
Abstract
In present study, we report on bone marrow (BM) mesenchymal stem cells (MSCs) that are isolated from giant pandas. Cells were collected from the BM of two stillborn giant pandas. The cells were cultured and expanded in 10% fetal bovine serum medium. Cell morphology was observed under an inverted microscopy, and the proliferation potential of the cells was evaluated by counting cell numbers for eight consecutive days. Differentiation potentials of the cells were determined by using a variety of differentiation protocols for osteocytes, adipocytes, neuron cells, and cardiomyocytes. Meanwhile, the specific gene expressions for MSCs or differentiated cells were analyzed by RT-PCR. The isolated cells exhibited a fibroblast-like morphology; expressed mesenchymal specific markers such as cluster of differentiation 73 (CD73), SRY (sex determining region Y)-box 2 (SOX-2), guanine nucleotide-binding protein-like 3 (GNL3), and stem cell factor receptor (SCFR); and could be differentiated into osteocytes and adipocytes that were characterized by Alizarin Red and Oil Red O staining. Under appropriate induction conditions, these cells were also able to differentiate into neuroglial-like or myocardial-like cells that expressed specific myocardial markers such as GATA transcription factors 4 (GATA-4), cardiac troponin T (cTnT), and myosin heavy chain 7B (MYH7B), or neural specific markers such as Nestin and glial fibrillary acidic protein (GFAP). This study demonstrated stem cells recovery and growth from giant pandas. The findings suggest that cells isolated from the BM of giant pandas have a high proliferative capacity and multiple differentiation potential in vitro which might aid conservation efforts.
Introduction
M
Due to the self-renewing and pluripotent properties, MSCs have generated a great deal of interest for their potential use in regenerative medicine and tissue engineering, and medical researchers believe that stem cells therapy has the potential to dramatically change the treatment of human diseases [12]. Moreover, MSCs from endangered species may be potentially useful for rescuing the species from extinction because of the possibility of producing MSC-derived germ cells, which could be used in conjunction with assisted reproduction efforts to increase the size and diversity of the population in the future.
The giant panda (Ailuropoda melanoleuca) is one of the most widely recognized endangered species in the world due to habitat loss, degradation, and fragmentation, as well as disorders such as parasitic roundworm infections [13]. It has been estimated that only 1,600 individual animals survive in the wild, living in six isolated mountain ranges within China [14]. Due to the critical status of the giant panda, in the present study, we isolated MSCs from the BM (BM-MSCs) of giant pandas to investigate their proliferative characteristics and differentiation potentials, which may provide valuable knowledge for future studies on possible stem cells applications in the endogenous species.
Materials and Methods
Isolation and culture of MSCs
MSCs were collected from two stillborn giant pandas at the Chengdu Research Base of Giant Panda Breeding. All experimental procedures involving animals were approved by relevant administrative departments.
The isolation and culture of MSCs were performed according to the protocol described by Soleimani and Nadri [15] with minor modifications. Briefly, under sterile conditions, the femur and tibiae of the dead cubs were excised, with special attention made to remove all connective tissue attached to bones. BM was excluded from theses bones by flushing the BM cavity using a syringe with 5-gauge needle filled with a complete culture medium. The complete culture medium was composed of low-glucose Dulbecco's modified Eagle medium (LG-DMEM; Gibco), supplemented with 10% fetal bovine serum (FBS; Hyclone), 10 ng/mL epidermal growth factor (Invitrogen), 5 ng/mL basic fibroblast growth factor (bFGF; Invitrogen), 7.5 mM MGlutaMAX-1 (Invitrogen), 100 U/mL penicillin G (Gibco), and 100 μg/mL streptomycin (Gibco). The BM capacity was flushed repeatedly, and the marrow was flushed into a 45 mm petri dish. The harvested BM-MSCs were gently pipetted to break up cell clumps in order to obtain cell suspension. After homogenous cell suspension had been achieved, the cells were incubated at 37°C with 5% CO2. Forty-eight hours later, the nonadherent cells were removed by washing thrice with Dulbecco's Phosphate-Buffered Saline (Invitrogen) solution, and the adherent cells were cultured for an additional 5 days with medium changes every 2 days. The primary BM-MSCs were passaged with 0.25% trypsin-EDTA (Hyclone) when the cells reached about 80% confluence.
