
Abstract
The therapeutic potential of mesenchymal stem cells (MSC) has highlighted the need for identifying easily accessible and reliable sources of these cells. An alternative source for obtaining large populations of MSC is through the controlled differentiation of induced pluripotent stem cells (iPSC). In the present study, colonies of iPSC were cultured in MSC culture media for 2 weeks. Serial passaging then selected for fast growing MSC-like cells with a typical fibroblastic morphology and the capacity to proliferate on standard culture flasks without feeder cells. MSC-like cells were developed from iPSC lines arising from three different somatic tissues: gingiva, periodontal ligament (PDL), and lung. The iPSC-MSC like cells expressed key MSC-associated markers (CD73, CD90, CD105, CD146, and CD166) and lacked expression of pluripotent markers (TRA160, TRA181, and alkaline phosphatase) and hematopoietic markers (CD14, CD34, and CD45). In vitro iPSC-MSC-like cells displayed the capacity to differentiate into osteoblasts, adipocytes, and chondrocytes. In vivo subcutaneous implantation of the iPSC-MSC-like cells into NOD/SCID mice demonstrated that only the PDL-derived iPSC-MSC-like cells exhibited the capacity to form mature mineralized structures which were histologically similar to mature bone. These findings demonstrate that controlled induction of iPSC into fibroblastic-like cells that phenotypically and functionally resemble adult MSC is an attractive approach to obtain a readily available source of progenitor cells for orthopedic and dental-related tissue-engineering applications. However, a detailed characterization of the iPSC-MSC-like cells will be important, as MSC-like cells derived from different iPSC lines exhibit variability in their differentiation capacity.
Introduction
M
Due to the scarcity of specific cell surface markers available to positively identify and isolate MSC, the International Society of Cellular Therapy has formulated minimal criteria for defining multipotent MSC [19,20]. In order for cells to be classified as MSC, they have to meet three key criteria: First, they should be plastic adherent when maintained in standard culture conditions. Second, ≥95% of the MSC should express CD105, CD73, and CD90, as determined by flow cytometry; these cells should also lack (≤2% positive) expression of CD45, CD34, and CD14. Finally, the MSC should be able to differentiate to osteoblasts, adipocytes, and chondrocytes under standard in vitro differentiation protocols [19,20]. However, many of these criteria also describe other fibroblast populations with limited differentiation capacity and fail to address the stem cell property of self-renewal.
With an increasing interest in MSC, for use in clinical stem cell treatment strategies, the identification of an accessible and reliable source of these cells is a critical problem. Current methods for ascertaining MSC from patients are invasive, expensive, and labour intensive. Furthermore, MSC have limited capacity to expand in culture, as they generally undergo culture senescence after 8–10 passages, thus inhibiting the generation of the large cell numbers required for clinical therapies. It is, therefore, important that alternative sources of MSC are established.
The discovery that induced pluripotent stem cells (iPSC) can be generated from adult somatic cells using reprogramming techniques [21,22] represents a promising alternative for obtaining larger populations of adult stem cells for use in cell therapies and regenerative medicine. The iPSC are similar to embryonic stem cells (ESC) but since they are derived from adult cells, they do not attract the same ethical concerns as do ESC. In addition, iPSC are not limited in terms of their expansion in culture, meaning they could function as an inexhaustible source of MSC. Furthermore, iPSC-MSC have been found to have a greater proliferation capacity than bone marrow derived MSC (BM-MSC) [23]. For example, MSC from human iPSC have been reported to be capable of expanding for approximately 40 passages (120 population doublings) without obvious loss of plasticity or onset of replicative senescence [23]. The combination of the high proliferative capacity of iPSC and iPSC-MSC-like cells should facilitate the generation of large quantities of MSC for clinical treatment approaches.
In recent years, various groups have successfully derived MSC from iPSC using a range of methods [23 –30]. Two main strategies were previously employed to generate MSC from iPSC. One strategy consists of dissociating iPS colonies, which are platted out onto gelatine-coated flasks in the presence of growth factor media, into a single cell suspension. The resulting cells are enriched either for MSC-like cells through cell passaging protocols or through flow cytometric selection [23,28,29]. The alternative strategy involves the formation of embryoid bodies (EBs), where the iPSC are grown in suspension to promote differentiation into the three primary germ layers. The EBs are then cultured on gelatine-coated flasks, and various methods are employed to enrich for MSC-like cells [26,27]. Recently, fibrillar collagen or a small-molecule inhibitor has also been used by different groups to aid the differentiation of iPSC to MSC [25,30].
The aim of this study was to develop a simplified and reproducible method for inducing iPSC into MSC-like cells. We have achieved this through modification of a protocol previously used to generate MSC-like cells from ESCs [31]. In this study, we utilized a serial passaging protocol to establish MSC-like cells from iPSC generated from three different somatic tissue derived cell lines: gingival fibroblasts (GF). PDL, and lung fibroblasts. We demonstrate that the cell lines generated through this induction protocol met the minimal criterion for classification as MSC [19,20]. However, iPSC-MSC lines generated from PDL showed a superior differentiation capacity both in vitro and in vivo.
Materials and Methods
Cell culture
The GF-iPSC and PDL-iPSC were sourced from in-house supplies [32]; the iPSC were reprogrammed from human female gingival fibroblasts and PDL cells, respectively, through retro-viral transduction of OCT3/4, SOX2, KLF4, and c-MYC. The human lung iPSC line used was obtained from The National Stem Cell foundation of Australia (previously known as The Australian Stem Cell Center).
iPSC were cultured in iPSC cell medium containing the base media Dulbecco's modified Eagle's medium-Nutrient mixture F-12 (DMEM/F12) (Life Technologies Australia) that was supplemented with 20% Knockout Serum Replacement (Knockout™ SR; Life Technologies Australia), 2 mM
Induction of iPSC to MSC-like cells
Confluent flasks of non-induced iPSC were dissociated with 4 mg/mL of collagenase I; to remove mouse embryonic fibroblasts (MEFs) feeders and any differentiated cells, the resulting clusters of iPSC were partially dissociated via manual pipetting and then platted out onto 0.1% gelatine-coated tissue culture flasks (which did not contain MEFs). The passaged iPSC were cultured in MSC culture media (instead of the iPSC media that they had been growing in) for 2 weeks with occasional media changes, thus enabling the iPSC colonies to differentiate and produce outgrowths of heterogenous cell types. MSC culture media comprised Minimum Essential Medium Eagle - alpha modified (α-MEM) with full additives consisting of 10% fetal calf serum (FCS), penicillin/streptomycin, sodium pyruvate,
Differentiation assays were performed as detailed next to assess whether the MSC-like cells that were generated had multipotent differentiation potential.
