
Abstract
Dysferlinopathies are caused by mutations in the DYSF gene. Dysferlin is a protein mainly expressed in the skeletal muscle and monocytes. Cell therapy constitutes a promising tool for the treatment of muscular dystrophies. The aim of our study was to evaluate the effect of bone marrow transplantation (BMT) using the A/J Dysfprmd mouse model of dysferlinopathy. For that purpose, we studied dysferlin expression by western blot and/or immunohistochemistry in transplanted mice and controls. Computerized analyses of locomotion and electrophysiological techniques were also performed to test the functional improvement. We observed dysferlin expression in splenocytes, but not in the skeletal muscle of the transplanted mice. However, the locomotion test, electromyography studies, and muscle histology showed an improvement in all transplanted mice that was more significant in the animals transplanted with dysferlin+/+ cells. In conclusion, although BMT restores dysferlin expression in monocytes, but not in skeletal muscle, muscle function was partially recovered. We propose that the slight improvement observed in the functional studies could be related with factors, such as the hepatocyte growth factor, released after BMT that prevented muscle degeneration.
Introduction
Dysferlin is a type 2 protein of 2080 amino acids and a molecular weight of about 237 kDa, which is expressed in different tissues such as the kidney, heart, liver [1], and placenta [12]. However, most research is focused in skeletal muscle [13] and monocytes [14 –16].
Different studies have demonstrated a role of dysferlin in sarcolemma repair after injury [17,18], but it has also been shown to be involved in muscle differentiation in vitro [19] and in vivo [20].
The presence of inflammatory infiltrates, mainly constituted by macrophages, is a hallmark of dysferlinopathies [11]. The presence of inflammation has been observed in other muscular dystrophies such as Duchenne muscular dystrophy (DMD) [21] and facioscapulohumeral muscular dystrophy [11,21,22]. The absence of dysferlin has also been reported to enhance phagocytosis in murine monocytes [16], and macrophages from patients with dysferlinopathy show altered adhesion and motility [23]. These findings suggest that in dysferlinopathies, muscle inflammatory infiltrates could worsen and perpetuate the dystrophic process.
Cell therapy has become an important research line for the treatment of muscular dystrophies. The use of human umbilical cord cells in the SJL mouse model [24] resulted in low engraftment and low levels of dysferlin expression in muscle [25]. In another study, human adipose-derived stromal cells were used in the same mouse model with similar results [26]. In yet another study, the authors found that mesoangioblasts, a vessel-associated progenitor cell, can engraft and restore dysferlin expression in skeletal muscle in A/J mice [27], another mouse model of dysferlinopathy [18].
Bone marrow transplantation (BMT) constitutes a different approach of stem cell-based therapy in muscular dystrophies. Because allogeneic BMT has been extensively used for the treatment of different pathologies in humans, a possible beneficial effect in animal models of muscular dystrophy could be more easily translated to therapy in patients. The first BMT in a dystrophic animal was performed in the mdx mouse, a model for DMD [28,29]. The authors found that some transplanted cells were able to engraft in the skeletal muscle, but not to express dystrophin. In another study, BMT was assessed in a muscular mouse model of spinal muscular atrophy, which presents a mild dystrophic muscular phenotype [30]. In view of their results, the authors suggested that the biological activity of BM-derived cells was able to improve the myopathic phenotype by enhancing muscle regeneration.
It has been shown that BM-derived cells can fuse into skeletal muscle in vivo [31 –33]. However, those cells failed to express muscle-specific proteins. In the case of dysferlin myopathies, this kind of cell therapy could be more effective given that some hematopoietic cells such as monocytes express dysferlin. Furthermore, monocytes were reported to be able to fuse and regenerate other tissues such as the liver [34]. BMT has not been tested in animal models of dysferlinopathy,
The general aim of our study was to evaluate the therapeutic potential of syngeneic BMT in the A/J Dysfprmd mouse model of dysferlinopathy.
Materials and Methods
Mice
For this study, the mouse model for dysferlinopathy A/J Dysfprmd (The Jackson Laboratories) [27] was used as BMT recipients. A/J Dysf+/+ (A/JOlaHsd) mice (Harlan Laboratories) were used as donors, and as positive controls for dysferlin expression. All animals were genotyped as previously described [27].
We discovered the existence of A/J Dysf+/+ (A/JOlaHsd) mice accidentally. We initially purchased these animals from Harlan Laboratories as A/J mice bearing the previously described DYSF mutation [27]. However, when we performed a western blot (WB) analysis using a monoclonal antibody specific for dysferlin, we found that they displayed normal levels of dysferlin. These results prompted us to genotype this strain. Polymerase chain reaction (PCR) analysis of genomic DNA isolated from A/J Dysf+/+ mice and the A/J Dysfprmd, using the indicated primer pairs, yielded the following results: the primer pair that flanked the ETn insertion (F and R) yielded a 207 bp PCR product in mice from Harlan Laboratories, but not in A/J Dysfprmd from Jackson Laboratories. Conversely, only genomic DNA from the A/J Dysfprmd mice generated 416 and 237 bp products when primers specific for the ETn sequences were used (Fig. 1).
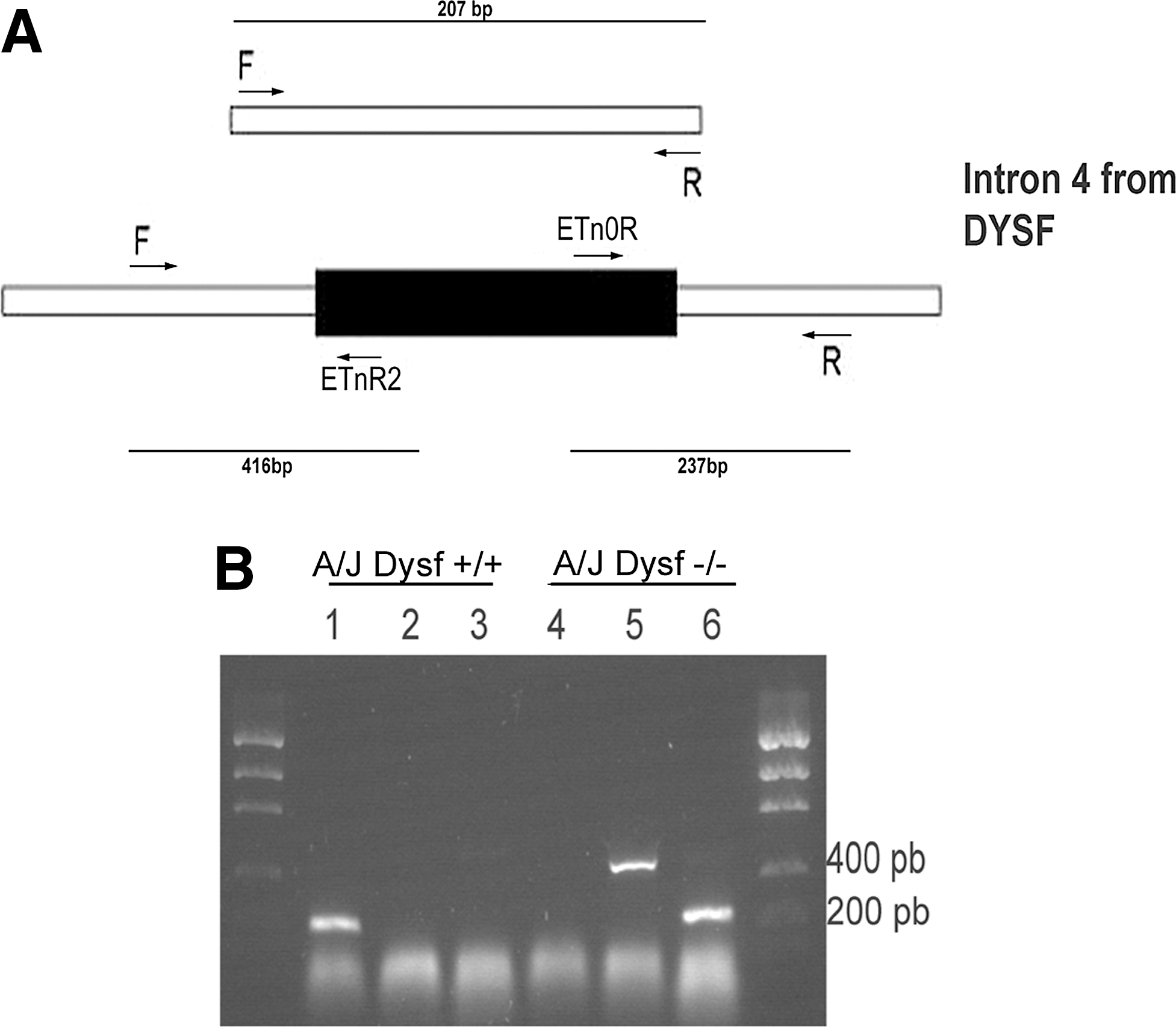
Identification of the dysferlin mutation in the A/J mice.