Proliferation determination
The second, fourth, and sixth passage of the BM-MSCs were used to investigate their proliferation potential. First, the BM-MSCs were suspended with 0.25% trypsin-EDTA, and they were centrifuged at 1,500 rpm for 5 min. The cells were resuspended in fresh medium and seeded in 24-well plates at a concentration of 0.5×104 per well. The number of the cells per three wells was counted every day for eight successive days. The population doubling time of the BM-MSCs in an exponential growth phase was calculated according to this growth curve.
RT-PCR analysis
Total RNA was extracted from the third-passage cells using RNApreppureCell/BacteriaKit (Tiangen Biotech Co., Ltd.) according to the manufacturer's instructions. The mRNA was reverse transcribed to cDNA using PrimeScript® RT reagent Kit (TaKaRa Bio Group) according to the manufacturer's instructions. Expressions of specific marker genes for BM-MSCs and induced differentiation into neuronal-like or cardiomyocyte-like cells were determined by using the system of TaKaRa Taq™ (TaKaRa Bio Group). The amplification cycling conditions were 94°C 30 s, 50°C–58°C 30 s, and 72°C 40 s for 35 cycles after initial denaturation at 94°C for 5 min. Primers used for the amplification are listed in Table 1.
CD73, cluster of differentiation 73; SOX-2, SRY (sex determining region Y)-box 2; GNL3, guanine nucleotide-binding protein-like 3; SCFR, stem cell factor receptor; GATA-4, GATA transcription factors 4; cTnT, cardiac troponin T; MYH7B, myosin heavy chain 7B; GFAP, glial fibrillary acidic protein.
Differentiation protocols
Osteogenic differentiation
To induce osteogenic differentiation, the fourth-passage cells were cultured with osteogenic differentiation medium that consisted of LG-DMEM supplemented with 10% FBS, 0.1 μM dexamethasone (Sigma-Aldrich), 10 mM β-glycerolphosphate (Sigma-Aldrich), and 50 μM ascorbic acid (Sigma-Aldrich). The induction lasted for 21 days with the medium changed twice a week. Untreated cells used as controls were maintained in LG-DMEM with 10% FBS. Formed mineralized matrix in the induced cultures was demonstrated by Alizarin red staining.
Adipogenic differentiation
To induce adipogenic differentiation, the fourth-passage cells were cultured with adipogenic differentiation medium that consisted of LG-DMEM supplemented with 10% FBS, 0.5 mM 3-isobutyl-1-methylxanthine (IBMX; Sigma-Aldrich), 1 μM dexamethasone (Sigma-Aldrich), 200 μM indomethacin (Sigma-Aldrich), and 10 μg/mL insulin (Sigma-Aldrich). The cells were treated for 21 days with the differentiation medium that was changed twice a week. Untreated cells used as controls were maintained in LG-DMEM with 10% FBS. Oil droplets secreted by induced cells were evaluated by Oil red O staining.
Cardiomyocyte-like cells differentiation
The differentiation of cardiomyocyte-like cells from the fourth-passage cells was performed by using a protocol described by Kumar et al. [16] with minor modifications. Briefly, subconfluent cells were treated with 10 μM 5-azacytidine for 24 h. Afterward, the cells were washed twice with Tyrode's balanced salt solution and cultured in 10% FBS complete medium without 5-azacytidine. The medium was changed twice a week until the termination of experiments at 4 weeks after the drug treatment. Cells without treatment that were used as control were maintained in LG-DMEM with 10% FBS during the induced experiments. Differentiations in cell morphological changes were observed under an inverted microscope (Olympus IX70), and images were captured by relevant software (Olympus DP25). Expressions of marker genes for myocardial cells were analyzed by using the RT-PCR.
