
Abstract
Human skin-derived precursors (hSKP) are postnatal stem cells with neural crest properties that reside in the dermis of human skin. These cells can be easily isolated from small (fore) skin segments and have the capacity to differentiate into multiple cell types. In this study, we show that upon exposure to hepatogenic growth factors and cytokines, hSKP acquire sufficient hepatic features that could make these cells suitable in vitro tools for hepatotoxicity screening of new chemical entities and already existing pharmaceutical compounds. Indeed, hepatic differentiated hSKP [hSKP-derived hepatic progenitor cells (hSKP-HPC)] express hepatic progenitor cell markers (EPCAM, NCAM2, PROM1) and adult hepatocyte markers (ALB), as well as key biotransformation enzymes (CYP1B1, FMO1, GSTA4, GSTM3) and influx and efflux drug transporters (ABCC4, ABCA1, SLC2A5). Using a toxicogenomics approach, we could demonstrate that hSKP-HPC respond to acetaminophen exposure in a comparable way to primary human hepatocytes in culture. The toxicological responses “liver damage”, “liver proliferation”, “liver necrosis” and “liver steatosis” were found to be significantly enriched in both in vitro models. Also genes associated with either cytotoxic responses or induction of apoptosis (BCL2L11, FOS, HMOX1, TIMP3, and AHR) were commonly upregulated and might represent future molecular biomarkers for hepatotoxicity. In conclusion, our data gives a first indication that hSKP-HPC might represent a suitable preclinical model for in vitro screening of hepatotoxicity. To the best of our knowledge, this is the first report in which human postnatal stem cells derived from skin are described as a potentially relevant cell source for in vitro hepatotoxicity testing of pharmaceutical compounds.
Introduction
T
Since the liver is a first-line target organ during toxicant assault, human-based in vitro hepatic cell systems are valuable tools to study, the potential hepatotoxicity of pharmaceuticals and other substances. Freshly isolated human hepatocytes represent the model that most appropriately reflects the in vivo situation and are considered as the gold standard in the area of liver-based in vitro modelling [11]. Human hepatocytes, however, are difficult to obtain because of intensified liver transplantation programmes. In addition, these cells are phenotypically unstable and have a limited life span [11]. Therefore, immortalized human liver cell lines, including HepG2 [12,13], Fa2N-4 [14], and HepaRG [15] have become increasingly popular as in vitro models to study human liver function and toxicity. These cell lines are readily available and can be kept in culture for long periods of time, but they suffer from genotypical instability and decreased or even absent metabolic capacity. In addition, currently available cell lines do not represent population diversity [16]. Stem cells could form a novel and attractive alternative human cell source to develop in vitro systems for predicting liver toxicity. Indeed, the establishment of continuous cell lines from embryonic stem cells [17] and induced pluripotent stem cells [18], the advancements in isolating and culturing adult stem cells (ASC) [19] and in particular the breakthrough in differentiating them into cells with particular functionalities brought huge expectations to the scientific and industrial community.
ASC reside in specialized stem cell niches [20] from where they contribute to tissue renewal and/or regeneration [21]. These cells have the ability to give rise to a range of cells of the tissue within which they reside.
ASC can be isolated from a wide range of tissues and are able to differentiate towards their hepatic progeny. Among others, ASC isolated from bone marrow [22], adipose tissue [23], umbilical cord matrix [24], umbilical cord epithelial cell layer [25], and placenta [26] have been successfully differentiated towards cells with hepatic characteristics. In this study, human skin-derived precursors (hSKP) and their hepatic derivatives are evaluated.
hSKP are a multipotent cell population isolated from human dermal (fore)skin [27 –29]. These neural crest related ASC can also be isolated from skin of the abdomen [30], breast [30], arm [31], face [32], and scalp [33] mostly upon aesthetic interventions. These cells have a high self-renewal and high multipotent differentiation capacity. More specifically, it was previously shown by our group that hSKP, besides their ectodermal and mesodermal differentiation potential, can be directed towards the hepatic lineage [34 –36] upon sequential exposure to growth factors and cytokines that mimic liver development in vivo [22].
The present study focuses on the actual application of hepatic differentiated hSKP as a potential novel in vitro model for hepatotoxicity screening of chemical substances, in casu pharmaceuticals. Acetyl-para-aminophenol, acetaminophen (APAP), which is a common over-the-counter analgesic considered to be safe when used at therapeutic doses, is used as a proof-of-principle reference compound. When taken in overdose, APAP becomes hepatotoxic and can cause ALF, the latter being a leading cause of DILI [37]. The toxic mechanism of APAP in adult hepatocytes is known to involve several toxicity pathways [38,39]. It is acknowledged that APAP is metabolically activated to N-acetyl-p-benzoquinone imine (NAPQI), which is normally detoxified by glutathione. Depletion of glutathione after APAP overdose leads to NAPQI accumulation that adversely binds to different cellular proteins causing toxicity. Here we evaluate to what extent APAP exposure modulates the whole genome expression of hepatic differentiated hSKP versus primary human hepatocytes by using a full genome microarray platform.
