3.1 software (MediaCybernetics).
Rejection of MSCs in vivo
To test the rejection of MSCs, in vivo 1.0×106 fLuc-positive CL-MSCs, CB-MSCs, P-MSCs, or WJ-MSCs in 60 μL PBS (Gibco, Invitrogen) were injected into the hind limb muscle of immunocompetent BALB/c mice. FLuc expression was examined on days 0, 1, and thereafter, every second day until the signal dropped to background levels; n=6/group.
In vivo ELISPOT assay in BALB/c mice
To test the immunogenicity of MSCs in vivo, 1×106 MSCs in 60 μL PBS (Gibco, Invitrogen) were injected into the gastrocnemius muscle of BALB/c mice. Spleens were harvested after 5 days and 1×107 splenocytes were used as responder cells with 1×106 mitomycin C treated-MSCs (Sigma Aldrich) as stimulators, to detect anti-mouse IL-4 and IFN-γ in an ELISPOT assay (BD Biosciences).
In vitro human ELISPOT assay
To test the potential of MSCs to modify the immune response of human peripheral blood mononuclear cells (PBMCs), 5×106 Ficoll-isolated (Ficoll-Paque PLUS solution, Amersham Biosciences) PBMCs were used as responder cells with 1×105 MSCs. Anti-human IL-4 and IFN-γ responses were detected in a 4-day ELISPOT assay (BD Biosciences).
The immunosuppressive properties of MSCs in vitro were assessed by their potential to suppress a one-directional allogeneic mixed lymphocyte response in an allogeneic setting. A total of 5×106 allogeneic Ficoll-isolated (Amersham Biosciences) human PBMCs was used as responder cells with 5×105 mitomycin C-treated (Sigma Aldrich) PBMCs and 1×105 MSCs. Anti-human IFN-γ responses were detected in a 4-day ELISPOT assay (BD Biosciences). All ELISPOT spots were counted using an ELISPOT plate reader (CTL).
Statistical analysis
Data are presented as the mean±standard deviation. Comparisons were done by analysis of variance between groups with Fisher's least significant difference Post Hoc tests. Probability values (p) of less than 0.05 were considered significant. Statistical analysis was performed using the SPSS statistical software package 15.0 for Windows (SPSS, Inc.).
Results
Characterization of extraembryonic gestational tissue-derived MSCs
All four cell types adhered to plastic and were spindle shaped (Fig. 1A). To further verify that the cells were MSCs, we tested for the expression of typical MSC markers, including CD44, CD90, and CD105 (Supplementary Fig. S1A; Supplementary Data are available online at

Extraembryonic gestational tissue-derived MSC morphology, migratory, and proliferative potential.
To confirm multipotency of the cells, we tried to differentiate them into osteocytes, chondrocytes, and adipocytes (Supplementary Fig. S1B). In accordance with previous studies CB-MSC [27] and P-MSC [20] did not readily differentiate into cells of the adipogenic lineage. In our hands also, WJ-MSCs did not show adipogenic differentiation within the 3-week time frame.
In summary, our data support the notion that only CL-MSCs fulfill the criteria for MSCs according to the criteria of the International Society for Cellular Therapy [28].
Migratory and proliferative potential of extraembryonic gestational tissue-derived MSCs
We evaluated the migration capabilities of MSCs in a scratch assay. CL-MSCs were able to migrate into the scratch area the fastest (P<0.001 compared to CB-MSCs, P-MSCs, and WJ-MSCs) (Figs. 1B, C). CB-MSCs showed a slightly lower level of migratory potential, but were still faster than P-MSCs (P<0.001) and WJ-MSCs (P<0.001). P-MSCs and WJ-MSCs demonstrated the lowest migratory potential, with the least cells entering the scratch area after 6 h.
To investigate whether the MSCs differed in their proliferation potential, we compared them in a MTS assay. CL-MSCs showed a significantly higher proliferation rate than P-MSCs and WJ-MSCs (P<0.01; Fig. 1D).
