
Abstract
Understanding the mechanisms regulating retinal stem cell (RSC) activity is fundamental for future stem cell-based therapeutic purposes. By combining gain and loss of function approaches, we addressed whether Notch signaling may play a selective role in retinal stem versus retinal progenitor cells in both developing and adult eyes. Inhibition of either Notch or fibroblast growth factor signaling reduced proliferation of retinal stem and retinal progenitor cells, and inhibited RSC self-renewal. Conversely, exogenous Delta-like 3 and direct intrinsic Notch activation stimulated expansionary symmetric divisions in adult RSCs with the concomitant upregulation of Hes5. Knocking down Hes5 expression specifically decreased the numbers, but not the diameters, of adult RSC primary spheres, indicating that HES5 is the downstream effector of Notch receptor in controlling adult RSC proliferation. In addition, constitutive Notch activation induced retinal stem-like asymmetric self-renewal properties, with no expansion (no symmetrical division) in perinatal neural retina progenitor cells. These findings highlight central roles of Notch signaling activity in regulating the modes of division of retinal stem and retinal progenitor cells.
Introduction
S
In addition, two laboratories have sparked the controversy and suggested that all the cells within the adult mammalian CE are identical pigmented epithelial cells with a limited potential to differentiate into mature non-pigmented rod photoreceptors [16,17]. Many recent studies clearly have demonstrated that RSCs can be isolated and grown as spheres from the adult CE of distinct mammals [18 –21]. Moreover, we previously identified a very rare population (1 in 500 cells, ie, 0.2%) of pigmented cells within the CE of embryonic to adult mammalian eyes that clonally exhibits the self-renewal and multipotentiality stem cell properties [22,23]. More convincing and more supportive of a stem hypothesis are the cell sorting methods that allowed us to prospectively isolate a unique and rare population of medium-sized, heavily pigmented, P-cadherin low-expressing cells within the adult CE that exhibits RSC properties. These adult CE-isolated RSCs from adult mouse and human eyes showed also a great potential to differentiate into mature non-pigmented rod photoreceptors [24].
The adult CE consists of two distinct cell layers: the inner non-pigmented CE, which represents a continuum of the neural retina (NR), and the outer pigmented CE, which is continuous with the retinal pigmented epithelium (RPE) [25]. RSCs in the mammalian eye clonally divide in vitro to generate self-renewing, pigmented spheres with or without exogenous growth factors [23]. NR progenitors, which exist only in the embryonic and perinatal retina [26], proliferate to form clonal, non-pigmented, and most important, non-self-renewing spheres. In the present study, the presence of pigment within clonal spheres was a clear and rigorous criterion to distinguish clonal pigmented RSC-derived colonies (containing pigmented cells, accounting for rare stem cells and a majority of RPE progenitors) from clonal nonpigmented NR progenitor-derived colonies (which contain only non-pigmented NR progenitors) [13,22,23].
The regulating roles of Notch signaling in neural and retinal development have been extensively described but not specifically in individual stem cells or progenitors [9,27 –31]. In the absence of reliable markers to purely and uniquely isolate a RSC from a retinal progenitor, the clonal sphere assay represents the best option to study the specific effects of Notch activity in individual RSCs or retinal progenitors [32,33].
We externally and internally modulated Notch signaling in vitro to determine the specific functions of this cell signaling pathway in single retinal progenitors or RSCs in the perinatal post-natal day 2 (PND2) period, and in single adult RSCs. We demonstrated that Dll3 and the overexpression of Notch1 IntraCellular Domain (N1ICD) both correlated with the upregulation of Hes5 that stimulated expansion of adult RSCs. Loss of function studies showed that inhibition of either Notch or fibroblast growth factor (FGF) signaling pathways reduced the proliferation of RSCs and retinal progenitors and blocked RSC self-renewal. More specifically, downregulation of Hes5 in adult RSCs resulted in decreased proliferation of adult RSCs, but not that of retinal progenitors since only the numbers, but not the diameters of adult RSC primary spheres were reduced in presence of Hes5 small interfering RNA (siRNA) compared to control. Most interesting, the constitutive activation of Notch signaling in perinatal NR progenitor cells conferred a retinal stem-like cell identity on these cells, as assessed by the acquired FGF-dependent self-renewal properties. These findings demonstrate that Notch signaling may play an important role to enhance RSC population in vitro to treat retinal diseases.
Materials and Methods
Mice
Timed pregnant and adult (6 to 8 week-old) C57Bl6 mice (Charles River) were housed and used in accordance with “The use and care of experimental animals” guidelines. Our animal protocols were reviewed and approved by the Animal Care Committee of the University of Toronto.
Primary cell cultures, clonal sphere assay, passaging techniques
RSCs were isolated from the presumptive ciliary epithelium (pCE) and from the CE of PND2 and adult mice, respectively (Fig. 1A). Each “n” experiment used tissues dissected from at least three different animals (six eyes). Culture conditions and the clonal sphere assay were performed as previously described [13,23]. Briefly, pCE- and CE-derived cells were plated at a low cell density (10 cells/μL) in standard culture conditions containing serum-free media (SFM) [23] supplemented with fibroblast growth factor (FGF2) (F) (10 ng/mL) and heparin (H) (2 μg/mL; Sigma Aldrich) (SFM+FH).
