
Abstract
Ryk is an atypical transmembrane receptor tyrosine kinase that has been shown to play multiple roles in development through the modulation of Wnt signaling. Within the developing ventral midbrain (VM), Wnts have been shown to contribute to the proliferation, differentiation, and connectivity of dopamine (DA) neurons; however, the Wnt-related receptors regulating these events remain less well described. In light of the established roles of Wnt5a in dopaminergic development (regulating DA differentiation as well as axonal growth and repulsion), and its interaction with Ryk elsewhere within the central nervous system, we investigated the potential role of Ryk in VM development. Here we show temporal and spatial expression of Ryk within the VM, suggestive of a role in DA neurogenesis and axonal plasticity. In VM primary cultures, we show that the effects of Wnt5a on VM progenitor proliferation, DA differentiation, and DA axonal connectivity can be inhibited using an Ryk-blocking antibody. In support, Ryk knockout mice showed reduced VM progenitors and DA precursor populations, resulting in a significant decrease in DA cells. However, Ryk−/− mice displayed no defects in DA axonal growth, guidance, or fasciculation of the MFB, suggesting other receptors may be involved and/or compensate for the loss of this receptor. These findings identify for the first time Ryk as an important receptor for midbrain DA development.
Introduction
W
Within the developing ventral midbrain (VM), Wnts have been shown to modulate neurogenesis, dopamine (DA) differentiation, and DA axon morphogenesis. In particular, Wnt1-mediated canonical signaling, through the stabilization of β-catenin, has been shown to regulate proliferation of DA progenitors [8,9], while Wnt5a, via activation of Rac1 in the Wnt/PCP pathway, plays important roles in DA differentiation and connectivity of DA neurons [8,10 –12]. Despite the extensive evidence for Wnts in VM development, until recently, little attention has been paid to the receptors transducing these effects.
In 2007, Rawal et al. demonstrated that the majority of Fz receptors were expressed within the developing VM [13]. Supportive evidence, by us and others, showed specific roles for both Fz3 and Fz6 in the birth and connectivity of DA neurons [14 –17]. Additionally, loss of LRP6, ubiquitously expressed within the central nervous system (CNS) [18], was shown to result in a delayed onset of DA precursor differentiation in the VM [19].
The first evidence of Ryk function in neural development was observed in loss-of-function studies in Drosophila, where Derailed (the drosophila homolog) was shown to repel developing axons [20], a role that was later attributed to Wnt5 binding [21]. Within the mammalian brain, Ryk (commonly through Wnt5a interactions) has similarly been shown to regulate neural connectivity, promoting axon extension of dorsal root ganglia, cortical and commissural axons [22], as well as inducing chemorepulsion of the retinotectal [23], cortical [24], and commissural [25] axons. As a result, Ryk −/− mice show major disruptions to axonal tracts, including defasciculation of cortical axons through the corpus collosum [24] and the commissural tract [26]. Within the developing nervous system, Ryk has also been shown to be necessary for neurogenic events, regulating the induction of GABAergic neurons and inhibition of oligodendrogenesis through the interaction with Wnt3a in the ventral forebrain [27], and promoting cortical neurogenesis and differentiation at the cortical plate after nuclear translocation of the receptor [28].
In light of these findings, here we examined the temporal and spatial expression of Ryk within the developing VM. Additionally, we assessed the potential for Ryk to mediate the known Wn5a functions in the birth and connectivity of midbrain DA neurons, using the Ryk-blocking antibody as well as the Ryk-deficient mouse.
Materials and Methods
Animals
All experiments conformed to the Australian National Health and Medical Research Council's published Code of Practice for the Care and Use of Animals for Scientific Purposes, and the experiments were approved by the Florey Neuroscience Institutes Animal Ethics Committee or the Peter MacCallum Cancer Centre Animal Experimentation Ethics Committee.
For primary VM cultures, embryos were isolated from time-mated Sprague-Dawley rats. For quantitative polymerase chain reaction (PCR), embryonic tissue was isolated from time-mated Swiss mice or mice expressing green fluorescent protein under the tyrosine hydroxylase promoter (TH-GFP) [29]. Animals were time mated overnight, and visualization of a vaginal plug the following morning was taken as embryonic day (E) 0.5. Ryk −/− embryos [7] were collected at E11.5, E14.5, and E18.5.
In situ hybridization and immunohistochemistry in vivo
Embryos were isolated in ice-cold phosphate-buffered saline (PBS) and fixed in 4% paraformaldehyde (PFA), followed by overnight immersion in 30% sucrose in PBS. E11.5, E14.5, and E18.5 embryos were cryosectioned on either a sagittal or coronal plane at a thickness of 16 μm. In situ hybridization (ISH) was performed as previously described [30], using a DIG-labeled single-stranded RNA probe for Ryk (a kind gift from Professor Yimin Zou, University of Chicago). After hybridization, the tissue was again fixed using 4% PFA before immunohistochemistry for tyrosine hydroxylase (TH; the rate-limiting enzyme in dopamine synthesis and marker of DA neurons and neurites).
Immunohistochemistry was performed on 4% PFA-fixed cultures and slide-mounted sections as previously described [31]. The following primary antibodies were used: rabbit anti-TH (1:500; PelFreez), sheep anti-TH (1:500; PelFreez); mouse anti-βIII-tubulin (1:1500; Promega), rabbit anti-Nurr1 (1:200; Santa Cruz), and goat anti-Sox2 (1:200; R&D Systems). Appropriate fluorophore-conjugated (DyLight 488, 549, and 694) secondary antibodies or a biotinylated secondary antibody together with the Vector Laboratories ABC immunoperoxidase kit was used for visualization.