Immunogenicity and survival of MSCs
The fLuc signal correlated with cell numbers after transduction (Supplementary Figs. S2 and S3). The ability of MSCs to evade a xenogeneic immune response in vivo was elucidated in immunocompetent BALB/c mice (Fig. 2A). FLuc positive CL-MSCs were detected up to 11 days after injection, while both P-MSCs and WJ-MSCs were rejected by day 7 (P<0.01 and P<0.01 for CL-MSCs versus P-MSCs and WJ-MSCs, respectively). The fLuc signal of CB-MSCs was detected until day 9 in BALB/c mice. To test whether the differences in MSC survival in immunocompetent mice were due to their ability to evoke T helper (TH)1 or TH2 responses, ELISPOT assays were performed 5 days after MSC injection into BALB/c mice (Figs. 2B, C). CL-MSCs evoked the weakest immune response of TH1 cells (IFN-γ: P<0.001, P<0.002, P<0.005 compared to CB-MSCs, P-MSCs, and WJ-MSCs, respectively), and TH2 (IL-4 assays: all P<0.001 compared to CB-MSCs, P-MSCs, and WJ-MSCs). CB-MSCs, P-MSCs, and WJ-MSCs evoked a similar cell-mediated immune response.
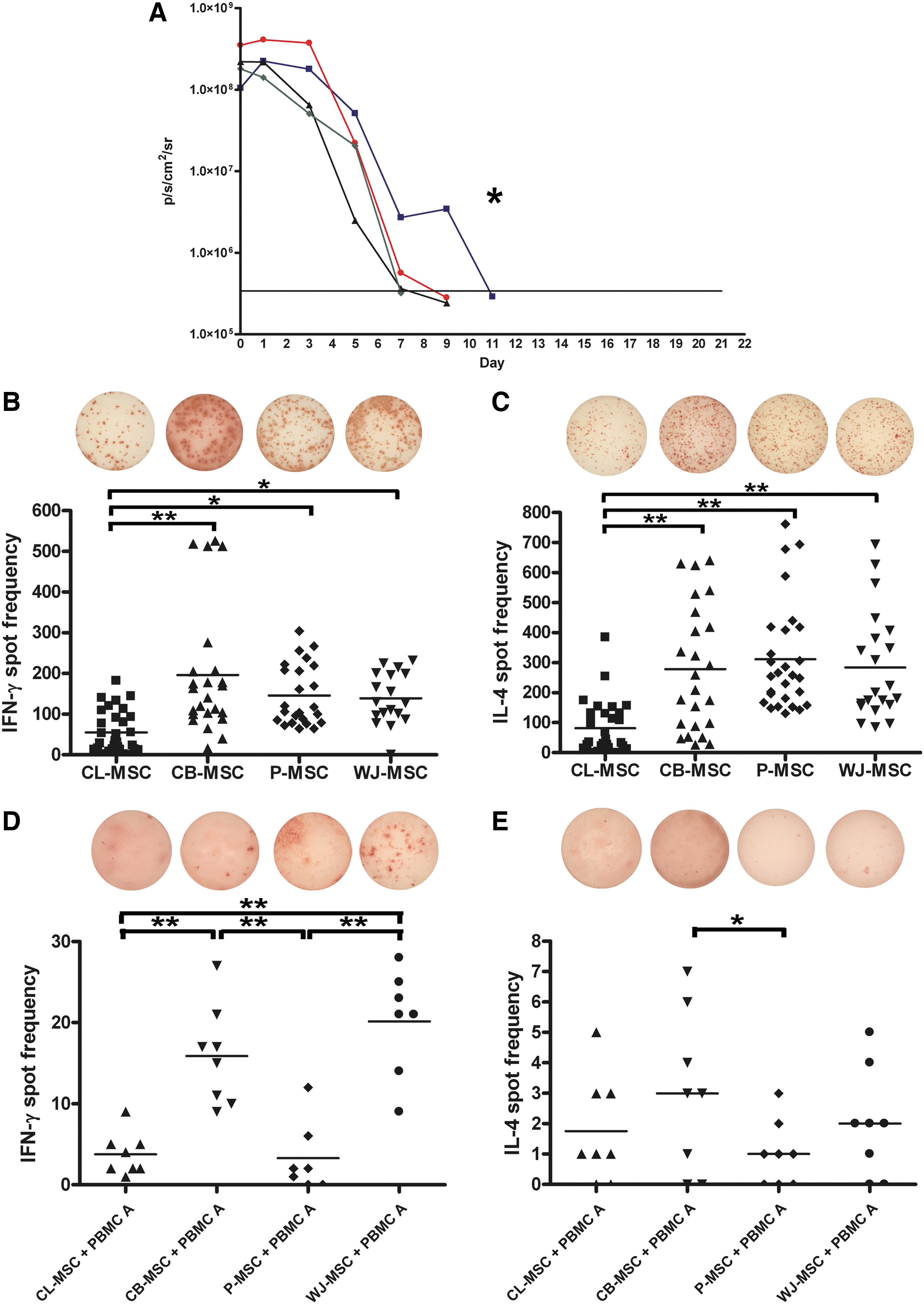
Survival and immunogenicity of extraembryonic gestational tissue-derived MSCs (n=6/group).