Materials and Methods
Isolation and cultivation of hSKP
hSKP were isolated from foreskin circumcision samples of 1- to 10-year old boys after informed consent of the parents. The cells were isolated as previously described [36]. After isolation, cells were seeded at a density of 7.5×104 viable cells/mL in growth medium composed of Dulbecco's modified Eagle medium (DMEM)+GLUTAMAX/F12 Nutrient Mixture (3:1; Life Technologies) supplemented with 7.3 IU/mL benzyl penicillin (Continental Pharma), 50 mg/mL streptomycin sulfate (Sigma-Aldrich), 2.5 mg/mL fungizone, 2% (v/v) B27 Supplement (Life Technologies), 40 ng/mL basic fibroblast growth factor (FGF) 2, and 20 ng/mL epidermal growth factor (both from Promega). The cells were cultured in suspension at 37°C in a humidified incubator under an atmosphere of 5% (v/v) CO2. Every 2–3 days the growth medium was replenished. After 2 weeks of culture, the hSKP spheres were passaged using 0.2 mg/mL Liberase DH solution (Roche Applied Science). Obtained single cells were seeded at a density of 1.3×104 cells/cm2. After a second passage (7 days later), sufficient cells were obtained to establish a small cryopreserved cell bank. hSKP were collected in a cryoprotectant solution composed of 90% (v/v) fetal bovine serum (Hyclone) and 10% (v/v) dimethylsulfoxide (cell culture grade; Sigma-Aldrich) at a density of 1×106 cells/mL and preserved in liquid nitrogen. All further experiments were based on these batches to rule out donor variability.
Hepatogenic differentiation of hSKP
hSKP were differentiated after a slightly modified protocol established previously by our group [36]. In essence, cells were seeded on collagen type 1 (BD Biosciences)-coated 24-well plates and T75 culture flasks (both BD Falcon) and cultured until 90% confluence in Basal Medium (BM) was reached. BM consisted of DMEM+GLUTAMAX/F12 Nutrient Mixture (3:1; Life Technologies) supplemented with 7.33 IU/mL benzyl penicillin (Continental Pharma), 50 mg/mL streptomycin sulfate (Sigma-Aldrich), 2.5 mg/mL fungizone (Life Technologies), 0.1 mM
Human liver samples
Human liver biopsy samples from deceased healthy persons were obtained from the UZ Brussel upon informed consent of the family and approval of the Ethics Committee. Small pieces of liver tissue were snap frozen and further stored in liquid nitrogen.
Cytotoxicity assessment and IC10 determination
After 24 days of hepatogenic differentiation, cells were exposed for 24 h to six concentrations of APAP (Sigma-Aldrich) ranging from 5 to 500 μg/mL (corresponding to 33 μM to 3.3 mM, respectively). Subsequently, the effect of APAP on cell viability was evaluated by a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) viability assay as previously described [40]. The 24-h IC10 value was determined by a four-parameter logistic nonlinear regression analysis of the obtained dose response curves, using Masterplex Readerfit 2010 software (Hitashi Solutions). The test was conducted in triplicate.
Isolation of total RNA
Treated and untreated cells were collected in a RNA protecting solution composed of RNA stabilization reagent (Qiagen) and BM (5:1). Cell cultures of three 75 cm2 culture flasks were used per condition. Total RNA was extracted from all samples using a GenElute Mammalian Total RNA Purification Miniprep Kit (Sigma-Aldrich), according to the manufacturer's instructions. Purity and quantification of the isolated RNA were determined by spectrophotometric analysis using a Nanodrop spectrophotometer (Thermo Scientific).
Microarray data analysis
Microarray technologies from Affymetrix (including reagents and instrumentation) were used for whole genome expression analysis. One hundred nanogram total RNA per sample was amplified using a Genechip 3′ IVT Express Kit, following the manufacturer's instructions (Affymetrix). The amplified RNA (aRNA) was purified with magnetic beads and 15 mg Biotin-aRNA was treated with the fragmentation reagent. 12.5 mg fragmented aRNA was subsequently hybridized to Affymetrix Human Genome U133 plus 2.0 arrays and placed in a Genechip Hybridization Oven-645 (Affymetrix) rotating at 60 rpm at 45°C for 16 h. After incubation, the arrays were washed on a Genechip Fluidics Station-450 (Affymetrix) and stained with an Affymetrix HWS kit as indicated by the manufacturer's protocols. The chips were scanned with an Affymetrix Gene-Chip Scanner-3000-7G, and quality control matrices were confirmed with Affymetrix GCOS software, following the manufacturer's guidelines. Microarray data of human hepatocyte cultures (hHEP) established from human cryopreserved hepatocytes were obtained from the toxicology database TG-GATEs. The latter being established by a 5-year collaboration between the National Institute of Health Sciences and 17 pharmaceutical companies in Japan [41,42]. Only data from cells exposed for 24 h to 5 mM APAP and the respective vehicle controls was used for comparative gene expression analysis with self obtained data. Background correction, summarization, and normalization of all data were carried out with Robust Multiarray Analysis [43]. For principal component analysis (PCA) and determination of differential gene expression, output data files were analyzed with GeneSpring GX v11.5 software (Agilent Technologies). Functional toxicological analyses were performed using Ingenuity Pathways Analysis (version SEP 2011; Ingenuity Systems) with a Fisher's Exact test (P value<0.05) to identify enriched functional gene classes.
Reverse transcriptase quantitative polymerase chain reaction
After total RNA isolation, as described above, mRNA was reverse-transcribed into cDNA using an iScript-cDNA Synthesis Kit (BioRad), followed by cDNA purification with a GenElute PCR Clean-Up Kit (Sigma-Aldrich). These cDNA products were then quantitatively amplified using Taqman probes and primers specific to the target genes of interest. The gene expression assays used in this study are listed in Supplementary Table S1 (Supplementary Data are available online at
qPCR data analysis
The qPCR efficiency was estimated by StepOne Plus System's Software, and only data with PCR efficiency between 0.85 and 1.15 was used. Four stable reference genes were identified to normalize the qPCR data as determined by geNorm (Biogazelle). These were glyceraldehyde 3-phosphate dehydrogenase (GAPDH), beta-2-microglobulin (B2M), hydroxy-methylbilane synthase (HMBS), and ubiquitin C (UBC). Results are expressed as fold changes normalized against the geometric mean of these four reference genes using qbasePLUS software (Biogazelle). Statistical analyses were performed by a one-way unpaired analysis of variance test with correction for multiple testing. Gene expressions with a fold change (up- or downregulation) of at least two and a corrected P value of 0.05 or lower were considered to be significantly different.