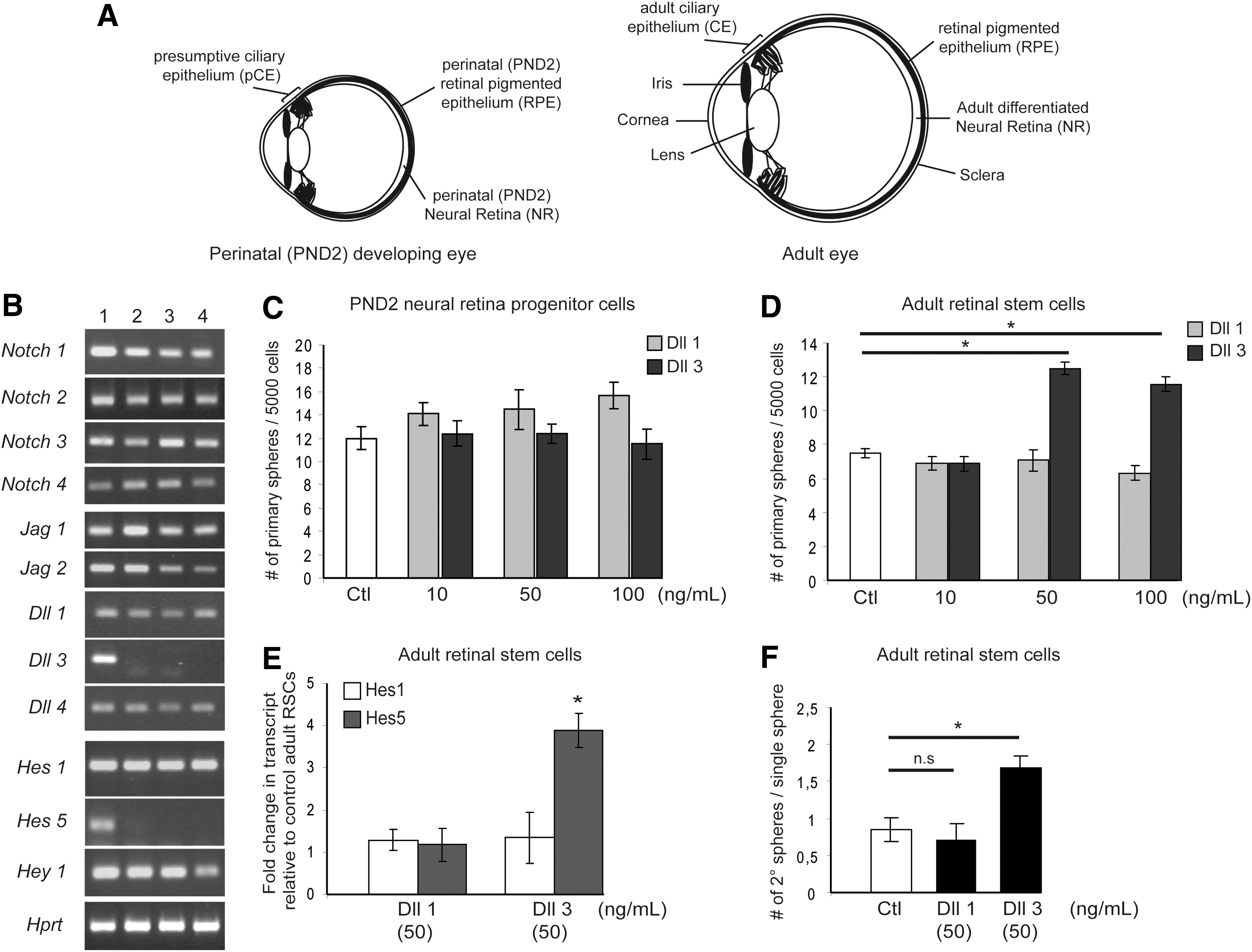
Exogenous Dll3 enhances proliferation and self-renewal of adult retinal stem cells (RSCs).
NR progenitor cells were extracted from the developing PND2 NR (Fig. 1A). NR and RPE were separated by placing eye tissues into Dispase (Collaborative Research, VWR) for 1 min (postnatal period) to 3 min (adult) at 37°C. Tissues were then mechanically triturated and dissociated to single cells. Cell suspensions were plated at 10 cells/μL in SFM+FH. Notch IntraCellular Domain (NICD)-infected NR progenitors were also passaged, then grown in a set of experiments (Fig. 4) in SFM supplemented with the epidermal growth factor (EGF, 10 ng/mL), or supplemented with both FGF2 and EGF (10 ng/mL each) after being grown in SFM+FH or SFM+EGF for the first 7 days.
To assess expansion and/or self-renewal abilities in vitro, primary colonies were passaged either in bulk or in single sphere manner. All of the colonies (bulk) or individual spheres (single sphere) were harvested and placed either in SFM only (NR progenitor-derived colonies) or in an enzyme solution (Trypsin 1.33 mg/mL, Hyaluronidase 0.67 mg/mL, Kinurenic acid 0.2 mg/mL, Collagenase type 1 and type 2 (0.5 mg/mL each), and Elastase (0.1%) (pCE- and CE-derived colonies). NR-derived colonies were readily mechanically dissociated to single cells and plated again in SFM+FH. Stem cell colonies (bulk or single) were kept in enzyme solution for 30 min at 37°C. Upon enzyme exposure spheres were triturated 30 times and then spun down for 5 min. Partially dissociated colonies were then resuspended in SFM-containing 1 mg/mL Trypsin inhibitor and triturated additional 30 times to single cells. Cells were then centrifuged again for 5 min at 1,500 rpm and resuspended in SFM+FH. Passaged spheres were counted after 7 days in culture.
Toxicity evaluation of pharmacological inhibitors
PND2 NR-derived progenitors, PND2 pCE- and adult CE-derived RSC cultures were cultured in SFM+FH in presence of the DMSO vehicle (Ctl) or in increasing doses of γ-secretase inhibitor DAPT (N-[N-(3,5-Difluorophenacetyl)-L-alanyl]-S-phenylglycine t-butyl ester; Calbiochem) from 1 to 20 μM as indicated, and/or increasing concentrations of FGF receptor inhibitor SU 5402 (Calbiochem) from 1 to 10 μM. Pharmacological inhibitors were repeatedly added every other day for 6 days. At this point, numbers of generated spheres were counted and sphere diameters measured. Inhibitor-induced toxic effects were further evaluated by harvesting the growing spheres and cells present in the different drug and dose groups, washing them twice in SFM and plating cells and forming spheres into SFM+FH. The new cultures were allowed to grow for additional 7 days and colonies were then counted again. Nonsignificant decreases in sphere numbers compared to vehicle controls after a total of 12 days in culture were considered as drug doses viable for cells. Subsequent experiments were performed using the highest non-toxic doses for both inhibitors.
