
Abstract
Progenitor expansion during development is a highly regulated process dictating the final organ size, while expansion of specific progenitor populators can adjust the final cellular composition of the organ. Understanding factors involved in these pathways is required to develop cell-based therapies such as β-cell transplantation for conditions such as diabetes mellitus. One versatile factor controlling both processes as well as a network of other proteins involved in pancreatic and duodenal development is the transcription factor SOX9. This review will focus on a comparison of SOX9 function during progenitor expansion and differentiation in the developing pancreas and duodenum with specific focus on endocrine development. During human pancreatic development, SOX9 functions in a dose-dependent manner to regulate epithelial progenitor expansion and endocrine differentiation. SOX9 expression is eventually limited to a subset of ductal and centroacinar cells, hypothesized to be the pancreatic stem cell compartment. Similarly, during duodenal development, SOX9 is expressed in most early epithelial progenitors and becomes gradually restricted to proliferative progenitors in the lower crypts, as well as mature Paneth and enteroendocrine cells indicating some differences in functional roles. However, in both developmental contexts, SOX9 is involved in pathways responsible for cellular proliferation and differentiation, such as Notch and Wnt. With its adaptable and central function in progenitor control, SOX9 represents an attractive target for manipulation for in vitro progenitor expansion and differentiation meriting further investigation.
Introduction
D
Numerous transcription factors, expressed differentially in the developing pancreas and duodenum, govern progenitor expansion and endocrine differentiation. Among this vast network is the Sry Box (SOX) factor, SOX9, which has been linked to progenitor proliferation and differentiation in both developmental systems [3 –7]. To date, literature reviews focusing on the roles of SOX9 transcription factor and signaling cascades involved in pancreatic and duodenum development are limited with very few publications highlighting its role in humans. This review will focus on the role of SOX9 in maintaining progenitor cell populations as well as its integral function of promoting proliferation and initiating differentiation in pancreatic and duodenal tissues via Notch and Wnt signaling pathways.
Structure of SOX9 and Roles During Development
The SOX9 protein belongs to the superfamily of high-mobility binding group (HMG) domain transcription factors, referring to the conserved 79 amino acid (AA) high-efficiency DNA binding domain. Distinctively, binding occurs in the minor groove of the double helix at specific consensus sequences, inducing a 70°–85° conformational change, which reveals previously hidden protein binding sites and allows close association with regulatory proteins (Fig. 1) [8 –12]. This unique DNA-binding mechanism allows SOX9 to assume a diverse and context-dependent role during development [13 –16].
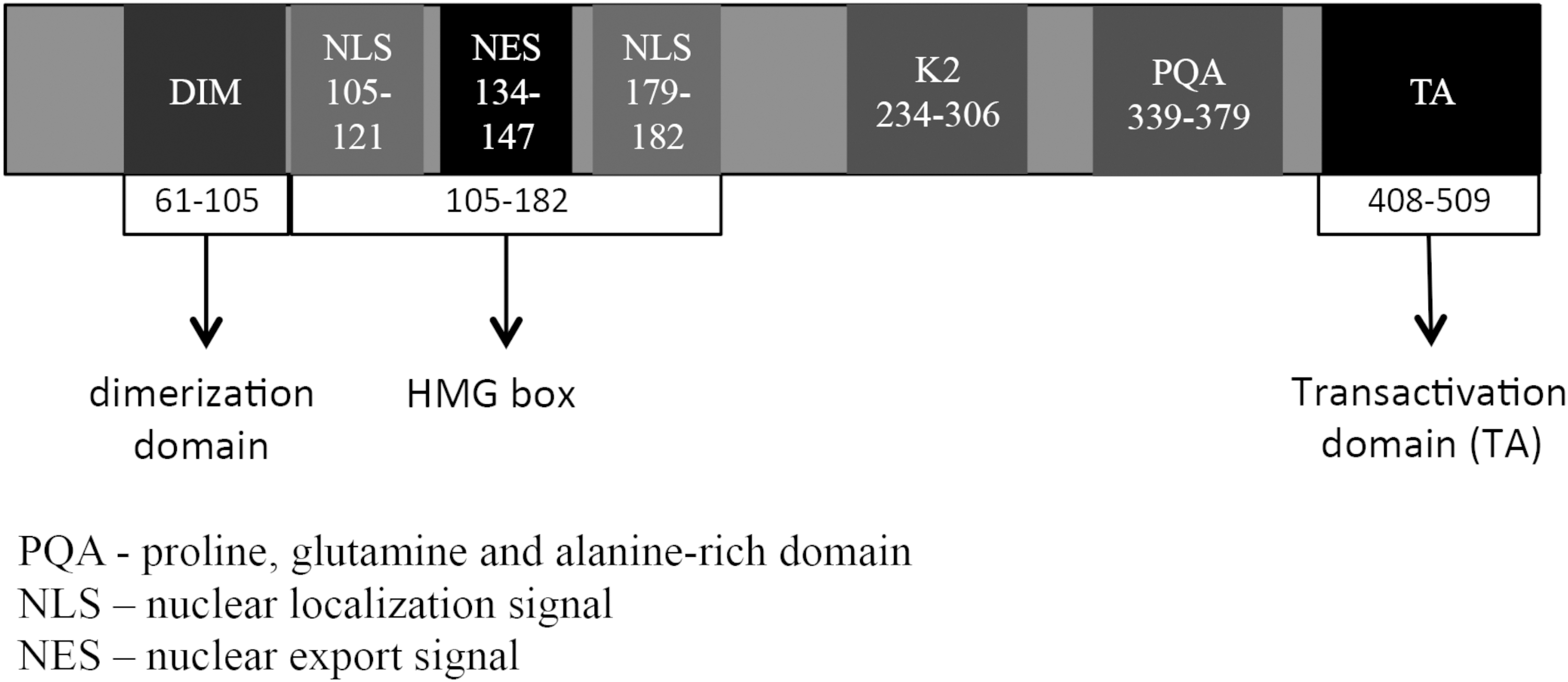