To further investigate the immunogenicity of MSCs, ELISPOT assays using human allogeneic PBMCs as responders were carried out. CB-MSCs and WJ-MSCs provoked the strongest cellular responses for TH1 cells (Fig. 2D) (both P<0.001). Interestingly, WJ-MSCs induced a significant TH1 immune response, despite having the least HLA mismatches with the PBMC responder A (Supplementary Table S1). Both CL-MSCs and P-MSCs showed a weak alloresponse only. The number of IL-4 secreting PBMCs was comparable among cell types and only differed between CB-MSCs and P-MSCs (P<0.05) (Fig. 2E).
Next, we investigated whether distinct immunogenicity might be related to HLA expression of the MSCs from different sources. In our study, native CL-MSCs and CB-MSCs expressed quite low, while P-MSCs and WJ-MSCs expressed a moderate level of HLA class I molecules on their surface (Fig. 3A). We also determined whether the presence of IFN-γ, associated with inflammation, increases the level of HLA class I expression, which would lead to an increase in MSC immunogenicity and ultimately make MSCs more susceptible to T-cell-mediated lysis. A 48-h stimulation period with a moderate concentration of IFN-γ lead to an increase in surface expression of HLA class I by all MSC types to quite similar levels, except P-MSCs (P<0.001, P<0.005, and P<0.02 for CL-MSCs, CB-MSCs, and WJ-MSCs, respectively).
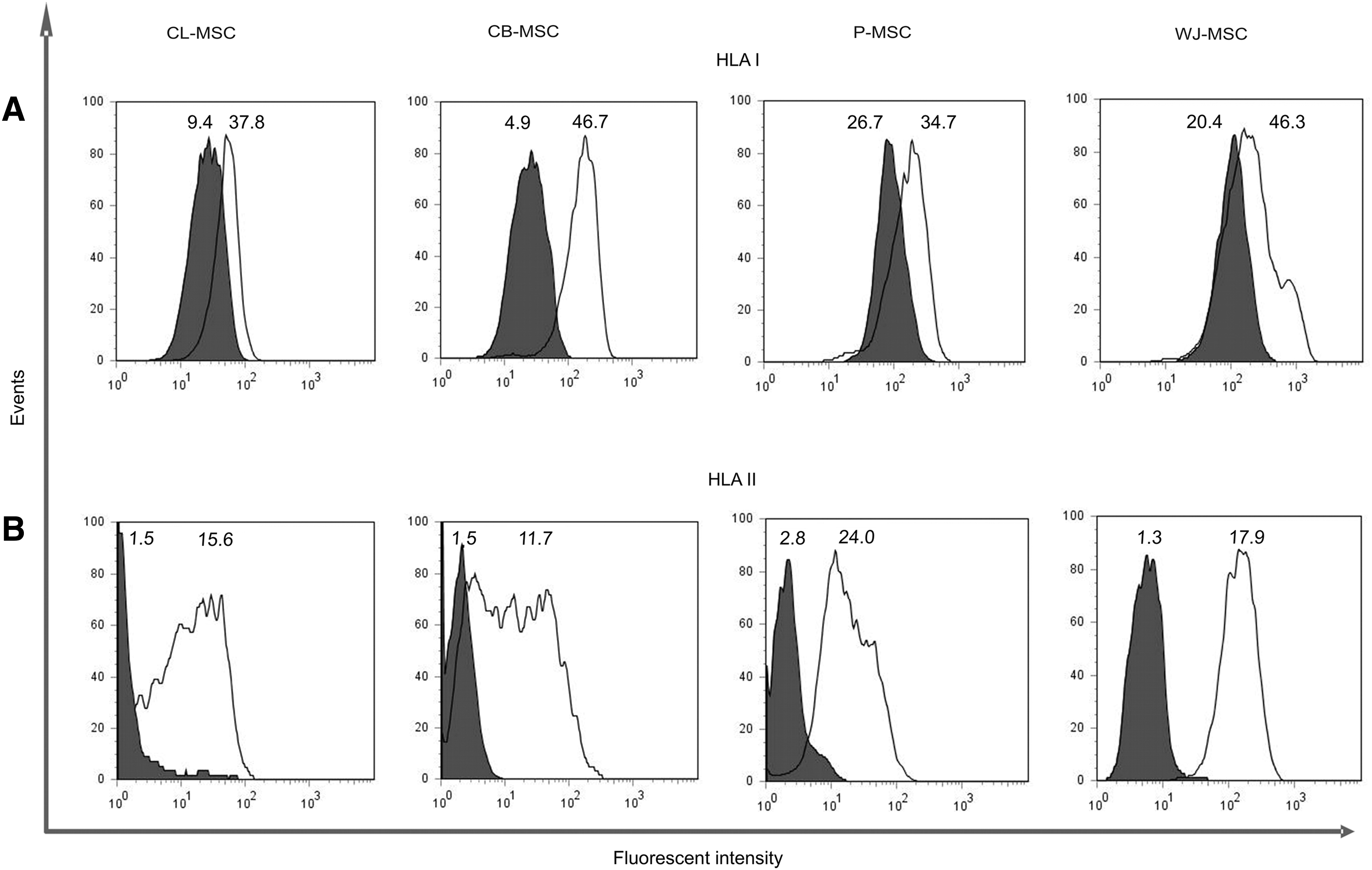
HLA I and II expression.
All native MSCs showed a very low HLA class II expression before the addition of IFN-γ (Fig. 3B). IFN-γ (25 ng/mL) induced the upregulation in HLA II expression in all MSCs (P<0.01 for both CL-MSCs and CB-MSCs, P<0.001 for both P-MSCs and WJ-MSCs). WJ-MSCs showed the highest expression level after stimulation (P<0.001 for all MSC types versus WJ-MSCs).
Immunomodulation by MSCs
HLA-G expression is associated with the immunosuppressive phenotype of MSCs [29,30]. The upregulation of surface HLA-G in the presence of IFN-γ with the exception of CB-MSCs (P<0.001) was not significant (Fig. 4A). Differences in expression were, however, observed among the MSC types studied (P<0.02 CL-MSCs versus CB-MSCs and P<0.05 for CB-MSCs versus P-MSCs).