Knockdown siRNA experiments
Targeted Hes5, and non-targeted control (scrambled) siRNA was used at 50 nM with DharmaFECT transfection reagent 1 (Dharmacon). Knockdown was confirmed by quantitative polymerase chain reaction (q-PCR) for Hes1 and Hes5. Hes1 levels were measured to assess the specificity of Hes5 siRNA. No significant difference in Hes1 expression levels were observed across three independent experiments. Hes5 levels were reduced to ∼65% of control.
Retrovirus preparation and infection
Replication-incompetent retroviral backbones pMX-IRES-GFP (pMXIE) and N1ICD-containing retroviral construct (pMXIE-N1ICD) were previously described [29]. PND2 NR progenitors, PND2 and adult RSCs were plated at clonal cell density (10 cells/μL) in SFM+FH. Viruses were added 6 h after plating with a Multiplicity of Infection=10 and left in culture media for 7 days. Numbers of clonal non-infected “GFP−” and infected “GFP+” spheres were determined.
RT–PCR and q-PCR
Developing (pCE) and adult CE and NR eye tissues were microdissected and RNA was extracted using RNAeasy mini kit (Qiagen) complemented with DNase treatment. RNAs were then reverse-transcribed using one-step reverse- transcription kit (Qiagen). Equivalent cDNA amounts were used to screen expression of Notch signaling components using the pairs of primers listed in Supplementary Table S1 (Supplementary Data are available online at
For q-PCR experiments, total RNA (50–100 ng, extracted from four to six spheres of about the same size) were reverse-transcribed using random hexamer primers (Invitrogen) and MoMLV reverse transcriptase Superscript III (Invitrogen) according to the manufacturer's instructions.
q-PCR were performed using Hes1, Hes5, and Hprt TaqMan Gene expression assays (Applied Biosystems) and run on 7900HT sequence Detection System (Fast real time PCR system; Applied Biosystems). Three independent experiments were performed and each sample was screened in triplicates per individual run. Quantification was performed using ΔΔCt analysis with specific reference samples as indicated in figure captions and normalized to hprt gene.
Statistics
Data are expressed as mean±SEM. GraphPad Prism 4.0 t-tests or analyses of variance were used for statistical comparisons where appropriate, with Dunnett's and Bonferroni's post hoc tests as required. Minimal statistical significance was defined at *P<0.05.
Results
In the current study, the proliferation of RSCs was monitored by evaluating the total numbers of primary or passaged clonal spheres. The average diameter of clonally expanded spheres was used as an estimate of the proliferation of retinal progenitors (given that more than 99% of cells in the spheres are progenitor cells) [23,34]. Self-renewal capacities were assessed by both bulk and single sphere passaging to address the involvement of non-cell or cell autonomous effects, respectively [13,22,23]. Moreover, the numbers of new spheres obtained from the dissociation of individual clonal spheres enabled the determination of the mode of division (symmetric giving rise to multiple secondary clonal spheres or asymmetric generating one single clonal secondary sphere) of the initial sphere-forming cell.
Exogenous Dll3 Notch ligand promotes proliferation of adult RSCs
Notch isoforms (Notch1 to 4) represent a class of transmembrane receptors that binds to transmembrane ligands (such as Delta-like 1, 3, and 4 or Jagged 1 and 2) present on the surface of adjacent cells. Receptor ligand-binding initiates two proteolytic events. The last one, mediated by the γ-secretase, releases the NICD, which is subsequently translocated to the nucleus where it binds to several partners and activates the expression of downstream target Hairy and enhancer of split (Hes) family genes [35]. Several reports in both the vertebrate and invertebrate nervous systems have established a critical role for Notch signaling in preserving a pool of undifferentiated precursor cells in developing eyes [36 –41]. Moreover, we previously showed that components of the Notch signaling pathway are critical for the maintenance (self-renewal) and expansion (proliferation) of embryonic neural and RSCs [29,42].
To investigate specific roles of Notch signaling in either RSCs or retinal progenitor cells, we first analyzed the expression patterns of Notch signaling components (ie, ligands, receptors, and target genes) within the RSC compartment (presumptive and adult ciliary epithelium, pCE and CE respectively), in dividing perinatal NR progenitors and in the differentiated adult NR. Notch 1, 2, 3, and 4 receptor isoforms and the majority of the Notch ligands, including Jagged 1, Jagged 2, Delta-like (Dll)1, and Dll4, were ubiquitously expressed in both developing (PND2) and adult CE and NR (Fig. 1B). By contrast, Dll3 is expressed only in PND2 NR progenitor cells and no longer in adult NR, nor in pCE or CE. Regarding the Notch target genes, only Hes5 was specifically expressed in the PND2 NR and not in the other tested tissue samples (Fig. 1B). This correlation between Hes5 and Dll3 expression patterns suggests a possible communication between neighboring PND2 NR progenitors as Dll3, binding to Notch receptor-expressing progenitors, which would activate downstream Hes5 expression. This result is in accordance with a previous study showing that Hes5 is expressed in dividing retinal precursors located in the germinal zone in developing mouse retina, before being gradually restricted to differentiating Müller glial cells later in the early postnatal (PND7) period [43].