Flow cytometric analysis—MSC panel+iPS markers
Single cell suspensions of the iPSC-MSC were prepared by digestion using trypsin/iPSC-MSC-like cells. A single cell suspension of iPSC was obtained through digestion with Cell Dissociation Buffer Enzyme-Free phosphate-buffered saline (Gibco), as we found this dissociation buffer lifted of the iPSC while leaving the MEFs attached to the flasks. The single cell suspensions were resuspended in blocking buffer containing Hank's balanced salt solution (Sigma-Aldrich) that was supplemented with 5% normal human serum (Australian Red Cross), 1% bovine serum albumin (BSA; ICN Biomedicals), and 5% FCS (Thermo Electron) and incubated on ice for 30 min. Approximately 1×105 cells were incubated with specific cell surface marker antibodies (10 μg/mL) or isotype control antibodies (10 μg/mL) on ice for 1 h. Antibodies specific for CD29, CD73, CD90, CD105, CD166 (BD Biosciences), CD44 (Clone H9H11; kindly provided by Prof. Andrew Zannettino, Division of Haematology, IMVS), CD106 (Clone QE4G9; kindly provided by Dr. Ravi Krishnan, Transplantation Immunology Laboratoy, The Queen Elizabeth Hospital), CD146 (CA12) [33] and alkaline phosphatise (STRO-3) [34], TRA160 (Millipore), TRA181(Millipore), CD14-PE (Beckman Coulter), CD34-PE (Beckman Coulter), CD45-PE (Beckman Coulter), or isotype control immunoglobulin, 1B5 (IgG1), 1D4.5 (IgG2), and 1A6.12 (IgM) (kindly provided by Prof. L.K. Ashman, The University of Newcastlea) were used. After washing, cells were incubated for 30 min on ice with their respective secondary detection antibodies: goat anti-mouse IgG-PE or IgM-PE (Southern Biotechnology Associates). After washing, the samples were fixed and then analyzed using an Epics-XL/MCL flow cytometer (Beckman Coulter). Analysis was performed using the FlowExpress software.
In vitro differentiation assays
Osteogenesis
Mineralization was induced as previously described [35]. Briefly, iPSC differentiated to MSC iPSC-MSC-like cells were seeded in triplicate in 24-well plates with 3×104 cells/well for use in imaging; cells were also seeded in six-well plates with 2×105 cells/well for RNA collection. Cells were cultured in osteoinductive media, which consisted of α-MEM supplemented with 5% FCS, 100 μM
After 28 days of induction, mineral deposit formation was identified in the 24-well plates with the use of Alizarin Red staining (Alizarin Red S; Sigma-Aldrich, Inc.); images were taken using standard light microscopy.
RNA was obtained from the cells grown in the six-well plates by Trizol isolation. RNA was isolated from cells at day zero, which was before osteogenic induction, as well as from cells that were grown in osteoductive conditions for 4 weeks and cells grown in the absence of osteoconductive conditions for 4 weeks.
Adipogenesis
Adipogenesis was performed as previously described [1,35]. For adipogenic induction, iPSC-MSC-like cells were seeded in 24-well plates at 3×103 cells per well and cultured for 4 weeks in adipogenic media, which was changed thrice a week [1,35]. Lipid deposits were observed after staining with Oil Red O (MP Biomedicals). iPSC-MSC-like cells were also plated in six-well plates as described earlier, and RNA was isolated with TRIzol after 28 days of adipogenic induction.
Chondrogenesis
Chondrogenesis induction was performed as previously reported [36]. Cell pellets from 5×105 iPSC-MSC-like cells were spun at 600g and cultured in polypropylene tubes in chondrogenic media that was changed thrice a week for 28 days. Pellet cultures were set up in duplicate with one pellet being used for histological assessment and the second pellet being used for reverse transcriptase (RT)-polymerase chain reaction (RT-PCR) analysis. Pellet cultures designated for histological assessment were fixed, paraffin embedded, sectioned, and stained with hematoxylin and eosin (H&E). Immunohistochemical staining was performed with anti collagen type II monoclonal antibody as previously described [37]. Pellet cultures designated for real-time RT-PCR analysis were washed and digested with collagenase I (3 mg/mL; Worthington Biochemical) and dispase II (4 mg/mL; Roche Diagnostics), and RNA was isolated with TRIzol.
Quantitation analysis was performed by assessing the level of glycosaminoglycan (GAG) synthesis achieved by the three iPSC-MSC-like cell lines. GAG synthesis was performed in quadruplicate high-density cultures (1×105 cells per well in a 96-well plate). The level of GAG synthesis was measured over a 5 day period through 35SO4 incorporation using a TopCount NXT Microplate Scintillation and Luminescence counter (Perkin Elmer Life and Analytical Sciences). The level of GAG synthesis was normalized to DNA content per well. DNA quantitation was performed using the Quant-iT™ PicoGreen dsDNA Assay Kit as per the manufacturer's instructions, with absorbance at 540 nm measured using a Polar Star Optima micoplate reader.
Real-time PCR analysis
Total cellular RNA was isolated using TRIzol (Invitrogen) extraction as per the manufacturer's recommendations. Complementary DNA was generated using SuperScript III Reverse Transcriptase Kit in accordance with the manufacturer's instructions (Invitrogen). The expression of genes of interest was assessed by real-time PCR using RT2 SYBR Green/ROX qPCR Master Mix (Qiagen) using a Rotot-Gene 6000 Real Timer Thermal Cycler (Corbett Research). The cycling protocol used was 50°C for 2 min followed by 95°C for 15 min; this was followed by 40 cycles consisting of 95°C for 15 s, 60°C for 26 s, and 72°C for 10 s, finished off with 72°C for 3 min.
In vivo transplantation of iPS-MSC-like cells into NOD/SCID mice
Approximately 5×106 iPSC-MSC-like cells were mixed with 40 mg of hydroxyapatite/tricalcium phosphate ceramic particles (Zimmer Corporation) and then implanted into subcutaneous pockets on the dorsal surface of 2-month-old SCID mice as previously described [6]; implants were performed in duplicate. These procedures were performed in accordance with an approved animal protocol (IMVS/SA Pathology AEC No. 139/09). Implants were recovered after 8 weeks, fixed in 4% paraformaldehyde for 2 days, and then decalcified for a further 14 days in 10% EDTA before embedding in paraffin. For histological analysis, 5 and 8 μm sections of the implants were prepared. The 5 μm sections were used for H&E staining as well as immunohistochemical staining was performed. The 8 μm sections were used in the modified tetrachrome staining. Modified tetrachrome staining was performed as previously described [38].
The iPSC-MSC-like cells implanted were BrdU labeled at 24 and 48 h before implantation to facilitate assessment of the survival and localization of the transplanted cells. Immunohistochemical staining using an anti-BrdU antibody at 3 μg/mL (6.1 mg/mL; Sigma-Aldrich) was used to identify the implanted cells. The mouse anti human IgG isotype control, 1B5, at 3 μg/mL (1 mg/mL; Sigma-Aldrich) was used as a negative control for the BrdU staining. The staining intensity was assessed by observation under a light microscope.
Statistical analysis
All values are expressed as mean±standard deviation. To test statistical significance, the Student's t-test for paired data was used. A P-value<0.05 was considered statistically significant.