These results confirmed that these two murine models differ in the expression of dysferlin, as the A/J Dysf+/+ lacks the insertion of a retrotransposon that disrupts the DYSF gene, resulting in the absence of this protein.
The mutation in the A/J strain must have occurred between the late 1970s and the early 1980s [27]. When we checked with Harlan Laboratories about the origin of their mice, they confirmed that they obtained their A/J strain in the early 1970s before the mutation was introduced.
Bone marrow transplantation
Ten untreated A/J Dysf+/+ mice were used as positive controls (group A), 10 untreated A/J Dysfprmd mice were used as negative controls (group B), and 20 A/J Dysfprmd animals were transplanted, 10 with dysferlin+/+ cells (group C) and 10 with dysferlin−/− cells (group D). All animals were 12 weeks old at the time the BMT was performed. The experiment was designed to evaluate a possible preventive effect of BMT in these animals.
The bone marrow cells of donor mice were obtained postmortem by flushing the femorae [35,36]. After sublethal irradiation with 4.5 Gy, 3×106 cells were injected intraperitoneally in the receptor mice during the first hours.
Ten weeks after transplantation, electrophysiological and locomotion analyses were performed in all animals. After functional tests, all mice were sacrificed and samples from quadriceps, gastrocnemius, and tibialis anterior (TA) muscle and the spleen were collected and frozen.
All the procedures using animals in this project were approved by the Animal Experimentation Committees of the Vall D'Hebron Institut de Recerca, Barcelona, Spain and the Universitat Autònoma de Barcelona, Spain.
Digigait analysis
Digigait analysis was performed using the Digigait Imaging system (Mouse Specifics). Briefly, digital video images of the underside of the mouse were collected with a high-speed video camera (80 frames/s) from below the transparent belt of a motorized treadmill. The mice were accustomed to the treadmill belt and the testing conditions before the recordings. Each mouse was allowed to explore the treadmill compartment, with the motor speed set to zero, for 5 min. Then, the motor speed was set to 20 cm/s to collect the videos. A minimum of 200 images was collected for each walking mouse so that five to seven strides were monitored in each run. Each video image representing 12.5 ms was digitized and the area (in pixels) of the paws was calculated with the DigiGait software [37 –39]. Four parameters were used for the locomotor assessment: (i) the percentage of animals that were able to run at 20 cm/s; (ii) the stride duration (time duration of one complete stride for one paw); (iii) the step angle (the angle that the paw makes with the long axis of the direction of motion of the animal); and (iv) the stance width (The perpendicular distance between the centroids of hindpaws during peak stance).
Electromyography tests
Electromyographic (EMG) recordings of motor unit action potentials (MUAPs) were obtained from the TA and plantar interossei muscles following a similar protocol to that previously described in mdx mice [40] with the mice under light anesthesia. Recordings were obtained in rest conditions and following light noxious stimuli delivered as light pinchs in the tail to provoke bursts of EMG activity. The signals were digitized (Powerlab 6T; ADInstruments) and fed into Chart software for post hoc analysis. From each mouse of the four groups, MUAPs were recorded and the duration, amplitude, and number of phases were measured. Furthermore, MUAPs were categorized into small, medium, and high amplitude as a representation of the three main types of motor units (S, FR, FF) [41] and to facilitate the interpretation of the results. At least six distinct, repeatedly firing MUAPs from each class were selected for analysis.
Western blot
To study protein levels in muscle and spleen, samples were treated with the Nicholson sample buffer (0.125 M Tris/HCl, pH 6.4, 10% of glycerol, 4% sodium dodecyl sulfate (SDS), 4 M urea, 10% of mercaptoethanol, and 0.001% of bromophenol blue) [42] The protein extracts were analyzed in 8% SDS-polyacrylamide gel electrophoresis (PAGE) gels transferred to PROTRAN (Whatman) nitrocellulose membranes. The unspecific binding sites were blocked using the odyssey blocking buffer (LI-COR). Dysferlin levels were detected using the Hamlet I monoclonal antibody (Novocastra) and hepatocyte growth factor (HGF) levels with a rabbit polyclonal antibody (AbCam). Protein levels in mice were normalized with the immunodetection of β-tubulin with a monoclonal antibody (clone B-5-1-2; Sigma). A goat anti-mouse conjugated with IRDye_800 (LI-COR) was used for detection with Odyssey Infrared Imaging System and protein levels were analyzed with the Odyssey 2.1 software (LI-COR). Analysis of dysferlin and HGF was performed in all animals.
Histochemistry and immunohistochemistry studies
For histological description of quadriceps and TA muscles, 7-μm-thick sections were stained with hematoxylin and eosin.
For dysferlin and the monocyte/macrophage marker [43,44] immunodetection, 7-μm-thick sections were fixed with acetone and blocked with Tris buffer saline supplemented with 0.5% normal donkey serum. Dysferlin was detected using a rabbit monoclonal antibody (Epitomics), and macrophages using a rat anti-mouse F4/80 (Acris Antibodies) arginase-1 (MyBioSource) fetal myosin (Novocastra) and rat anti-CD206 (LifeSpan Biosciences) before using the donkey anti-rabbit antibody conjugated to Fluor Alexa 488 (Invitrogen), goat anti-rabbit HRP or goat anti-rat HRP (Jackson Laboratories) in serial sections. The HRP was developed using the peroxidase substrate kit DAB kit (Vector Laboratories). Images were collected with an Olympus DP72 camera connected to a BX51 Olympus fluorescence microscope and analyzed with the Cell^D software. To quantify the number of central nuclei or macrophages in muscle, four fields were photographed at 200× from hematoxylin and eosin-stained frozen sections of at least three different animals per group.
RNA extraction and semiquantitative retrotranscriptase PCR
RNA from all animal groups (5–6 animals/group) was extracted using Ultraspec (Biotech Laboratories, Inc.). Total RNA (1250 ng) of each RNA sample was reverse transcribed into cDNA using MultiScribe reverse transcriptase (Applied Biosystems). Quantification of the mRNA encoding DYSF, and GAPDH as internal standard was performed using TaqMan Universal Master Mix technology (Applied Biosystems). Quantitative PCR was performed in a total reaction volume of 12 μL per well. The primers used for retrotranscriptase (RT)-PCR were designed by Applied Biosystems (DYSF, Mm00458042_m1, GAPD, 4352932E). The comparative CT method (ΔΔCT) for relative quantification of gene expression was used as previously reported [19], since the target and normalizer (GAPDH) have similar dynamic ranges.
Statistical analysis
For all functional studies, central nuclei counting, macrophage quantification, and real-time PCR, differences in results between groups were analyzed by one-way analysis of variance (ANOVA) followed by Bonferroni post hoc tests. Statistical significance was set at P<0.05.