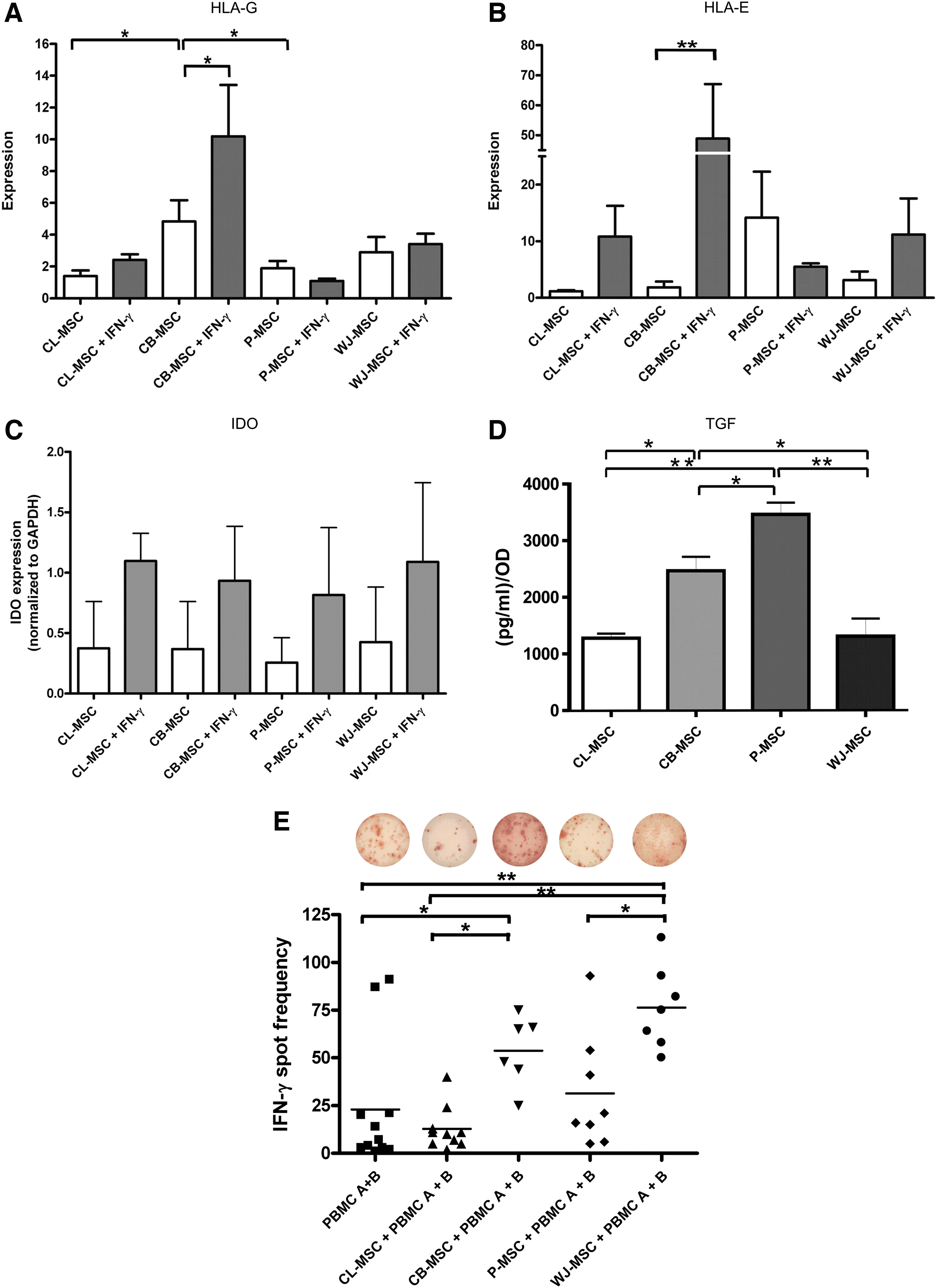
Immunomodulatory capabilities of extraembryonic gestational tissue-derived MSCs.
In addition, MSCs express the NK cell ligand HLA-E (Fig. 4B) [29]. HLA-E upregulation by IFN-γ reached significance only in CB-MSCs (P<0.001).
It has been demonstrated that IDO plays an important role in mediating the suppressive effect of MSCs on T-cells [31]. IDO, which is involved in the catabolism of tryptophan, an essential amino acid crucial for T-cell proliferation [32], was not detected in native MSCs (Fig. 4C). However, all MSC types showed a similar degree of IDO upregulation after a 48-h stimulation period with 500 ng/mL of IFN-γ, which in our experiments mimicked an inflammatory milieu.
In our previous study [15], we showed that resting CL-MSCs released IL-10 and TGF-β1 and that the release of both cytokines could be increased with IFN-γ stimulation. In our present study, we quantified the amount of cytokines released by MSCs using an ELISA assay. IL-10 and IL-2 were not detected in the conditioned supernatants from resting MSCs (assay detection limit 7.8 pg/mL) (data not shown), while TGF-β1 was secreted by all MSCs (detection limit 125 pg/mL) (Fig. 4D). After normalization to their cell count, P-MSCs secreted the highest amount of TGF-β1 compared to other MSCs (P<0.05).