Neuron-like cells differentiation
Neuronal-like cells differentiations were induced by using the protocol reported by Woodbury et al. [17] with minor modifications. The subconfluent cells (ie, the fourth-passage) were preincubated in LG-DMEM supplemented with 10% FBS, 1 mM β-mercaptoethanol (BME; Sigma-Aldrich), and 10 ng/mL bFGF (Invitrogen) for 24 h. The cells were then cultured for another 24 h in serum-free LG-DMEM supplemented with 2% dimethylsulfoxide (Sigma-Aldrich) and 200 μM butylated hydroxyanisole (Sigma-Aldrich). Cells without treatment were used as controls, which were maintained in LG-DMEM with 10% FBS during the induced experiments. Differentiations in cell morphological changes were observed under the inverted microscope, and images were captured by the software as described earlier. Expressions of marker genes for neuron cells were analyzed by using the RT-PCR.
Data analysis
All results were generated from at least three independent experiments and were presented as mean±SD. Statistical significance was determined using Student's t-test, and differences were considered significant when P<0.05.
Results
Morphology and gene expressions of undifferentiated cells
After the initial 48 h of primary culture, the cells sparsely attached to culture dishes displayed a spindle and triangle shape (Fig. 1A-a), and gradually grew to form colonies. By the fifth day, the cells became confluent, showing spindle-shaped cells with a whirlpool-like array (Fig. 1A-b). Subconfluent cells were treated with 0.25% trypsin-EDTA, and replaced at 1:3 under the same culture