
Abstract
Bone marrow-derived mesenchymal stem cells (BM-MSCs) play an important role in Helicobacter pylori-induced gastric carcinogenesis. While the mechanism is not well understood, BM-MSCs have been shown to contribute to the immunosuppressive response found in a number of diseases. Here, BM-MSCs were transplanted into the stomach of mice with a 44-week mouse-adapted H. pylori infection. At day 28 post-transplantation, BM-MSCs migrated from the subserosal to the mucosal layer of the stomach. The grafted BM-MSCs significantly stimulated systemic and local interleukin-10 (IL-10)-secreting T cell and regulatory T cell (Treg) functions. This observation was correlated with an increased percentage of CD4+IL-10+ cells and CD4+CD25+FoxP3+ cells in splenic mononuclear cells compared with H. pylori-infected mice not receiving BM-MSCs. Moreover, inhibitory cytokines IL-10 and transforming growth factor-β1 increased in the gastric tissue, while there was a decrease in inflammatory interferon-γ (IFN-γ). BM-MSC-transplanted mice also developed elevated IL-10/IFN-γ secreting and Treg/Th17 ratios. A coculture system in the presence or absence of BM-MSCs was also established to evaluate the immune responses in vitro. An increase in IL-10-secreting T cells and Tregs, associated with increased expression of Gata-3 and FoxP3, generation of IL-10 in the supernatant, and proliferation of gastric epithelial cells (GECs) was observed. These findings demonstrate that transplantation of BM-MSCs into a chronic H. pylori-infected mouse model results in the generation of an immunosuppressive environment. The local and systemic immunosuppression mediated by BM-MSCs likely contributed to an environment that is compatible with the development of H. pylori-induced gastric cancer.
Introduction
The persistence of chronic inflammation plays a pivotal role in almost all inflammation induced-tumor progression [3,4]. Thus, a complete understanding of how immune cells interact with the tumor microenvironment is necessary if we hope to understand how H. pylori induce gastric carcinogenesis. Importantly, H. pylori is known to modulate both the mucosal and systemic immune response, as evidenced by the local secretion of cytokines and the recruitment of distinct inflammatory cells, respectively [5,6]. Previous studies have demonstrated that T cell responses that are critical to the pathogenesis of H. pylori infection are present in different temporal patterns during disease progression. In the early stages of infection, H. pylori is known to induce a mixed T helper 1 (Th1)/Th17 cell response characterized by high expression of interferon-γ (IFN-γ) and IL-17 [7,8], while in advanced disease, especially in gastric carcinogenesis, the immune responses have been shown to be predominantly Th2/regulatory T cell (Treg) [9 –11]. By altering the DC-polarized Th17/Treg balance, Kao et al. have demonstrated that H. pylori induce a Treg-biased response that favors their immune escape. This skewing of the immune response equilibrium has been found in a variety of diseases and has been shown to be a key factor in antitumor immunity [12,13].
In recent years, there is increasing evidence in support of a role for bone marrow-derived cells (BMDC) in the progression of gastric carcinogenesis. Varon et al. have demonstrated that BMDCs are recruited to the gastric glands of H. pylori-infected mice and that the presence of BMDCs is positively correlated with the development of pseudointestinal neoplasia [14]. Houghton et al. demonstrated that chronic inflammation induced by H. pylori infection is required for appreciable bone marrow-derived mesenchymal stem cells (BM-MSCs) engraftment in the stomach and subsequent carcinogenesis; acute injuries by chemical treatment and transient parietal cell loss could not induce BM-MSC recruitment [15]. In addition, a recent in vitro study demonstrated that BM-MSC recruitment can be induced by H. pylori-infected gastrointestinal epithelial cells. This recruitment was found to require the secretion of multiple cytokines via an NF-κB-dependent pathway [16]. Taken together, these data suggest that there are interactions between MSCs and H. pylori-induced gastric cancer. However, information remains limited regarding the specific contribution of BM-MSC to gastric inflammation and carcinogenesis.
How might BM-MSC contribute to the development of H. pylori-induced gastric carcinogenesis? One potential mechanism may be in altering the local immune responses in the tumor microenvironment. BM-MSCs have attracted considerable attention for their potential use in the clinical therapy of many diseases because, in addition to their multipotent differentiation properties and low immunogenicity, they have a proven ability to modulate immune function [17,18]. BM-MSCs have been shown to significantly inhibit naïve and memory T cell proliferation, and to block the cytotoxicity and inflammatory cytokine production associated with the mixed lymphocyte reaction [19,20]. Other studies have demonstrated that BM-MSCs can successfully improve solid organ transplantation by downregulating immune effector function, such as by suppressing Th1 and Th17 activity and increasing the generation of Tregs [21 –23]. Parekkadan et al. have also observed that BM-MSC transplantation as a cellular therapy ameliorates autoimmune enteropathy in a multiorgan autoimmunity model by increasing CD4+CD8+ thymocyte production and serum interleukin-10 (IL-10) and decreasing serum IFN-γ [24]. Moreover, numerous studies have shown that, as immunosuppressive agents, BM-MSCs can promote a variety of types of tumor growth, including breast cancer, Barrett's adenocarcinoma of esophagus, and lung adenocarcinoma, among others [8,25 –27]. It may be that, through inappropriate cell–cell contacts in the persistent inflammatory/injury environment, BM-MSCs exert immunomodulatory effects that enhance the ability of tumors to evade immune surveillance.
To better understand the specific molecular and cellular mechanisms involved in the immunomodulatory activities of BM-MSCs in H. pylori-induced gastric carcinogenesis, it is necessary to elucidate the systemic and local immune responses in this abnormal inflammatory environment. In the current study, we investigated the immune response following BM-MSC transplantation in a mouse model of chronic H. pylori infection. We demonstrate for the first time that BM-MSC can function as an anti-inflammatory component of the immune system in an H. pylori-induced gastric cancer microenvironment.
Materials and Methods
Culture of mouse-adapted H. pylori
The H. pylori strain TN2GF4 (kindly provided by Jianzhong Zhang, Chinese Center for Disease Control and Prevention of Infectious Diseases, Beijing, China) was used in this study. H. pylori TN2GF4 was cultured at 37°C under microaerophilic conditions on brain heart infusion agar plates (Oxoid) supplemented with (10% v/v) sheep blood and a standard Campylobacter selective supplement (Oxoid).
To establish a BALB/c mouse-adapted H. pylori strain of TN2GF4, H. pylori TN2GF4 were inoculated into BALB/c mice [28]. After being infected for 4 weeks, mice were sacrificed and the gastric mucosal tissue was collected for H. pylori microaerophilic culture. The first generation of BALB/c mouse-adapted H. pylori strain TN2GF4 was called TN2GF4 BS1. Using the same method, TN2GF4 BS1 was inoculated into BALB/c mice and acclimated through serial passages in BALB/c mice for procuring a better colonizing H. pylori strain. In this study, the fifth generation of BALB/c mouse-adapted H. pylori strain (TN2GF4 BS5) was used.
For coculture experiments, BALB/c mouse-adapted H. pylori strain TN2GF4 BS5 was cultured in liquid media containing 5% fetal bovine serum (FBS; Gibco) for 2–3 days, centrifuged, resuspended in culture medium, and then used immediately. H. pylori concentration was estimated by measuring the OD600nm, where OD600nm corresponds to ∼2×108 CFU/mL. H. pylori were added to gastric epithelial cells (GECs) at a multiplicity of infection (MOI) of 10.