Quantitative real-time PCR
The temporal expression of Ryk within the developing midbrain dopamine system was examined by quantitative real-time PCR (qPCR). The VM, dorsal midbrain, and rest of embryo were isolated from E10.5, E11.5, E14.5, and P1 mice. For each developmental stage (E10.5, E11.5, and E14.5), the tissue was pooled from >4 embryos, and 4 independent litters were isolated for replicates. P1 tissue was isolated from 4–5 animals. Additionally, qPCR was used to determine if Ryk was expressed on DA neurons, or other cell populations, within the VM. For this purpose, the VM was isolated and dissociated from the E12.5 TH-GFP embryos. Subsequently, GFP+ DA neurons were sorted from other cell populations within the VM (GFP−), using fluorescence-activated cell sorting [31].
After tissue isolation, total RNA was extracted using the RNeasy Microkit (Qiagen). RNA was reverse transcribed using Superscript III First-Strand Synthesis Supermix for qRT-PCR (Life Technologies), and qPCR was carried out using the SYBR GreenER™ qPCR SuperMix Universal (Life Technologies) on an ABI7700 sequence detection system (Applied Biosystems) using the comparative ΔΔCT method [32]. Oligonucleotide sequences were as follows:
HPRT forward, 5′-CTTTGCTGACCTGCTGGATT-3′
HPRT reverse, 5′-TATGTCCCCCGTTGACTGAT-3′
Ryk forward, 5′-CGCTCTGTCCTTTAACCTGC-3′
Ryk reverse, 5′-CCAGTTCAATCCTTTTCATGC-3′
Wnt5a forward, 5′-AATAACCCTGTTCAGATGTCA-3′
Wnt5a reverse, 5′-TACTGCATGTGGTCCTGATA-3′
TH forward, 5′-CAGCTGGAGGATGTGTCTCA-3′
TH reverse, 5′-GGCATGACGGATGTACTGTG-3′
Generation of Ryk-blocking antibody (RWD1)
An Ryk-blocking antibody was generated to test the function of the receptor in dopaminergic-enriched VM primary cultures. A phage-display antibody library screen was performed by CD BioSciences, Inc. The screening protein, purified hRYKWD.Fc, was used on a human single-chain fragment variable (scFv)-naïve phage display library in three rounds of screening. The final (third) round of screening was performed using recombinant mouse Wnt3a protein (R&D Systems) in competitive ELISA.
The cDNAs of the heavy-chain and light-chain-variable regions from the phage clone able to compete with Wnts for binding to hRYKWD.Fc (scFv3) were subcloned into two separate vectors: one encoding human IgG-κ light-chain (pSTDLH3.RYK) and the other human IgG1-γ heavy-chain (pSTDHH3.RYK; CD BioSciences, Inc.). Large-scale purification of the anti-RYK IgG antibody, named RWD1, was performed using a stable cell line. CHO-K1 cells were transfected with pSTDHH3.RYK and pSTDLH3.RYK (6:4 ratio) using Lipofectamine 2000 (Life Technologies), and selection was applied after 24 h by adding 300 μg/mL zeocin (Life Technologies) to the medium. Colonies were picked after 7–9 days. The RWD1/CHO stable cell line was seeded into a medium FiberCell cartridge, 20 kDa (FiberCell Systems), using DMEM + 10% FBS + 200 μg/mL zeocin, and then maintained in DMEM + 10% CDM-HD serum replacement (FiberCell Systems) + 150 μg/mL zeocin. The extracapillary space medium was collected every 2–3 days.
Antibodies were purified using a Protein A sepharose column (GE), followed by IgG elution with 100 mM glycine, pH 2.8, into tubes containing 1:5 volume of 1 M Tris, pH 8.0. Antibody fractions were buffer-exchanged with PBS and concentrated using Amicon Ultra centrifuge filters (Millipore).
Immunoblotting
The activity of the Ryk-blocking antibody RWD1 was verified by coadministration with Wnt5a to the clonal VM DA neuronal progenitor cell line SN4741, as previously described [14]. In brief, SN4741 cells were cultured overnight in the absence of serum. The following day, the cells were treated with 10 μg/mL RWD1 (or human IgG, 10 μg/mL; R&D Systems; i.e., control) 1 h before the addition of recombinant mouse Wnt5a (300 ng/mL; R&D Systems). After 3 h, the cell lysates were collected, and immunoblotting was performed as previously described [33]. The following primary antibodies were used: rabbit anti-Dishevelled-2 (Dvl-2 1:500; Santa Cruz Biotechnology) and mouse anti-β-actin (1:3000; Sigma).
VM primary cultures
E13.5 represents a critical time in the development of the VM DA neurons in rats, as it corresponds to a period of DA neurogenesis as well as the onset of axonal growth and guidance. Consequently, this age was utilized to examine the ability of the Ryk-blocking antibody RWD1 to antagonize the effects of Wnt5a on the birth and axonal growth of primary VM DA neurons. Immunoblotting results demonstrated that 10 μg/mL of RWD1 was able to prevent Wnt5a- (150 ng/mL) induced hyperphosphorylation, and was therefore utilized in subsequent primary cultures.
The VM of E13.5 rat embryos were microdissected in a chilled L15 medium (Life Technologies). The isolated VMs were enzymatically dissociated in HBSS containing 0.05% trypsin and 0.1% DNase for 15 min at 37°C. Cells were subsequently centrifuged and resuspended in a serum-free N2 medium consisting of a 1:1 mixture of F12 and MEM supplemented with 15 mM HEPES buffer, 1 mM glutamine, 6 mg/mL glucose (Sigma-Aldrich), 1 mg/mL bovine serum albumin and N2 supplement (all purchased from Life Technologies). Cells were seeded at a density of 125,000 cells per well in a 48-well plate at 37°C, 5% CO2 with RWD1 antibody (or IgG) added 30 min before the addition of Wnt5a. After 72 h in culture, the cells were fixed with 4% PFA for 20 min before immunocytochemistry. All cultures were counterstained with Hoechst 33258 for 5 min (1:1000; Life Technologies). Cells immunoreactive for TH, βIII-tubulin, and Nurr1, and also stained with Hoechst, were counted from 10 fields of view in three wells per culture. Counts were obtained from four to five independent cultures. Neurite morphology of TH-immunoreactive neurons within the cultures was analyzed as previously described [14]. Assessments included the number of neurites per DA neuron, the number of neurite branches, length of the dominant neurite (axon), and total length of all neurites per neuron.