conditions. The subcultured cells behaved similar to the primary cultures, and could be steadily cultured to more than 10 successive passages (Fig. 1A-c). RT-PCR assay revealed that the cells were positive for cluster of differentiation 73 (CD73), guanine nucleotide-binding protein-like 3 (GNL3), and stem cell factor receptor (SCFR) (Fig. 1B), all of which are specific markers for MSCs or many types of stem cells [18,19]. In addition, the cells were found to be positive for SRY (sex determining region Y)-box 2 (SOX-2), which is a marker for pluripotent stem cells [20].
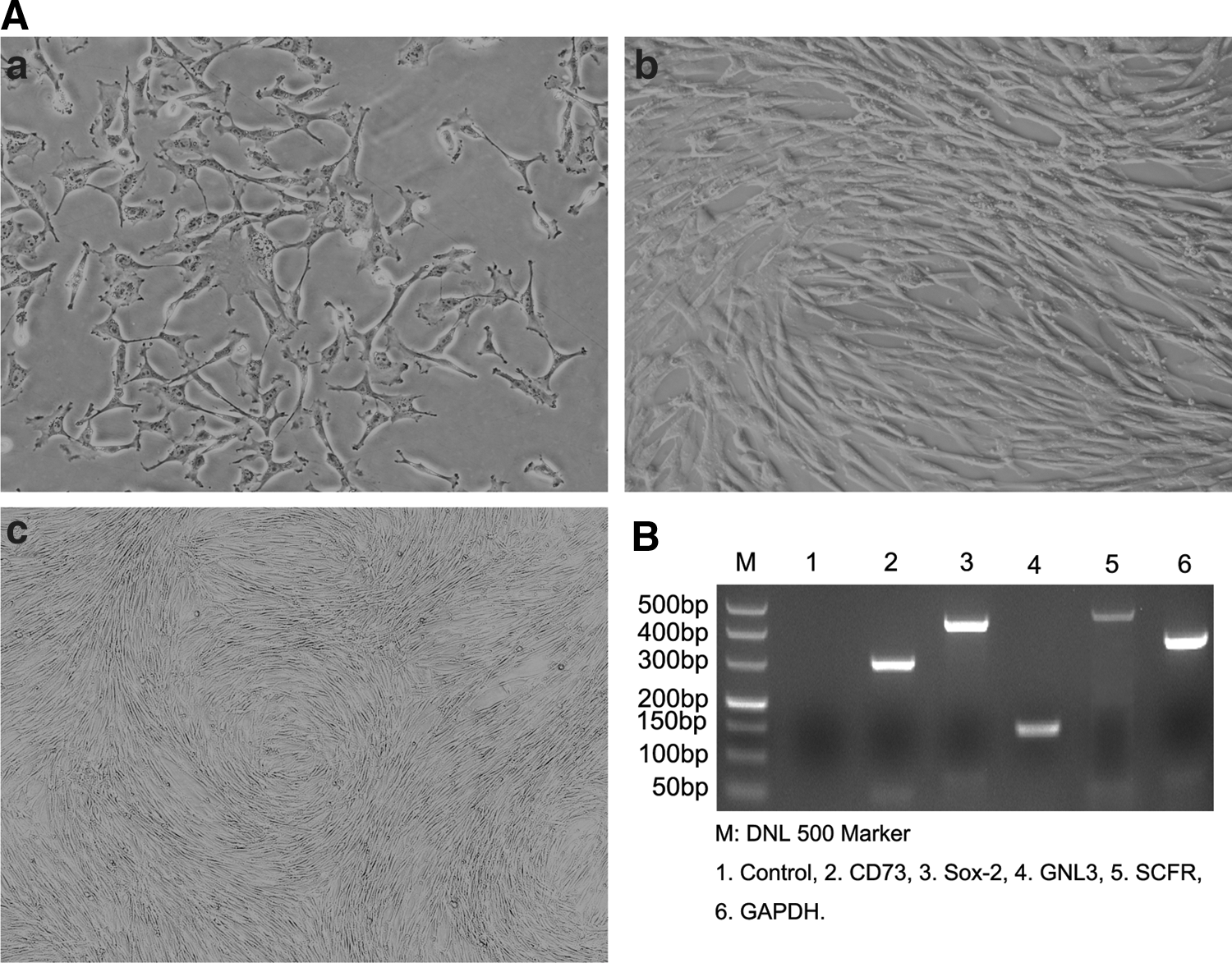
Morphologic and RT-PCR analyses of expressions of stem cells-specific genes.
Growth properties of the cells
The passage cells grew faster than the primary cells and generally undergo 3 days to reach 90% confluent rate after seeding. The cell growth curves of the second-, fourth-, and sixth-passage cells showed a typical S type. After re-seeding, the cells underwent 1–2 days of an adaptive phase, and began to expand rapidly by moving into a logarithmic phase of growth. After 5 days, cell counts reached their highest levels followed by a plateau phase (Fig. 2A). The extrapolated analysis of the accumulated cell number for the second, fourth, and sixth passage revealed that the second and fourth passages had greater proliferative capacity than the sixth passage (P=0.034) (Fig. 2B). These results indicated that the cells have a stem cell property to self-renew at a relatively higher rate [21].
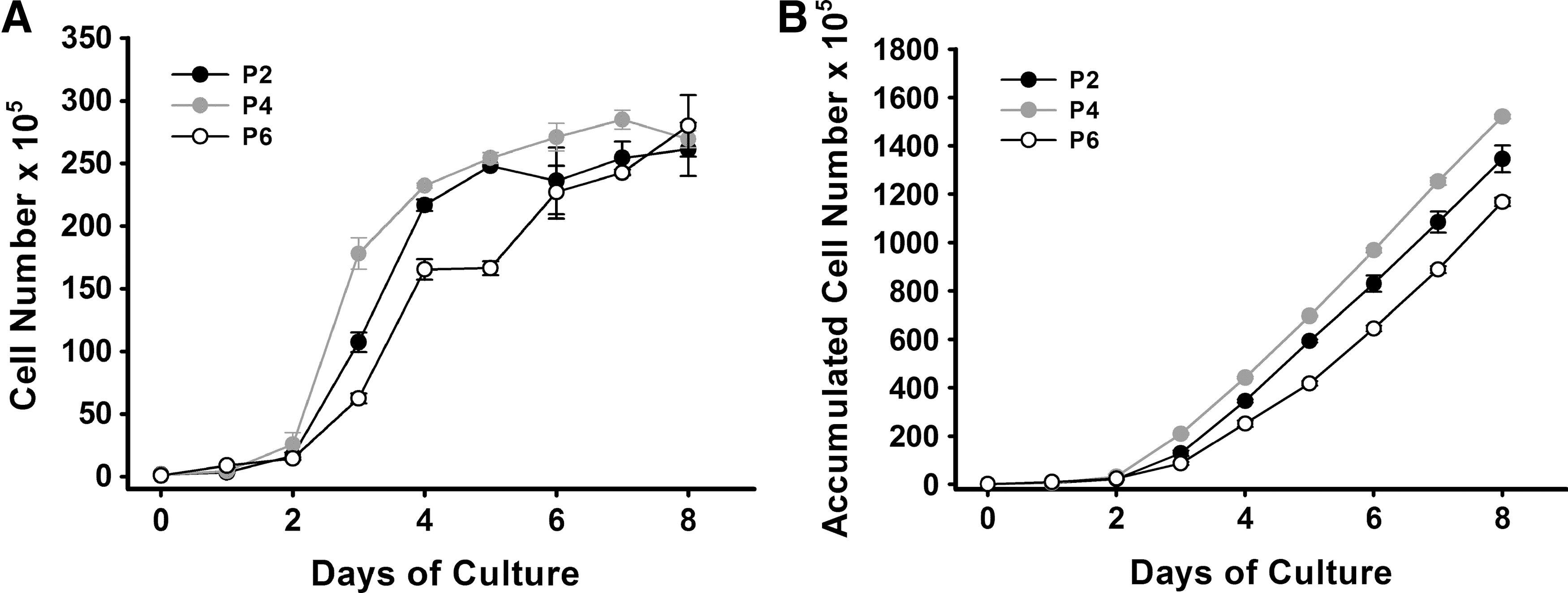
Growth curves of BM-MSCs.