Results
In vitro induction of iPSC into cells that appear phenotypically similar to MSC-like populations
The differentiation method used to generate MSC-like cells from three different human iPSC lines was based on the flat culture, serial passage method previously used for human ESC lines [31]. This methodology utilizes selective passaging with TrypLE, which favored rapidly cycling, plastic adherent cells grown in the absence of feeder cells, thereby selecting for iPSC-MSC-like cells and eliminating slow growing differentiated cell types as well as non-induced iPSC. After 14 days of culture in MSC inductive media, GF-iPSC, PDL-iPSC, and lung-iPSC had undergone significant differentiation (Fig. 1A[I and II]). After sub-culture onto new gelatine-coated flasks, the cells exhibited a mixed morphology with a high proportion developing an epithelial-like appearance (Fig. 1A[III]). In subsequent passages onto standard culture flasks (without gelatine coating), there was a decrease in the proportion of cells with an epithelial-like morphology, with a concurrent increase in the proportion of cells with a fibroblastic-like morphology (Fig. 1A[IV], [V]). By passage 8 (post differentiation), the induced iPSC displayed a uniform fibroblastic-like morphology which was similar to that previously described for MSC (Fig. 1A[VI]), dramatically different when compared with non-induced iPSC (Fig. 1A[I], B[I]).

Cell morphology changes in the induction of induced pluripotent stem cell (iPSC) into mesenchymal stem cell (MSC)-like cells.
While all the MSC-like cell populations generated from the three different iPSC lines exhibited a fibroblast-like morphology, differences between the three populations were observed (Fig. 1B). In particular, the MSC-like cells derived from the lung-iPSC were smaller in size than the MSC-like cells derived from the GF-iPSC and PDL-iPSC lines. Therefore, the MSC-like cells generated from GF-iPSC and PDL-iPSC most closely resembled those of BM-MSC.
Flow cytometric analysis using a panel of MSC and pluripotency markers was used to determine whether the iPSC had successfully differentiated into MSC-like cells. The three non-induced iPSC lines were found to express a number of the MSC-associated markers before induction into MSC-like cells (Fig. 2 left panel), including CD90 (GF-iPSC—92.76%, PDL-iPSC—95.62%, and lung iPSC—96.35%), CD146 (GF-iPSC—85.91%, PDL-iPSC—86.5%, and lung iPSC—96.47%), and CD166 (GF-iPSC—86.32%, PDL-iPSC—57.62%, and lung iPSC—90.29%). The non-induced iPSC also expressed the pluripotency markers TRA-160 (GF-iPSC—96.21%, PDL-iPSC—94.2%, and lung iPSC—98.07%), TRA-181 (GF-iPSC—93.51%, PDL-iPSC—90.64%, and lung iPSC—98.25%), and ALKPHOS (GF-iPSC—86.29%, PDL-iPSC—56.3%, and lung iPSC—64.04%), but did not express the hematopoietic markers CD14 (GF-iPSC—0.8%, PDL-iPSC—0.15%, and lung iPSC—0.19%), CD34 (GF-iPSC—1.21%, PDL-iPSC—0.24%, and lung iPSC—0.05%), and CD45 (GF-iPSC—1.6%, PDL-iPSC—0.2%, and lung iPSC—0.03%). Of note, non-induced iPSC were negative for two key MSC markers, CD73 (GF-iPSC—0.42%, PDL-iPSC—0.72%, and lung iPSC—0.66%) and CD105 (GF-iPSC—1.27%, PDL-iPSC—0.28%, and lung iPSC—10.4%). The three non-induced iPSC lines were also negative for the MSC markers CD106 (GF-iPSC—1.17%, PDL-iPSC—6.65%, and lung iPSC—2.43%). After induction, more than 95% of cells expressed the MSC markers CD73 and CD105, while less than 5% of cells expressed the pluripotency markers TRA-160, TRA-180, and alkaline phosphatase. In addition, the MSC-like cells derived from the three iPSC lines maintained expression of the MSC markers CD90, CD146, and CD166. These cells also maintained their negative expression of the hematopoietic markers CD14, CD34, and CD45 (Fig. 2 right panel). Importantly, the MSC-like cells demonstrated little or no expression of the pluripotency markers TRA-160, TRA-180, and alkaline phosphatase. While there were noticeable differences in the cell morphology of the MSC-like cells derived from the three iPSC lines, CD106 was the only cell surface marker that showed differences in expression by flow analysis across the three MSC-like cell lines generated. The MSC-like cells derived from GF-iPSC were the only population that showed an up-regulation of CD106 expression (52.18% in the GF-iPSC-MSC-like cells compared with 1.17% in non-induced iPSC) (Fig. 2).
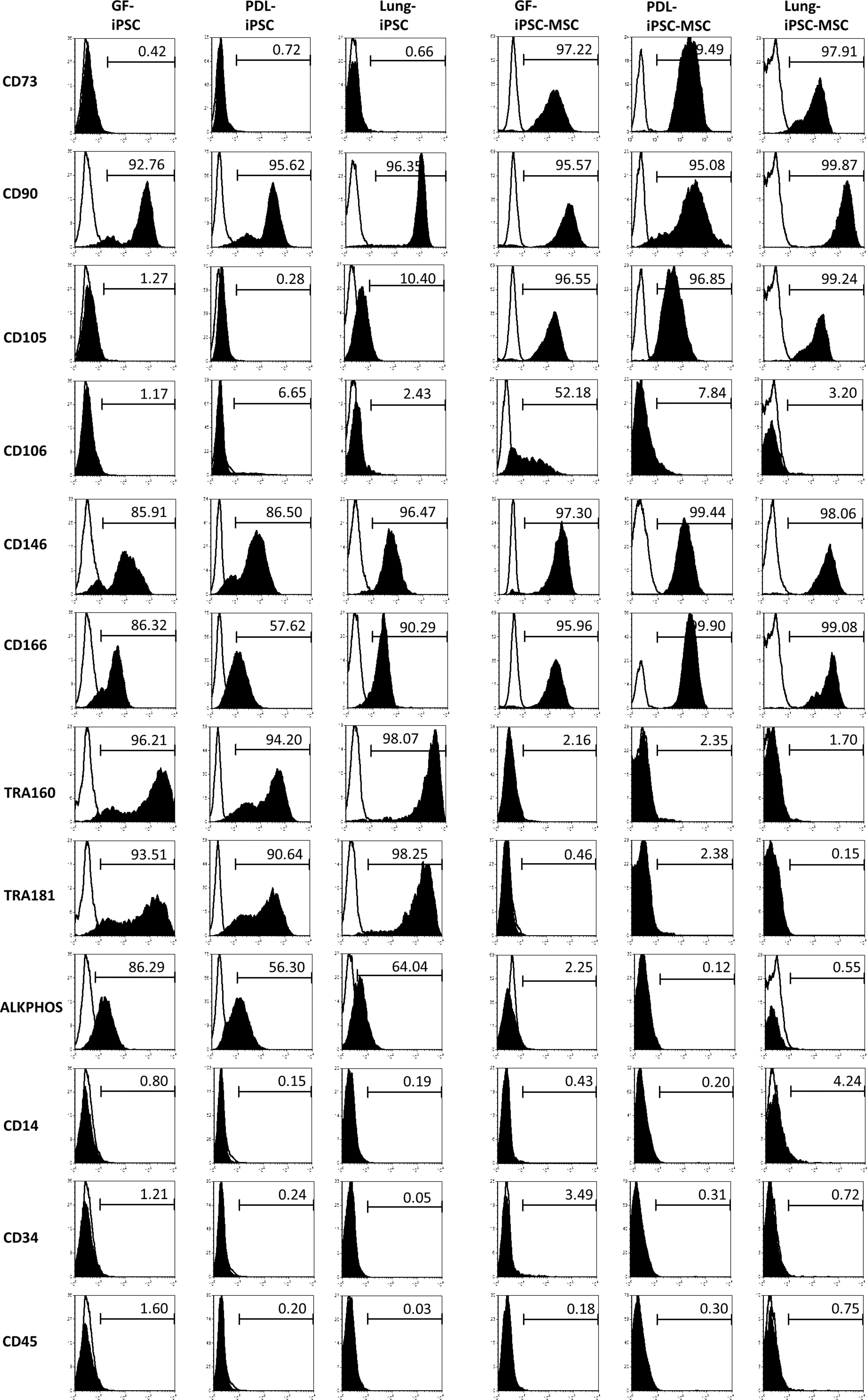
Immunophenotype of the three iPSC-MSC-like cell lines generated. Representative flow cytometry analysis of non-induced iPSCs [gingiva (GF)-iPSC-MSC, periodontal ligament (PDL)-iPSC-MSC, and lung-iPSC] on the left of the panel and the iPSC-MSC-like cells (GF-iPSC-MSC, PDL-iPSC-MSC, and Lung-iPSC-MSC) on the right of the panel. MSC-related markers CD73, CD90, CD105, CD106, CD146, and CD166 were assessed (solid histogram). Pluripotency-associated markers TRA160, TRA181, and alkaline phosphatise (ALKPHOS) as well as the hematopoietic markers CD14, CD34, and CD45 (solid histogram) were also assessed. Antibodies for immunoglobulin, 1B5 (IgG1), 1D4.5 (IgG2), and 1A6.12 (IgM) were used as isotype controls (open histogram).