Explants
Three-dimensional collagen matrix explants of the VM were prepared as previously described [34]. In brief, explants were isolated from E12.5 embryos and embedded in close proximity to (∼300 μm) aggregates of HEK293 cells transiently transfected with the expression vectors for Wnt5a or mock-transfected as control. Ryk antibody (25 μg/mL; a kind gift of Yimin Zou, University of California, [23]) or IgG (Calbiochem) was added in an effort to antagonize the chemorepulsive effects of Wnt5a on DA neurites. Explants remained in culture for 56–58 h, before fixation and coimmunostaining for TH and βIII tubulin (TUJ1). The length of the 20 longest TH+ neurites emanating from the proximal and distal quadrants of each explant was measured, and the proximal/distal ratio calculated [34].
Analysis of Ryk−/− mice
All Ryk −/− embryos were compared to the littermate controls (E11.5, n=5–6/genotype; E14.5, n=6; E18, n=3). Cell counts within the E11.5 and E14.5 developing VM were estimated using the previously described fractionator methods [35 –37], with immunostaining (for sox2, nurr1, or TH) used to delineate the region of interest. Sox2 was used to label the VM stem cell/progenitor population seen lining the ventricle; Nurr1 labeled the dopaminergic precursor population within the intermediate zone of the VM, while TH was used to identify differentiated DA neurons within the marginal zone. Cell counts were derived by means of a grid program (StereoInvestigator software), through which a systematic sample of the area occupied by the cells was made from a random starting point. Counts were made at regular predetermined intervals (x=30 μm; y=30 μm), and an unbiased counting frame of known area (8 μm×8 μm=64 μm2) was superimposed on the image of the tissue sections, viewed under a 100×N.A. 1.30 oil immersion objective. Counts were made from 14-μm-thick 1:10 serial sections.
At E18, chromogenic staining of TH+ neurons was performed to quantify the number of TH+ cells within the VM, volume of the medial forebrain bundle (MFB), DA fiber density within the MFB, and dopaminergic striatal innervations, using previously described methods [14]. The volume of the MFB was estimated by delineating the area of the TH+ fiber bundle in the first section rostral to the midbrain TH+ neurons until the final section before the arrival of TH+ fibers in the striatum. The MFB was delineated in ∼7 sections (16-μm thickness, 1:10 series, i.e., ∼1120 μm in length) from each brain, with the area and total volume estimated.
The number of dopaminergic fibers (TH+) in the MFB and the density of TH+ varicosities in the lateral striatum were estimated using the fractionator methods. The density of TH+ fibers was assessed at a point along the MFB, 320 μm rostral to the midbrain DA neurons. TH+ fibers were counted at regular predetermined intervals (x=50 μm; y=50 μm), using a counting frame of known area (7 μm×7 μm=49 μm2), and viewed under a 100×N.A. 1.30 oil immersion objective. The number of TH+ fibers was counted in 16-μm-thick 1:10 serial sections.
TH+ varicosities in the lateral 600 μm of the striatum (Fig. 6F) were counted from 16-μm serial sections, 1:10 series, with four sections sampled from each striatum. TH+ varicosities were counted at regular predetermined intervals (x=150 μm; y=150 μm) using an unbiased counting frame of known area (6 μm×6 μm=36 μm2). TH+ varicosities were identified as predominantly round swellings in association with axonal processes. TH+ varicosity counts were expressed as terminal density, with comparisons made between Ryk+/+ and Ryk −/− mice. For all cell and fiber counts, the coefficients of error (CE) and coefficients of variance (CV) were calculated as the estimates of precision, and values of <0.1 were accepted [35,36,38,39].
Statistical analysis
One-way ANOVA with Tukey's post-hoc test or Student's t-test was used to identify statistically significant changes. Statistical significance was set at a level of P<0.05. Data represent mean±SEM.
Results and Discussion
Midbrain dopaminergic neurons play critical roles in the control of voluntary movement and the regulation of emotion- and reward-related behaviors. Further, a number of neurological disorders are associated with disturbances of dopaminergic neural networks. Consequently, understanding the birth and connectivity of these neurons may shed new light on understanding disease etiology and aid in the development of new therapies. In this regard, midbrain development has been shown to involve a tightly orchestrated sequence of molecular events to ensure the birth and appropriate connectivity of these dopamine neurons. The Wnt family of proteins has been shown to modulate DA proliferation, differentiation, and neuronal connectivity. Of particular interest, Wnt5a regulates proliferation of DA precursors, their differentiation, and, finally, appropriate trajectory and forebrain targeting. While an increasing number of receptors have been attributed to these functions, here we reveal that several Wnt5a-mediated effects on DA progenitors are regulated via the atypical tyrosine kinase receptor, Ryk.
Ryk shows temporal and spatial expression indicative of a role in VM development
To ascertain whether Ryk plays a role in the developing VM, we examined the expression of the receptor during periods of DA neurogenesis and axon morphogenesis. Within the developing mouse VM, dopaminergic neurogenesis commences at approximately embryonic day (E) 10.5, peaking at E12.5, and finally ceasing at E14.5. Shortly after birth of these neurons, DA neurite extension commences (approximately E11.5). In accordance with these developmental events, we examined Ryk expression, by ISH, at E11.5 and E14.5. At E11.5, during ongoing DA neurogenesis and initiation of neurite outgrowth, Ryk was strongly expressed in the VM. Expression was observed throughout the layers of the neural epithelium, extending from the proliferative ventricular zone to the intermediate progenitor zone and into the marginal zone (Fig. 1A–C). By E14.5, expression was notably downregulated within the ventricular and intermediate zone, yet remained within the marginal zone, overlapping with TH+ DA neurons (Fig. 1D–F).