Diagram of Sox9 protein with functional domains labeled. The HMG box domain binds DNA at consensus sites transcribing relevant targets. The dimerization (DIM) and transactivation domains (TA) can bind other proteins modifying transcriptional targets, while the functions of the K2 and PQA domains are still unclear. [20,21,76 –80]
SOX9 was first described as the causative factor in Campomelic Dysplasia (CD; OMIM #114290) in which, haploinsufficiency manifests with 75% XY sex reversal, skeletal malformations, and pancreatic endocrine defects, thought to be due to reduced progenitor proliferation and differentiation [17 –21]. Since then, SOX9 has been found to greatly contribute to the organogenesis and development of many tissue types, including the gastrointestinal tract organs: the stomach, pancreas, and duodenum [5,14,20,22].
As demonstrated by symptoms of CD, SOX9 functions in a dose-dependent manner to regulate progenitor expansion and differentiation during chondrogenesis, sex determination, and pancreatic endocrine development [4,18,20]. In both chondrogenesis and sex determination, this has been attributed to decreased dimerization due to decreased SOX9 expression. Through interactions with numerous transcription factors, SOX9 ensures proper organogenesis, maintenance of stem cell compartments, and contributes to extracellular matrix modelling [14,23]. For each role during development, SOX9 interacts with a small subset of genes such as Col2a1 for chondrogenesis and organ fibrosis, anti-mullerian hormone in genital ridge development, and Ngn3 in pancreatic endocrine differentiation to carry out its specific functions for the development of organs and tissues [7,23,24].
SOX9 expression is controlled through several different processes. Cell autonomous expression of SOX9 is accomplished through long-range control of transcription. Numerous binding elements have been found up to 1 Mb upstream and 1.5 Mb downstream with global and tissue-specific enhancers [19]. One such enhancer, E1, is located 28 kb upstream of Sox9, and has been shown to regulate Sox9 expression in the notochord, bronchial epithelium, pancreas, and throughout the gut tube [25]. An additional major, highly conserved enhancer for Sox9 is located 70 kb upstream and promotes Sox9 expression in selective somatic tissues during development and regeneration, including the pancreas and gut [26]. Interestingly, this enhancer is responsive to Sox9 dimers, indicating a positive autoregulation of Sox9 and providing an additional level of control of expression [26]. This enhancer has been show to be necessary for both pancreatic development and regeneration following cearulin-induced acute pancreatitis [26]. In addition, Sox9 regulates and is regulated by several signaling pathways, including Wnt, providing regulatory control to and from the surrounding environment, which will be reviewed more extensively in the sections below [27 –30]. Finally, both protein partners and modifications of SOX9 (phosphorylation, acetylation, and sumoylation) provide posttranslational control through the disruption of stability, modification of interactions, and regulation of localization [13].