Human iPSC-derived MSC-like cells demonstrate multi-lineage differentiation in vitro
One of the defining characteristics of MSC is an ability to undergo tri-lineage differentiation into the three lineages of mesoderm origin: osteocytes/cementocytes, adipocytes, and chondrocytes. Therefore, tri-lineage differentiation was performed on the cells in vitro to confirm the derivation of MSC-like cells from the three iPSC lines.
The capacity of the iPSC-MSC-like cells to undergo osteogenic differentiation in vitro was investigated by growing the iPS-MSC-like cells in osteoinductive media containing dexamethasone,
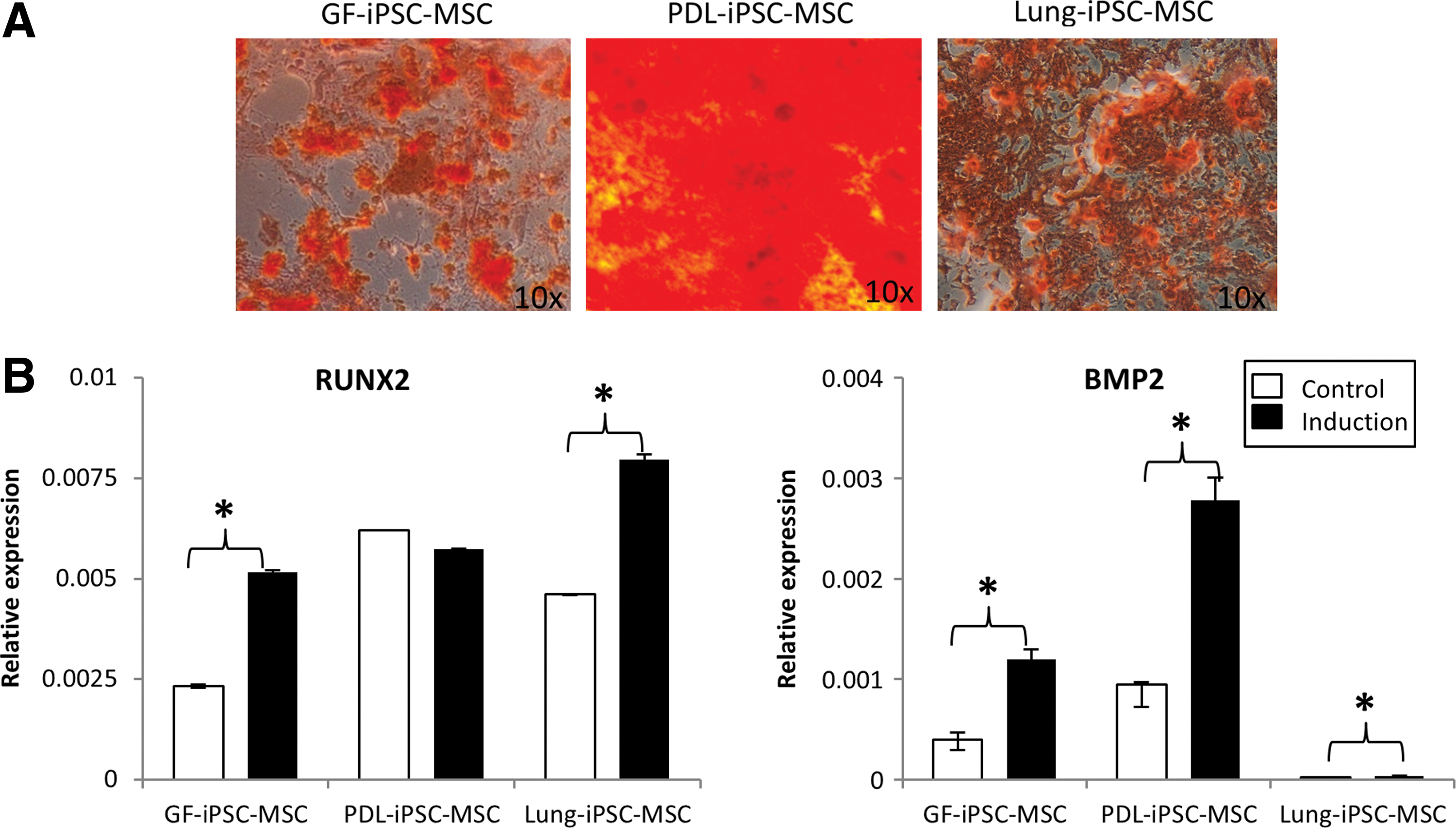
Osteogenic differentiation potential of the three iPSC-MSC-like cell lines generated.
The capacity of the iPSC-MSC-like cells to undergo adipogenic differentiation in vitro was investigated by growing the iPSC-MSC-like cells in adipogenic media containing hydrocortisone, indomethacin, and 3-isobutyl-1-methyl-xanthine. The iPSC-MSC-like cells differentiated into Oil Red O-positive lipid-containing adipocytes at similar levels for all three tissue types (Fig. 4A). Real-time RT-PCR analysis demonstrated up-regulated expression of peroxisome proliferator-activated receptor gamma-2 (PPARγ2) and adipogenic-associated markers, complement factor D (ADIPSIN) (Fig. 4B) (n=3, P<0.05), after adipogenic induction of all iPSC-MSC-like cell lines.