Preparation and characterization of BM-MSCs and GECs
Bone marrow cells were obtained by lavage of the femurs of 4-week-old male BALB/c mice. Cells were suspended in low glucose Dulbecco's modified Eagle's medium (DMEM; Gibco), supplemented with 10% FBS, and cultured in a humidified atmosphere containing 5% CO2 at 37°C. Nonadherent cells were removed after 48 h and the culture media was replaced every 3 days. After 2 weeks, 70% to 80% confluent cells were dissociated with 0.25% trypsin (Sigma) and passaged. BM-MSCs at passage 6 were identified by staining with antibodies to CD11b, CD45, CD90, CD105, and stem cell antigen (Sca)-1 (eBioscience). GECs were obtained from the body region of the stomach using BALB/c mice. The gastric body area was excised and rinsed thrice, cut into explants, and dissociated using collagenase (Sigma) at 37°C in a shaking water bath for 30–60 min depending on the amount of tissue dissociation required. At the end of this incubation, large explants were removed and the cell suspension was centrifuged. Cells were suspended in DMEM with 10% FBS, penicillin (100 U/mL), and streptomycin (50 μg/mL) and cultured in a tissue culture incubator. Nonadherent material was discarded after 48 h and the culture media was replaced every 3 days.
Establishment of mouse model of chronic H. pylori infection
Pathogen-free female BALB/c mice (4–6 weeks old) were purchased from the Experimental Animal Center of Tongji Medical College, Huazhong University of Science and Technology (Wuhan, China) and housed under standard laboratory conditions. All protocols regarding animal care and experimental procedures were reviewed and approved by the Animal Care and Use Committee at Huazhong University of Science and Technology. To induce H. pylori chronic inflammation, 0.1 mL of BALB/c mouse-adapted strain of H. pylori TN2GF4 BS5 suspension (∼1–2×109 CFU/mL) was orogastrically inoculated thrice over a 5-day period using a mouse gavage needle. Control mice were mock-inoculated with H. pylori liquid culture medium.
BALB/c mice that were successfully infected with mouse-adapted H. pylori strain TN2GF4 BS5 for 10 months were subserosally tranplanted with male-donor BM-MSCs (F6, 1×106 cells/mouse) into the antrum area of the greater curvature. Control groups were given cell culture medium alone. Thus, experimental mice were randomly divided into four groups: cell culture medium transplantation alone group (SHAM, n=25), BM-MSC transplantation alone group (MSC, n=25), cell culture medium transplantation with chronic H. pylori infection group (Hp, n=25), and BM-MSC transplantation with chronic H. pylori infection group (Hp+MSC, n=25). To track the transplanted BM-MSCs in vivo, a small part of the mice in each group were transplanted BM-MSCs prelabeled the bis-benzimide (Hoechst 33258; Sigma). Fifty-six days after BM-MSC transplantation, spleen samples and various segments of the stomach were collected. Tissue segments were immediately frozen in a −80°C freezer for later use in histologic studies. Protein extraction and cytokine determination were performed as described elsewhere in the Materials and Methods section.
Isolation of splenic mononuclear cells
Splenic mononuclear cells (MNC) were obtained by Percoll-Paque gradient centrifugation (specific gravity 1.077 g/mL; GE Healthcare) under aseptic conditions. Erythrocytes were removed by hypotoniclysis and the remaining cells were cultured in RPMI-1640 medium supplemented with 10% FBS. Cell viability was evaluated by using 0.1% trypan blue. To measure the immunomodulatory activity of BM-MSC transplantation in vivo, splenic MNCs were isolated, stained, and analyzed by flow cytometry analysis.
In vitro culture system
To evaluate the role of BM-MSC on immunomodulation of T cells and proliferation of GECs, a series of in vitro cell coculture experiments were performed using a transwell system (pore size 3 μm; Corning, Inc.) [16,29,30]. Briefly, 1×104 mouse GECs were seeded on the upper chamber of cell culture inserts and 1×105 BM-MSCs were separately seeded into the lower culture plates. After overnight incubation, the ultrasonic crushing of H. pylori was then added to the upper chamber at an MOI of 10, 1×106 naïve splenic MNCs were added to the lower culture plates, and the cells were cocultured. Thus, experimental MNCs were divided into 4 groups: MNC+GEC group (SHAM), MNC+GEC+BM-MSC group (MSC), MNC+GEC+ultrasonic crushing of H. pylori group (Hp), and MNC+GEC+ultrasonic crushing of H. pylori+BM-MSC group (Hp+MSC). In some experiments, GECs were prelabeled with 2 μM 5,6 carboxyfluorescein diacetate succinimidyl ester (CFSE; Invitrogen) for 10 min at 37°C in the dark and washed twice in RPMI 1640 supplemented with 10% FBS before seeding and later coculture. Twenty-four hours past coculture, splenic MNCs, GECs, and supernatants from the lower chamber were collected for further analysis.
Flow cytometric analysis
BM-MSCs were positively identified by staining 100 μL aliquots of cells with fluorescein isothiocyanate (FITC)-conjugated rat anti-mouse CD11b, CD45, phycoerythrin (PE)-conjugated rat anti-mouse CD90, CD105, and Sca-1 (eBioscience) at 4°C for 30 min. Fluorescence intensity of positively stained samples was compared with cells that had been either left unstained or stained with an isotype control antibody. Cells were then washed with phosphate-buffered saline (PBS) containing 1% FBS and resuspended in flow cytometry (FACS) buffer (PBS supplemented with 1% FBS and containing 0.05% NaN3). At least 5×105 events were analyzed by flow cytometry (FACScan; Becton Dickinson).
Splenic MNCs were harvested and restimulated for another 6 h with H. pylori sonicate in the presence of Brefeldin A. Cells were then stained at a concentration of 5×105 cells per 100 μL of PBS with 1% FBS. One set of aliquots was labeled with FITC-conjugated anti-CD4 mAb combined with PE-conjugated anti-CD25 mAb (eBioscience). A second set of aliquots was labeled with FITC-conjugated anti-CD4 mAb, fixed, permeabilized, and stained with PE-conjugated anti-IFN-γ, peridinin chlorophyll protein (PerCP)-Cy5.5-conjugated anti-IL-10 and allophycocyanine (APC)-conjugated anti-IL-17 mAb (eBioscience) at room temperature in the dark for 30 min. For labeled CD4+CD25+ cells, cells were permeabilized and stained with APC-conjugated anti-Foxp3 mAbs (eBioscience). At least 5×105 events in the lymphocyte gate were analyzed. All mAbs were used at concentrations recommended by the manufacturer.
After 24 h of coculture in vitro, CFSE-labeled GECs were collected and washed in 0.1% BSA in PBS, and assessed CFSE fluorescence at 488 nm by flow cytometry. The dye dilution occurred with cell divisions, and cell proliferation could be easily analyzed by separating dot plots with CFSE fluorescence intensity.