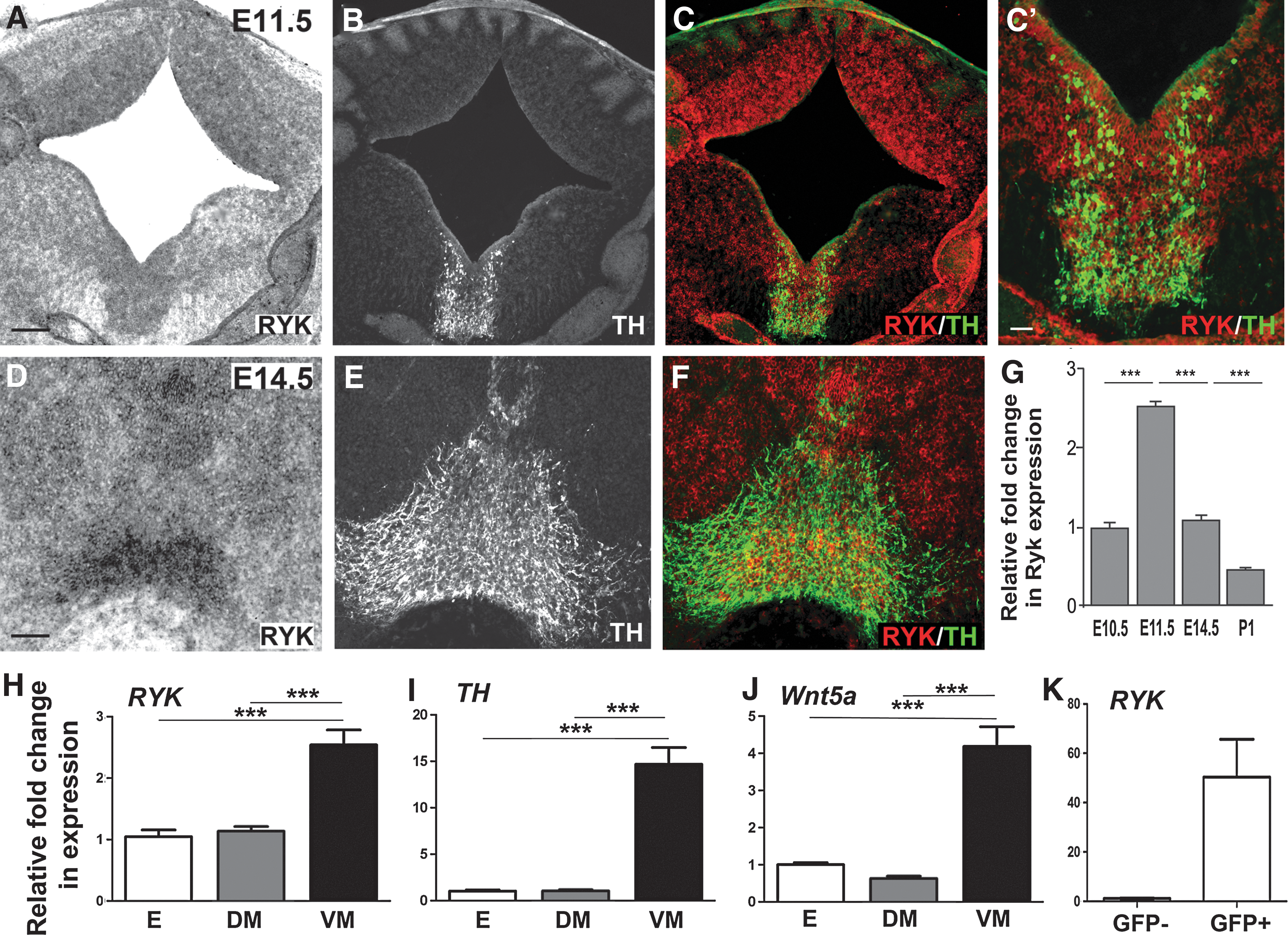
Ryk is temporally and spatially expressed within the developing ventral midbrain (VM), suggestive of a role in dopaminergic neurogenesis and/or axon morphogenesis.
The expression of Ryk within the VM was confirmed using quantitative PCR. We examined the relative temporal expression levels from the onset of DA neurogenesis (E10.5) until establishment of the dopaminergic pathway (postnatal day 1; P1). Results showed that Ryk expression peaked at E11.5 and was downregulated by P1 (Fig. 1G). At E11.5, Ryk expression was significantly higher in the VM than either the dorsal midbrain (DM) or the entire embryo (E; Fig. 1H), similar to both TH (Fig. 1I) and Wnt5a (Fig. 1J) expression.
To establish whether Ryk was a receptor regulating Wnt responsiveness of DA neurons, we additionally examined whether Ryk was expressed on DA neurons or other cell populations within the developing VM. We utilized the TH-GFP mouse to isolate dopaminergic neurons (GFP+ cells) from other cell types within the VM (i.e., GFP− cells) by fluorescence-activated cell sorting. At E12.5, quantitative PCR showed that the GFP+ fraction contained the majority of the Ryk transcript (Fig. 1K), indicating that the Ryk receptor was strongly expressed on VM TH+ DA neurons, and may therefore be a candidate receptor mediating Wnt signaling in DA neurons.