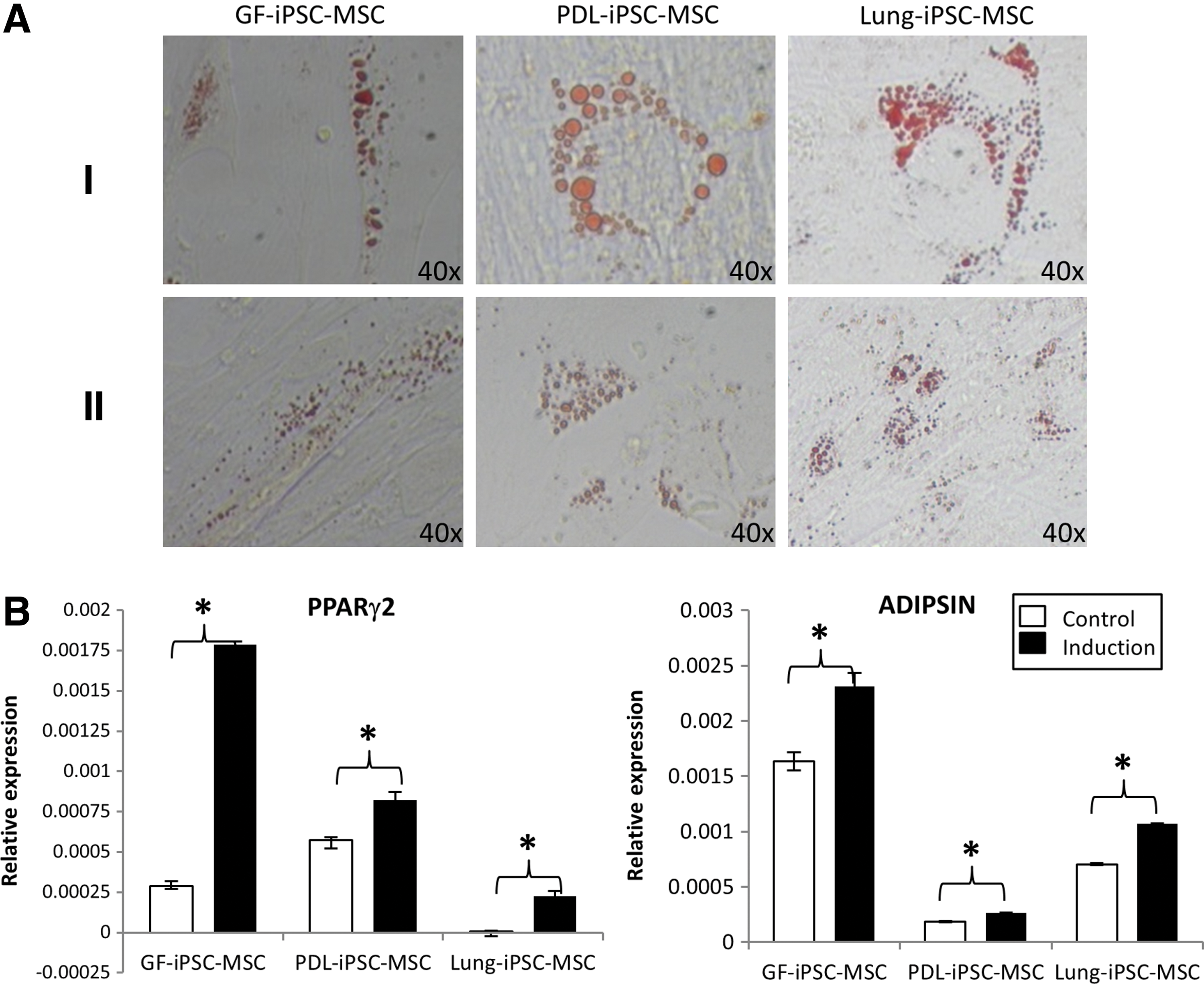
Adipogenic differentiation potential of the three iPSC-MSC-like cell lines generated.
Finally, the potential of the three iPSC-MSC-like cell lines to differentiate the chondrogenic lineage was examined. The three iPSC-MSC-like cell lines were cultured in three-dimensional cell aggregates in the presence of transforming growth factor-β3, BSA, dexamethasone, and ITS+Premix. Two of the three iPSC-MSC-like cell lines (GF-iPSC-MSC and PDL-iPSC-MSC) formed tight compact pellets, while the lung-iPSC-MSC-like cells formed a larger and less compact pellet (Fig. 5A). Histological staining of the chondrogenic pellets collagen II revealed that the pellets were composed of cells that were embedded within a sulfate-proteoglycan-rich extra-cellular matrix comprising collagen type II protein (Fig. 5A[I and II]).
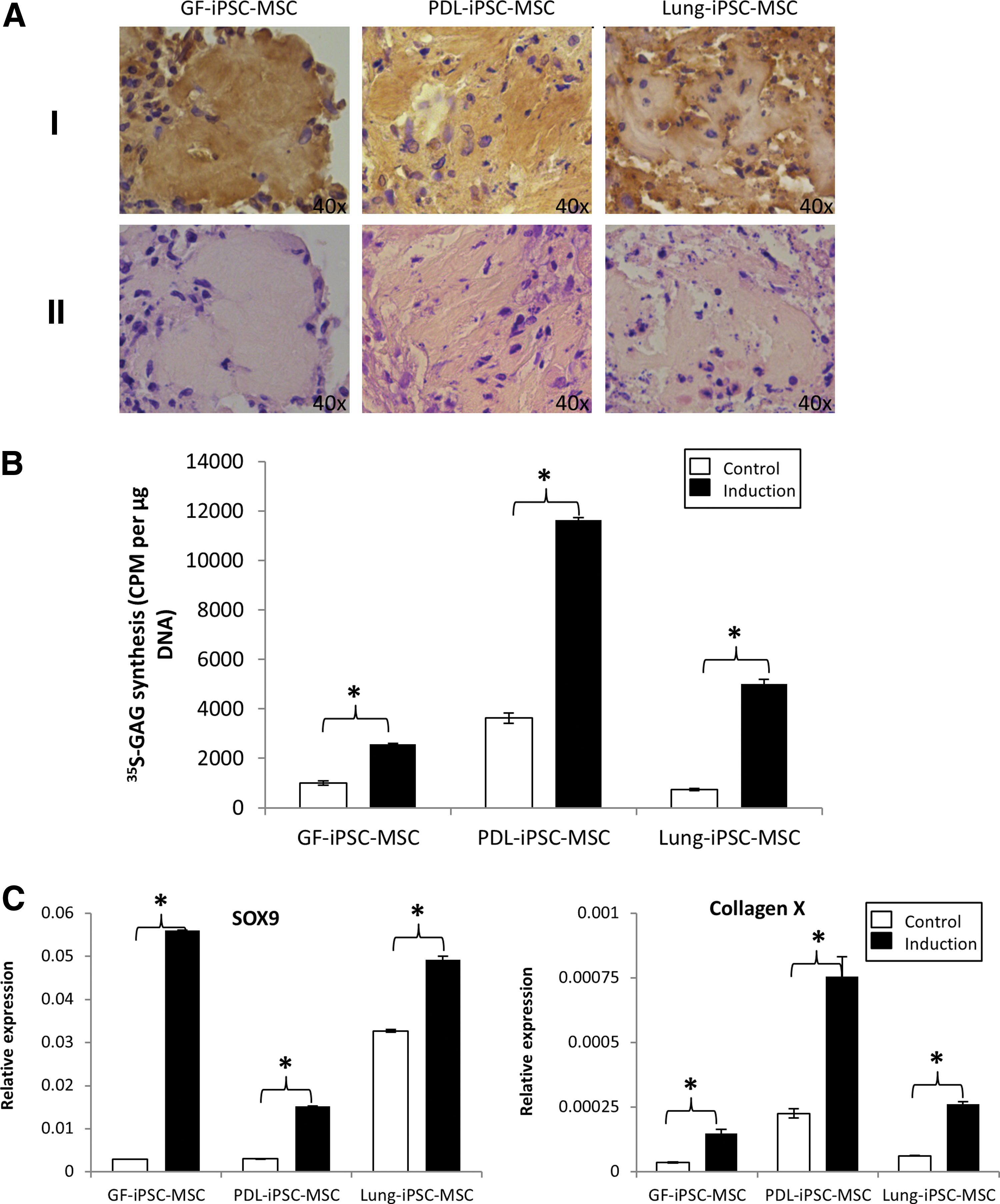
Chondrogenic differentiation potential of the three iPSC-MSC-like cell lines generated.