Induced differentiation
Osteocytes and adipocytes
A feature of the MSCs is their ability to differentiate toward mesenchymal lineages, including adipogenic, chondrogenic, and osteogenic lineages [18]. Figure 3A-b shows that the cells were able to differentiate into osteocytes treated with osteogenic differentiation medium. Figure 3B-b shows that the cells were able to differentiate into adipocytes when treated with adipogenic differentiation medium. After 21 days of osteogenic induction, the majority of monolayer cultures had undergone differentiation toward osteocytes, as indicated by the formation of mineralized matrix that was demonstrated by the Alizarin red staining (Fig. 3A-b). Similar to osteogenic induction, after 21 days of induction, adipogenic differentiation was evidenced by presenting multivacuolar cells with the secretion of neutral lipid droplets that were confirmed by oil red O staining (Fig. 3B-b). Undifferentiated cells were not stained by either the Alizarin red or the oil red O, but still showed a stable fibroblast-like phenotype during the experimental period (Fig. 3A-a, B-a).
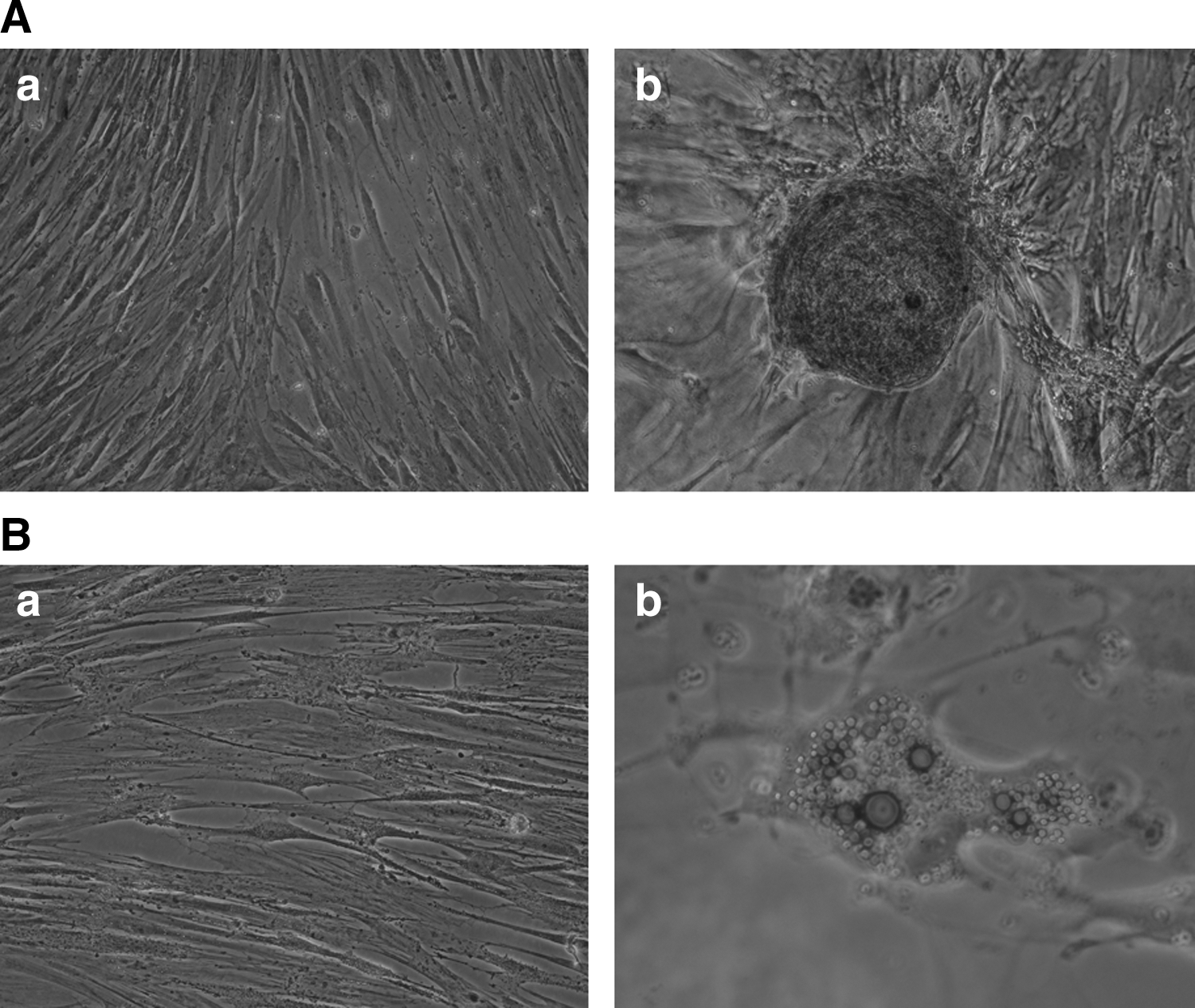
BM-MSCs of giant pandas can differentiate into osteogenic and adipogenic of the main mesenchymal lineage cell types.
Cardiomyocyte-like cells
After the subconfluent cells (Fig. 4A-a) had been exposed to 5- azacytidine for 24 h, some adherent cells died. However, surviving cells continued to proliferate and differentiate. At 1 week of the induction, the cells showed elongated and extended cytoplasmic processes with adjacent cells in which ∼20% of them aggregated and gradually increased in size to form a stick-like