Results
Functional improvement was observed in animals treated with Dysf+/+ syngeneic BM cells
We performed functional studies before the animals were euthanized for histological and dysferlin expression study. Regarding the locomotor evaluation, we first assessed the percentage of animals that were able to run at 20 cm/s as a measure of general locomotor performance [45]. This analysis revealed that both Dysf+/+ and Dysf−/− cell transplantation promoted a partial recovery of the general locomotor performance (Fig. 2). Second, we assessed a detailed gait analysis. A/J Dysfprmd mice showed mild locomotor abnormalities, evidenced by a significant increase in the stride duration and the step angle with respect to A/J Dysf+/+ mice. A/J Dysfprmd mice transplanted with Dysf+/+ cells showed a partial recovery of the stride duration compared to nontreated and autologous transplanted A/J Dysfprmd animals, suggesting a mild improvement in gait performance. Moreover, both Dysf+/+ and Dysf−/− cell transplantation promoted a partial recovery in terms of step angle (Supplementary Table S1; Supplementary Data are available online at
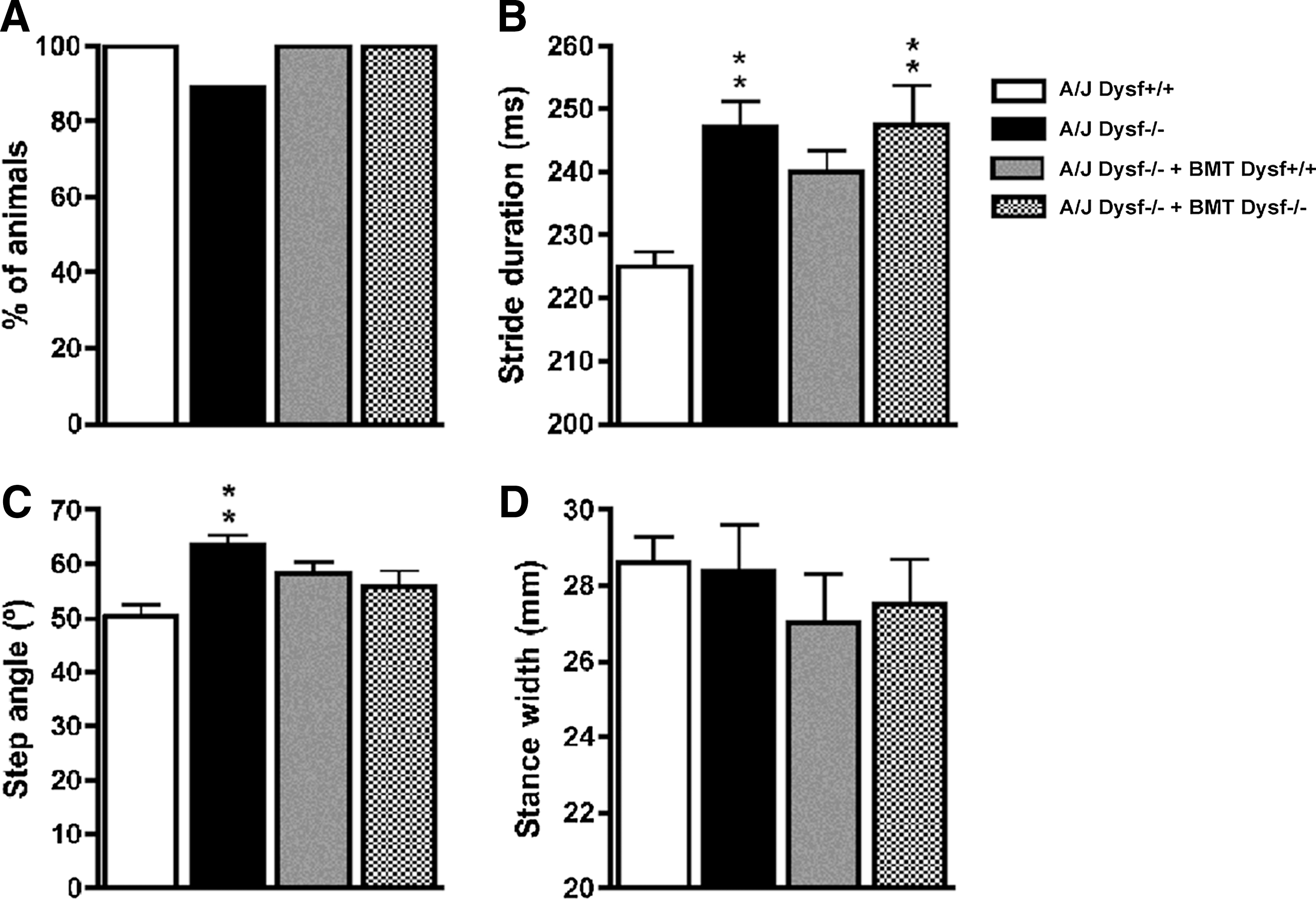
Results of locomotion analysis (Digigait). Abnormalities manifested by reduction of general locomotion
EMG analysis showed a decreased amplitude, and an increased duration and number of phases in the MUAPs recorded from the muscles of A/J Dysfprmd mice muscles, all of them hallmarks of myopathic condition [46]. In groups C (A/J Dysfprmd transplanted with Dysf+/+) and D (A/J Dysfprmd transplanted with Dysf−/−), the mean MUAP amplitude was completely recovered to normal values. However, MUAP duration and number of phases was only partially improved, slightly more in the group C than in group D (Fig. 3, Supplementary Table S2 and Supplementary Fig. S1).
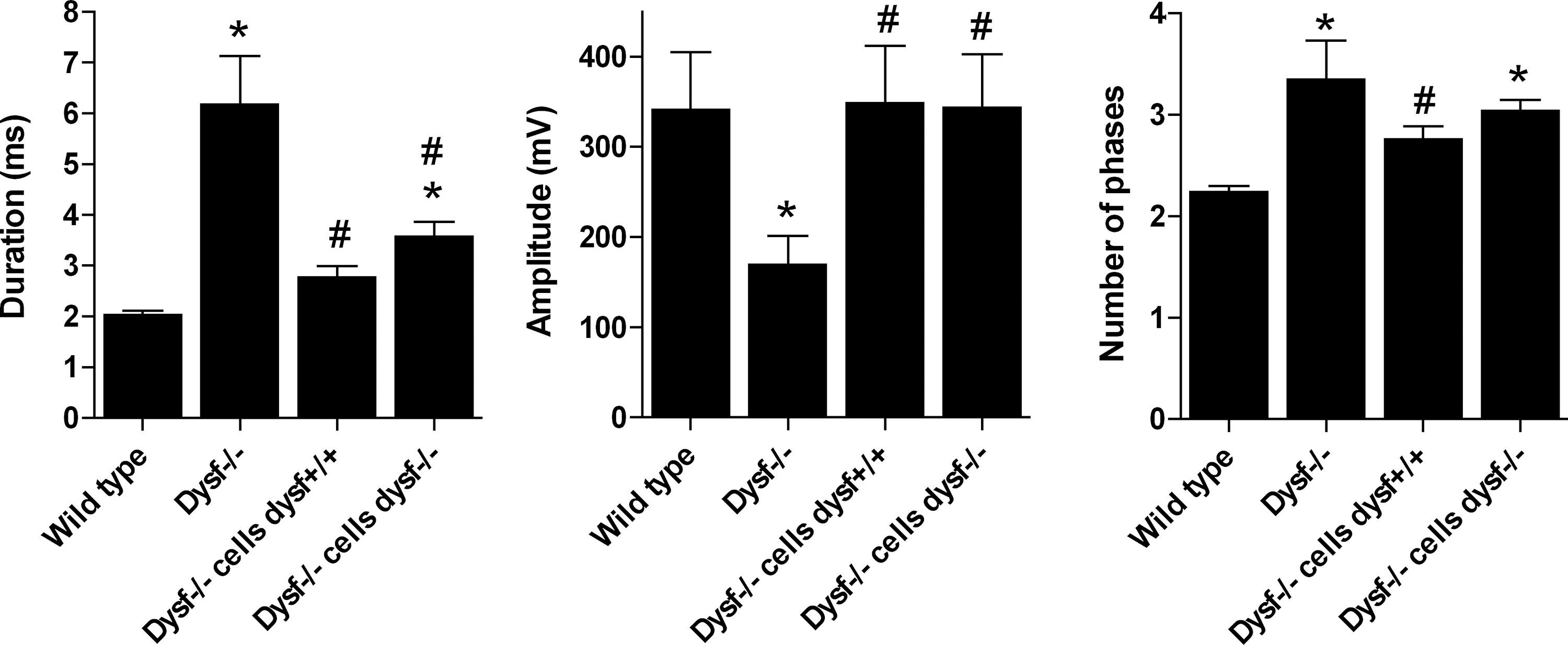
Most relevant EMG results. A/J Dysf+/+ mice as the control group for normal dysferlin expression, A/J Dysf−/− as the control group for dysferlin deficiency (A/J Dysfprmd), A/J Dysf−/−+BMT Dysf+/+ as A/J Dysfprmd transplanted with Dysf+/+ BM cells, and A/J Dysf−/−+BMT Dysf−/− as A/J Dysfprmd transplanted with Dysf−/− BM cells. *P<0.05 versus Dysf+/+, # P<0.05 versus Dysf−/−. EMG, electromyographic.