Ryk-blocking antibody, RWD1, is capable of antagonizing Wnt signaling in a dopaminergic cell line
To examine the function of Ryk on VM DA neurons, we used an anti-Ryk monoclonal antibody (subsequently referred to as RWD1) capable of binding to the extracellular domain of Ryk and preventing Wnt-mediated signaling. To confirm the specificity of RWD1, we demonstrated that treatment with Wnt5a protein (150 ng/mL) alone induced downstream phosphorylation of intracellular Dishevelled-2 in a dopaminergic cell line, SN4741 (observed as a band shift by immunoblotting; Supplementary Fig. S1; Supplementary Data are available online at
RWD1 antibody prevents Wnt5a-mediated DA neurogenesis
Given the interaction of Wnt5a and Ryk elsewhere within the CNS, next we examined whether Ryk was the receptor responsible for mediating the effect of Wnt5a on DA progenitors. Therefore, we treated E13.5 dopaminergic neuron-enriched rat VM primary cultures with Wnt5a in the presence and absence of the RWD1 (or control IgG) antibody. Quantification of total cells in culture (Hoechst+ nuclei) revealed no significant difference across the treatments (Fig. 2A–D, Q), indicating that RWD1 was not toxic to the cells, nor had an effect on overall cell proliferation. Additionally, treatments had no effect on the number of TUJ1+ neurons in culture (Fig. 2E–H, R).
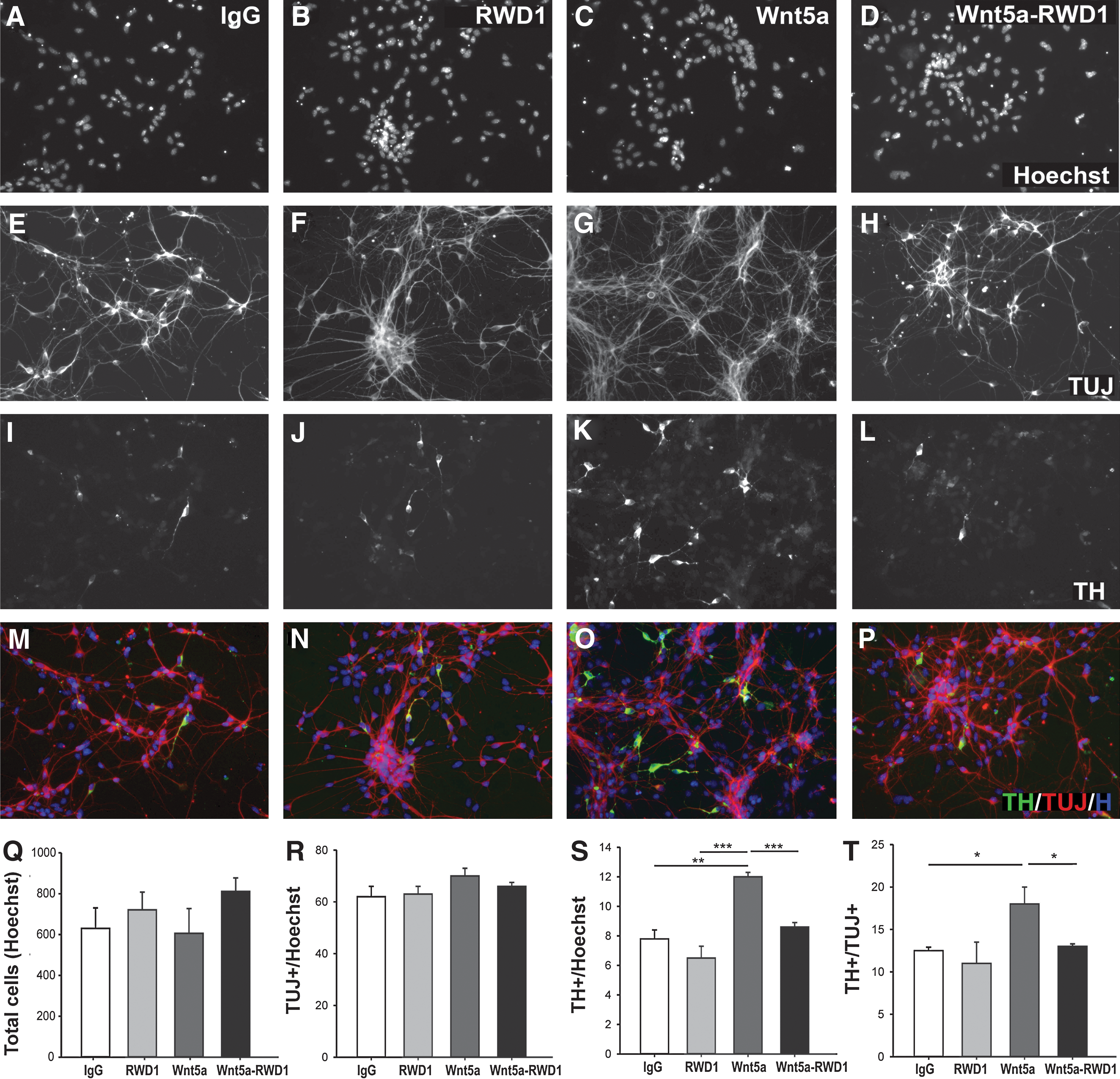
Ryk mediates Wnt5a-induced increases in DA neurons in vitro. Photomicrographs of Hoechst-labeled cells from E13.5 rat VM cultures treated with
Subsequently, we examined the number DA neurons in the culture. Under control conditions, RWD1 had no effect on the number of TH+ cells (Fig. 2I, J, S). As expected, and previously reported [8,10], the addition of Wnt5a resulted in a significant increase in TH+ DA cells compared to control (IgG), as a proportion of total Hoechst-labeled cells (12%±0.4% and 8%±0.6%, respectively; Fig. 2K, S) and TUJ1+ neurons (17%±1.0% and 12%±0.3%, respectively; Fig. 2T). Exposure of the cultures to RWD1 in addition to Wnt5a abolished this increase in TH+ neurons, with DA neuronal counts not significantly different from those observed in the control-treated cultures (Fig. 2L, M–P, S, T).