Immunocytochemistry
Cells were fixated with 4% (w/v) paraformaldehyde, and then further exposed to 100 mM glycin. After permeabilization with 0.1% (v/v) Triton X, nonspecific sites were blocked with 10% (v/v) donkey serum. Immunolocalization was obtained by incubation of the cells with the following primary antibodies: antialbumin (Alb; Bethyl Laboratories-A80-129F), antiaryl hydrocarbon receptor (Ahr; Santa Cruz-8089c), antihepatocyte nuclear factor (Hnf) 4α (Santa Cruz—8987), anti-Hnf1α (Santa Cruz—10791), anticytokeratin (Ck) 18 (Abcam—ab93741), anti-Ck19 (Abcam—ab76539), anti-GATA motif binding factor (GATA) 6 (Abcam—ab22600) antiepithelial cell adhesion molecule (Epcam; Sigma-Aldrich—SAB3300055), anticytochrome P450 (CYP) 1b1 (Sigma-Aldrich—HPA026863), antiflavin containing monooxygenase (Fmo) 1 (Sigma-Aldrich—HPA023680), antineural cell adhesion molecule (Ncam) 2 (Sigma-Aldrich—HPA030900) and antiprominin (Prom) 1 (Miltenyi Biotec—130-090-851). The secondary antibodies used were: anti-rabbit DyLight 488 (711-485-152), anti-mouse DyLight 488 (715-485-150) and anti-goat DyLight 488 (705-485-147) obtained from Jackson ImmunoResearch Europe. After washing with phosphate buffered saline, the cells were mounted with Vectashield with 4′,6-diamidino-2-phenylindole (Vector Laboratories) for nuclear staining and bleaching protection. Images were obtained by fluorescence microscopy (Nikon Eclipse Ti-S).
Results
Characterization of hSKP-derived hepatic progeny
Hepatogenic differentiation strongly modulates the gene expression of hSKP. After 24 days of sequential exposure to hepatogenic growth factors and cytokines, 6685 and 4245 genes are at least twofold up or downregulated, respectively (Student t-test with P value<0.05). A 10-fold upregulation or more is observed for 299 genes, whereas 86 genes are at least 10-fold downregulated. As illustrated by the PCA plots shown in Fig. 1A, samples of both undifferentiated (hSKP) and hepatic differentiated hSKP, further described as hSKP-derived hepatic progenitor cell (hSKP-HPC), group well together. The latter shows the robustness of the cell culture conditions and the high reproducibility of the microarray datasets. These plots also highlight a shift of the hSKP-HPC towards hHEP and human liver samples (LIVER). Further analysis of the hSKP-HPC shows that, compared to undifferentiated hSKP, these cells express significantly higher levels of typical hepatic progenitor cell markers, including EPCAM, GATA motif binding protein 6 (GATA6), prominin 1 (PROM1), NCAM, SMAD family member 4 (SMAD4), stem cell factor receptor (KIT), and thymocyte differentiation antigen 1 (THY1) (Fig. 1B). For some of these markers, the observations made are confirmed at the protein level (Fig. 2). Immunocytochemistry analysis also shows that hSKP-HPC express markers typical of more mature hepatic cells, such as albumin, HNF 1α and 4α (HNF1α, HNF4α) and Ahr (Fig. 2). 