Reconstitution of dysferlin expression in peripheral blood monocytes does not prevent inflammation in skeletal muscle
After functional tests were performed, mice were euthanized and tissue samples were collected to assess the efficacy of treatment. WB analysis of spleen cells from A/J Dysfprmd mice treated with Dysf+/+ showed expression of dysferlin (Fig. 4) with an average expression of 9.2% of normal values. In contrast, nontreated A/J Dysfprmd and A/J Dysfprmd mice transplanted with Dysf−/− BM cells showed no dysferlin expression. Histological studies showed the presence of central nuclei in A/J Dysfprmd (Fig. 5). We observed a statistically significant decrease in the number of central nuclei in all treated mice compared to nontreated A/J Dysfprmd (P<0.05) both in the TA muscle and quadriceps muscle (Fig. 6). Prominent inflammatory infiltrates were also observed in all treated and not treated A/J Dysfprmd groups (Fig. 7). Some inflammatory cells were positive for dysferlin in Dysf−/− mice treated with Dysf+/+ cells, but not those treated with Dysf−/− cells (Fig. 7).
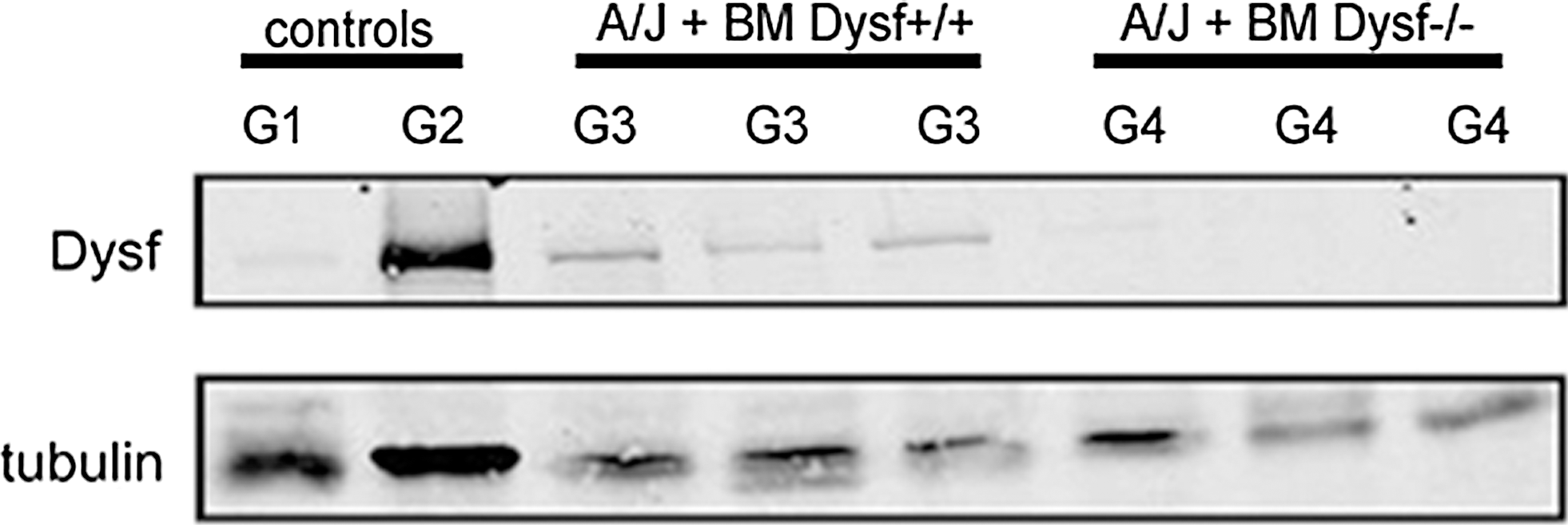
WB analysis of dysferlin expression in spleen cells. G1: A/J Dysfprmd; G2: A/J Dysf+/+; G3: A/J Dysfprmd transplanted with Dysf+/+ BM cells; G4: A/J Dysfprmd transplanted with Dysf−/− BM cells. WB, western blot.