In mammals, both the pancreas and duodenum share a common endodermal origin and exhibit many parallels in transcriptional profile during development [31 –33]. Transcription factors such as Pdx1 have been found to be expressed in all islet and exocrine cell populations as well as the epithelial layer of the intestines during early development. Later in development, Pdx1 expression becomes restricted to β-cells, δ-cells, and duodenal tissues [34]. A common Sox9-expressing progenitor was recently identified in both pancreatic and intestinal epithelium of rodents and was shown to provide a continual supply of cells throughout development and during adulthood. However, these Sox9+ progenitors were found to contribute only to the pancreatic endocrine compartment during development, illustrating the complex and adaptable role of Sox9 [35]. This is also indicative of the importance of SOX9 in endodermal stem cell maintenance.
SOX9 in Pancreatic Development
A significant role of the SOX9 transcription factor has been confirmed in the developing and adult human pancreas [17]. Patients with SOX9 haploinsufficiency (CD) present a significantly disrupted islet structure (including loosely packed endocrine cells), a distinct reduction in numbers, an expression of late markers (IAPP and PC1/3), and a reduction in islet size with no changes in the exocrine compartment [17]. This suggests an essential role of SOX9 in pancreatic endocrine development.
During murine pancreatic development, Sox9 expression is first detected at E9 and colocalizes with Pdx1 at the future site of the ventral and dorsal pancreatic buds. Expression continues in the majority of Pdx1+ epithelial cells from e10.5 to e12.5 [36]. At E14, Sox9 is restricted to a subpopulation of Pdx1+ cells located within the central epithelial cords [3]. At this site, proliferating Notch-responsive progenitors give rise to migrating Ngn3+ endocrine precursors [37,38]. In agreement, the Notch target Hes1 was found in ∼40% of Sox9+ cells, indicating that they represent a supply of proliferating progenitors [3]. Additionally, Sox9 maintains the pancreatic progenitor identity through cell autonomous expression of Fgfr2b. Ablation of Sox9 from Pdx1+ cells at e10.5 during murine pancreatic development (Pdx1-Cre:Sox9 fl/fl) resulted in a loss of Fgfr2b expression with pancreatic hypoplasia and expression of liver markers, such as albumin [39]. Mesenchymal Fgf10 signaling acts to expand and maintain the identity of the pancreatic progenitor population in a feedforward loop through induction of Sox9 expression [39]. Later in development (>e18.5), Sox9 is restricted to a subset of ductal epithelial cells, including centroacinar cells [36]. Sox9 deletion (Sox9 −/−) results in developmental failure of the pancreas and death of the mice shortly after birth [3]. Sox9 −/− embryos' pancreas at e11.5 demonstrate an increased ratio of glucagon+/Pdx1+ cells, increased Isl1+ cells, reduced Hes1 expression with decreased proliferation, and increased apoptosis of pancreatic cells [3]. Mosaic deletion of Sox9 from Pdx1+ cells (Pdx1Cre:Sox9 fl/fl) resulted in decreased expression of Pdx1 by e12.5, suggesting that Sox9 is involved in Pdx1 expression to maintain pancreatic progenitors [40]. However, the early promoter used in the study by Seymour et al. (2007) [3] did not demonstrate the changes in Pdx1 expression, pointing out that Sox9 assumes different roles during early and late stages of pancreatic development. In addition, Sox9 plays a role in Ptf1α+ cells during murine embryonic development [41]. Cells that are Ptf1α+Sox9+Hnf1β+ make up a possible multipotent progenitor pool that is present in the pancreatic epithelium. Pft1α+Sox9+Hnf1β+ cells are able convert their expression to Ptf1αHISox9LO, which favours an acinar fate. However, the cells that expressed high levels of Sox9 and Hnf1β in the pancreatic trunk are reported to assume an endocrine or ductal cell fate [41]. Furthermore, Ptf1α+ acinar cells could be activated during an injury-induced factultative restoration to become Ptf1α+Sox9+Hnf1β+ progenitors, which undergo rapid reprogramming from acinar-to-duct-to-endocrine transdifferentiation [41]. Taken together, these studies indicate that Sox9 plays a crucial role in early pancreatic development and islet cell differentiation in the mouse (Fig. 2).