27% (±5%) of hSKP-HPC (counted in three independent experiments) are albumin-positive. This value represents the yield of hepatogenic differentiation. Additionally, it can be observed that Gata6, Hnf1α, Hnf4α, C-Kit, Ahr, and Fmo1 are mainly located in the nucleus. Albumin, Prom1, Ncam2, and Cyp1B1 are as expected mainly cytoplasmatic or membrane-bound.
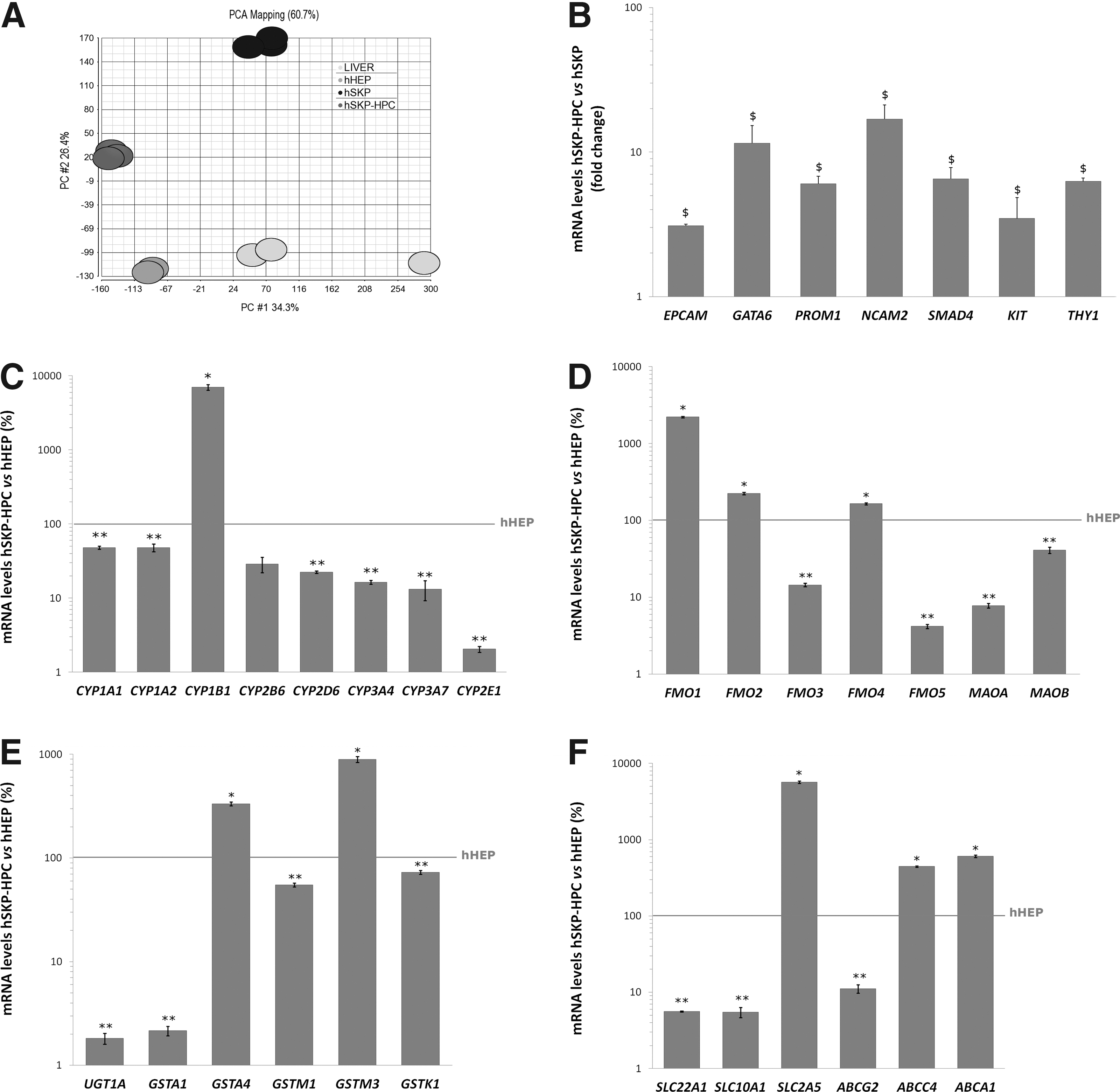
Characterization of human skin-derived precursors (hSKP)-hepatic progenitor cells.
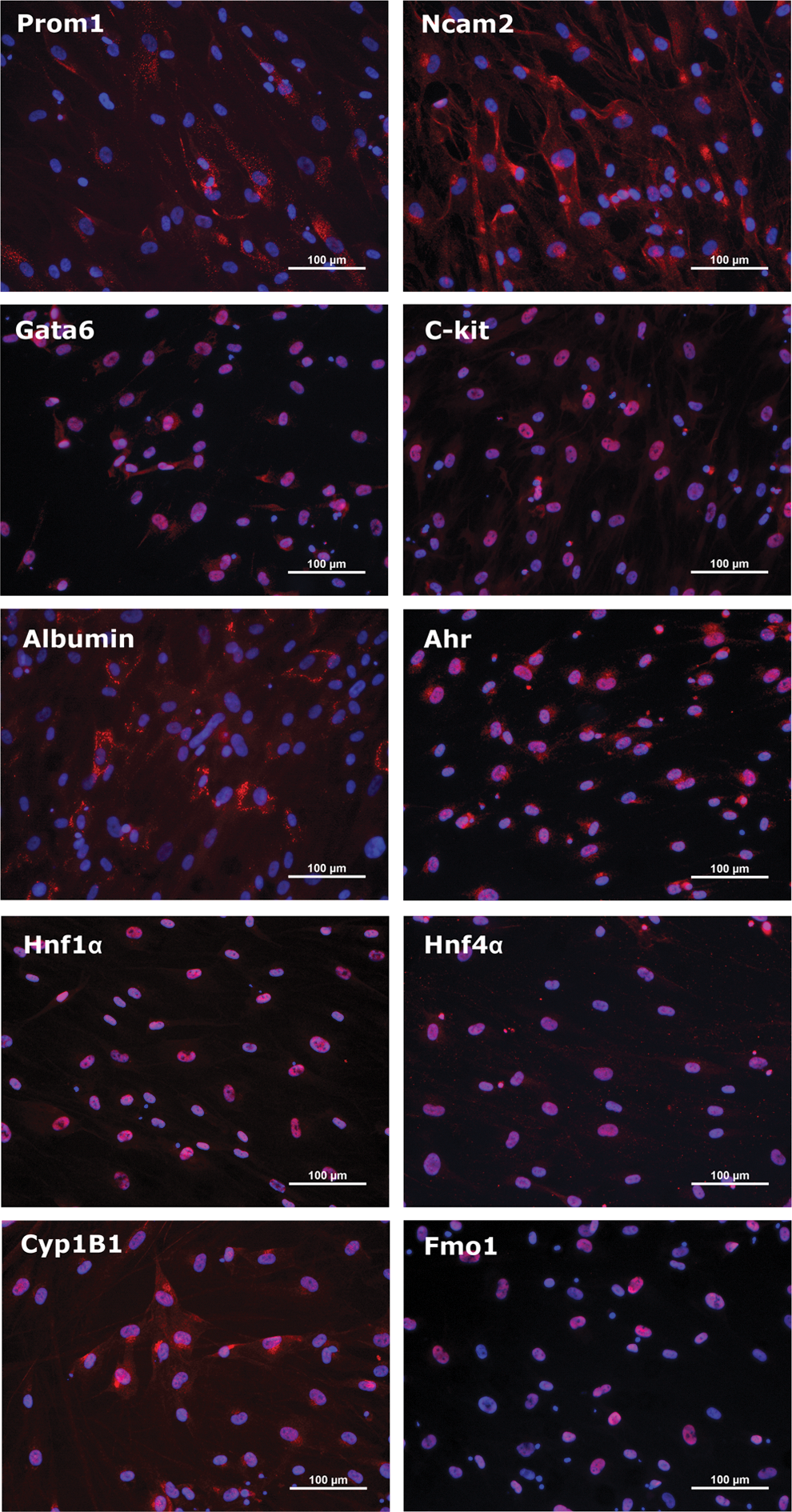
hSKP-HPC immunocytochemistry stainings of hepatic progenitor cell markers, as well as specific markers of mature hepatocytes. Color images available online at
Comparison between hSKP-HPC and hHEP with respect to the expression levels of typical hepatic markers shows that the major cytochrome P450 enzymes (CYP) are expressed at a lower level in hSKP-HPC than in hHEP (Fig. 1C). CYP1A1 and CYP1A2 reach 48% of the hHEP expression levels and CYP2D6, CYP3A4, CYP3A7 and CYP2E1 get to 23%, 16%, 13%, and 2%, respectively. Interestingly, the expression of CYP1B1 is a 100-fold higher in hSKP-HPC than in hHEP (Fig. 1C). The expression of the flavin-containing monooxygenase 1 (FMO1), which is a typical fetal liver phase I enzyme, is ten times higher in hSKP-HPC than in hHEP (Fig. 1D). FMO2 and FMO4, which are in general found in adult human liver, achieve levels comparable to those of hHEP (Fig. 1D). FMO3 expression on the contrary stays 10 times lower than in hHEP (Fig. 1D). The gene expression of monoamine oxidases A and B (MAOA, MAOB), another class of phase I oxidation enzymes, stays lower in hSKP-HPC than in hHEP (Fig. 1D). The phase II hepatic enzymes glutathione S-transferase GSTA4 and GSTM3 are found to be highly expressed in hSKP-HPC and reach levels that are, respectively, three and ninefold higher than those found in hHEP (Fig. 1E). Other phase II enzymes that is, GSTM1 and GSTK1 reach 52% and 72% of the expression levels of hHEP, respectively. UDP glucuronosyltransferase 1A (UGT1A) and GSTA1 expression is dramatically lower in hSKP-HPC than in hHEP (Fig. 1E). Typical phase 0 hepatic uptake drug transporters, including solute carrier family 10A1 (SLC10A1 or NTCP) and SLC22A1 (OCT1) are barely expressed at the gene level in hSKP-HPC. On the contrary, other influx transporters, as for instance SLC2A5 is up to 50-fold higher expressed in hSKP-HPC than in hHEP (Fig. 1F). Phase III efflux transporters, including ATP-binding cassette G2 (ABCG2 also referred to as BCRP), ABCC4 (MRP4), and ABCA1, known to be present in both adult and perinatal liver, are also significantly higher expressed in hSKP-HPC (Fig. 1F).