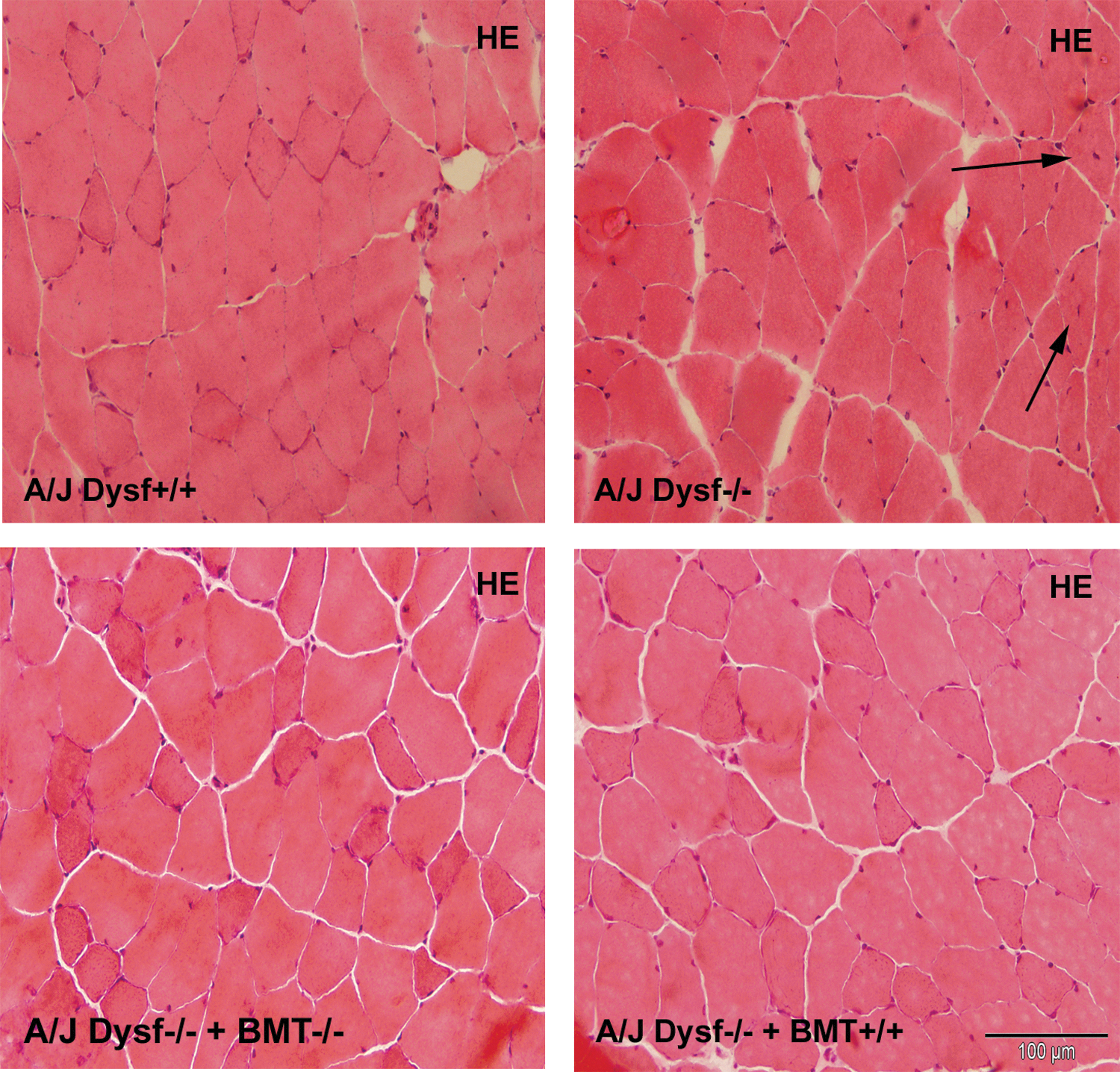
Histological analysis of skeletal muscle from 6-month-old mice of all groups. HE stainingof quadriceps muscle from each group (200×). Arrows show central nuclei. Scale bars represent 100 μm. HE, hematoxylin and eosin. Color images available online at
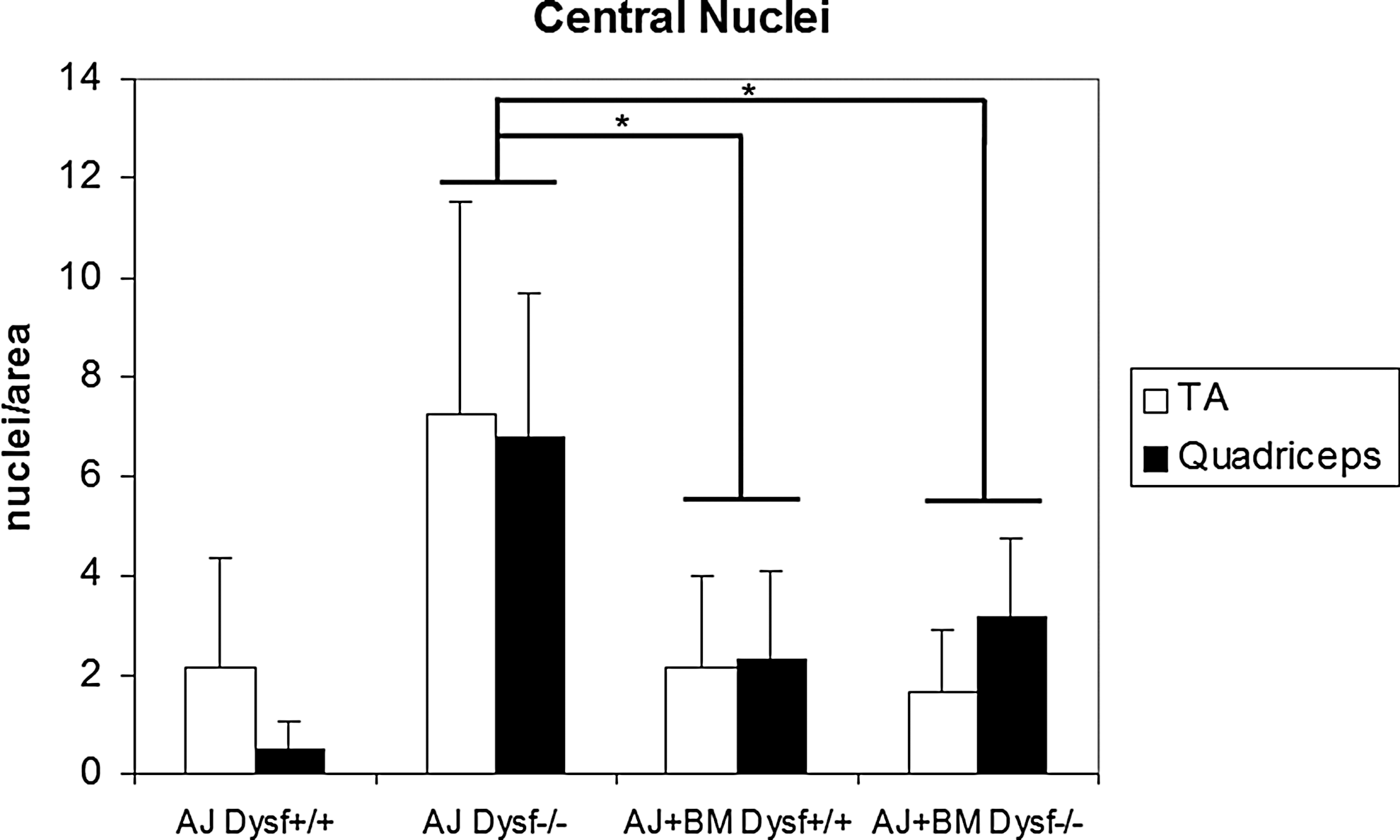
Central nuclei quantification. Bar graph showing the number of central nuclei/area in all animal groups (*P<0.05) in TA and quadriceps muscles. The number of central nuclei is reduced in all treated animals. TA, tibialis anterior.