Real-time quantitative polymerase chain reaction
Messenger RNA (mRNA) expression analysis from gastric tissue and splenic MNCs was performed by reverse transcriptase–polymerase chain reaction (RT-PCR). Total RNA was extracted using TRIzol reagent (Invitrogen) and reverse transcribed to complementary DNA using Multiscribe reverse transcriptase (Applied Biosystems). Real-time quantitative PCR was performed using the SuperScript® III One-Step RT-PCR System with Platinum® Taq (Invitrogen) and a StepOneTM Real-Time PCR system (Applied Biosystems). For quantification, cytokine mRNA copy numbers were normalized to the housekeeping gene β-actin. Relative mRNA expression was measured using the 2-ΔΔCt method [31]. All gene primer sequences are shown in Table 1.
IFN-γ, interferon-γ; IL-10, interleukin-10; IL-17, interleukin-17; TGF-β1, transforming growth factor-β1.
Immunofluorescence, enzyme-linked immunosorbent assay, western blot analysis, and MTT assay
Immunofluorescence, enzyme-linked immunosorbent assay (ELISA), western blot analysis, and MTT assay were performed as described previously [32,33]. Briefly, immunofluorescence studies were performed by growing epithelial cells on glass cover slips. Cells were fixed with 4% paraformaledehyde, blocked using PBS with 10% donkey serum, and incubated with antibodies against pancytokeratin (C11) (Santa Cruz), followed by the indicated secondary antibody. Nuclei were stained with Hoechst 33258 and imaged using a IX70 Olympus microscope. ELISA was performed using ELISA kits for murine IFN-γ and IL-10 according to the manufacturer's instructions (eBioscience). Antibodies used for western blotting included anti-IFN-γ, anti-IL-10, anti-IL-17 anti-transforming growth factor-β1 (TGF-β1), and anti-β-actin (all from R&D System). The imaging analysis was carried out by an automated electrophoresis gel imaging system, and quantitative expression of proteins was calculated as the ratio of integrated optical density of target protein to that of β-actin. The IL-10/IFN-γ and Treg/Th17 ratios of the same specimen in stimulation of T cell proliferation, cytokine production, and protein expression levels were calculated [32,34,35]. MTT (3-(4, 5-dimethylthiazol-z-yl)-2, 5-diphenyltetrazolium bromide) assay were performed to assess the proliferation of GECs at 6, 12, 24, and 48 h. After coculture in vitro, GECs were incubated with MTT reagent (5 mg/mL; Sigma) for 4 h at 37°C before solubilization in dimethyl sulfoxide (DMSO; Sigma). The formazan was quantified by detecting absorbance at 570 nm (630 nm as a reference).
Statistical analysis
Statistical analysis was performed using either a one-way ANOVA or the nonparametric Dunnet's T3 test depending on the dataset. The data were presented as mean±SEM. A P value of less than 0.05 indicated statistical significance.
Results
Generation of highly homogenous CD105/Sca-1-positive and CD11b/CD45/CD90-negative BM-MSCs
Bone marrow-derived adherent cells were isolated from BALB/c mice and cultured as described in the Methods section. After passage 6, the purified cells exhibited the spindle-shaped and well-spread morphology that is characteristic of these cells. Flow cytometry analysis confirmed that the cells at passage 6 were a group of highly homogenous cells, positive for CD105 and Sca-1 surface markers and negative for the hematopoietic markers CD11b, CD45, and the monocytic marker CD90, the molecules attributed to BALB/c murine BM-MSC (Fig. 1A, B) [36,37].
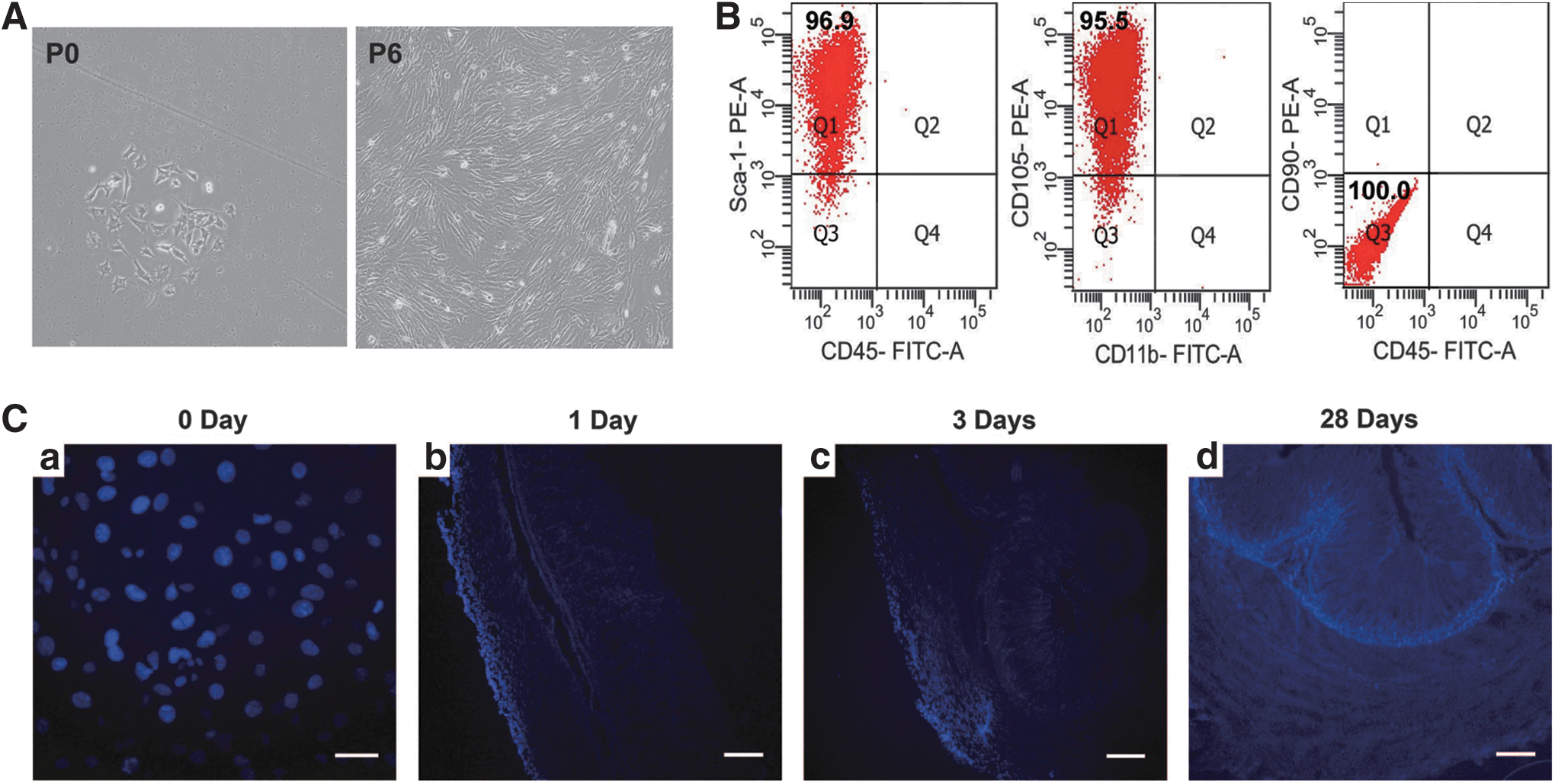
BM-MSCs primarily migrate to the mucosa in mice with chronic Helicobacter pylori infection 28 days after transplantation.