In general, when we evaluate the downstream effects of the modulated genes in hSKP-HPC, an enrichment of the “Gene Classes” related to “Development of Liver” is observed. The latter is composed of 116 known genes. Compared to undifferentiated hSKP, 56 of these are modulated in hSKP-HPC, out of which 37 are upregulated (Table 1). The Venn-diagram in Fig. 3 shows the liver development class genes that are upregulated in hSKP-HPC, hHEP and LIVER in comparison to undifferentiated hSKP. Eight genes are commonly upregulated in the three systems. GATA6 is 15-fold upregulated in hSKP-HPC. Other highly upregulated (>10-fold) genes are Toll-like receptor 4 (TLR4), suppressor of cytokine signaling 3 (SOCS3) and protein tyrosine phosphatase 12 (PTPN12). The top downregulated genes (>10-fold decrease) are SLC20A1 and vascular endothelial growth factor A (VEGFA).
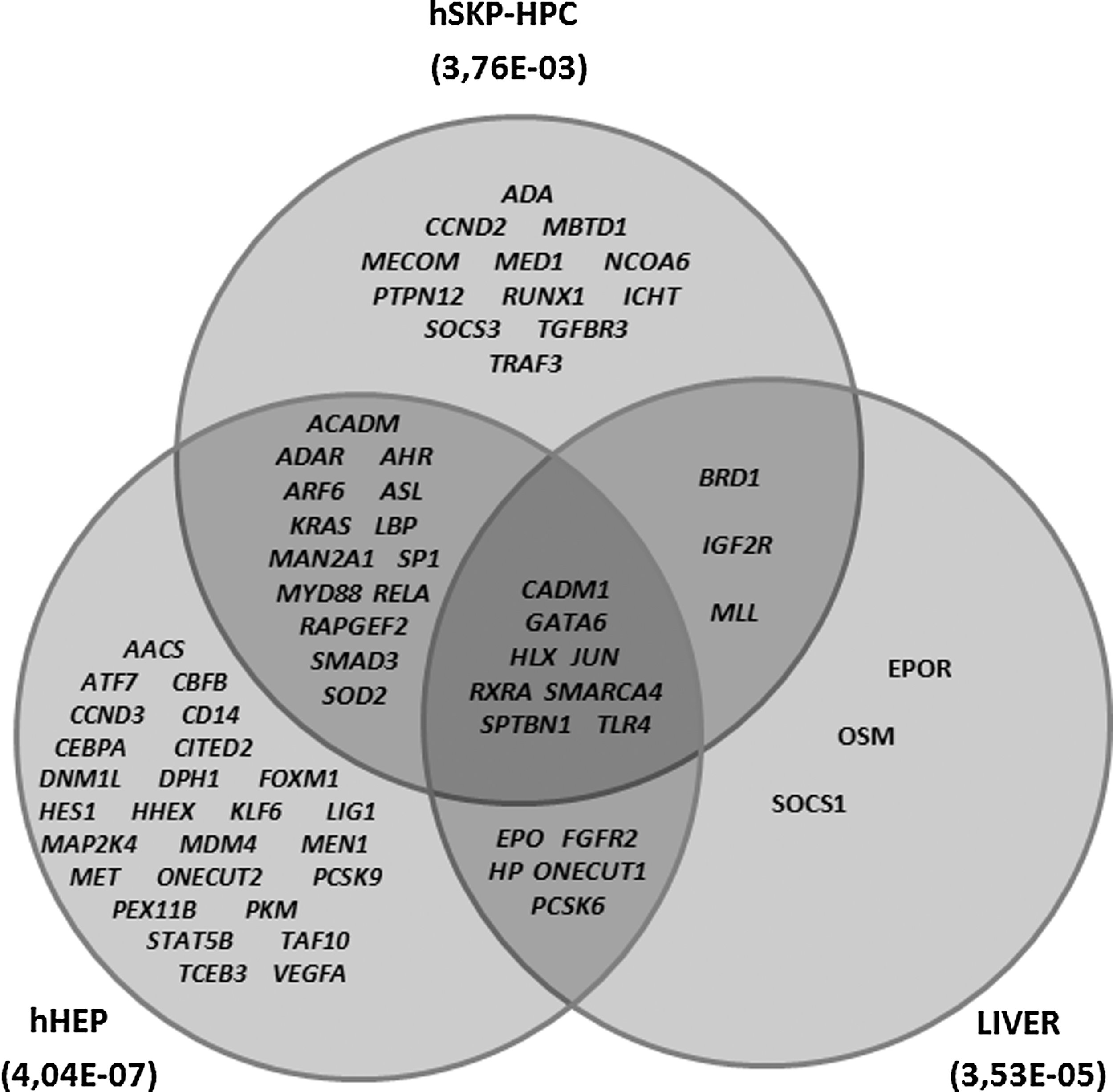
Venn diagram of the genes of the liver development class in hSKP-HPC, hHEP, and LIVER when compared to undifferentiated hSKP (Fisher's Exact P value is mentioned between brackets).
Genes expected to be upregulated for enrichment of Development of Liver Class.
Predictive capacity of hSKP-HPC for APAP-induced hepatotoxicity
By exposing hSKP-HPC to a range of APAP concentrations for 24 h and applying MTT viability tests, subcytotoxic concentrations of APAP could be determined. From a total of three tests an average IC10 value of 18 mM was calculated. The subcytotoxic concentration of APAP used in hHEP was 5 mM, which corresponds to the value previously described [42]. PCA of the microarray data shows highly reproducible results for the in vitro samples (Fig. 4). HSKP-HPC and hHEP exposed to IC10 of APAP [hSKP-derived hepatic progenitor cells exposed to acetaminophen (hSKP-HPC+APAP) and human hepatocyte cultures exposed to acetaminophen (hHEP+APAP)], as well as the respective control samples cluster individually. From the same PCA plot, it can also be noticed that APAP exposure results in a more pronounced shift of hSKP-HPC as compared to hHEP. Upon APAP exposure, a total of 5,591 genes is significantly (Student t-test with P value<0.05) modulated (minimal twofold) in hSKP-HPC, out of which 2,646 and 2,945 genes are up or downregulated, respectively. In hHEP, only 511 and 868 genes are significantly (Student t-test with P value<0.05) up and downregulated, respectively (minimal twofold). Functional analysis of the differentially