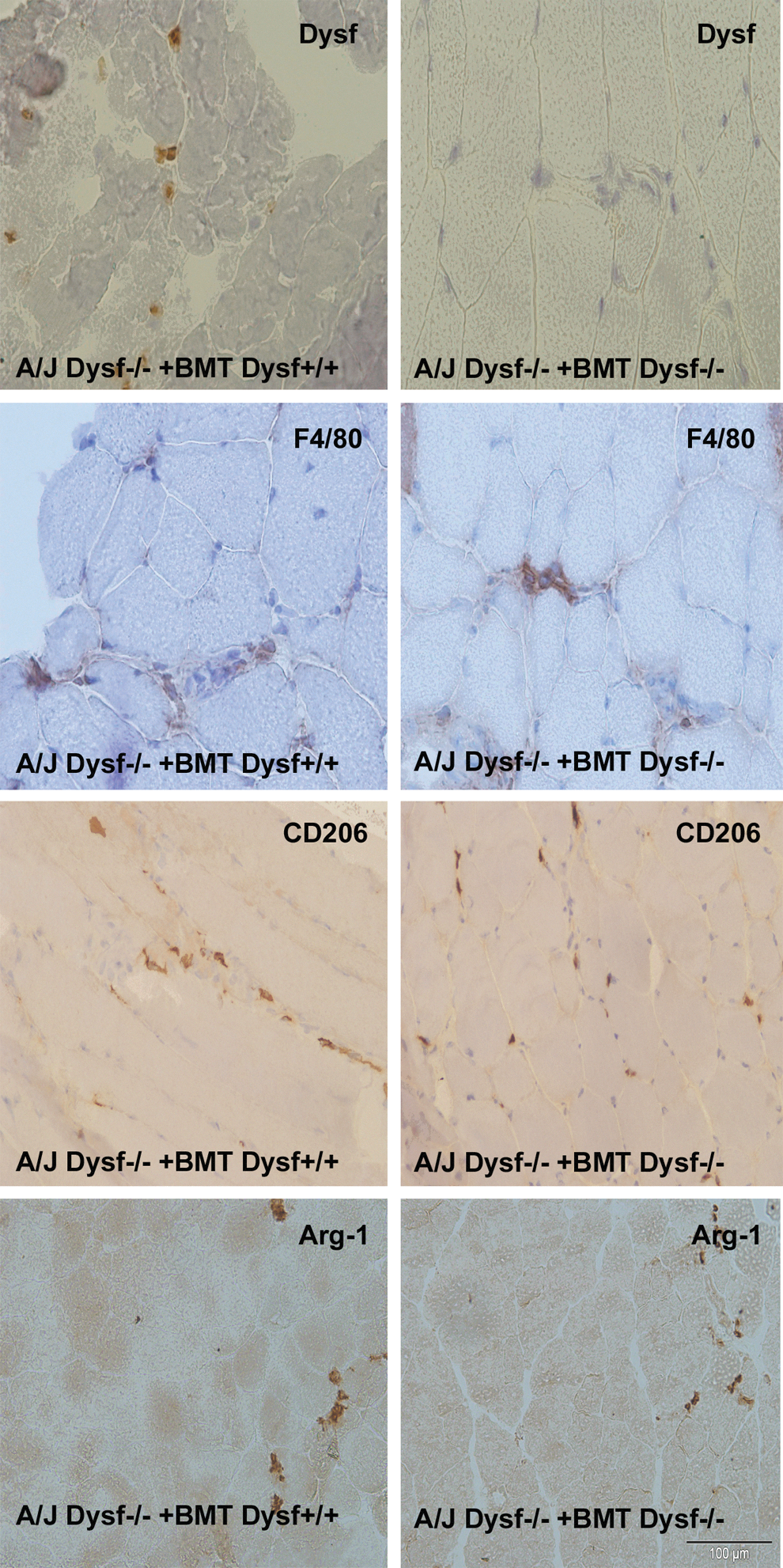
Characterization of infiltrating cells in skeletal muscle. Dysferlin immunostaining in transplanted mice showing positive inflammatory cells (400×) only in those animals transplanted with dysferlin+/+ cells. Immunohistochemistry of F4/80, a general monocyte/macrophage marker (200×), CD206 immunostaining and Arg-1, two anti-inflammatory monocyte/macrophage markers (200×). Scale bars represent 100 μm. Arg-1, arginase-1. Color images available online at
The inflammatory infiltrates were characterized as macrophages using the F4/80 marker (Fig. 7). There were no significant differences between the number of F4/80+ infiltrates in each group of transplanted mice and the A/J Dysfprmd (P>0.05) (Fig. 8). When we studied the expression of CD206 and arginase-1 markers of an anti-inflammatory subset of macrophages, only some infiltrating cells were positive. We found no statistically significant differences between the groups (P>0.05) (Fig. 7).
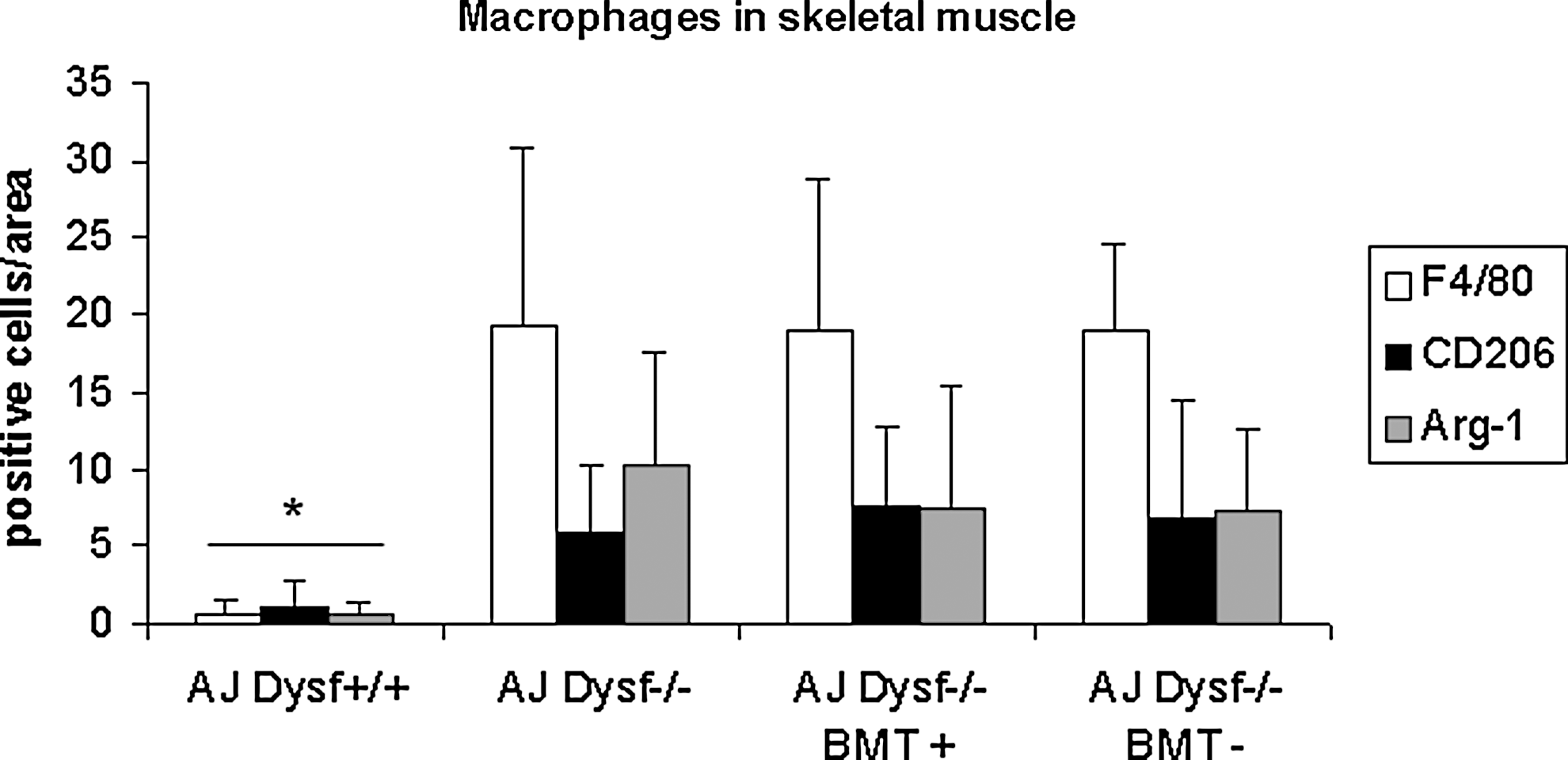
Macrophage quantification. Bar graph showing CD206, Arg-1, and F4/80-positive cell counting in the muscle tissue inflammatory infiltrates from skeletal muscle of all the experimental groups.
BMT does not restore dysferlin expression in skeletal muscle, but prevents muscle degeneration
When we analyzed the skeletal muscle from treated mice by WB, we did not find dysferlin expression in any group of A/J Dysfprmd mice either treated or not (Fig. 9A). However, in the immunolocalization studies, we observed some isolated fibers showing a patchy expression of dysferlin in mice treated with dysferlin-positive cells (Fig. 9B). The real-time PCR analysis for Dysf mRNA expression showed increased levels in mice treated with Dysf+/+ BMT. This increase in dysferlin mRNA expression was not statistically significant when compared to nontreated A/J Dysfprmd mice (P>0.05) (Fig. 9C).