Quantitative GAG synthesis analysis confirmed that all three iPSC-MSC-like cells lines had the capacity to undergo chondrogenesis, as all three cell lines generated significantly more GAG when cultured in inductive conditions than in control conditions. Overall, the PDL-iPSC-MSC-like cells generated the most GAG, reinforcing the histology results by which the PDL-iPSC-MSC chondrocyte pellets appeared the most chondrocyte like (Fig. 5B).
Gene expression studies confirmed up-regulated expression of the chondrocyte-associated markers SOX9 and collagen type X after 4 weeks of chondrogenic differentiation in all three iPSC-MSC-like cell lines assessed (Fig. 5C).
Human iPSC-MSC-like cells derived from PDL tissue demonstrate the capacity to regenerate bone in vivo
To assess the in vivo differentiation capacity of the iPSC-MSC-like cells, ∼5×106 BrdU-stained cells were combined with an HA/TCP carrier and implanted subcutaneously into the dorsal surface of NOD/SCID mice. After 8 weeks, the implants were retrieved and assessed histologically with H&E, modified tetrachrome, and anti-BrdU staining. A histomorphic analysis revealed the presence of mineralized structures (Fig. 6A), adipocytes (Fig. 6B), and fibrous connective tissues (Fig. 6C) in the implants containing the iPSC-MSC-like cell lines. The H&E and modified tetrachrome staining were performed to identify regions of mineralized tissue and to characterize their composition, respectively. While bright pink H&E staining (Fig. 6A[I]) is indicative of mineralized tissue, the modified tetrachrome stain (Fig. 6A[II]) can be used to distinguish osteoid tissue (deep blue), the mineralization front (pale blue-pink), woven bone (blue with red patches), and cartilage and connective tissue fibers (pale blue) from each other [38]. Together, the H&E and modified tetrachrome staining demonstrated that all three iPSC-MSC-like cell lines had the capacity to form mineralized tissue in vivo, as the modified tetrachrome revealed the presence of a mineralization front in the implants from all three cell iPSC-MSC-like cell lines. Furthermore, the H&E staining demonstrated the presence of adipocytes and fibrous tissue in the implants from all three iPSC-MSC-like cell lines. Together, these results demonstrate that all three iPSC-MSC-like cell lines have the capacity to undergo tri-lineage differentiation in vivo.
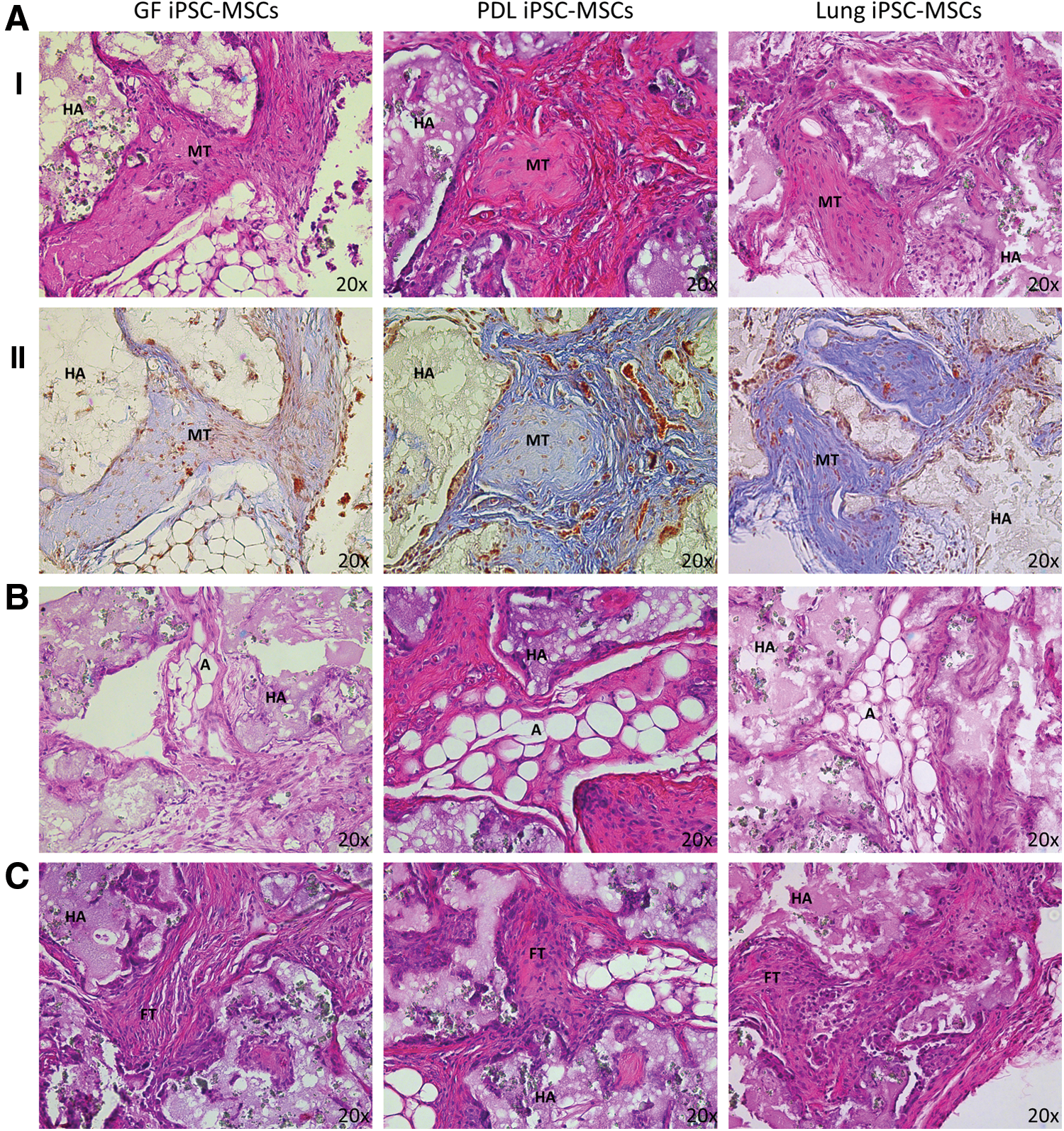
In vivo tri-lineage differentiation capacity of iPSC-MSC-like cells. MSC-like cells derived from iPSC were seeded onto hydroxyapatite tricalcium phosphate (HA/TCP) carriers and then implanted subcutaneously into immunocompromised mice. Representative images of cross-sections are shown of implants at 8 weeks post transplantation.