The immunomodulatory effect of MSCs on T-cells can be demonstrated in a one-way MLR [14,15]. For this assay, we included mismatched donor and responder PBMCs, as well as MSCs (for HLA-typing results, please refer to Supplementary Table S1A, B). The number of IFN-γ releasing human PBMCs in response to allogeneic PBMCs was significantly increased when CB-MSCs and WJ-MSCs were added (Fig. 4E; P<0.025 and P<0.001 compared to PBMCs A+B, respectively). CB-MSCs and WJ-MSCs induced a significantly stronger IFN-γ response compared to CL-MSCs (P<0.05 and P<0.001, respectively). TH1 responses of WJ-MSC MLR was also significantly higher than using P-MSCs (P<0.002).
Discussion
To date, MSCs have been isolated from organs such as bone marrow, liver, spleen [33,34], and adipose tissue [35]. MSCs derived from extraembryonic gestational tissue in contrast to those mentioned above, show great promise in MSC-based therapies. Their isolation is simple and does not require any invasive procedures and typically involves the isolation of cells from tissues discarded after birth. To identify the most suitable cell type for allogeneic cell-based therapies, we compared the immunogenicity and immunomodulatory capabilities, as well as the proliferative and migratory potential of MSCs isolated from different extraembryonic gestational tissues.
Although the cell types examined in our study fulfilled most of the criteria accepted for MSCs [28], only CL-MSCs were truly multipotent as they were able to differentiate into adipocytes. Moreover, MSCs differed in terms of their cell culture requirements. These differences could be explained by the fact that the cells were isolated using different techniques, from different extraembryonic tissue compartments, and from different donors and from different laboratories. MSC-type and donor-specific differences in adipocyte differentiation have in the past been observed by other groups [20,27]. It is conceivable that MSCs are not a homogenous population and the tissue from which they are isolated distinguishes them [27,34,36] and directs their potential therapeutic use. This study aimed to determine whether the extraembryonic gestational tissue-derived MSCs differed in their suitability for cell-based therapy.
MSC migration plays a pivotal role in MSC-mediated wound healing. Significant differences in migration rates were observed between MSC types, showing CL-MSCs migrating the fastest into the scratch area. Large numbers of MSCs are needed for regenerative therapies, which prompted us to examine whether the MSCs differed in their proliferative potential in vitro. In this assay, CL-MSCs and CB-MSCs showed the highest rates, making their ex vivo expansion easier.
There is a growing body of evidence suggesting that MSCs induce tolerance and so can survive in vivo for prolonged periods of time in an allogeneic setting [37]. On the other hand, previous studies have also suggested that allogeneic MSC transplantation can lead to the induction of immune responses in the recipient and lead to the rejection of the implanted cells [38]. In our present study, we demonstrated the low immunogenicity of extraembryonic tissue-derived MSCs and especially CL-MSCs. All MSCs showed a low to moderate HLA class I, low HLA II, and no costimulatory molecule expression. Therefore, we investigated the MSC targeted xenogeneic immune response in vivo using immunocompetent BALB/c mice. CL-MSCs, which showed the latest rejection, demonstrated also the lowest TH1 and TH2 cell activation in our ELISPOT assay using BALB/c sensitized splenocytes as responders. Similarly, when we used human PBMCs as responders, the spot frequency was low, which suggests that human CL-MSCs are able to limit murine as well as human immune responses. Interestingly, P-MSCs although they induced a high xenogeneic response, were able to limit human TH1 and TH2 cell activation. The reason for this might be the absence of responsiveness to human IFN-γ (Fig. 3). In the murine system, the response to IFN-γ is missing as human MSCs are not reactive to mouse IFN-γ. Further studies should demonstrate the immunogenicity of human MSCs in the mouse system under inflammatory (human IFN-γ) conditions.