MSCs transplanted into H. pylori-infected mice are primarily localized to the mucosa of the gastric wall 28 days after transplantation
Allogenic BM-MSCs, labeled with Hoechst 33258, were subserosally transplanted into the gastric wall of mice that had a 44-week mouse-adapted H. pylori TN2GF4 infection. At 1, 3, and 28 days post-transplantation, the pyloric wall was dissected, cut into serial sections, and analyzed by a blinded observer. At 1 day post-transplantation, grafted BM-MSCs were localized to the serosa and subserosa. At 3 days post-transplantation, the BM-MSCs had predominantly migrated to the longitudinal/circular muscle layer. At day 28, the BM-MSCs had survived and predominantly migrated to the mucosa (Fig. 1C). To ensure these findings, 56 days after BM-MSC transplantation, fluoresence in situ hybridization (FISH) was performed to detect Y chromosome in the mouse stomach. In BM-MSC transplantation mouse with chronic H. pylori infection (Hp+MSC), quantification of the transplanted epithelial area containing 22% of Y-positive GECs was performed. However, Y chromosome fluorescein signal in the nuclei was not stained in cells from mouse that was transplanted BM-MSCs without chronic H. pylori infection (BM-MSC transplantation alone, MSC; Supplementary Fig. S1; Supplementary Data are available online at
BM-MSC transplantation upregulates systemic anti-inflammatory cytokine expression and induces Treg responses in mice chronically infected with H. pylori
To test the role for BM-MSC in regulation of the immune response in vivo, splenic MNCs were obtained from mice that had been infected with H. pylori for 52 weeks. Some of these mice had been transplanted with BM-MSCs for a period of 8 weeks while a control group did not. Chronic H. pylori infection systemically increased the percentage of IL-10-secreting CD4+ T cells (3.878±0.312 for Hp vs. 1.117±0.093 for SHAM, P<0.01), which could be further upregulated by 8 weeks of BM-MSC transplantation (5.614±0.530 for Hp+MSC vs. 3.878±0.312 for Hp, P<0.01), shown as shown in Fig. 2A and B. The percentage of CD4+ IFN-γ+ T cells was also stimulated in the splenic MNCs of mice that were chronically infected with H. pylori (14.629±0.425 for Hp vs. 5.933±0.96 for SHAM, P<0.001, Fig. 2A, B). However, there was no significant difference in CD4+ IFN-γ+ T cell percentage in mice with a chronic H. pylori infection that did not receive BM-MSC transplantation, indicating that MSC transplantation heightens the anti-inflammatory response.
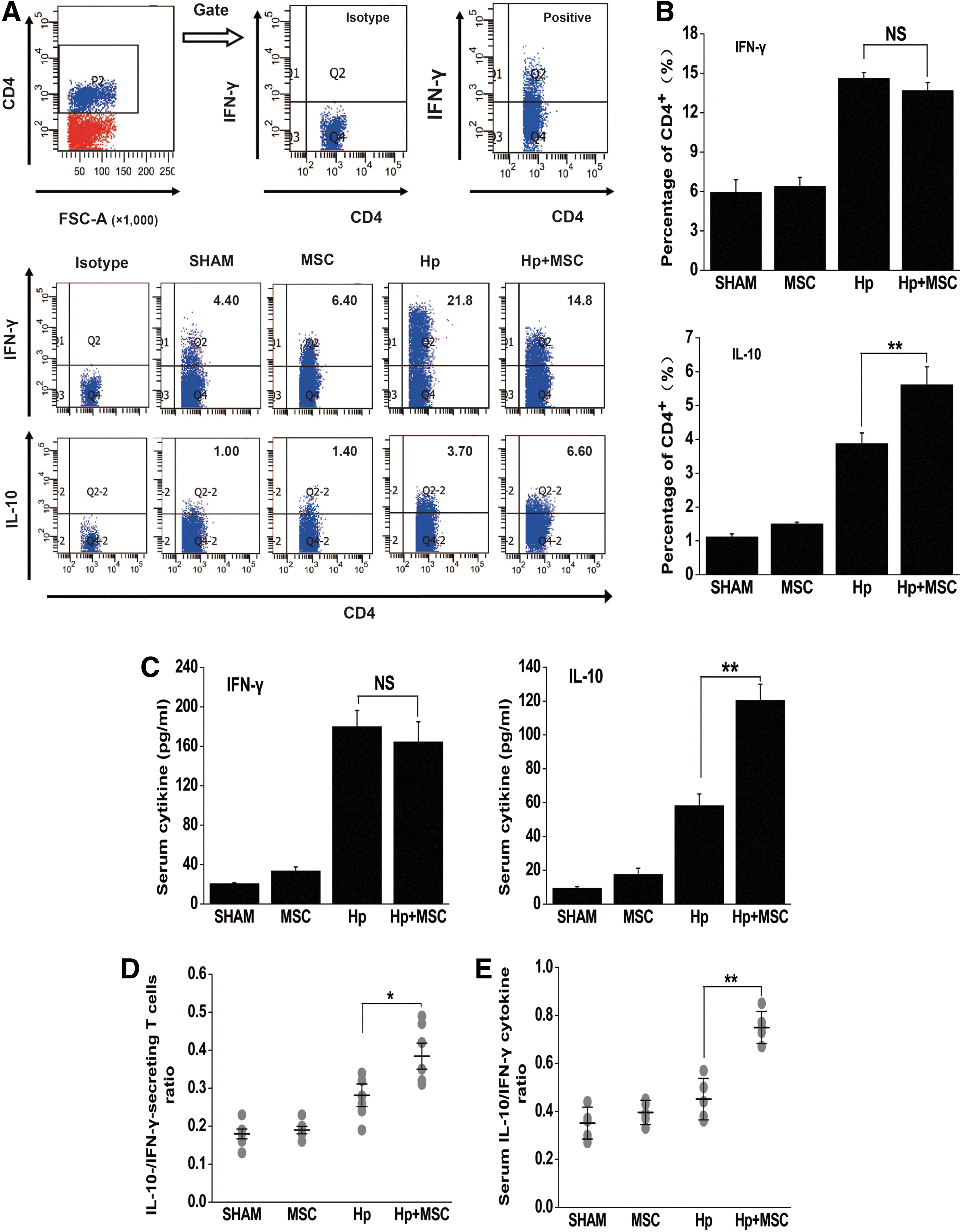
BM-MSC transplantation systemically upregulates IL-10-secreting T cell responses in mice with chronic H. pylori infection.
The presence of the Th1-type cytokine IFN-γ and the anti-inflammatory cytokine IL-10 in the serum was evaluated by ELISA. Levels of IL-10, which have been mechanistically linked to H. pylori-induced gastric cancer, were found to be significantly increased in Hp+MSC group mice (120.44±9.58 pg/mL) compared with Hp group mice (58.20±6.899 pg/mL), (P<0.01, Fig. 2C). The expression of IFN-γ in the serum did not change in response to BM-MSC transplantation (Fig. 2C).