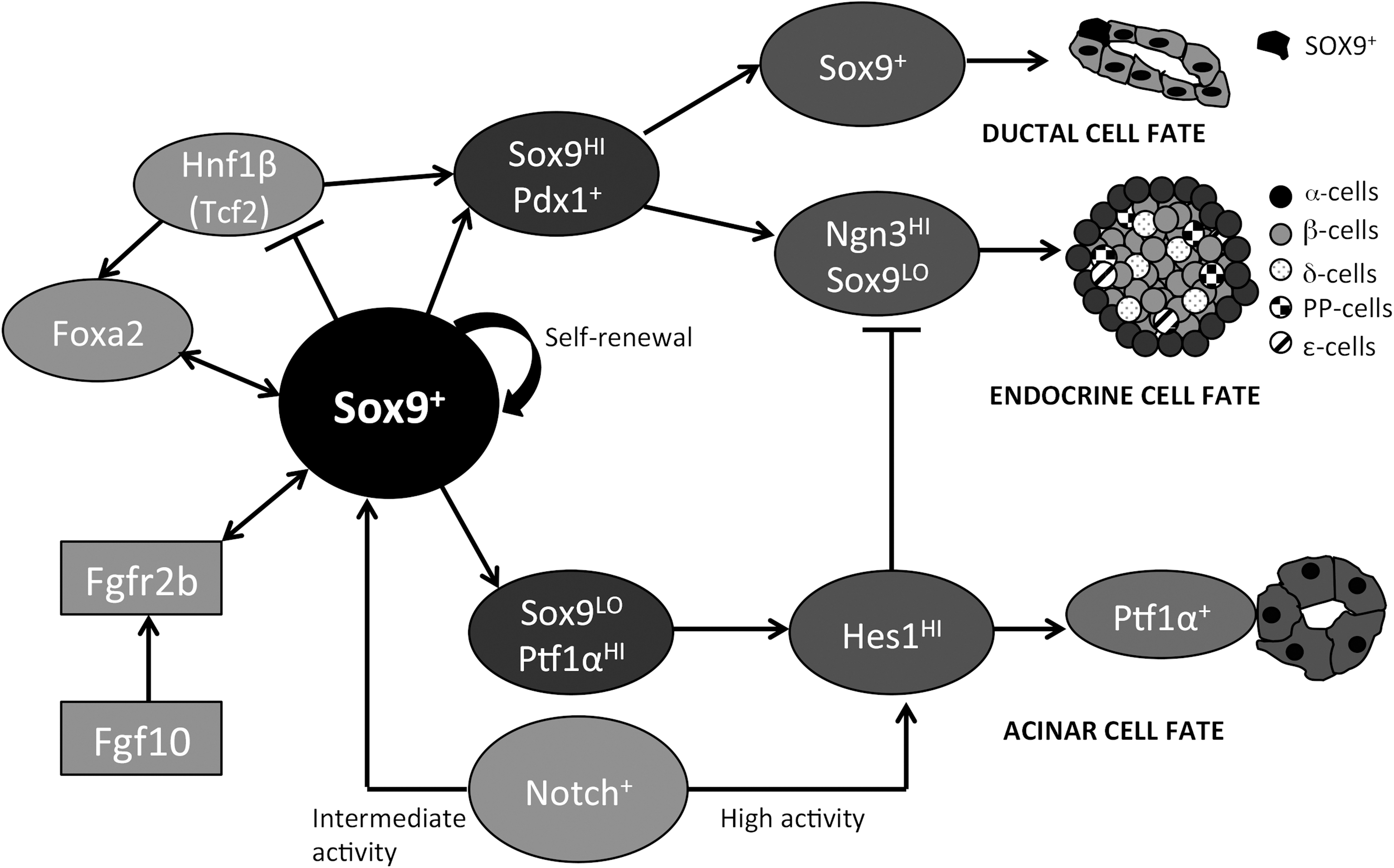
Simplified lineage schematic of Sox9's role in the developing murine pancreas. Sox9+ cells have the ability to self renew and give rise to different cell types during the murine pancreas development. Sox9 directly binds to and regulates Foxa2 and Hnf1β, forming a stable transcription factor network through either direct or indirect feedback interactions to stabilize progenitor cell identity. The Fgfr2b/Fgf10 interactions affect the expression of Sox9 cells in the developing pancreas. Intermediate Notch activity maintains Sox9 expression and differentiation into endocrine lineage, while high Notch activity leads to increased Hes-1 and subsequent exocrine lineage differentiation. [3,7,39,41,55]
During human pancreatic development, SOX9 expression is detected from 8 to 21 weeks of fetal age. Protein levels of SOX9 significantly decreased at 19–20 weeks compared to 10–11 weeks. Colocalization analysis of SOX9 and various pancreatic transcription factors indicated a consistent pattern. A high degree of colocalization was observed early in development; however, reduced coexpression patterns were detected in later developmental periods. Specifically, PDX-1 was found to be highly expressed in SOX9+ cells at 8–11 weeks, but a significant reduction was observed at 14–16 weeks [42,43]. A similar pattern of expression was observed with CK19, a marker of ductal cells. Interestingly, SOX9+ cells were also found to express numerous endocrine progenitor markers, including NGN3, FOXA2, NKX2.2, NKX6.1, and PAX6, but SOX9 expression was absent from mature endocrine cells, a finding that was also noted in studies of murine pancreatic development [3,7,42]. Finally, proliferation, as indicated by KI67 labelling, was also found to be high in the SOX9+ population when compared to other pancreatic cell types, providing further evidence that SOX9 is expressed in proliferating progenitors [42].