Wnt5a has previously been described to expand the Nurr1+ dopaminergic precursor pool and promote differentiation of this population of cells from Nurr1+TH− into Nurr1+TH+ dopaminergic neurons [8,10]. We wished to determine whether the effects of the Ryk-blocking antibody on DA neuronal numbers were a consequence of reduced precursor cells and/or failed differentiation. Here, Wnt5a treatment resulted in a modest increase in the number of Nurr1+ cells in culture, with a trend for RWD1 in the presence of Wnt5a to reduce this effect (Fig. 3A–D, I). In support of previous findings [8,10], dopaminergic differentiation (i.e., the proportion of TH+ cells relative to Nurr1+ cells) was significantly increased after Wnt5a application, an effect that could be notably reduced by coadministration of Wnt5a and RWD1 (Fig. 3E–H, J). These findings suggest that Wnt5a may signal through Ryk to regulate DA differentiation.

Ryk regulates the differentiation of Nurr1+ dopaminergic progenitors. Photomicrographs of Nurr1-immunoreactive cells from E13.5 rat VM cultures treated with
Within the VM, it is plausible that Ryk may also be mediating other Wnt signaling. Previously, Wnt1 and Wnt2 have also been shown to influence the VM DA population in vitro and in vivo [8,9,40], predominantly by increasing the precursor population available for differentiation. However, within the present cultures, the Nurr1+ precursor population remained unchanged, suggesting that RWD1 was specifically blocking Wnt5a differentiation, rather than altering proliferation or cell survival. Supporting this level of specificity, blocking of Ryk (in the absence of Wnt5a) had no effect on the other cell populations within the VM (data not shown).
Ryk mediates Wnt5a-induced axon morphogenesis
Recently, we described the ability of Wnt5a to regulate dopaminergic axon growth and guidance [14,17]. Additionally, Wnt5a, through Ryk binding, has been shown to mediate neurite outgrowth and guidance events elsewhere within the CNS [41 –43]. To establish whether Wnt5a signals through Ryk to alter the DA neurite morphology, E13.5 VM rat primary cultures were treated with Wnt5a (100 ng/mL)±RWD1 (or control IgG). As previously observed, Wnt5a significantly reduced the neurite number and branching of DA neurons, as well as promoted neurite extension. While RWD1 alone had no effect on the DA neurite morphology, it was capable of ablating the effects of Wnt5a on the neurite number and length (Fig. 4A–G), suggesting that Ryk, at least in part, is responsible for mediating the effects of Wnt5a on DA axon morphogenesis.

Ryk mediates Wnt5a-induced axonal growth and guidance of dopaminergic axons. Images of TH-immunoreactive cells from the E13.5 rat VM cultures, depicting dopaminergic neurite morphology, after treatment with
In addition, Wnt5a provides chemotaxic cues for DA neurites, causing repulsion of TH+ axons emanating from VM explants in vitro, a mechanism mediated via the Wnt/PCP pathway [14,17]. In other CNS systems, the Ryk receptor has been linked with Wnt-mediated chemorepulsion; hence, we wished to determine whether Ryk was also associated with Wnt5a repulsion of the midbrain DA neurites during development. Mouse VM explants (E12.5) were cocultured with either mock-transfected or Wnt5a-overexpressing cell aggregates, and the length of TH+ fibers in the distal (D) and proximal (P) quadrants of the explant, with respect to the cell aggregate, was determined, as previously described [17]. TH+ neurites from explants cocultured with mock-transfected cell aggregates radiated out in all directions from the explant, showing a proximal-to-distal ratio of neurites close to 1 (1.02±0.03, n=9; Fig. 4H, L). In VM explants cultured with Wnt5a-overexpressing cell aggregates, the majority of TH+ neurites were repelled from the proximal aspect of the explant, with a proximal-to-distal ratio of 0.74±0.03 (n=9, P<0.01; Fig. 4I, L). This effect was inhibited by the addition of Ryk antibody (0.95±0.04, n=9, Fig. 4K, L), confirming the involvement of Ryk in DA axon chemotaxis.
Within the developing cortex, the seemingly opposing effects of Wnts controlling growth and repulsion in the same cell populations have been shown to be mediated by different calcium signaling pathways through Ryk acting independently (to control outgrowth) and Ryk acting in conjunction with Fz (to achieve chemorepulsion) [44,45]. It remains to be determined whether similar calcium signaling pathways via Ryk alone or Ryk/Fz dimers mediate these opposing growth and repulsion effects on DA axons.
Ryk-mediated signaling affects VM progenitors and TH+ dopaminergic populations in vivo
In an effort to support our in vitro findings demonstrating that Ryk mediates Wnt5a effects on DA neurons, we examined the midbrain dopaminergic neuron population, MFB, and striatal innervation in Ryk-deficient mice.
As Wnts have been shown to influence VM proliferation and differentiation, we examined the number of Sox2+ stem cells (within the ventricular zone, VZ), the number of Nurr1+ precursors (in the intermediate zone, IZ) and the number of TH+ DA neurons (in the marginal zone, MZ) during a period of ongoing VM DA neurogenesis (E13.5), and the number of TH+ cells within the VM upon completion of DA neurogenesis (E18.5). We observed a significant decrease (17%) in the number of Sox2+ progenitor cells within the ventricular zone of the VM of Ryk-deficient mice (Ryk +/+ 10180±370 and Ryk −/− 8471±127; Fig. 5A, D, E), as well as a significant reduction in the Nurr1+ population (Ryk +/+ 59213±2135 and Ryk −/− 44213±1195, 25% decrease; Fig. 5B, D, E). The decrease in the precursor pool correlated with a similar decrease in TH+ DA neurons (Ryk +/+ 22973±1262 and Ryk −/− 18328±770, 20% decrease; Fig. 5C, D–G).