We next examined the functional effects of exogenous Dll-induced Notch activation on developing PND2 NR progenitors and PND2 RSCs respectively, and on adult RSCs. Neither the addition of Dll1 nor Dll3 produced significant effects on the numbers of PND2 NR progenitor- or PND2 RSC-derived primary spheres (Fig. 1C, Supplementary Fig. S1A). These results might be explained by the fact that at PND2, both retinal stem and progenitor cells are still proliferating [13,23,44]. However, when added to adult RSC cultures, Dll3, but not Dll1, led to a 1.8-fold increase in the numbers of clonal adult pigmented RSC primary spheres compared to control (Fig. 1D). This clonal sphere assay suggests that Notch signaling activation promotes non-cell autonomous survival or proliferation of adult RSCs. In contrast, no functional effects of exogenous Dll3 were observed on retinal progenitors since both Dll3-treated PND2 and adult RSC spheres appeared to exhibit equivalent sphere diameters compared to controls (data not shown).
To further understand the underlying mechanisms upon Notch stimulation, the expression levels of Hes1 and Hes5 Notch target genes were quantified by q-PCR analysis. Dll3 stimulation specifically up-regulated Hes5 in primary adult RSC spheres, whereas Dll1 did not influence downstream Hes1 and Hes5 expression compared to control (Fig. 1E).
Using single-sphere passaging, we demonstrated that, in control conditions, single clonal PND2 RSC spheres have the ability to divide both symmetrically and asymmetrically as one initial sphere gave rise to 1.5 (from 0 up to 6 new spheres) new secondary pigmented spheres on average (Supplementary Fig. S1B). This means that some sphere-forming stem cells divided at least once symmetrically. In contrast, in the same conditions, single clonal adult RSC spheres did not generate more than one (either zero or one new sphere) new secondary pigmented sphere, indicating that adult RSCs only divided asymmetrically in our culture conditions (Fig. 1F). Single dissociated PND2 RSC spheres cultured in the presence of exogenous Dll1 or Dll3 did not exhibit any differences in the numbers of passaged pigmented spheres compared to control (Supplementary Fig. S1B). Nonetheless, single adult RSCs, in presence of Dll3, but not Dll1, produced 1.8 times more secondary pigmented spheres thus confirming effects of exogenous Dll3 on adult RSC survival or proliferation. The proportion of individual adult RSC primary spheres that did not passage in Dll3-treated (28%±2%) versus control (30%±3%) groups was equivalent, thus ruling out Dll3-induced survival effects on adult RSCs and rather suggesting enhanced symmetric divisions. We hypothesized that this variability in response to ligand stimulation between PND2 and adult RSCs may be due to the fact that, in early in vivo postnatal development, Notch signaling activity already is maximal and could not therefore be more activated with the addition of Dll ligands, thus enabling RSCs to proliferate both symmetrically and asymmetrically to contribute to retina formation [13,26] (Supplementary Fig. S4D).
Inhibition of the Notch signaling pathway reduces the proliferation of RSCs and retinal progenitors
To test whether endogenously active Notch signaling is functional in developing and adult eyes, PND2 NR progenitors, PND2 pCE-derived RSCs, and adult CE-derived RSCs were cultured in presence of DAPT in clonal sphere assays. This drug is known to inhibit the γ-secretase enzyme activity responsible for the cleavage of Notch receptor following ligand binding, thus preventing the release of the N1ICD [45]. First, the toxicity of this inhibitor on the various cell types was evaluated by performing a dose–response curve to increasing concentrations of DAPT (1 to 20 μM). DAPT dose-dependently diminished the numbers of pigmented PND2 and adult RSC primary spheres, indicating a reduced proliferation or survival of RSCs compared to vehicle control groups (Supplementary Figs. S2B and S3A). However, no toxic effects of DAPT were observed on PND2 and adult RSCs up to the 10 μM DAPT dose, as the 7 day-rescue period allowed the generation of equivalent numbers of pigmented primary spheres compared to controls. Hence, we conclude that inhibition of Notch diminishes the proliferation of PND2 and adult RSCs. This result tallies with our previous data demonstrating that perturbation in Notch signaling (absence of the presenilin1 gene) impaired the numbers of embryonic RSC primary spheres [42]. Moreover, DAPT-treated PND2 and adult RSC primary spheres were significantly smaller in size compared to controls (Supplementary Figs. S2C and S3B). This smaller size phenotype could be due to either proliferation or survival defects on retinal progenitors. However, the analysis of DAPT treatments on PND2 NR progenitors led us to rule out the death of stem cell-derived progenitors in spheres, as the clonal sphere-forming ability of PND2 NR progenitors exposed to the 10 μM DAPT dose could still be rescued by transfer to standard culture conditions (Supplementary Fig. S2A). Therefore, these data demonstrate that Notch signaling regulates proliferation of retinal stem and retinal progenitor cells in vitro.
Additionally, to control the pharmacological specificity to Notch signaling, PND2 NR progenitors and adult CE-derived cells were stimulated by Dll1 or Dll3 in presence of DAPT. Neither of the Notch ligands restored the numbers of progenitor- nor stem cell-derived primary colonies when DAPT was included in the cultures (Fig. 2A, B). We thus conclude that the Dll3-mediated proliferation on adult RSCs (Fig. 1C) occurred through activation of the canonical Notch signaling pathway, as this proliferative effect was inhibited in presence of DAPT (Fig. 2B). Since the highest non-toxic dose of DAPT did not completely suppress the formation of PND2 NR progenitor- and adult RSC-derived primary spheres, we hypothesized that at least one other cell signaling pathway may be responsible for this remaining proliferation.