SOX9 in Islet Development
Similar to the phenotype of patients with CD, reducing Sox9 expression through monoallelic deletion in mice (Sox9 ±) produced a normal exocrine compartment with islets displaying hypoplasia (50% reduction in mass) along with a reduction in late differentiation markers [4].
Further analysis of islets revealed that all cell types and the total pancreatic insulin content were reduced by 50% [4]. The reduction in endocrine cell numbers was due to a diminished progenitor population, as shown by a 50% reduction in Ngn3+ cells along with low levels of Sox9 expression [4]. However, the existing β-cells expressed markers of functional insulin-producing cells (Glut2, PC1/3, IAPP), indicating that Sox9 expression is not required for proper functional β-cell development, but is more important for the maintenance of progenitor cell populations. Furthermore, in vitro studies reveal that Sox9-dependent expression of Ngn3 is required to induce endocrine cell development [4,7]. NGN3 is essential for the development of all endocrine cell types [44]. Furthermore, the direct control of Ngn3 expression is achieved through direct binding of Sox9 to the murine Ngn3 promoter [4,7]. In murine studies, Sox9 has also been shown to regulate the expression of factors upstream of Ngn3. The knockdown of Sox9 (50%) resulted in a proportional reduction of FoxA2 and Onecut1 expression, along with a 50% increase in Hnf1β (TCF2) expression [7]. This indicates that Sox9 plays a role in controlling the transcription factor regulatory network involved in islet cell progenitor proliferation and maintenance (Fig. 2). Interestingly, Dubois et al. demonstrated the results that conflict with the findings of previously reported studies. They reported that biallelic deletion in Pdx1+ cells (Pdx1Cre:Sox9 −/−) resulted in a dramatic decrease in Pdx1 expression with no effect on FoxA2, Onecut1, and Hnf1β expression [40]. A similar result was also observed in a tamoxifen-induced Sox9 deletion (CreTM;Sox9fl/fl ) mouse model [40]. Taken together, these studies signify that Sox9 plays a critical and time-dependent role during pancreatic development [40].
Investigations of human fetal islet cells in vitro have produced similar results to murine studies, with some subtle differences. In human fetal islet epithelial cells, the knockdown of SOX9 revealed a 50% decrease in NGN3+ and KI67+ cells with a significant loss of insulin+ cells accompanied by a twofold increase in glucagon+ cells [42]. However, the knockdown of SOX9 did not result in any change in expression of PDX1 or FOXA2 when compared with controls, whereas mRNA expression of progenitor factors HNF1α (TCF1) and HES1 increased twofold. Interestingly, overexpression of SOX9 in human fetal islet epithelial cells showed inverse changes in expression of these factors [42]. Opposite results were seen in murine studies, where the percentage of Hes1+ cells was reduced in e10.5 embryos with Sox9-specific deletions in pancreatic cells [3]. This difference could be due to the methods used to supress the SOX9 activity (knockdown of islets versus pancreatic-specific deletion). These findings indicate that SOX9 participates in a vast network of progenitor factors, including Pdx1 in murine pancreatic cells and HNF1α and HES1 in human pancreatic cells. In both species, during pancreatic development, SOX9 functions to maintain proliferative progenitors and initiate endocrine differentiation through NGN3 in a dose-dependent manner. Additionally, increased expression of HES1 observed in human cells indicates that SOX9 plays a role in maintaining progenitor cells by antagonizing differentiation. Although the genetic program in which SOX9 controls pancreatic development is fairly understood, the pathways regulated by SOX9 are still unclear and will be discussed in the following sections.