expressed genes reveals the enrichment of gene classes of specific toxicological functions. As such, five liver-related “Toxicological Gene Classes” (Liver Damage, Liver Proliferation, Liver Necrosis/Cell Death, Liver Steatosis and Liver Hepatitis) could be identified with high accuracy (Fisher's Exact P value<0.05) in both hSKP-HPC and hHEP exposed to APAP (Table 2). The hSKP-HPC+APAP data could also identify two additionally relevant toxicological functions (Liver Steatohepatitis and Liver Edema). The percentage of modulated genes in each function was consistently higher in the hSKP-HPC+APAP samples than in the hHEP+APAP samples. The identified toxicological classes can be further divided in subfunctions. Though, these subfunctions do not always correlate among hSKP-HPC+APAP and hHEP+APAP (Table 3). More specifically, the Liver Damage gene class is divided in four subfunctions in hSKP-HPC+APAP and two different subfunctions in hHEP+APAP. The Liver Proliferation Class on the other hand has two common subfunctions in both cell types, but an extra three subfunctions in hHEP+APAP. Six subfunctions of the Liver Necrosis/Cell Death Class are enriched in hSKP-HPC+APAP and four in hHEP+APAP. Only one of these, however, is common to both cell systems. Four subfunctions of Liver Hepatitis are identified in hHEP+APAP out of which one is common to hHEP+APAP and hSKP-HPC+APAP. Liver Steatosis is composed of two subfunctions, of which each is identified by a different cell system. The number of modulated genes of each subfunction is consistently higher in hSKP-HPC+APAP compared to hHEP+APAP, resulting in higher subfunction enrichment in the former cell type. The gene lists of the different subfunctions of the toxicological classes have been summarized in the Supplementary Data (Supplementary Tables S2–S6). When looking at the gene level, it is found that a number of genes of the previously identified “Toxicological Classes” are commonly modulated in both hSKP-HPC and hHEP exposed to APAP. Five genes, including Bcl-2 interacting mediator of cell death (BCL2L11), cellular oncogene c-fos (FOS), heme oxygenase 1 (HMOX1), TIMP metallopeptidase inhibitor 3 (TIMP3) and AHR, are significantly upregulated (fold change>2, Fisher's Exact P value<0.05) and three genes, including insulin-like growth factor 1 (IGF1), regucalcin (RGN) and inhibin beta A (INHBA) are significantly downregulated (fold change>2, Fisher's Exact P value<0.05) (Fig. 5). These results were confirmed by qPCR analysis for both hSKP-HPC and hSKP-HPC+APAP samples (Supplementary Fig. S1), validating the obtained microarray data. The upregulation of AHR could not be observed in a significant way.