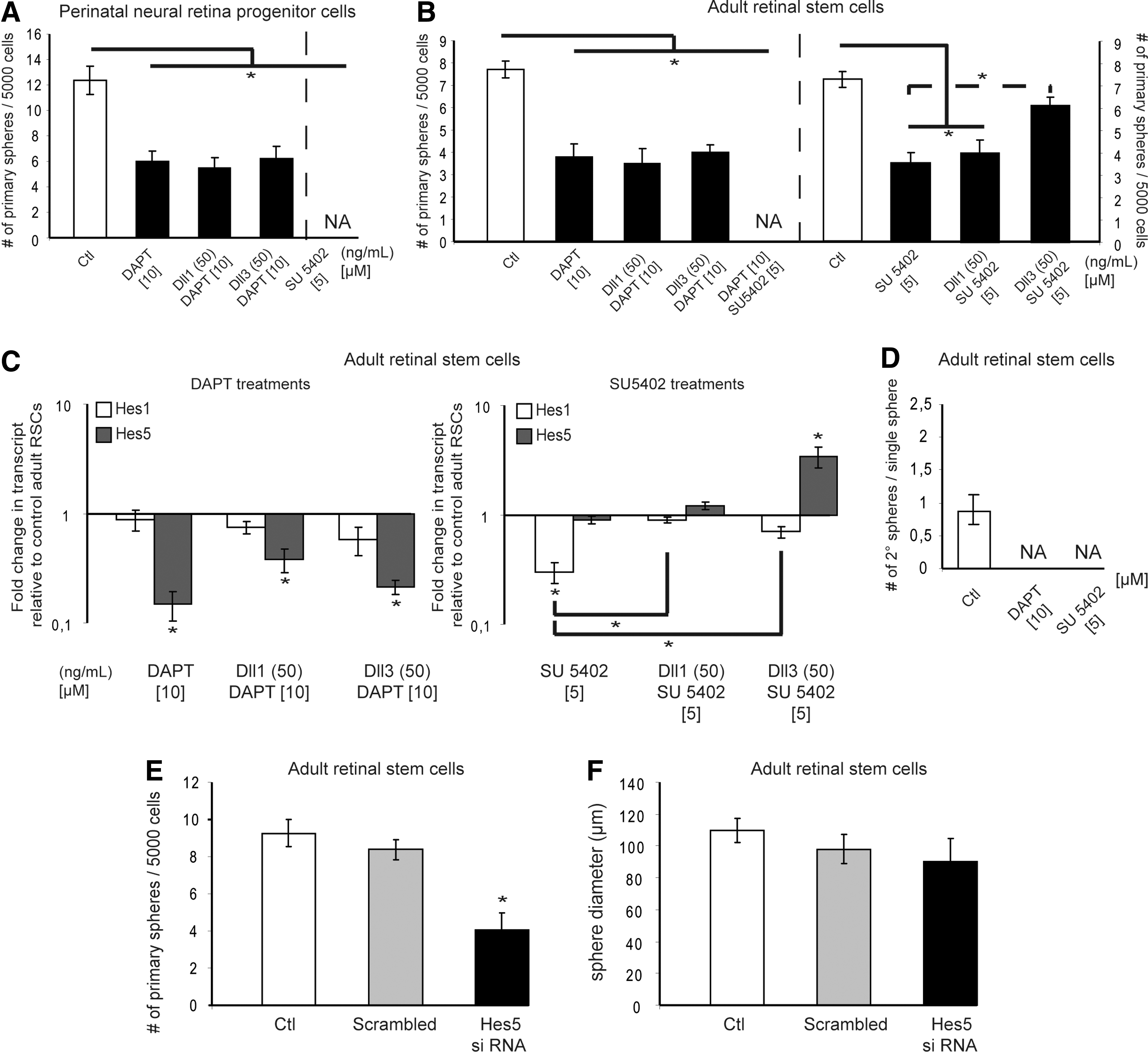
Notch and fibroblast growth factor (FGF) signaling pathways are involved in the proliferation of retinal stem and retinal progenitor cells and both are required for stem cell self-renewal.
FGF signaling pathway regulates RSC proliferation and is required for retinal progenitor proliferation
Although an initial study demonstrated that a rare subset of pigmented cells from the adult CE is able to form clonal pigmented spheres in the absence of exogenous growth factors, this work also clearly established that exogenous FGF2 promotes RSC proliferation [23]. Further, exogenous Dll3 was shown to promote proliferation of adult RSCs in our standard FGF2-containing culture medium (Fig. 1D). This additional growth led us to anticipate that both Notch and FGF signaling pathways may participate in RSC proliferation. A specific FGFR1 inhibitor, SU5402, was used to block FGF signaling and its toxicity was assessed through a dose–response curve on adult CE-derived cells. SU5402 exposure led to decreased numbers and diameters of adult clonal RSC colonies (Fig. 2B) in a dose-dependent manner (Supplementary Fig. S3C, D). At the 5 μM dose, the numbers of adult pigmented RSC primary spheres could be restored following the 7-day rescue period in control media, thus excluding any potential toxic effects. We conclude that inhibition of FGF signaling alters adult RSC proliferation.
The reduced sphere diameters can be due to impairment of retinal progenitor proliferation since blocking the FGF signaling with 5 μM of SU5402 completely blocked the primary sphere formation of PND2 NR progenitors (Fig. 2A and Supplementary Fig. S3D). The fact that adult RSC primary spheres were observed at the 5 μM SU5402 dose (Fig. 2B and Supplementary Fig. S3C) eliminated a potential toxicity on retinal progenitors and suggested that FGF signaling is involved in retinal progenitor proliferation. These results support the idea that FGF signaling stimulates adult RSC proliferation, while being required for the expansion of PND2 NR progenitor cells. All together, these data demonstrate the involvement of both Notch and FGF signaling pathways in RSC proliferation. Inhibiting both Notch and FGF signaling pathways together resulted in no primary sphere formation from adult RSC cultures (Fig. 2B), providing a direct piece of evidence for the additive effect of these two signaling pathways.