SOX9 in Duodenal Development
Formation of the duodenum originates with the primitive midgut endoderm in close proximity to the region of pancreatic specification. The entire gut tube in early development exists as a homogenous cell population, which differentiates into appropriate structures based on surrounding cues [45]. During intestinal development, Sox9 expression can be detected in most duodenal epithelial cells at e13.5 (7 weeks of fetal age in humans), gradually restricted to proliferative progenitor, Paneth, and enteroendocrine cells by e18.5, and by adulthood is mostly located at the base of mucosal folds called the crypts of Lieberkühn [23,27,35,46]. Progenitors residing at the base of these crypts migrate toward the top of the intestinal villi providing a continual cell supply. These progenitors differentiate into various intestinal cell types: absorptive enterocytes, mucus-producing goblet cells, antimicrobial-producing Paneth cells, and hormone-producing enteroendocrine cells (Fig. 3) [27,46]. Sox9 maintains undifferentiated cell populations in the intestinal crypt through direct repression of Cdx2 and Muc2, which are required for general epithelial differentiation and specific goblet cell maturation [27]. Although Sox9 is associated with proliferating epithelial progenitors, inactivation of its expression in the murine intestinal epithelium (using the villin promoter, VilCre; Sox9 −/−) resulted in altered morphology and increased proliferation in the crypts [22]. Moreover, Paneth cells were absent with no change observed in goblet, enteroendocrine, or enterocyte cell populations [22]. To clarify expression of SOX9 during intestinal development, lineage tracing of Sox9+ cells using EGFP labeling (Sox9 EGFP) was performed. Expression was seen mostly at the base of the crypts, as well as in scattered cells along the villus [6]. Although no expression was detected in Paneth cells using EGFP, immunolabeling of the Sox9 protein found expression that could be due to an alternative regulatory element controlling Sox9 expression in this cell type.

Simplified diagram of morphology of duodenal crypt development. Early Sox9+ progenitors originate at the base of the crypts of Lieberkuhn. As cells migrate toward the intestinal villi, Sox9 expression is lost resulting in loss of repression of Cdx2 and Muc2 gene precipitating differentiation. Cells at the crypt base include progenitors, and mature Paneth cells retain Sox9 expression.
Another study using a different promoter construct with eGFP labeling (Sox9 IRES-eGFP) revealed expression in both Paneth and enteroendocrine cells, further indicating that the Sox9 promoter provides a cell-specific expression pattern [35]. Detailed analysis of expression levels revealed two distinct populations of Sox9HI and Sox9LO cell types in the crypts. Further characterization of these cell types revealed that Sox9HI cells have a fivefold increase in Sox9 mRNA expression when compared to Sox9LO cells. They also express substance P and chromogranin A, indicating that Sox9HI cells express a mature enteroendocrine phenotype. Moreover, the expression of stem cell markers, Notch1, Hes1, Lgr5, and Musashi1, were all found to be increased in Sox9LO cells, suggesting that these cells are more likely to represent a proliferative progenitor population [6]. Overexpression studies of Sox9 in duodenal epithelial cells resulted in decreased proliferation and morphological changes supporting findings of increased Sox9 expression in nonproliferative cells. This indicates that Sox9 plays a role in the transition from proliferating progenitor to mature cell, although markers of mature cells were absent suggesting that other factors are required for complete differentiation. Interestingly, no intestinal-specific effects have been observed in the CD patients, revealing that SOX9 may assume a different role during development when compared to the maintenance of adult stem cell populations. Alternatively, either monoallelic SOX9 expression is sufficient for intestinal development or other SOXE factors compensate for the partial loss of SOX9 [47].