To further analyze the cytokine balance in this experimental mouse model, we assessed the ratio of IL-10/IFN-γ-secreting-T cells. In nuclear cells, the CD4+ IL-10+ T cells/CD4+ IFN-γ+ T cell ratio was markedly elevated in Hp+MSC mice in comparison with H. pylori group mice (0.374±0.034 for Hp+MSC vs. 0.281±0.030 for Hp, P<0.05; Fig. 2D). Similarly, the IL-10/IFN-γ ratio was found to be higher in Hp+MSC mice than in Hp group mice (0.749±0.299 for Hp+MSC vs. 0.451±0.038 for Hp, P<0.01; Fig. 2E). These data suggest that BM-MSC transplantation skews the T cell response in vivo to an immunosuppressive IL-10-secreting response. This shift in immune response may contribute to gastric cancer progression.
Because elevated levels of Th17 and Treg cells have been found in some tumor tissue and the mucosal microenvironment [38 –41], we measured the percentage of CD4+IL-17+ helper T cells and CD4+CD25+Foxp3+ Tregs in the spleen. We found that chronic H. pylori infection significantly increased the percentage of CD4+IL-17+ helper T cells and CD4+CD25+Foxp3+ Tregs in the spleen (P<0.05, Fig. 3A). Further, BM-MSC transplantation induced Treg production significantly more than chronic H. pylori infection alone (10.309±1.108 for Hp+MSC vs. 5.744±0.22 for Hp, P<0.001, Fig. 3B). No significant differences were seen in the percentages of Th17 cells (Fig. 3A, B). Thus, 8-week BM-MSC transplantation was found to dramatically elevate the Treg/Th17 ratio in mice that are chronically infected with H. pylori (Fig. 3C).
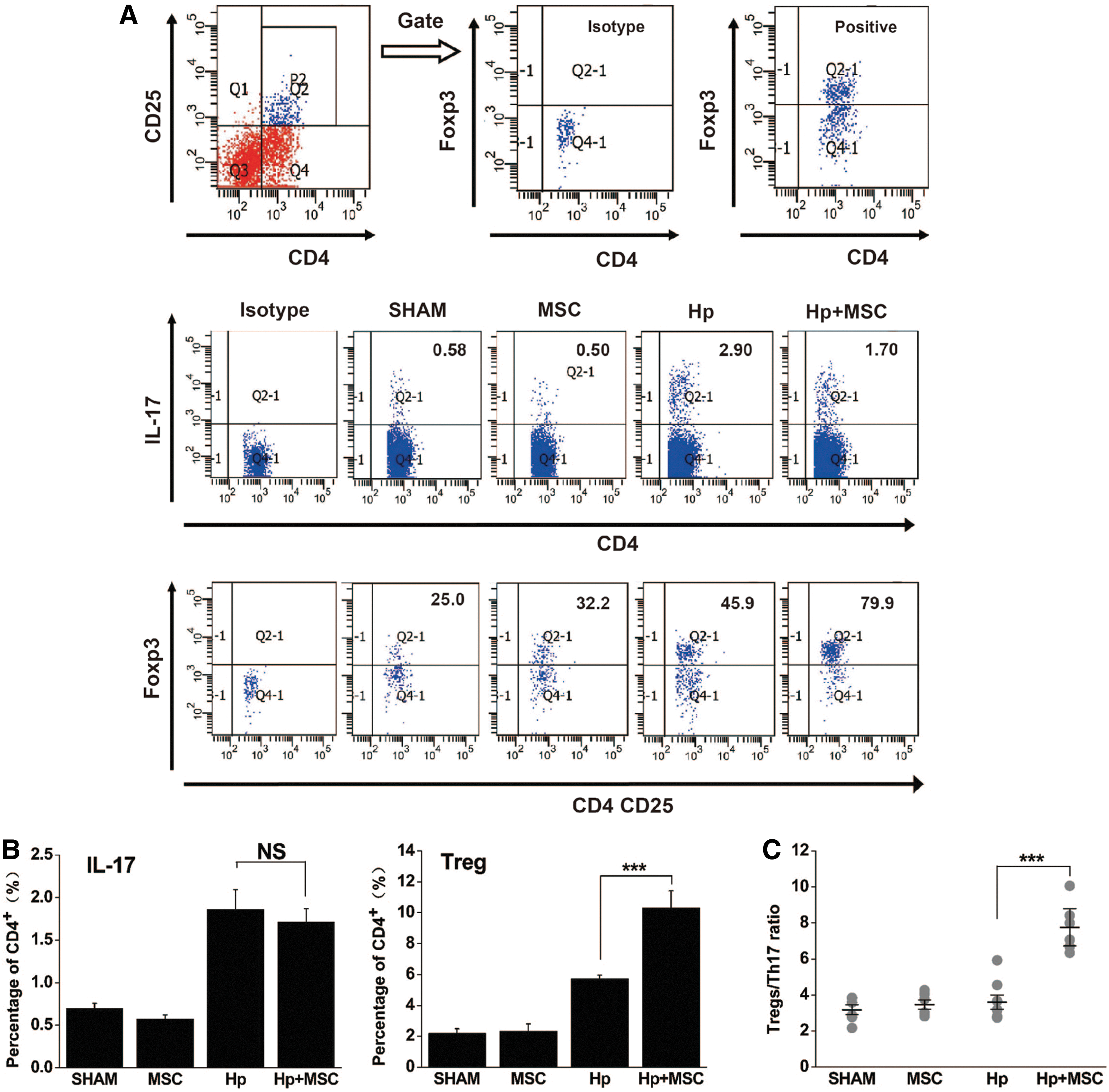
BM-MSC transplantation induces Treg responses in mice chronically infected with H. pylori.
BM-MSC transplantation induces gastric anti-inflammatory responses in mice that are chronically infected with H. pylori
To further define the influence of BM-MSC transplantation on cytokine expression, we sought to characterize the local immune response in the stomach. First, we measured the mRNA expression of cytokines IFN-γ, IL-10, IL-17, and TGF-β1 in gastric tissue using real-time quantitative PCR. The expression levels of the anti-inflammatory cytokines IL-10 and TGF-β1 in the stomach of Hp+MSC mice were significantly higher than those seen in Hp mice (P<0.001; Fig. 4A). In contrast, the expression levels of pro-inflammatory cytokines IFN-γ and IL-17 in Hp+MSC mice were reduced compared with those in the Hp mice (P<0.05).