appearance (Fig. 4A-b). In the 3rd week, the cells became enlarged and showed a number of branches (Fig. 4A-c). At the fourth week, the cells formed a myotube-like structure by connecting with adjoining cells (Fig. 4A-d). However, spontaneous beating was not observed. Untreated cells maintained their fibroblast-like morphology.

The cardiomyogenic potential of the BM-MSCs of giant pandas.
To verify whether or not the differentiated cells were cardiomyocyte-like cells, the mRNA expressions of cardiomyogenic specific markers [22] such as GATA transcription factors 4 (GATA-4), cardiac troponin T (cTnT), and myosin heavy chain 7B (MYH7B) were assessed by using RT-PCR. Figure 4B shows that GATA-4, cTnT, and MYH7B were strongly expressed in the induced cells, although there were weak expressions for these genes in the undifferentiated cells. The differentiated cells had 1.5, 2.4, and 1.9 times higher expressions of GATA-4, cTnT, and MYH7B than the undifferentiated cells, respectively (Fig. 4C). These data suggest that the cells have cardiomyogenic potential.
Neuroglial-like cells
To assess whether or not the cells have any proclivity toward neural ectodermal lineage, experiments were performed to induce neural differentiation. After preinduction of 24 h, some treated cells changed morphology into neuron-like cells (Fig. 5A-a). With the time of exposure to SFM/BME, the cells started to show the change into neuroglial-like cells. The cytoplasm of treated cells retracted toward the nucleus, forming contracted multipolar cell bodies that assumed neuronal morphological characteristics (Fig. 5A-b–d). Untreated cells maintained their fibroblast-like morphology.