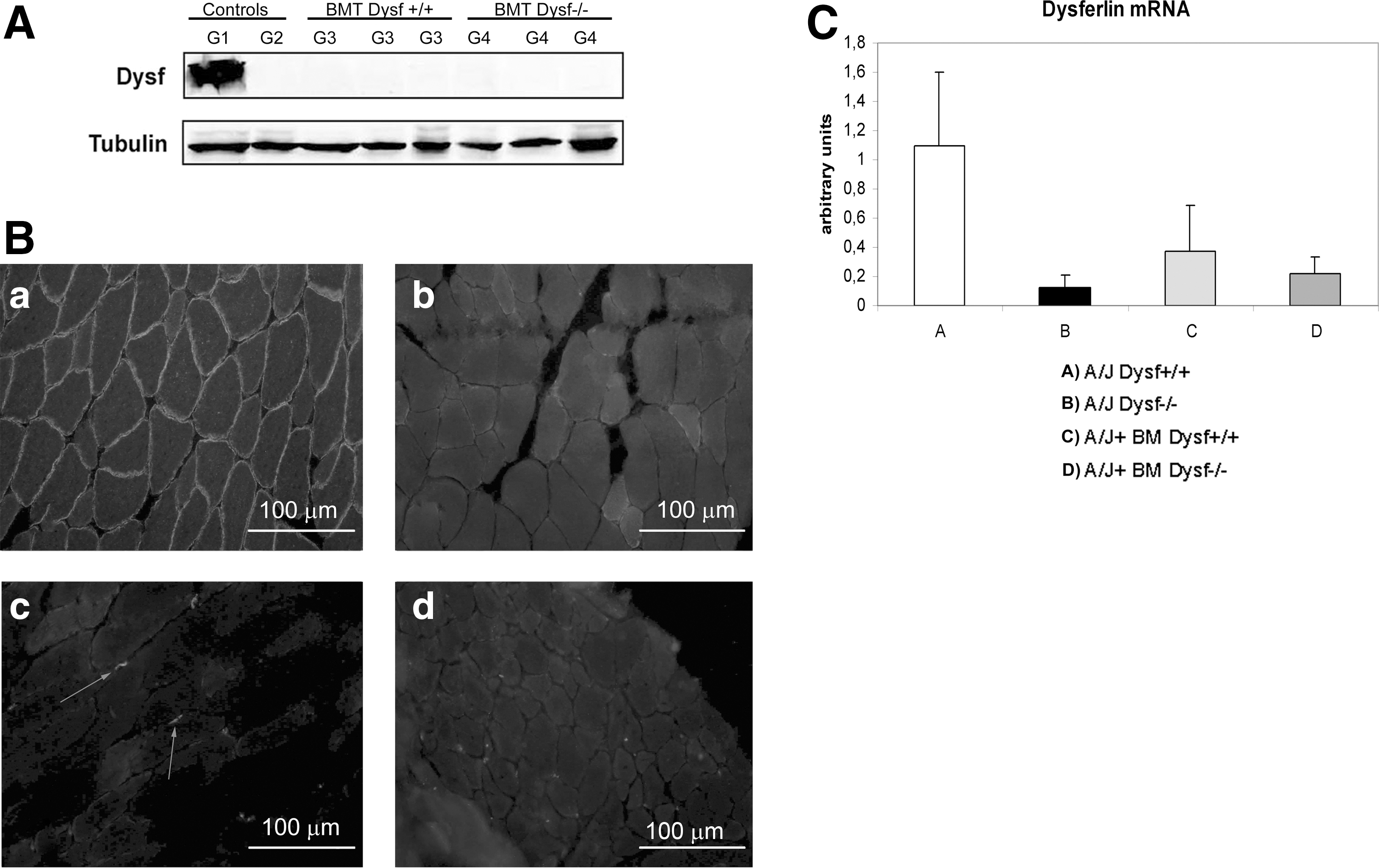
Dysferlin expression in skeletal muscle.
We also analyzed the presence of fetal myosin-positive fibers in all mice groups. We observed isolated fetal myosin-positive fibers in all groups.
HGF expression is reduced in the A/J Dysfprmd mouse model and restored after partial reconstitution of dysferlin in peripheral blood
The study of protein extracts from quadriceps muscles by WB showed a significant reduction of HGF expression in nontreated A/J Dysfprmd mice as compared to that of A/J Dysf+/+ mice.
When we analyzed animals from treated groups, we could observe restoration of HGF expression to normal levels only in those animals treated with Dysf+/+ cells. The HGF protein levels of mice transplanted with Dysf+/+ BM cells were similar to those from A/J Dysf+/+ mice, but were significantly higher than those of nontreated A/J Dysfprmd animals (P<0.02) or A/J Dysfprmd mice treated with Dysf−/− BM cells (P<0.01) (Fig. 10).
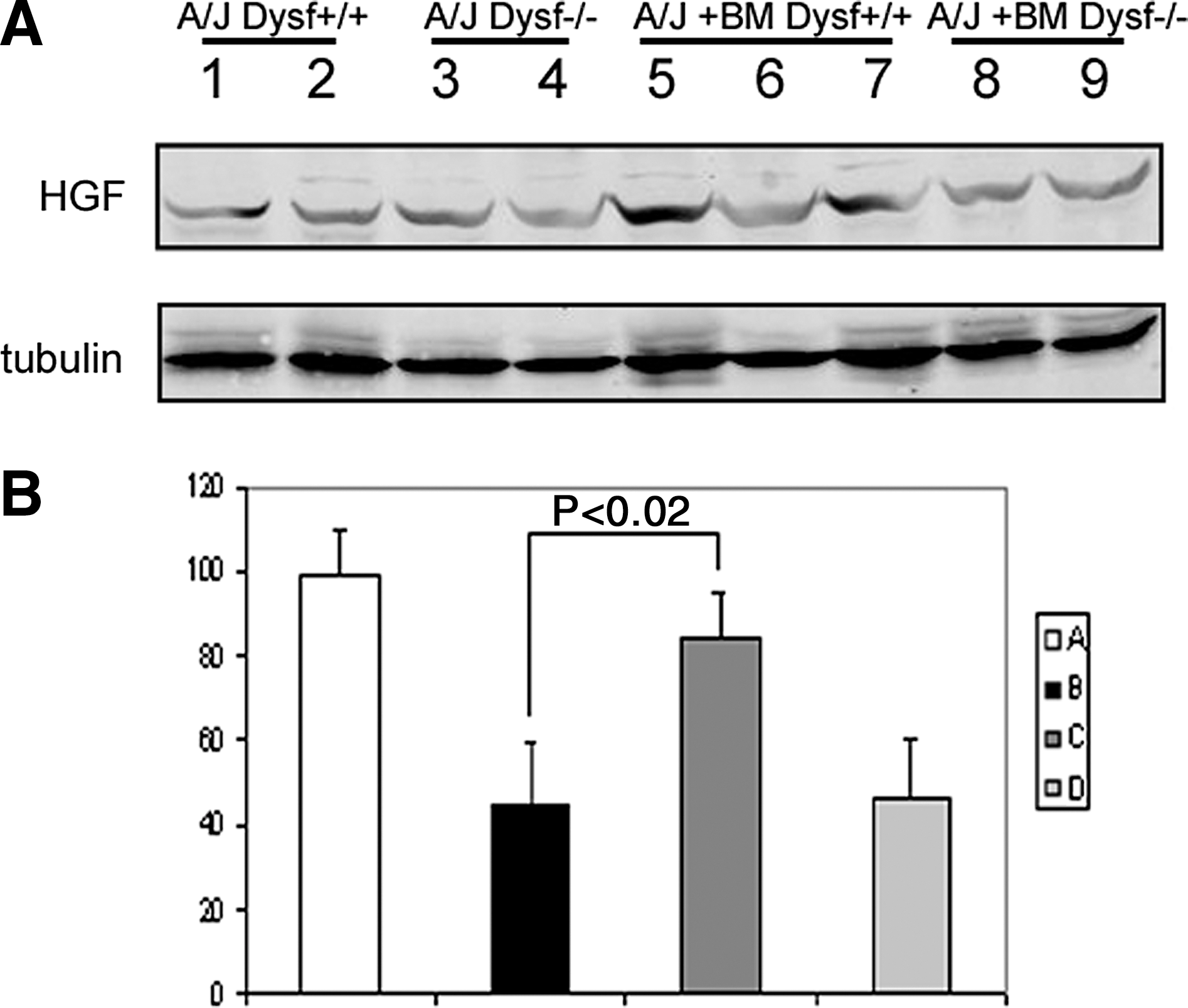
Expression of HGF by WB.