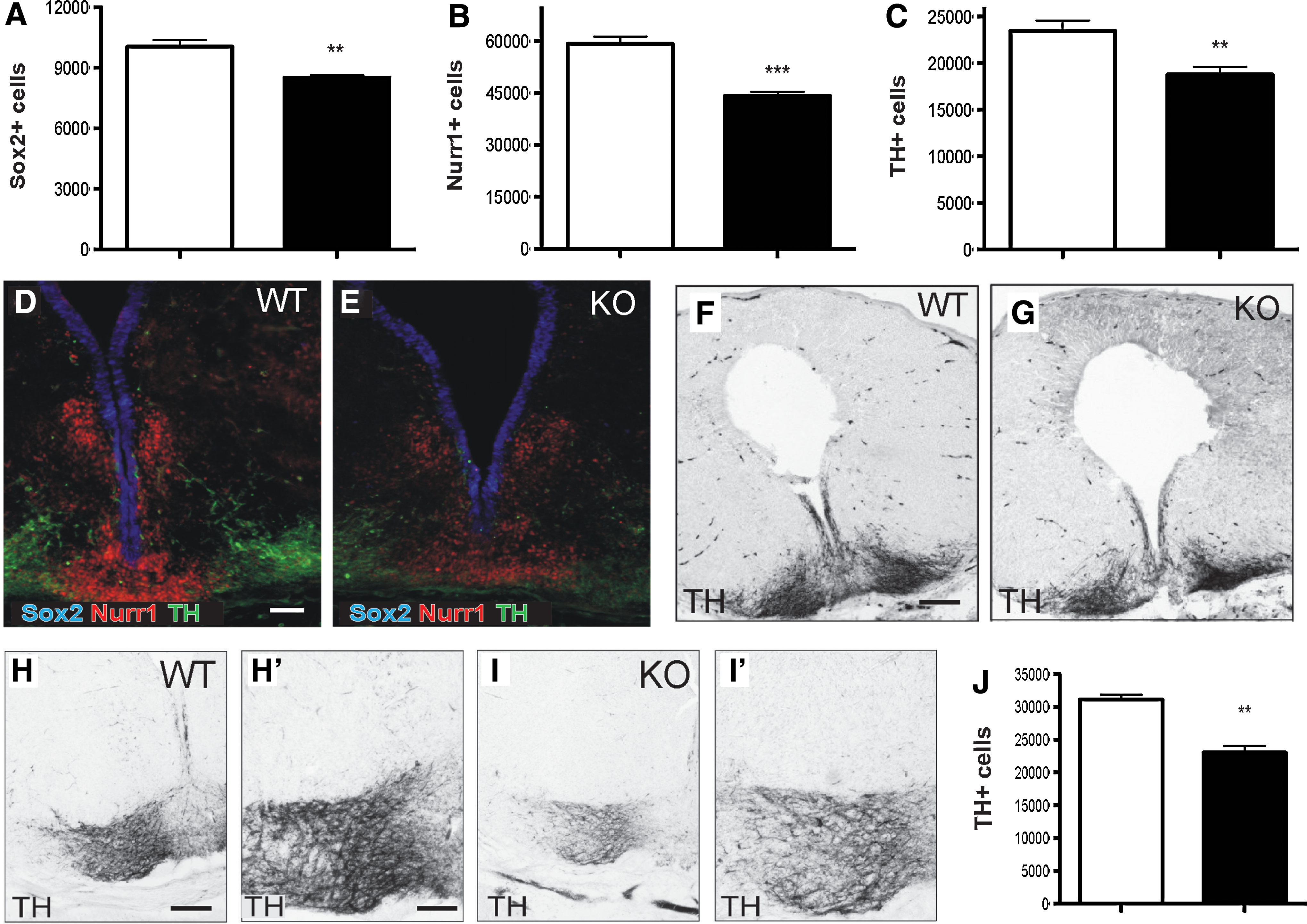
Ablation of the Ryk receptor affects progenitor, precursor, and mature DA neuronal populations in the VM.
Unlike many other Wnt-related deficient mice (including Wnt5a [11] and LRP5/6 [19]), where developmental compensation normalizes TH counts, at E18.5, Ryk−/− mice maintained a significant decrease in the number of DA neurons. At this later age, there was a 32% decrease in the TH+ numbers (Ryk +/+ 31377±719, and Ryk−/− 21493±1043; Fig. 5H–J), likely reflecting the 25% decrease in Nurr1+ cells seen at E14.5 as well as failure of Nurr1+ progenitors to differentiate. These findings demonstrate that Ryk is responsible, at least in part, for the regulation of DA progenitor proliferation and differentiation. They also suggest that Wnts other than Wnt5a, such as Wnt1 and Wnt2, which have been shown to regulate DA neurogenesis, may also signal through this receptor. Further studies examining Ryk antagonism of exogenous Wnt1 and the phenotype of double-knockout mice (such as Wnt5a–Ryk, Wnt1–Ryk, or other Wnts–Ryk) may shed more light on the involvement of this receptor in VM neurogenesis.
Ryk−/− mice show normal midbrain dopaminergic axonal development and striatal targeting
The Ryk −/− mouse has previously been reported to show major defects in axonal guidance and axon fasciculation, particularly of cortical axons traversing the corpus callosum [24] and of corticospinal tract axons [25]. In both instances, these effects were shown to be mediated by the interactions with Wnt5a. Further, we previously reported defasciculation of the TH+ fibers within the MFB in Wnt5a −/− mice, as well as increased fiber density within the bundle and premature entry into the striatum, indicative of inappropriate neurite number, branching, and loss of axonal stalling before striatal targeting [14]. Added to this, here we show that Ryk mediates Wnt5a effects on the DA neurite length in vitro. We therefore wished to examine the morphology of the developing midbrain dopaminergic pathways in Ryk −/− mice, and to assess whether there were similarities to the Wnt5a −/− mouse.