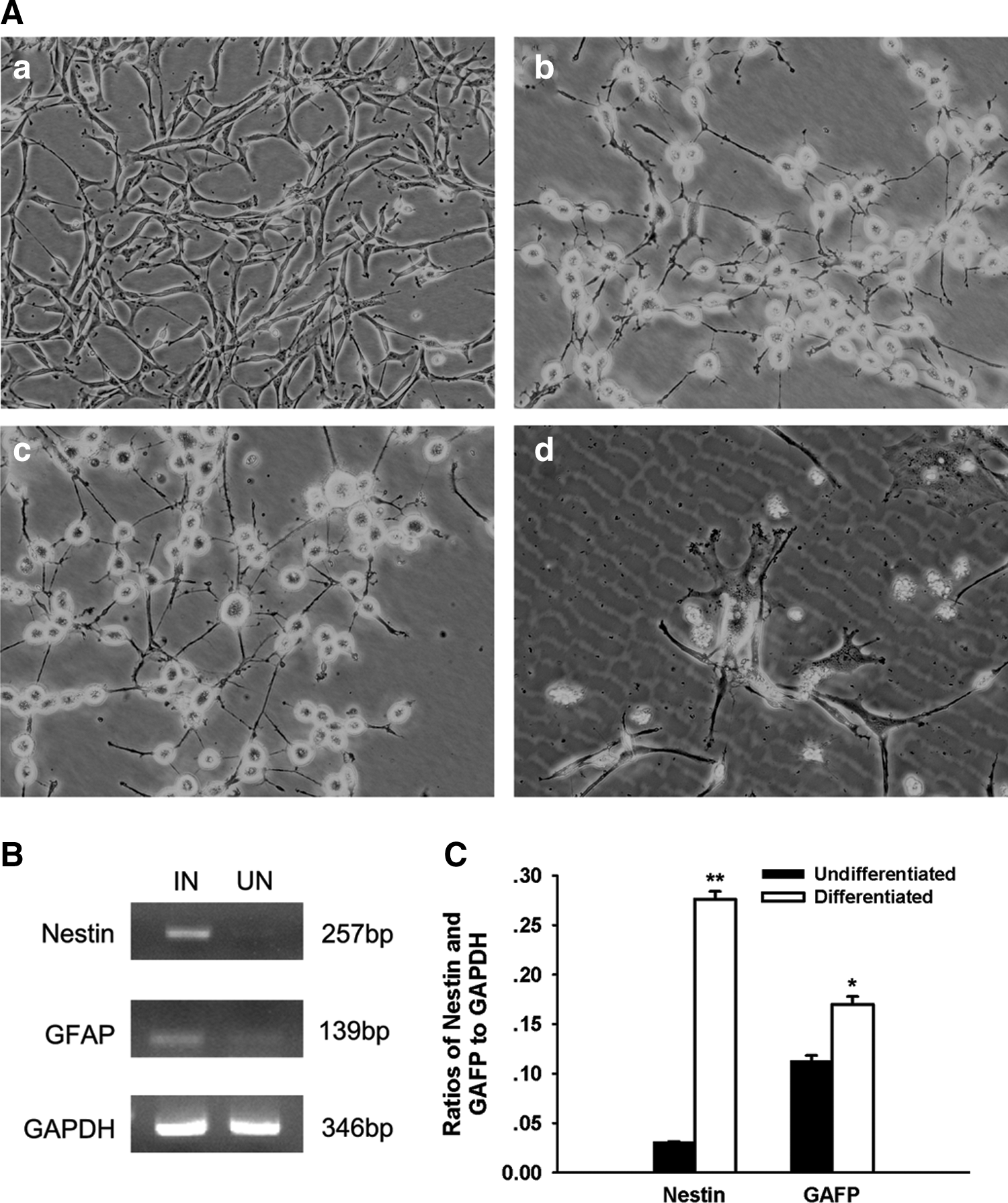
The neural potential of the BM-MSCs of giant pandas.
RT-PCR analysis was performed to demonstrate whether or not the neuron-like cells expressed neuro-glial specific markers [23] such as nestin (the neuronal precursor marker) or glial fibrillary acidic protein (GFAP, astrocyte marker). Figure 5B shows that undifferentiated cells expressed weak but detectable expressions of these genes. However, the expressions for nestin and GFAP were 8.94 and 1.5 times higher in the neuroglial-like cells than those in the undifferentiated cells, respectively (Fig. 5C).