Discussion
Our study demonstrates that BMT slightly improved functional and histological outcome in the A/J Dysfprmd mouse model, even though the expression of dysferlin was not rescued in muscular tissue. Although our results indicate that Dysf+/+ BMT promotes a functional recovery, they also suggest that the improvement is due, in part, to the BMT procedure by itself, since animals treated with BMT cells from A/J Dysfprmd mice also displayed some improvement.
We would like to remark that EMG studies performed in the present work to assess muscle function are equivalent to those used for the diagnosis of myopathic patients. In accordance with these results, it has been recently published that there is a lack of correlation between the results obtained in vitro using skeletal muscle membrane repair assays and the amelioration of dystrophic changes observed in vivo in the same mouse models of dysferlinopathy after different treatments. The authors concluded that the selection of assays to demonstrate the efficiency of a treatment has to be done carefully [47]. We believe that the use of EMG may become a test of choice in some experimental settings.
Although it needs to be further investigated, our results suggest that BMT enhanced skeletal muscle regeneration by a process likely mediated by a biological activity of BM-derived cells as proposed by other authors [30,48]. In the mdx mouse model for DMD, BM-derived cells can fuse to skeletal muscle, but this does not result in a significant increase in dystrophin expression [28,29]. Unfortunately, in these articles, the authors did not perform functional studies to analyze if BMT produced any benefit to the animals. The low levels of dystrophin expression observed by these authors might be due to the fact that hematopoietic cells do not express dystrophin. However, it has been shown that at least some BM-derived cells, including monocytes/macrophages, express dysferlin [15]. This led us to hypothesize that BMT could be more effective in restoring dysferlin expression in muscle in a dysferlinopathy mouse model. Although BMT rescued dysferlin expression in spleen cells, it did not restore dysferlin expression in skeletal muscle in our model. Only in animals transplanted with Dysf+/+ BM cells we found scattered patches of dysferlin-positive sarcolemma that could be explained by fusion of transplanted cells with muscle fibers. Although we observed an increased expression of dysferlin mRNA in the muscle of mice treated with Dysf+/+ BM cells, these results could be explained mainly by the presence of BM-derived Dysf+/+ macrophages infiltrating the muscle.
The origin of inflammation in dysferlinopathies is still under debate. Two laboratories have generated transgenic mice expressing dysferlin only in skeletal muscle. These mice do not present dystrophic features and interestingly, they do not show inflammation. This suggests that restoring dysferlin expression in skeletal muscle could be sufficient to avoid inflammatory cells infiltrating the muscle tissue [49,50]. Accordingly, our results demonstrate that reconstitution of dysferlin expression only in hematopoietic cells is not sufficient to avoid inflammation in dysferlin-deficient skeletal muscle, as we observed inflammatory infiltrates in all treated mice regardless of the origin of BM cells transplanted.
Nevertheless, we observed evidences of locomotor and EMG recovery in A/J Dysfprmd treated with dysferlin-positive cells compared to nontreated mice. Autologously treated mice also improved, but to a lesser extent. All treated mice also showed a reduction in central nuclei although the number of myosin-positive fibers was very low in our groups, suggesting that BMT had a preventive effect on the progression of the disease. Previous reports by other authors could explain, at least in part, these findings [16,30,51]. Our immunohistochemistry experiments showed dysferlin-positive macrophages in the muscle of mice transplanted with dysferlin+/+ BM. As expected, muscle infiltrates in mice transplanted autologously, showed dysferlin−/− macrophages. Nagaraju et al. demonstrated that dysferlin-deficient monocytes have increased the phagocytic activity and that this could play a relevant role in the disease [16]. These findings could explain the milder improvement observed in the group of mice treated with dysferlin−/− cells since dysf−/− macrophages could have some deleterious effect on skeletal muscle. As we observed similar numbers of anti-inflammatory macrophages (CD206 and arginase-1-positive cells) in all AJ Dysfprmd animal groups, based on these results, we cannot conclude that the beneficial effect of BMT is due to infiltrating macrophages switching to an anti-inflammatory profile.
The restoration of HGF expression to normal levels in our mice treated with Dysf+/+ BM cells could also play an important role in the functional recovery of those animals. Salah-Mohellibi et al. showed, in a murine model of spinal muscular atrophy, an important upregulation of HGF after BMT and consequent motor improvement of the animals [30]. However, the authors did not explain the mechanisms by which HGF was upregulated. HGF is known to activate satellite cells [52 –55] and to promote tissue-derived mesoangioblast mobilization, a vessel-derived progenitor cell involved in muscle regeneration [56]. In fact, after cardiac surgery, the number of circulating tissue-derived mesoangioblasts in peripheral blood significantly increases after a peak in HGF expression levels. The same authors demonstrated that administration of HGF in rats increased the number of circulating mesoangioblasts [57]. It has also been reported that the absence of the HGF activator, urokinase plasminogen activator, worsens the mdx phenotype [58]. We believe that the lower rates of HGF observed in the A/J Dysfprmd mouse model may contribute to the muscle pathology, and that restoration of normal HGF expression following BMT with Dysf+/+ cells contributed to mild improvement observed. We hypothesize that normal levels of HGF promote the mobilization of satellite cells and mesoangioblasts for tissue repair, which although do not express dysferlin can reduce damage in the short term. The slight functional improvement in mice transplanted with Dysf−/− BM cells with not significant upregulation of HGF suggests the involvement of factors other than HGF that are released by the new BMT cells.
We do not know if irradiation per se could have an effect on muscle regeneration. However, irradiation produces animal morbidity due to tissue damage, which in turn triggers inflammatory responses mediated mainly by TNF-α. This cytokine produces fever, hypotension, shock, and respiratory distress. All these effects together with the inherent transient period of immunodeficiency lead to an important risk of death secondary to infection. All these issues make it really difficult to get a protocol accepted by animal ethics committees [59].
Finally, an additional relevant contribution of our study is the description of A/J mice that do not bear the mutation previously described in DYSF [27]. Consequently, this local strain can be used as a suitable parental control for the AXB and BXA RI strains [27] and as a control for A/J Dysfprmd mice in future studies, since they share all their genetic background except for the mutation in DYSF.
We conclude that (i) BMT does not reduce skeletal muscle inflammation; (ii) BMT after sublethal irradiation improves MUAPs, locomotor performance, and histopathological features in our mouse model for dysferlinopathy; (iii) this improvement is higher in animals transplanted with Dysf+/+ cells, suggesting that the absence of dysferlin in peripheral blood may also contribute to disease progression; and (iv) The description of A/J mice with normal dysferlin is a useful control for future studies using A/J dysferlin-deficient mice. (v) Autologous BMT in patients with dysferlinopathy may be of some clinical benefit.
Footnotes
Acknowledgments
This work was supported by grants from “Fundación Isabel Gemio” and “Ministerio de Sanidad del Gobierno de España” (FIS 09/1944) and TERCEL funds (FIS, Spain). We thank Carolyn Newey for editorial support and Dr. Rojas-Garcia for his insightful comments and Mireia Genebriera for technical support.
Author Disclosure Statement
The authors have nothing to disclose.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.