Wild-type and littermate Ryk −/− mice were examined at E18.5, given the postnatally lethal phenotype of these mice [7], and that by this age, DA neurons have extended their axons along the MFB and begun to innervate the lateral ganglionic eminence/striatum [46]. Quantification revealed that the volume of the MFB in Ryk −/− mice was not significantly different from Ryk +/+ littermate controls (1.68×108±0.426×107 μm3 and 1.55×108±0.511×107 μm3, respectively; Fig. 6A, E, G, I). Therefore, the defasciculation phenotype present in the Wnt5a −/− mouse is not reflected in the Ryk mutant, indicating that either this receptor is not responsible for fasciculation of the MFB or other receptors present on the DA neurons are capable of compensating.

Ryk-deficient mice maintain normal forebrain targeting and innervations.
Next, we examined the effect of Ryk ablation on the number of TH+ axons traversing along the MFB. Stereological examination of the fiber density within the MFB showed no significant difference between Ryk knockout and wild-type mice (0.124±0.009 and 0.127±0.016 TH+ fibers/μm3, respectively; Fig. 6B, G, G′, I, I′). Similarly, no difference was observed in the density of TH+ synaptic varicosities within the ventrolateral striatum (Ryk −/− mice 8.12×10−3±0.36×10−3 terminals/μm3, Ryk +/+ 7.85×10−3±0.44×10−3 terminals/μm3; Fig. 6C, F, H, J). However, in light of the 25% reduction in TH+ cells at this age, Ryk −/− mice showed increased TH+ fibers/TH+ cells compared to wild-type mice (0.95±0.11 and 0.74±0.12, respectively, data not shown) and significantly increased TH+ varicosities/TH+ cells (29.35±1.70 and 21.97±0.73, respectively, P=0.016, Fig. 6D). Hence, these findings demonstrate that while the number of TH+ DA neurons was reduced, Ryk−/− mice were able to compensate through increased axonal branching and varicosities to enable normal levels of forebrain innervation.
Interestingly, while an Ryk-blocking antibody prevented Wnt5a-induced chemorepulsion of TH+ neurites in vitro (Fig. 4), TH+ axons in Ryk−/− mice were capable of following the appropriate trajectory through the MFB and innervating the target striatum, with no evidence of misrouted TH+ fibers. Of note, other Wnt-related receptor knockouts have similarly showed no, or only modest, phenotypes. In the case of Fz3 −/−; Fz6−/− double mutants, the mDA neuronal deficits were significantly more profound than in the single mutants, where Fz6 −/− mice showed no evident phenotype on the mDA cell number, and the Fz3−/− mice, while missing the nigrostriatal tract, only had a transient decrease in the mDA cell number [15,16]. Given that we have previously shown that Fz3 influences Wnt5a-induced neurite outgrowth in culture [14,17], and that neither Fz3 nor Ryk appeared to be involved in the effects of Wnt5a on neurite number and branching, it is possible that Ryk−/− mice show limited changes in path finding and innervation of the striatum due to compensation by other Fzs or Ror2. In this regard, the Ror2 receptor was previously shown to act as a Wnt5a receptor [5], where it regulates convergent extension movements during gastrulation [47]. Thus, future studies examining the role of other Wnt receptors and the generation of Ryk–Ror2 or Ryk–Fz double mutants may elucidate their roles within the DA populations.
While Ryk lacks intrinsic catalytic tyrosine kinase activity, findings have revealed that this receptor is capable of signaling via multiple mechanisms, including the use of canonical pathway components and targets (including Dishevelled and Tcf/Lef [22,48]), recruitment of catalytically active downstream effects (such as the Src kinases [49]), as well as translocation of the internal fragment of the receptor to the nucleus [28]. In the Drosophila, Derailed (the Ryk ortholog) is also capable of sequestering Wnt, thereby preventing the interaction with other receptors [49,50]. Hence, the Ryk receptor is capable of independently mediating Wnt signaling as well as functionally interacting, either by forming heterodimers (e.g., Fz/Ryk), or via downstream pathway components. In general, Wnt binding to Fz and the LRP coreceptors activates the Wnt/β-catenin canonical pathway [22], while Ryk and Ror2 (alone or as a dimer with Fz) are more commonly associated with β-catenin-independent Wnt signaling [47,51], via the Wnt/PCP pathway or the Wnt/Ca2+ pathway. The focus of the present study was to examine whether Ryk-mediated Wnt5a effects on DA development. Previous work, by us and others, has demonstrated that the effects of Wnt5a on DA neurons are mediated by downstream PCP signaling [52,53]. However, given that Ryk can signal via various Wnt-mediated pathways, it is possible and likely that this receptor has a number of other regulatory functions within the VM.
In summary, these findings identify Ryk as an important mediator of VM DA development. While we have attributed Ryk to a number of Wnt5a-related roles in VM development, a number of key questions remain. Specifically, studies that clearly differentiate between the effects attributable to Ryk mediation of Wnt1/2 and Wnt5a signaling would further demonstrate whether Ryk is necessary for both proliferation and differentiation. Further, the use of conditional Ryk−/− mice may yield clearer results by decreasing the capacity for developmental compensation, as would double-mutant mice of Ryk together with Wnt5a, Wnt1, or other Wnt-related receptors.
Footnotes
Acknowledgments
We thank Ms. Mong Tien for technical assistance. This research was supported by funding from the National Health and Medical Research Council (NHMRC), Australia. This study was performed within the framework of Dutch Top Institute Pharma project T5-207 and supported by the Dutch Parkinson Foundation (to RJP). B.D.B was supported by an Australian Postgraduate Award. C.R.B was supported by an NHMRC Peter Doherty Research Fellowship. C.L.P was supported by an NHMRC Career Development Award, and subsequently a Senior Medical Research Fellowship provided by the Viertel Foundation, Australia.
Author Disclosure Statement
The authors declare no conflict of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.