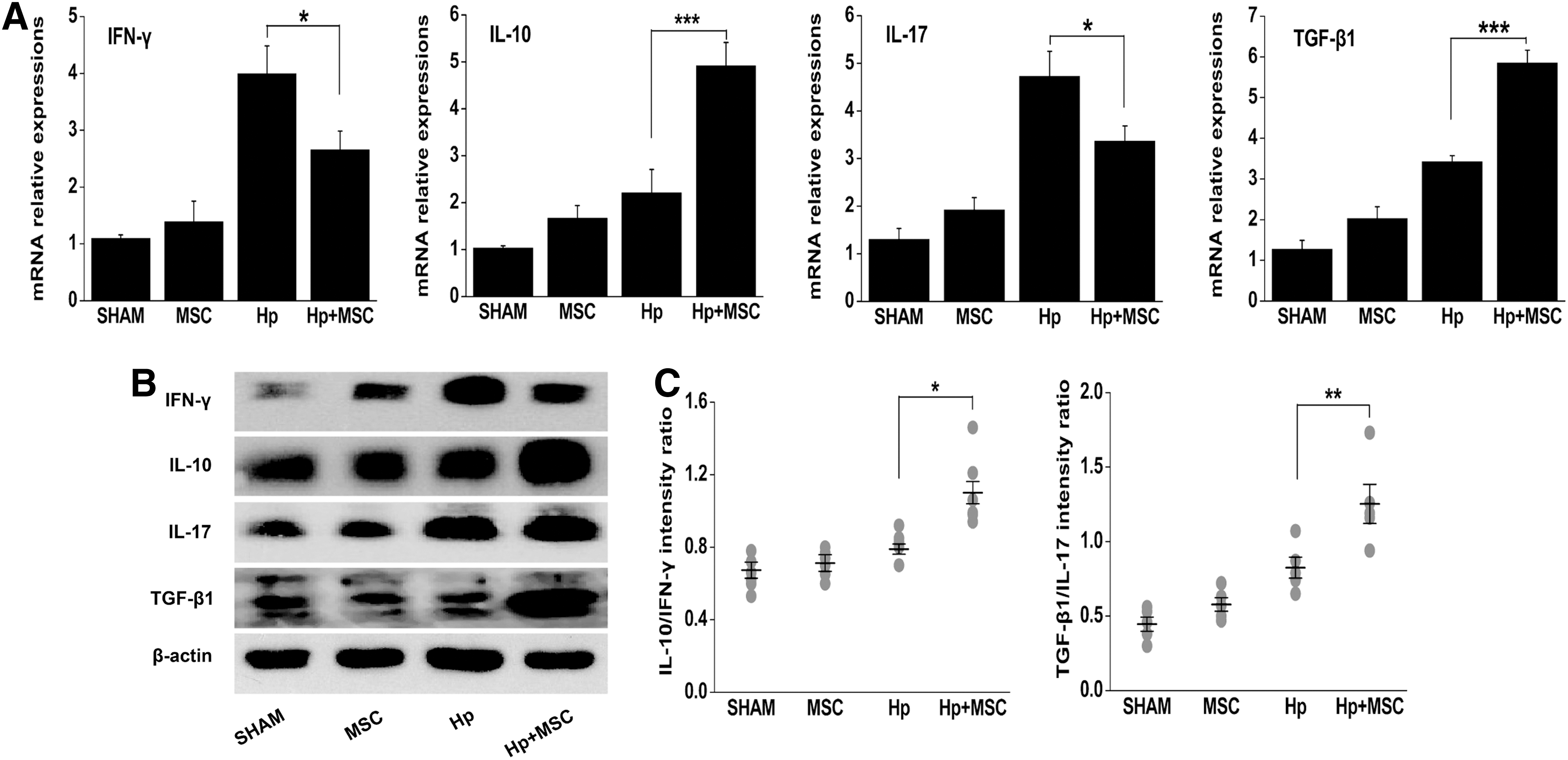
BM-MSC transplantation elevates gastric anti-inflammatory cytokine responses in mice chronically infected with H. pylori.
To further evaluate these results, we quantified the protein expression levels of cytokines in the stomach by western blot. Following BM-MSC transplantation, the protein expression levels of anti-inflammatory cytokines IL-10 and TGF-β1 were significantly increased, while the proinflammatory cytokine IFN-γ was significantly reduced, compared with mice that were chronically infected with H. pylori alone (P<0.05; Fig. 4B). However, there was not a significant decrease in expression of inflammatory cytokine IL-17. As expected, the IL-10/IFN-γ ratio and TGF-β1/IL-17 ratio in the stomach of Hp+MSC mice were strongly enhanced (P<0.05; Fig. 4C). These results are similar to the elevated levels of anti-inflammatory IL-10 that are seen in the serum following BM-MSC transplantation. Thus, BM-MSC transplantation into mice chronically infected with H. pylori induce an anti-inflammatory response that is seen both locally and systemically. These data suggest that the anti-inflammatory cytokines IL-10 and TGF-β1 that favor H. pylori persistence contribute to both systemic and local immune responses to BM-MSC transplantation during the development of gastric diseases.
BM-MSC induces immunosuppressive responses in vitro
To further clarify the effect of BM-MSC in mice with a chronic H. pylori infection, we determined the immunomodulatory roles of BM-MSC in vitro. A total of 1×106 naive splenic MNCs and 1×104 GECs were cocultured in the presence or absence of 1×105 BM-MSCs and the ultrasonic crushing of H. pylori for 24 h in transwell system. As shown in Fig. 5A and, the percentage of CD4+IL-10+ T cells and CD4+CD25+FoxP3+ T cells were significantly elevated, indicating that BM-MSCs stimulate the emergence of IL-10-producing T cells and Tregs in the microenvironment of H. pylori infection (P<0.05; Fig. 5A, B). Consistent with our flow cytometry results, the gene expressions of Gata-3, Foxp3, and IL-10 were all upregulated (P<0.05; Fig. 5C, D). These results may partially explain the increased IL-10 and TGF-β1 expression in the stomach. Together with our in vivo observations, these studies confirm that BM-MSCs induce potent anti-inflammatory and Treg responses that favor an immunosuppressive response.
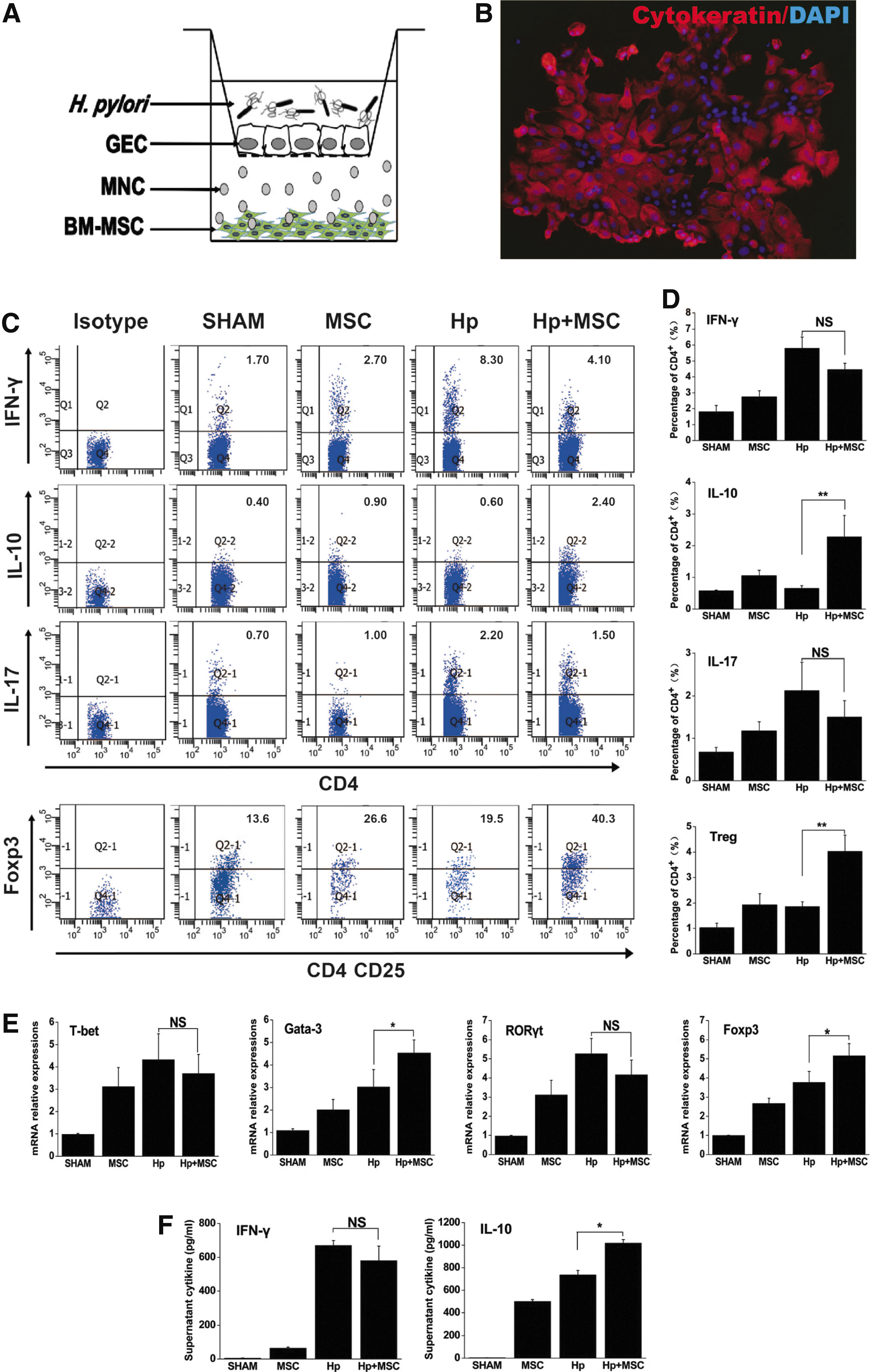
BM-MSC mediates immunosuppressive responses in splenic MNCs in vitro.