Discussion
In the present study, we successfully isolated MSCs from BM of giant pandas. The cells showed fibroblast-like morphology and have a stem cells property to self-renew at a relatively higher rate in vitro. The cells are positive for MSCs markers such as CD73, SCFR, and GNL3, and pluripotent stem cells markers such as SOX-2. Furthermore, the cells have proclivity toward mesodermal origins such as osteoblasts, adipocytes, and cardiomyocytes in culture, as well as toward neural ectodermal lineages. The present study, to the best of our knowledge, is the first study to isolate, expand, and analyze BM-MSCs of giant pandas.
According to the definitions announced by the International Society for Cellular Therapy, the minimal criteria for MSCs are their capacity for plastic adherence during growth, specific surface antigen (Ag) expression, and multipotent differentiation potential [18]. Our results were able to meet these minimal criteria, demonstrating that the cells isolated from the BM of stillborn giant pandas were indeed MSCs.
MSCs have been isolated and characterized from various species, including humans [4], small experimental animals such as rats [17], mice [24], and rabbits [25], and large animals such as dogs [26], cats [27], pigs [28], monkeys [29], bovines [30], and horses [31]. Our results showed that MSCs from giant pandas displayed similar characteristics to the cells of humans and other animal species.
However, MSCs can be obtained from a variety of organs/tissues, including fetal liver, BM, blood and amniotic fluid, umbilical cord tissues and umbilical blood at birth, developing tooth at youth, and BM tissues and fat tissue during adult life [32]. It has been reported that MSCs from different sources of the same species showed different characteristics in terms of proliferation capacity, differentiation ability, and/or other biological features [33 –36]. Drawing on these evidences, it could be speculated that MSCs of giant panda may be different from those of other species in certain properties, although the essential properties and functions of stem cells are conserved through evolution [37]. However, no study has been published so far on the comparison of MSCs among species. Therefore, the hypothesis needs to be verified by comparative studies in the future.
Since no specific antibodies against giant panda are commercially available at the present, we developed the use of antibodies against humans or rats CD105, CD73, CD90, and CD45 to analyze the giant panda cells by flow cytometry. Unfortunately, no specific bindings were observed (data not show). Therefore, we only used morphologic observations and RT-PCR to analyze the giant panda cells. Moreover, since the genome mapping of giant pandas is not fully developed, we only chose some marker genes for our study that had sequences which were available in the giant panda genome database. Thus, the preparation of specific antibodies against giant pandas will be very important in future studies.
In the present study, we also found that markers for cardiomyocyte or neural-glial cells were weakly expressed but detectable in undifferentiated cells. The finding is similar to the results reported by Antonitsis et al. [22] and Tseng et al. [38]. Nevertheless, differentiated cells showed strong expressions of the markers, indicating that the cells have been directionally differentiated to the expected cell types. Furthermore, we found that the cells expressed pluripotent stem cells markers such as SOX2, which along with neural ectodermal lineage differentiation suggest that the cells might have more multipotency beyond the results we have studied so far.
Despite extensive research into the roles of MSCs, there are still limited reports on wild animals. To date, only one report by Fink et al. [39], in which adipose-derived MSCs were obtained from the brown bears, revealed the spontaneously forming extracellular matrix characteristic of bone and cartilage in the absence of specific inducers. Their finding could have implications for using hibernating brown bears as models to study disuse osteoporosis. Owing to self-renewal and multipotency, BM-MSCs from giant pandas could be potentially useful in rescuing the species from extinction because of the possibility of producing MSC-derived germ cells, as mature gametes have been derived in vitro from mice BM-MSC, ovarian surface epitheliums, and porcine fetal skins [8 –11]. This can be used in conjunction with assisted reproduction efforts to increase the size and diversity of the giant panda population in the future.
In conclusion, the BM-MSCs isolated and analyzed from giant pandas are capable of in vitro extensive multiplication and multilineage differentiation that are similar to other species, and can be regarded as a promising cell population for aiding in conservation efforts.
Footnotes
Acknowledgments
This work is supported by the Prophase Project of the National Key Basic Research Program funded by MOST (2012CB722207), the program of the Chengdu Science and Technology Bureau (12GGYB810SW-002), and the program of the Giant Panda Breeding Research Foundation (CPF2010-03).
Author Disclosure Statement
The authors declare no competing financial interest.