In our study, all MSC types increased HLA class I expression in the presence of a moderate concentration of IFN-γ. We expect that the reason for WJ-MSCs inducing a very high IFN-γ response in MLR assays is caused by their high HLA class I and II expression.
The increase in MSC immunogenicity due to HLA I or II upregulation could potentially be counter-balanced by various immunomodulatory molecules, such as HLA-G, HLA-E, or IDO, which can influence the recipient's immune responses. Nonclassical HLA expression (HLA-G) has been suggested in playing a role in MSCs evading immune responses [30]. More importantly, it has been shown that both HLA-E and HLA-G expression, on the target cell are needed for the inhibition of NK cell lysis [39]. Therefore, we compared HLA-G and HLA-E expression among the different types of MSCs with and without IFN-γ stimulation. We found that only CB-MSCs show a significant increase in both HLA-G and HLA-E surface expression. However, all MSCs studied were able to significantly upregulate IDO after a 48-h stimulation period with high-dose IFN-γ.
Since MSCs exert immunomodulatory effects not only via direct cell–cell contact, but also by releasing soluble factors such as IL-10 and TGF-β1 [40], a varied expression of these factors will lead to differences in exerting immunosuppressive effects. We found that P-MSCs released significant amounts of TGF-β1.
In our hands, extraembryonic tissue-derived MSCs were unable to weaken the immune reaction in MLRs. Indeed, CB-MSCs and WJ-MSCs even significantly enhanced the release of IFN-γ by PBMCs. Similarly, human first trimester liver-derived MSCs are unable to suppress an MLR, which has been attributed to their immunological immaturity [41]. Contrary findings have been reported for second trimester MSCs from gestational tissues, which suppress MLR responses in an IL-10-dependent manner [42]. In our study, we investigated extraembryonic gestational tissue MSCs from full-term births, which clearly differ from second trimester-derived MSCs. The lack of MLR suppressive potential observed here may be due to their low IL-10 release. Second trimester MSCs from the fetal–maternal interface could be better equipped to suppress an MLR. However, MSCs could possibly lose this capability around the time of birth. Roelen et al. reported that gestational tissue-derived MSCs of fetal origin (amnion and amniotic fluid) show a stronger inhibition of MLRs compared to MSCs of maternal (decidua) origin [42]. In this study, MSCs from the umbilical blood and cord were of fetal [23], whereas placental MSCs were of maternal origin [20], which could possibly explain some of the observed differences in our immunological assays.
Although CL-MSCs expressed only moderate levels of HLA-G, HLA-E, and TGF-β1 and also did not differ in IDO production from other MSCs, they showed the longest in vivo survival rates and the lowest immune stimulation in our xenogeneic murine and allogeneic human ELISPOT assays. More importantly, these findings are further supported by the result of our human MLR as only CL-MSCs slightly reduced the production of IFN-γ by PBMCs in the allo-response.
In summary, we found that extraembryonic gestational tissue-derived MSC populations show a varied potential to evade immune responses as well as exert immunomodulatory effects. CL-MSCs showed the most promising potential for a cell-based therapy, as the cells showed low immunogenicity, but they also showed enhanced proliferative and migratory potential. Future research should concentrate on the best disease models in which CL-MSCs could be administered. Furthermore, our study highlights that although immunomodulatory molecules play an important role in MSC immune evasion, HLA expression is in some way related to the immunogenicity of MSCs, and therefore basal and induced HLA-expression should be examined when choosing suitable cell types for cell-based off-the-shelf therapies.
Footnotes
Acknowledgments
S.S. has received research grants from the Deutsche Forschungsgemeinschaft (DFG) (SCHR992/3-1 and SCHR992/4-1) and the ISHLT Shumway Career Development Award 2010. T.D. received a research grant from the ISHLT 2009. We thank Christiane Pahrmann for her excellent technical assistance and Edwin Chow (CellResearch Corporation Pte Ltd.) for providing CL-MSCs and cell culture medium. We also thank Joanna Kawalkowska for her participation (CL-MSC and CB-MSC) in the study.
Author Disclosure Statement
T.T.P. is affiliated with the CellResearch Corporation. The other authors have no conflict of interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.