BM-MSC promotes the proliferation of GECs in vitro
To assess the role of BM-MSC in gastric cancer induced by H. pylori infection, we investigated the effects of BM-MSC on the proliferation of GECs with the presence of MNC and H. pylori infection in vitro. MTT assay showed BM-MSC increased the number of GECs in a time-dependent manner compared with control (P<0.05; Fig. 6A). To further confirm the results, CFSE dilution assay was used. According to the fluorescence intensity, cells were defined as those that had proliferated (M1). We observed a significant promotion of GECs proliferation in Hp+MSC group after 24 h coculture (P<0.01; Fig. 6B, C). These results suggested that BM-MSC promoted the proliferation of GECs with H. pylori infection.
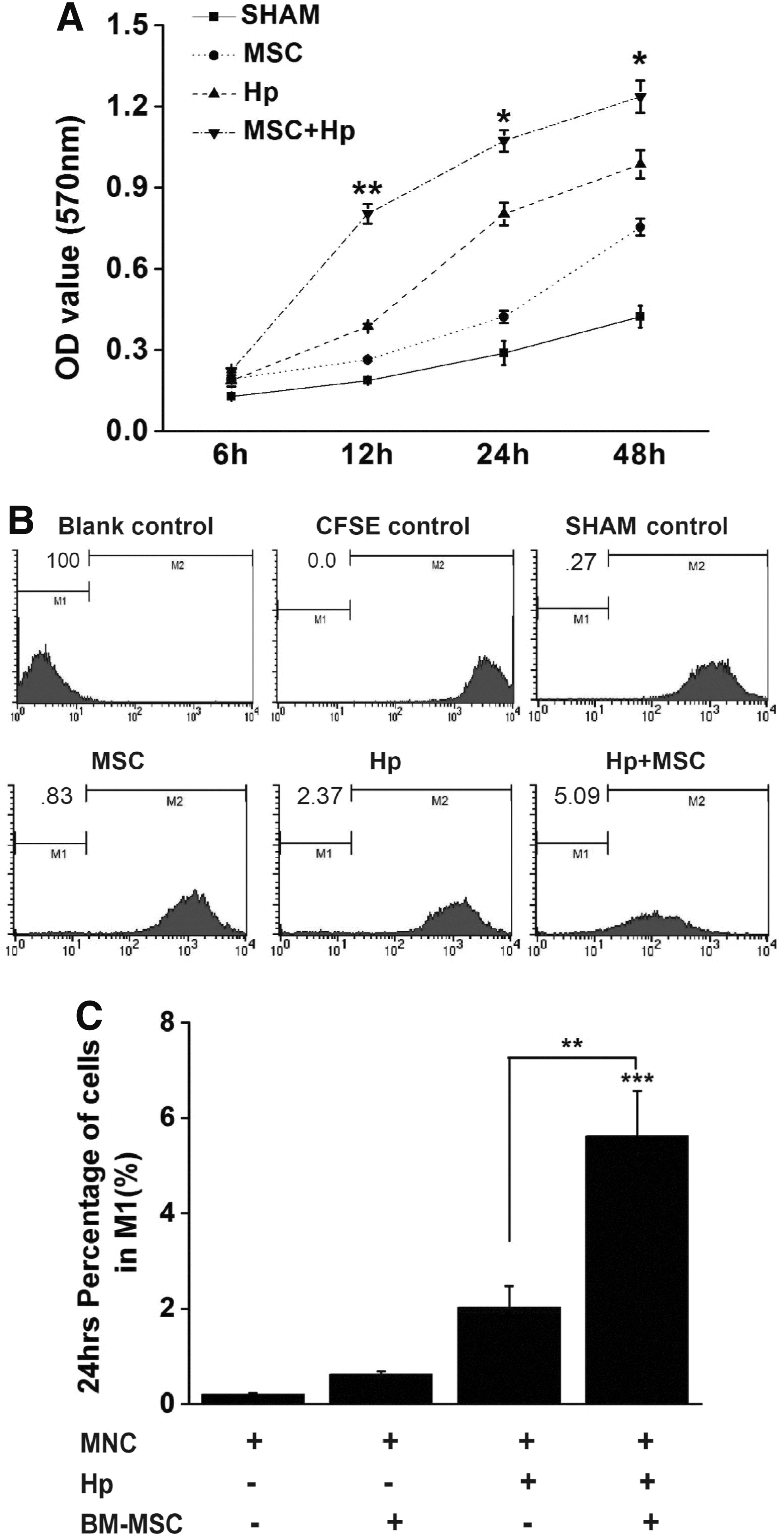
BM-MSC promotes the proliferation of GECs.
Discussion
In the current study, we evaluated the immunomodulatory role of BM-MSCs in the development of H. pylori-induced gastric cancer. This is the first in-depth report of a BM-MSC-induced immunosuppressive response that promotes H. pylori-induced gastric cancer development. Our results suggest that BM-MSCs are recruited to inflamed areas of mice chronically infected with H. pylori mice. BM-MSCs then act locally and systemically to suppress the immune response and thus promote carcinogenesis.
Gastric cancer is a common gastrointestinal tumor with a pathogenesis that is incompletely understood. BM-MSC has been shown to promote gastric carcinogenesis in a mouse model of chronic H. pylori infection [14 –16,42]. Our research also demonstrates that local BM-MSC transplantation significantly promotes gastric cancer progression. BM-MSC transplantation induces a near 3-fold increase in the incidence of gastric dysplasia in mice chronically infected with H. pylori, and the incidence of gastric cancer is elevated to more than 10%, which are significantly higher than in the Hp group (Supplementary Fig. S2). Especially, we find that BM-MSC significantly promotes the growth rate of GECs with the presence of MNCs and H. pylori infection in vitro. However, the molecular mechanisms prompting BM-MSC participation in the pathogenesis of gastric cancer remain unclear.