Anti-BrdU staining revealed the presence of the implanted iPSC-MSC-like cells in all the structures mentioned earlier (Fig. 7A), indicating that the implanted iPSC-MSC-like cells contributed to the formation of all three tissue lineages. It should be noted that intrinsic cells are likely to have also contributed to the formation of the structures mentioned earlier. Previous research has demonstrated that MSCs are capable of not only directly differentiating into a required tissue but also secreting bioactive molecules which stimulate endogenous progenitors to also differentiate [40].
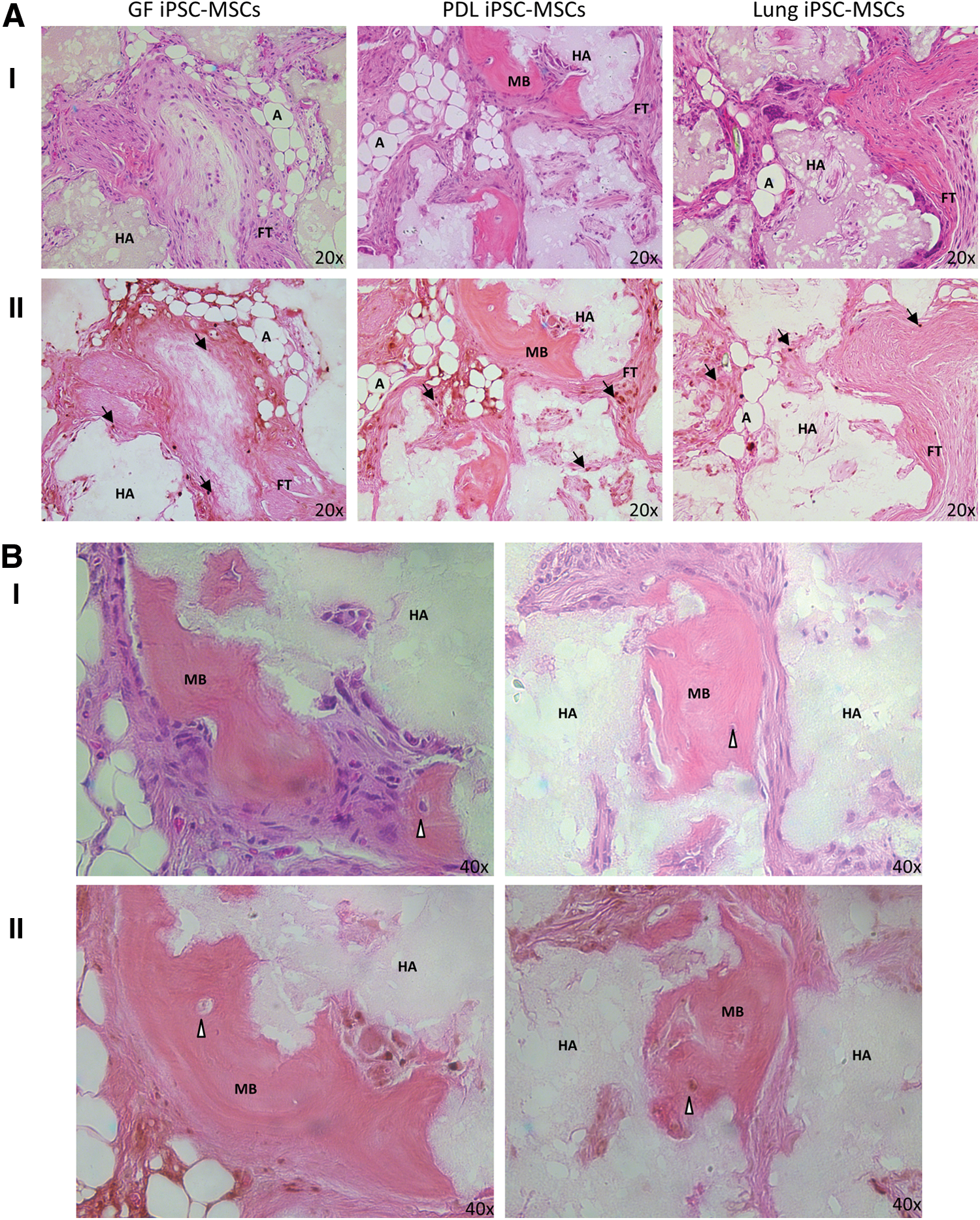
In vivo implantation of iPSC-MSC-like cells. H&E
Interestingly, only iPSC-MSC-like cells derived from PDL tissues formed physiological mature bone structures in vivo (Fig. 7B). Anti-Brdu staining showed the presence of BrdU-positive osteoblasts and cells embedded within the bone which morphologically appear to be osteocyte-like cells within and surrounding the mature bone, indicating that the PDL-iPSC-MSC-like cells had contributed to the formation of the bone.
Discussion
With an ever-increasing interest in the use of MSC for therapeutic purposes, it is imperative that new sources of MSC are identified. Current methodologies used to obtain MSC for clinical treatments are invasive, laborious, and expensive. Furthermore, MSC have a limited expansion capacity in vitro, as extensively cultured primary MSC tend to lose their stem cell properties, including their ability to undergo multi-lineage differentiation. Thus, obtaining sufficient quantities of MSC cells for therapeutic purposes is a significant challenge and limits their use in mainstream treatment approaches. A number of research groups have successfully derived MSC-like cells from iPSC, and these cells are emerging as an attractive alternative to primary MSC.
In this study, we developed a simple and reproducible method for inducing iPSC into MSC-like cells, without the requirement of any exogenous differentiation factors. Our protocol consists of two main stages, the first of which was a 2 week induction period followed by serial passaging to select for MSC-like cells. By utilizing this methodology, we successfully generated MSC-like cells from iPSC generated from three different somatic tissues: PDL, gingiva, and lung.