PCA-plots of the microarray data of hSKP-HPC and hHEP exposed to acetyl-para-aminophenol, acetaminophen (APAP) [respectively hSKP-derived hepatic progenitor cells exposed to acetaminophen (hSKP-HPC+APAP) and human hepatocyte cultures exposed to acetaminophen (hHEP+APAP) samples] and their respective control samples (hSKP-HPC and hHEP).
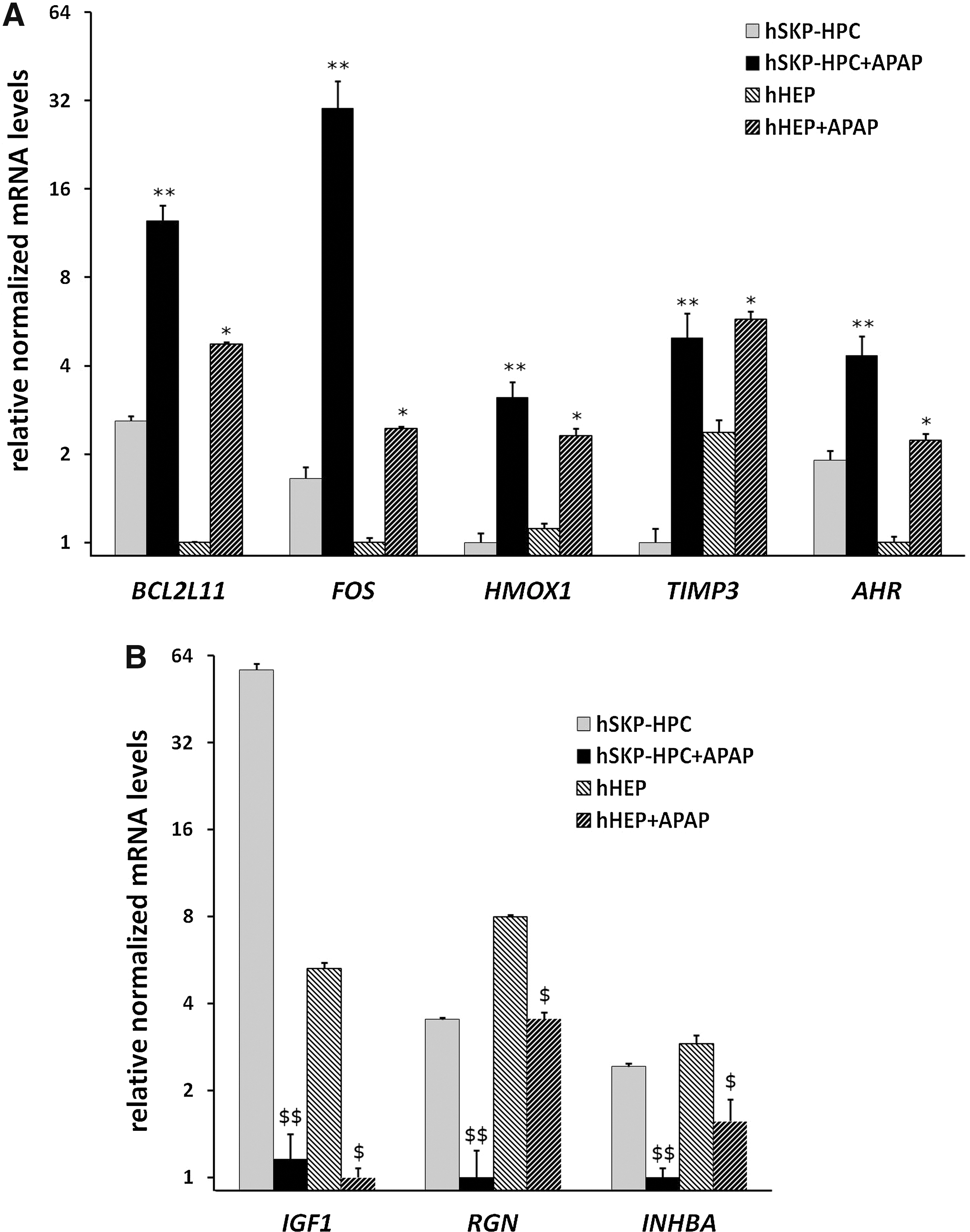
Microarray expression of Toxicological Classes genes that are commonly modulated in hSKP-HPC and hHEP treated with APAP.
APAP, acetyl-para-aminophenol, acetaminophen; hHEP+APAP, human hepatocyte cultures exposed to acetaminophen; hSKP-HPC+APAP, hSKP-derived hepatic progenitor cells exposed to acetaminophen.
Discussion
DILI is the main cause of ALF in the western world [3,44]. It is also one of the main reasons for post marketing withdrawal of drugs and discontinuation of the development of NCE, thereby representing a substantial cost for the pharmaceutical industry [3,44]. Current human cell models are scarce or not fully adequate for toxicity screening of pharmaceuticals and other chemical compounds [16,45]. Therefore, novel cell models need to be explored. The ability of ASC to self-renew and to differentiate into cell types of mesodermal, endodermal, or ectodermal origin makes them attractive as therapeutic agents to replace cell loss due to injury or disease [46]. In addition, ASC represent a potential new cell source for toxicity screening and the use of human ASC in this field could have a pivotal effect on the efficacy and predictability of in vitro screening methods [47].