Notch Signaling in SOX9+ Progenitors
In both intestinal and pancreatic Sox9+ progenitors, Notch signaling via Hes1 has an inhibitory role on proliferation and differentiation [37,48,49]. Hes1 expression maintains cells in an undifferentiated state, specifically by antagonizing Ngn3 and Math1 expression, which are required for the differentiation of endocrine cells in the pancreas and secretory cell lineages in the intestine (ie, cells other than enterocytes) [38,50 –52]. Notch activation is accomplished through direct contact of cell receptors initiating nuclear translocation of the Notch intracellular domain (NICD) and associating with cofactors (RBJκ in mammals) to regulate Notch targets [53,54]. Inhibition of Notch signaling in both the pancreas and duodenum allows progenitor cell populations to differentiate into mature phenotypes [53]. Recent studies have shown the Notch-dependent pathway in the developing pancreas regulating endocrine and ductal differentiation. With high Notch activity, expression of Sox9 is promoted, but endocrine differentiation is repressed due to Hes1 expression. As Notch activity decreases to an intermediate level, expression of Hes1 is lost, promoting endocrine differentiation, but Sox9 expression is maintained (Fig. 2) [55]. Loss of Notch activity through cell migration results in a loss of Sox9 expression and activation of endocrine differentiation via Ngn3 expression. Conversely, cells in contact with ducts retain Notch-induced Sox9 expression and differentiate into ductal cells [55].
SOX9/Wnt Pathways in Pancreatic and Duodenal Development
Stem cell niches have been shown to exist in multiple tissues and are regulated by a delicate balance of signaling pathways induced by the surrounding environment. In particular, Wnt signaling has been shown to play a role in promoting self-renewal and proliferation of multiple stem cell types [56 –61]. Interestingly, Sox9 also regulates the Wnt pathway in various stem cell environments [27,29,30,62]. Wnt ligands are lipid-modified glycoproteins (∼20 isoforms) responsible for short-range paracrine and autocrine signaling. These ligands bind the frizzled receptor (FZD) (10 isoforms in humans) and LRP5/6 coreceptors for canonical signaling and RYK/ROR receptors for noncanonical signaling [58,63,64]. The canonical Wnt signaling pathway occurs when the FZD receptor interacts with the Wnt ligand, which in turn inhibits GSK3β and prevents the phosphorylation and degradation of β-catenin. β-catenin is able to translocate to the nucleus, where it associates with cofactors, TCF7, 7L1, 7L2, and LEF1, to regulate Wnt target genes [64,65]. Noncanonical signaling occurs when FZD interacts with RYK and ROR coreceptors and functions through the JNK or Ca2+/NFAT signaling [58,63]. Receptor–ligand interactions in this pathway provide a wide degree of variability in cellular responses based on the identity of ligand and receptors present as well as competition between ligands [63,66 –68]. Furthermore, Sox9/Wnt interactions have been well studied in chondrocytes demonstrating that Sox9 exerts a negative feedback on the Wnt pathway through direct binding with both β-catenin and GSK3β, preventing the transcriptional activity and increasing β-catenin degradation [28 –30,69].
In the intestinal epithelium, the Wnt signaling pathway was first identified as a key regulator of proliferation through investigation into the hereditary colon cancer syndrome, familial adenomatous polyposis. The clinical condition manifests as extensive intestinal polyp formation, due to constitutive activation of the Wnt pathway via loss of the repressor protein adenomatous polyposi coli [70 –73]. High levels of Sox9 expression have also been detected in human colorectal carcinoma cell lines with known Wnt activating mutations, indicating possible Wnt-induced expression and roles in intestinal proliferation [27]. Moreover, murine Tcf4 deletion (Tcf4 −/−) results in undetectable Sox9 expression, providing evidence for expression through Wnt pathway, as well as reduced proliferation and an absence of both Paneth and enteroendocrine cells [27,72,74]. Taken together, these studies suggest that the increased proliferation observed may be due to unregulated Wnt expression and function and that SOX9 also contributes to the increased rates of proliferation observed.
Interestingly, examination of the Wnt pathway in Sox9-deficient animals (Sox9−/− ) did not show a significant change in nuclear β-catenin. However, overexpression of SOX9 in colon carcinoma cell lines resulted in decreased β-catenin/TCF reporter activity. Using transfection of full length or C-terminal truncated SOX9 (ΔCSOX9), differential effects on Wnt signaling were also observed. The Wnt target genes, c-Myc, and cyclinD1 protein levels were found to be downregulated upon transfection with full-length SOX9 and upregulated with ΔCSOX9 [5]. Mutation of the DNA binding domain abolished these results, indicating that this domain is required for Wnt modulation [6].