The MSC-like cells generated in this study met the International Society of Cellular Therapy's criteria for defining an MSC population. These criteria consist of three components: (i) adherence to tissue culture plastic under standard culture condition; (ii) an immunophenotype similar to that of bone marrow-derived MSC with ≥95% of the MSC expressing CD105, CD73, and CD90 combined with low expression of the hematopoietic markers of CD45, CD34, and CD14; and (iii) the ability to undergo tri-lineage differentiation to osteoblasts, adipocytes, and chondrocytes under standard in vitro differentiation protocols [19,20]. However, these criteria are not specific to MSC, as some properties are common with non-MSC fibroblast populations exhibiting limited differentiation potential [41]. More importantly, these minimal criteria fail to address the issue of stem cell self-renewal and tissue regeneration in vivo.
These findings add further support to previous works demonstrating that iPSC are an attractive source for obtaining large numbers of MSC required for numerous therapeutic applications [23 –30]. While the results outlined in this article support previous findings, they also demonstrate that MSC-like cells derived from different iPSC lines display different characteristics. This indicates that differences in the developmental capacity and cell surface marker expression profile exist between MSC derived from iPSC generated from different starting tissues. Although all three iPSC-MSC-like cell lines that were generated demonstrated the capacity to form mineralized deposits in vitro, the MSC-like cells generated from the PDL-iPSC showed the greatest capacity to form mineralized structures. These results imply that differences in reprogramming may occur after MSC induction of different iPSC lines. In the present study, we extended the assessment of the differentiation capacity of the iPSC-derived MSC-like cells in vivo, after subcutaneous implantation with an osteoconductive scaffold carrier into immunocompromised mice. These studies confirmed the in vitro results that iPSC-MSC-like cells derived from PDL tissue exhibited a superior capacity to form physiological bone and connective tissue.
The PDL tissue comprises a group of specialized connective ligament fibers that are embedded between cementum on the surface of the tooth root and the alveolar bone socket of the jaw bone. The PDL functions to attach the tooth into the alveolar bone, while enabling a degree of movement, and it is involved in tooth nutrition, homeostasis, and repair [42 –44]. In vitro studies have demonstrated that specific cells isolated from PDL exhibit an osteoblast-like phenotype and have the capacity to form mineralized deposits [45 –47]. In addition, MSC-like cells isolated from PDL have the capacity to form ligamentous structures resembling Sharpey's fibers and mineralized tissues similar to bone and cementum in vivo [8]. The finding that the PDL-iPSC-MSC-like cells had the greatest capacity to form mineralized tissues both in vitro and in vivo has led us to hypothesize that even after iPSC are induced into MSC-like cells, they may still maintain epigenetic memory of their tissue of origin. However, a number of other factors could have contributed to the differences in differentiation capacity of the iPSC-MSC-like cells. It is important to note that the iPSC lines used in this study were generated in two different laboratories using two different cocktails of transcription factors. The lung iPSC were generated with OCT4, SOX2, NANOG, and LIN28; while the PDL and GF-iPSC were generated with OCT4, SOX2, cMyc, and KLF4. The iPSC lines used have also been generated from different individuals, so there will be inherent differences in the lines and while this may not affect the differentiation capacity of the resulting iPSC-MSC-like cells, we are unable to eliminate this as a possibility at this time. Therefore, it would be informative in the future to repeat this work on iPSC-MSC-like cells generated from iPSC derived from different tissues of a single individual and with the same cocktail of transcription factors.
There are contradictory findings in the literature with regard to whether iPSC possess a memory for their tissue of origin, and whether this leads to an increased propensity to develop back into their original cell type. Various studies have described the presence of the epigenetic memory of iPSC for their somatic tissue of origin that can influence the in vitro differentiation capacity of the iPSC [48 –50]. This memory was, subsequently, lost through a continuous subculture of iPSC [48 –50]. More recent reports have, however, shown that iPSC retain the epigenetic memory of their tissue of origin even after an extensive passage [51 –54], where the epigenetic memory skews the developmental potential of iPSC toward their previous state of differentiation [51 –54]. The findings of this study could suggest that even after iPSC are induced into MSC-like cells, they may still have a memory of their tissue of origin. However, a number of additional factors may have led to the differences in differentiation capacity seen between the iPSC-MSC-like cells; so, at this time, we cannot definitively attribute the differences to memory alone.
While we are currently unable to definitively show that the differences in the differentiation capabilities of the iPSC-MSC-like cells are a result of the cells retaining a memory of their original cell type, our results highlight the importance of a thorough characterization of iPSC-MSC-like cells before they can be considered for therapeutic use. Our results demonstrate that iPSC-MSC-like cells derived from different iPSC lines have subtle differences which may have significant implications for their therapeutic use.
Although the MSC-like cells derived from GF-iPSC had the capacity to form significant mineralized calcium deposits in vitro and mineralized tissues in vivo, primary MSC derived from GF tissue did not have the same mineral-forming capacity. MSC derived from primary gingival tissue were capable of undergoing multi-lineage differentiation in vitro, whereas they were only capable of forming connective tissue-like structures in vivo [12,55,56]. Our results show that the generation of MSC-like cells from GF-iPSC produced cells with a greater capacity to form mineralized tissues, adipocytes, and connective tissue-like structures in vivo. These findings imply that while iPSC-MSC-like cells may maintain an epigenetic memory of their original tissue type, the in vitro induction process conferred additional differentiation capabilities onto these cells.
While the iPSC-MSC-like cells displayed the capacity to undergo tri-lineage differentiation in vitro, the osteogenic and chondrogenic differentiation appeared to be more pronounced than adipogenic differentiation. These observations are supported by previous studies describing the properties of MSC derived from different populations [57,58]. Therefore, it remains to be determined whether the differences in differentiation propensity result from the derivation of MSC from iPSC or whether they are dictated by the tissue of origin of the parental iPSC. The results outlined in this article indicate that all the MSC-like cells generated from three different iPSC lines had a greater capacity for undergoing osteogenic and chondrogenic differentiation than adipogenic differentiation. This suggests that the propensity toward osteogenic and chondrogenic differentiation could result from the derivation of MSC from iPSC; however, further investigations are required before this can be determined. The increased propensity of the iPSC-MSC-like cells to form osteoblasts and chondrocytes makes these cells attractive for use in regenerative therapies requiring the generation of bone and connective tissues in dental and skeletal applications.
Conclusion
In this study, three different human iPSC lines were successfully induced to form MSC-like cells and while all of the lines generated satisfied the selection criteria for MSC, they had different propensities to differentiate into certain lineages. The results support the notion that iPSC could be a valuable source of MSC for use in a range of clinical applications. The results also highlight that differences in the regenerative capacity of iPSC-MSC-like cells derived from different iPSC lines exist. Detailed characterization and careful consideration will be needed to determine the most suitable population of iPSC and resulting iPSC-MSC-like cells for use in the regeneration of a desired tissue.
Footnotes
Acknowledgments
The authors would like to acknowledge funding received from the National Health and Medical Council (Australia) grant no. 627143 and the Australian Dental Research Foundation grant no. 72/2011.
Author Disclosure Statement
The authors declare no conflicts of interest with regard to the publication of this article.