In this study, we focused on hSKP and more specifically on their stepwise-differentiated hepatic progeny, as a potential predictive human-based cell system for hepatotoxicity screening of drugs. As illustrated in the results' section, the expression of known hepatic progenitor cell markers was upregulated upon sequential exposure to liver-specific hepatogenic growth factors and cytokines. These markers included EPCAM, a surface marker of both human hepatic progenitor cells [48] and newly-derived hepatocytes [49]; NCAM2, known as a liver progenitor cell marker and also expressed in acute and chronic damaged livers [50]; PROM1 and KIT, both having been reported as markers for liver progenitor cells [51]. Markers, such as SMAD4 and THY1 have also been shown to be expressed in liver progenitor cells [51 –54]. TLR4 is known to be involved in liver fibrogenesis [55] and SOCS3 in hematopoiesis in fetal liver [56]. Both GATA6 and PTPN12 are essential during liver development [57,58]. The expression of all these specific hepatic progenitor cell markers, show that hSKP have acquired hepatic progenitor cell-like properties after in vitro differentiation. hSKP-HPC also expressed, both at the protein and gene expression level, more mature hepatic markers, including albumine, Hnf1α, Hnf4α, and Ahr. Recently, the latter has been reported to play an important role in hepatocytes responding to a toxic assault [59]. Interestingly, hSKP-HPC also seems to have acquired a metabolic machinery. An increased expression at the gene level is observed for several phase I and phase II biotransformation enzymes, as well as for influx and efflux drug transporters. Indeed, important phase I metabolizing enzymes, such as FMO1, FMO2, FMO3 and FMO4 and MAOB are highly expressed in hSKP-HPC. The gene expression of the phase II hepatic enzymes GSTA4 and GSTM3 was also increased in the differentiated cells. The drug transporters that were found to be highly expressed in hSKP-HPC are among others the liver basolateral transporters ABCC4 and ABCA1, as well as the apical transporter ABCG2. They are all known to be present during liver development [60]. Although the expression of some liver specific influx transporters, including SLC22A1 and SLC10A1, was not increased in hSKP-HPC, several other influx transporters, such as SLC2A5 [61] were highly upregulated. It seems, therefore, reasonable to assume that the expression of this metabolizing capacity at the gene level assigns hSKP-HPC adequate cellular and metabolic competences to ultimately serve as a potential novel human-based in vitro cell system useful for screening out liver toxic compounds. Here, we evaluated the toxicological response of hSKP-HPC after exposure to APAP, a hepatotoxic substance accountable for most cases of drug-induced ALF, due to its known hepatotoxicity [62]. Full genome transcriptomics was chosen as the main holistic method to understand hepatotoxic responses. This technology has proven to generate high quality, robust and reproducible data [63]. Because of its high sensitivity, evaluation of early toxicological responses is possible. Indeed, several in vivo transcriptomics studies have reported the predictive character of this technology to detect hepatotoxicity [64 –69] in human cell cultures of both primary hepatocytes and hepatic cell lines [64,70,71]. As mentioned before, these cell models have serious drawbacks regarding availability and functionality, respectively.
In this study, functional analysis of the differentially expressed genes elucidates the enrichment of gene classes that are involved in specific toxicological functions. Strikingly, the same hepatotoxic responses could be identified in both hSKP-HPC and hHEP exposed to APAP. Further evidence is found by the observation that a number of genes of the identified “Toxicological Classes” is commonly modulated in both hSKP-HPC and hHEP exposed to APAP. The upregulated genes, BCL2L11, FOS, HMOX1, TIMP3 and AHR are all associated either to cytotoxic responses or induction of apoptosis [72 –77] and might be useful molecular biomarkers for hepatotoxicity.
As such, this data gives a first indication that human stem cell-derived hepatic cells could be valuable tools in the early in vitro prediction of hepatotoxicity. In addition, it was also found that the percentages of the genes involved in each APAP-triggered “Toxicological Class” are consistently higher in hSKP-HPC than in hHEP. A possible explanation could be the immature status of hSKP-HPC, suggesting an alternative toxicological mode of action in comparison to adult human hepatocytes. This is predominantly evoked by a low expression of CYP2E1 in hSKP-HPC, which is known to be one of the main enzymes involved in the bioactivation of APAP by postnatal parenchymal liver cells. On the other hand, APAP is also known to induce fetal hepatotoxicity [78]. Yet, the presence and activity of CYP2E1 in developing liver remains a topic of debate [79]. Furthermore, it is known that the expression of adult CYP isoforms, including CYP1A2, CYP12C, CYP12D6, and CYP3A4, mainly occurs after birth [80]. This leads to important differences in metabolic capacity between embryonic/fetal liver and adult liver, which could suggest that APAP exhibits its hepatotoxicity through an alternative mode of action within developing hepatocytes. Yet, no specific in vitro system is currently available that is able to evaluate human hepatotoxicity of compounds during the ontogeny of parenchymal liver cells. As such, due to their similar characteristics with in vivo developing hepatocytes, hSKP-HPC could be a particularly interesting human screening model with focus on for developmental hepatotoxicity.
Footnotes
Acknowledgments
The authors would like to thank Ms. Margit Henry and Ms. Susan Rohani for their excellent technical assistance.
This research has received funding from grants of the Research Council (OZR) of the Vrije Universiteit Brussel, the Fund for Scientific Research in Flanders (FWO-Vlaanderen, GO16312N), the European Community's Seventh Framework Programme (FP7/2007–2013) under grant agreement no. 20161 (ESNATS), no. 266838 (DETECTIVE), no. 266777 (HEMIBIO), and from ISRIB (Brustem) and BELSPO (IAP-HEPRO).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.