Since clonal adult pigmented RSC primary spheres were generated in FGF2-supplemented culture media and in the presence of DAPT, (Fig. 2B), we next examined the respective importance of both FGF and Notch signaling pathways in adult RSCs. To do this, we cultured adult RSCs in the reverse culture conditions, thereby stimulating Notch while inhibiting FGF signaling pathways. In presence of SU5402, Dll3 but not Dll1 increased the numbers (Fig. 2B) but not the sizes (data not shown) of adult pigmented RSC primary spheres compared to adult RSC spheres grown in the FGFR inhibitor alone. Such an increase in sphere numbers was not observed when adult RSCs were grown in the presence of DAPT with an excess of exogenous FGF2 compared to the spheres cultured in DAPT alone (data not shown). Therefore, Notch signaling demonstrates preferential effects on RSCs, and FGF signaling on retinal progenitors.
Molecular crosstalk between FGF and Notch signaling pathways in clonal adult RSC colonies
To elucidate the downstream molecular effects of the inhibition of the separate Notch and FGF signaling pathways, the levels of the Hes1 and Hes5 Notch target genes were quantified in adult RSC primary spheres. qPCR analyses revealed that DAPT-exposed adult RSC primary spheres, with or without the addition of Dll1 or Dll3 Notch ligands, downregulated Hes5 expression compared to control (Fig. 2C). Besides, adult RSC spheres cultured in presence of FGFR inhibitor alone show decreased Hes1, but not Hes5 expression compared to controls. Moreover, SU5402-treated adult RSC spheres restored Hes1, but not Hes5 expression to normal levels when Dll1 was added, whereas Dll3 induced the specific up-regulation of Hes5 (Fig. 2C). These results underline a crosstalk between FGF and Notch signaling pathways in regulating retinal progenitor proliferation, since inhibiting FGF signaling affects the downstream Hes1 Notch target gene. These findings also suggest the combination of the Dll3 (ligand)-Notch receptor-HES5 as a preferential pathway to stimulate adult RSC proliferation. To confirm this assumption, we assessed the knockdown of Hes5 expression on adult RSC cultures. The 65% reduction in Hes5 transcripts led to a 50% decrease in numbers, but not in diameters of clonal pigmented adult RSC primary spheres (Fig. 2E, F). All together, these results and the fact that Hes5 expression was not detectable in adult CE (Fig. 1B) reinforces the idea that Notch-HES pathway is predominantly present in rare pigmented RSCs in vivo.
Notch and FGF cell signaling pathways are both required for adult RSC self-renewal
To address the roles of the FGF and Notch signaling pathways in the self-renewal of adult RSCs, CE-derived cells were initially cultured for 7 days in the presence of either DAPT or SU5402. Individual spheres with similar diameters from the control and inhibitor-treated groups were subsequently passaged into inhibitor-free standard culture conditions. As already shown (Fig. 1E), single clonal adult RSC colony gave rise to no more than 1 new sphere on passaging, thus demonstrating that adult RSCs solely divide asymmetrically (Fig. 2D). However, in the presence of either DAPT or SU5402 for 7 days, individual clonal adult RSC primary spheres did not form any secondary spheres in drug-free conditions, suggesting that the Notch and FGF signaling pathways both are necessary for adult RSC self-renewal.
Constitutive activation of Notch signaling pathway confers retinal stem-like cell features on PND2 NR progenitor cells
High Notch signaling activity appears to distinguish neural stem cells from their downstream intermediate neural progenitors [32]. To look at this notion in the retina, we addressed whether the constitutive activation of Notch signaling in single NR progenitors may revert them back to a stem cell state. PND2 NR progenitors were infected with retroviruses containing the N1ICD, the portion that is cleaved and translocated to the nucleus when Notch receptor is activated. No difference was observed in the total numbers of primary colonies obtained from both infected groups compared to the non-exposed group. Moreover, control GFP- and N1ICD-GFP-overexpressing cells produced equivalent numbers of clonal non-pigmented primary GFP+ spheres (Fig. 3A). All of the colonies from the three [non-exposed (−), GFP (pMXIE), N1ICD (pMXIE-N1ICD)] groups were then passaged in bulk. The overexpression of N1ICD induced self-renewal, as shown by the formation of clonal non-pigmented secondary spheres compared to control groups that did not show such self-renewal properties (Fig. 3B). A biased survival effect of FGF2 on infected NR progenitors can be ruled out as NICD-infected NR progenitors initially grown in EGF alone gave rise to equivalent numbers of primary spheres (data not shown). Moreover, if grown in serum-containing media for 3 weeks, these NICD-infected NR progenitor spheres showed multipotentiality as they differentiated into retinal neurons and glial cells (Balenci and van der Kooy, unpublished data) [41].