Paneth cells located at the crypt base, in close association with progenitors, express a high level of SOX9 and are required for intestinal stem cell maintenance (Fig. 3). They secrete a variety of factors, including the Notch ligand Dll4 and Wnt ligand (Wnt3), pointing to the essential role these Sox9+ cells have in stem cell regulation [5,22,75]. During pancreatic development, various FZD receptors and Wnt ligands are expressed, but precise spatiotemporal expression patterns are unclear due to limited antibody availability. To help clarify expression patterns of the Wnt pathway during human pancreatic development, our research group has performed microarray analysis of pancreata at 8–21 weeks of fetal age. Our microarray data demonstrate significant decreases in the expression of FZD2, FZD7, and WNT3 at 20–21 weeks when compared to 9–10 weeks (Table 1). Interestingly, approximately 50% of SOX9+ cells express WNT3A at 8–12 weeks. This decreases significantly by 14–16 and 19–21 weeks of fetal age, as determined by immunostaining. Half of the SOX9+ cells also stably express nuclear β-catenin from 14 to 21 weeks. The CK19+/SOX9+ cell populations are a proposed site of early endocrine progenitors. In this population, the expression of WNT3A expression was observed in 50% of cells at 8–12 weeks, but was found to be decreased by19–21 weeks. Furthermore, knockdown of SOX9 via siRNA in vitro in isolated human islet-epithelial clusters reduced SOX9 protein levels by 50%, which resulted in a 50% decrease in pGSK3β, an inactive β-catenin degradation enzyme, and subsequently reduced Wnt-associated target genes determined by decreased cyclinD1 expression and reduced nuclear β-catenin [42]. In line with these results, overexpression of SOX9 increased SOX9 protein expression 1.5-fold, producing a twofold increase in pGSK3β and a threefold increase in cyclinD1 expression. As expected, the proportion of KI67+ cells also increased twofold. These results show involvement of the Wnt pathway in SOX9+ progenitors during human pancreatic development. These studies suggest that SOX9 positively regulates Wnt through GSK3β, whereby SOX9 expression increases the Wnt pathway activity by directly or indirectly inhibiting GSK3β, reducing β-catenin degradation.
Human fetal pancreata (9–21 weeks of fetal age) were collected according to protocols approved by the Health Sciences Research Ethics Board at the University of Western Ontario, in accordance with the Canadian Council on Health Sciences Research Involving Human Subjects guidelines. Fetal samples were grouped into 9–10 weeks and 20–21 weeks, with at least three replicates per group. Total RNA from human fetal pancreata was isolated using TRIZOL reagent (Invitrogen, Burlington, ON). Biotinylated cRNA was generated and hybridized to Affymetrix HGU133-plus2 GeneChips. Data are expressed as mean fold changes±SEM and normalized using the Robust Multichip Analysis to show expression relative to 9–10 weeks data. Significantly (P<0.001) upregulated genes are highlighted in bold.
Summary and Future Research Directions
SOX9 is expressed in the duodenum and pancreas during both murine and human development. In these tissues, it serves a common function, in a dose-dependent manner, to regulate proliferation and differentiation of progenitors. To accomplish this function, SOX9 regulates the expression of numerous transcription factors along with the Wnt signaling pathway in both contexts. High levels of SOX9 are indicative of early endocrine differentiation demonstrated in murine pancreatic development [4] and shown in mature enteroendocrine cells in both mice and human duodenum. These differences can be explained, in part, by the activity of SOX9 on the Wnt pathway in both tissue types. In pancreatic epithelial cells, SOX9 increases the Wnt signaling activity, but represses it during duodenal development. Truncation of the C-terminal transactivation domain changes this effect to stimulatory, pointing to a partner protein modulating SOX9 in the duodenum. Despite this difference, loss of SOX9 in both tissues leads to a reduced number of progenitors. This makes SOX9 an ideal candidate for manipulation in vitro to expand progenitor populations. Further work should focus on characterizing proteins that bind SOX9 and their pattern of coexpression during development (Fig. 1). This will provide insight into the particular transcriptional profile of SOX9 in each cell type and how pathways are affected by differences in partners expressed. Additionally, differences in how SOX9 affects signaling pathways could explain the variation in function during duodenal and pancreatic development. An increased understanding of progenitor expansion and differentiation controlled by SOX9 could help improve current β-cell differentiation protocols.
Footnotes
Acknowledgment
This work was supported by grants from the Canadian Diabetes Association.
Author Disclosure Statement
No competing financial interests exist.