Cancer immunosurveillance appears to be an important host protection process that inhibits carcinogenesis and maintains cellular homeostasis [43]. BM-MSC possesses both immune-privileged and immune-inhibitory properties, which allow them to exert an immunosuppressive effect by interacting with the innate and adaptive immune systems [17,44,45]. Based on their immunosuppressive properties, BM-MSC was initially used in the treatment of immune disorders, prevention of organ transplantation rejection and tissue injury. However, several recent studies have demonstrated that BM-MSC can enhance the ability of tumors to evade immunesurveillance [46 –48]. In animal models of Ewing's sarcoma, colon adenocarcinoma, and lung cancer, the presence of exogenous MSCs has been shown to enhance tumor formation [27,49,50]. However, the ability of BM-MSC to regulate the immune environment in H. pylori-induced gastric cancer has not previously been addressed.
An experimental model of gastric cancer was established in BALB/c mice using a long-term infection (52 weeks) of the mouse-adapted H. pylori strain TN2GF4. The TN2GF4 strain is both CagA- and VacA-positive and is known to cause chronic inflammation and gastric ulcer in humans. The BALB/c mice mouse-adapted H. pylori strain was used in our study to domesticate a higher colonization rate and a more severe gastritis [28]. The sequence of events seen in this mouse model is very similar to that observed in the humans in response to H. pylori infection [51].
The recruitment of BM-MSC to sites of H. pylori-induced inflammation is crucial for the participation of BM-MSC in the development of gastric cancer. To assess the localization of BM-MSC to gastric tissue, we used the fluorescent dye Hoechst 33258, which has been used in several studies to track implanted stem cells after prelabeling just in a small part of mice [52]. The labeled BM-MSCs were further serosally transplanted into the stomach of chronically infected mice to amplify the potential role of allogenic BM-MSCs in the progression of H. pylori-induced gastric cancer. In our study, we observed that a considerable number of fluorescently labeled BM-MSCs had migrated from the serosal side of the stomach toward sites of inflammation in the gastric mucosa. This demonstrated that, in mice chronically infected with H. pylori, transplanted BM-MSCs were efficiently and specifically incorporated into the inflamed stomach.
Several studies have demonstrated that the initial immune response to H. pylori is predominantly a pro-inflammatory one. Shi et al. observed that short-term H. pylori infection induces theTh1/Th17 polarization of the immune response and blockade of IL-17 contributes to H. pylori clearance and reduces gastric inflammation [53]. Also, the groups of Bamford et al. and Wu et al. each showed high levels of pro-inflammatory cytokines IFN-γ and IL-17 during early H. pylori infection in humans [54,55]. The function of Th1 and Th17 cells during the early stage of H. pylori infection is well established [56].
However, a growing number of studies have found that the anti-inflammatory response becomes dominant during the chronic phase of H. pylori infection, including during the progression to gastric carcinogenesis [34,57]. Studies have shown that the production of suppressive cytokine IL-10 and the accumulation of CD4+CD25+Foxp3+ T (Tregs) cells in the tumor microenviroment of the H. pylori-induced gastric cancer positively correlates with disease progression. Moreover this suppressive immune response may inhibit protective antitumor immunity [10,58]. Tregs, which have been proposed to be one of the major contributions to tumor immune evasion, effectively hamper the anti-tumor immune responses during a chronic H. pylori infection [59]. The shift from a pro-inflammatory to an anti-inflammatory pattern of response may be a critical factor in the development of intestinal metaplasia and gastric carcinogenesis [10,60].
We observed that BM-MSC transplantation in mice chronically infected with H. pylori stimulated the anti-inflammatory activity of IL-10-secreting T cells and CD4+CD25+Foxp3+ T cells in splenic MNCs. Further, production of inhibitory cytokines IL-10 and TGF-β1 was enhanced while production of inflammatory IFN-γ was reduced under these conditions. The results are consistent with previous reports demonstrating that transplantation of BM-MSC induces immune tolerance associated with Treg expression after heart transplantation [23]. Thus, our study suggests that the local and systemic immunosuppressive effects of BM-MSC transplantation could be mediated through the production of cytokines that may directly act on effector T cells.
In addition, we used an in vitro system to show that BM-MSC increased the mRNA expression of the IL-10-specific transcription factor Gata-3 and the Treg-specific transcription factor Foxp3 in naïve splenic MNCs in the presence of H. pylori. As a consequence, BM-MSC showed profound immunosuppressive actions in our in vitro coculture system that were dependent on increased numbers of IL-10-secreting T cells and CD4+CD25+Foxp3+ T cells, and the production of cytokine IL-10 in supernatant. These data further indicate that BM-MSC has the potential to regulate the immune response of effector T cells in vitro and, once transplanted to H. pylori-infected mice, induce anti-inflammatory responses. These results are in agreement with an earlier study demonstrating that MSCs provide immune protection to breast cancer cells by increasing the emergence of Tregs [47]. Thus, these results could partially explain how BM-MSC is able to skew the T cell responses to anti-inflammatory response in the microenviroment of H. pylori infection.
Our results are consistent with a previous report that showed a positive correlation between skewing toward an immunosuppressive response and the progression of the H. pylori-induced gastric pathologies [60]. Our results are particularly intriguing in the light of the recent observation that a subset of tumors may exist in an “equilibrium state” specifically maintained by the adaptive immune system [25]. Most intriguingly, this raises the interesting possibility that IL-10-secreting T cells and Tregs might mediate a tumor-promoting effect in other tumor types when BM-MSC transplantation is involved.
BMDCs have been shown to promote in gastric carcinogenesis, however, the mechanisms remain unclear. Here, we demonstrate that transplantation of BM-MSCs promotes H. pylori-induced gastric carcinogenesis by regulating the balance of helper T cells. Although BM-MSC transplantation obviously does not recapitulate the exact sequence of events naturally occurring during gastric carcinogenesis in vivo, it does demonstrate that BM-MSC transplantation is capable of exerting anti-inflammatory effects through induction of IL-10-secreting T cells and generation of Foxp3 Tregs. In addition, we demonstrate that BM-MSCs are responsible for the immunosuppression required to promote the development of gastric cancer. In this regard, the skewing of the helper T cell balance induced by BM-MSC transplantation may explain their association with the pathologic progression in H. pylori-induced gastric cancer. This study also reminds us of the possible hazards involved in the use of BM-MSCs to treat patients. A better understanding of how BM-MSCs can regulate immune function is likely to provide important insights into the pathogenesis of gastric cancer and may suggest new therapeutic approaches to this disease.
Footnotes
Acknowledgments
This study was supported in part by the National Natural Science Foundation of China (no. 30670957; 30900664; 81272656; 81000160). We thank Dr. Jianzhong Zhang (Chinese Center for Disease Control and Prevention of Infectious diseases, Beijing, China) for kindly providing the H. pylori and Dr. Guanxin Shen (Department of Immunology, Tongji Medical School, Huazhong University of Science and Technology) for critical discussion of this work.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.