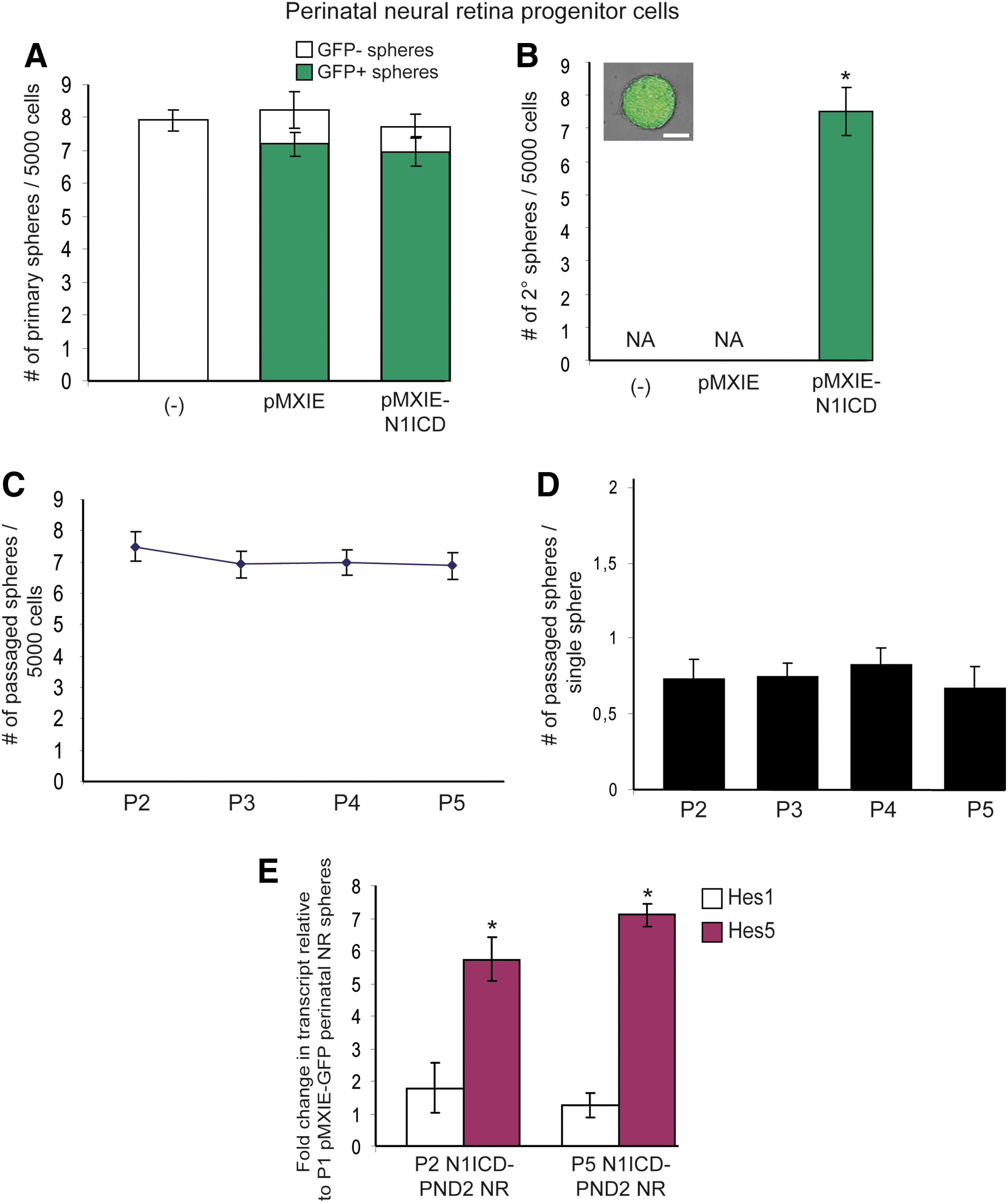
Constitutive activation of Notch signaling pathway induces self-renewal properties to perinatal NR progenitor cells.
To further examine the acquired self-renewal properties of PND2 NR progenitors in response to sustained Notch activity, non-pigmented N1ICD-infected NR progenitor colonies were passaged either in bulk or as single spheres. In bulk, we observed that the frequency of sphere forming cells was consistent over passages, suggesting that each single N1ICD-infected NR progenitor gives rise to a single new sphere-forming cell. These sphere-forming cells do not expand their numbers by symmetric division, but only asymmetrically divide to maintain the single sphere-forming cell in each colony (Fig. 3C). In support of this hypothesis, individual N1ICD-infected spheres generated no more than one new sphere on passaging, confirming that the initial sphere-forming cells did not symmetrically divide as this would have led, at least, to two new clonal spheres (Fig. 3D). Most interesting, neither N1ICD-infected primary colonies nor N1ICD-induced secondary spheres were larger in size than GFP-infected spheres (pMXIE), thus reinforcing the conclusion of Fig. 1C that activation of Notch signaling does not stimulate proliferation of retinal progenitors.
To identify the molecular components involved in this N1ICD-induced self-renewal ability, q-PCR analyses were carried out. N1ICD-overexpressing PND2 NR progenitor-derived spheres revealed a seven-fold increase in Hes5 expression compared to primary PND2 NR progenitor colonies, whereas Hes1 expression remained unchanged (Fig. 3E). Further, the up-regulation of Hes5 expression was maintained over time in N1ICD-infected spheres until at least the fifth passage (the last passage tested) (P5 N1ICD-PND2 NR, Fig. 3E). The constitutive activation of Notch signaling in PND2 NR progenitors correlated with an increase in Hes5 expression during the process of turning NR progenitors into retinal stem-like cells, thereby changing the mode of division of the sphere-forming cell from symmetric progenitor divisions to strictly asymmetric stem cell-like divisions. Noteworthy, none of the N1ICD-infected NR progenitor-derived spheres became pigmented, meaning that, they did not acquire all of the features that RSCs normally possess. However, we previously found that pigmentation is not crucial for RSC function, since adult albino CD1 mice harbor normal numbers of non-pigmented, colony sphere-forming RSCs that self-renew in vitro [23].
To further investigate these induced self-renewal properties, primary PND2 NR progenitor spheres overexpressing N1ICD were passaged in bulk and plated in EGF, FGF2, or EGF+FGF2, two growth factors shown to promote retinal progenitor and adult RSC proliferation [23]. Removal of exogenous FGF2 during passaging led to the formation of less secondary colonies and prevented the emergence of tertiary spheres (Fig. 4A, C). In addition, in the absence of FGF2, spheres were smaller in diameter emphasizing the role of FGF2 in promoting proliferation of retinal progenitors (Fig. 4B, D). These findings demonstrate that constitutive activation of Notch signaling led single PND2 NR progenitor cells to a stem-like cell identity, since these cells were induced to self-renew, a function for which exogenous FGF2 was required.
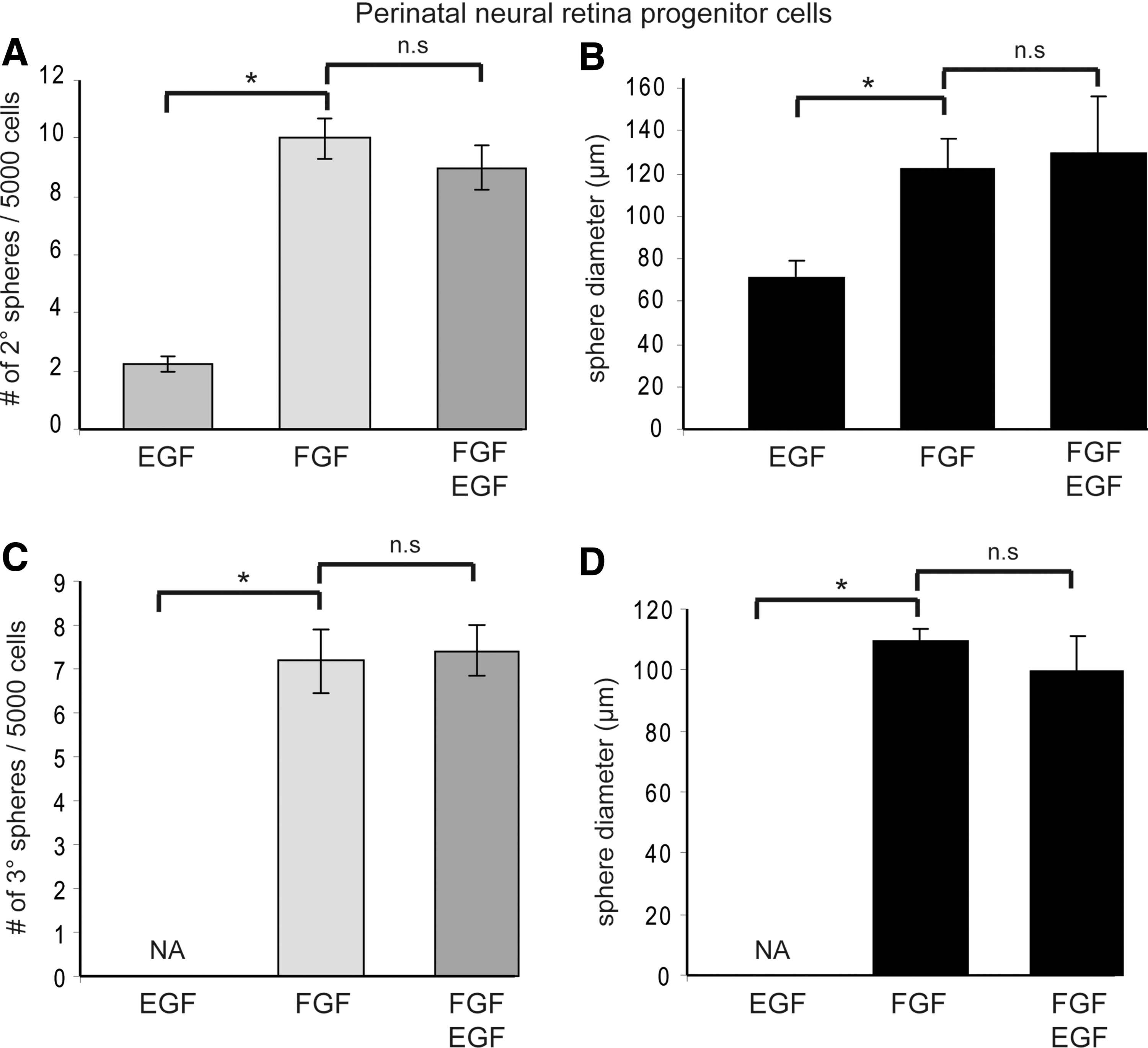
Gained self-renewal abilities of NR progenitors are FGF-dependent.
Constitutive activation of Notch signaling pathway induces symmetrical divisions in adult RSCs
Since Notch activation induced self-renewal in PND2 NR progenitors, we next addressed the potential role of Notch activation in PND2 and adult RSCs that possess self-renewal abilities. GFP(pMXIE)- and N1ICD(pMXIE-N1ICD)-overexpressing PND2 RSCs generated the same proportions of non-infected (GFP−) and infected (GFP+) clonal pigmented primary and secondary colonies (Supplementary Fig. S4A–C). The overall numbers of colony-forming cells were comparable between both infected groups (pMXIE and pMXIE-N1ICD) and the non-exposed group (−) over the first two passages ruling out any potential survival and proliferative effects of Notch activation on single PND2 RSCs. This result is consistent with the observation that exogenous Dll3-mediated Notch activation did not influence PND2 RSC proliferation (Supplementary Fig. S1A, B). It is important to note that individual non-infected (−) or infected GFP+ (from both control GFP- and N1ICD- groups) colonies generated more than one new sphere at each passage suggesting that PND2 RSCs divide both symmetrically (to generate two identical daughter RSCs) and asymmetrically (to maintain RSC population while also generating retinal progenitor cells) (Supplementary Fig. S4C).
Adult CE-derived cells submitted to N1ICD overexpression produced equivalent numbers of clonal infected (GFP+), pigmented primary spheres of comparable sizes to adult RSC cultures grown in the presence of the control GFP virus (Fig. 5A, B). Nevertheless, once passaged in bulk, N1ICD-infected clonal adult RSC-derived spheres generated twice as many pigmented clonal secondary colonies (which were also smaller in diameter) as the control GFP-infected colonies (Fig. 5C, D). The proportion of non-infected (GFP−) spheres between GFP- and N1ICD-overexpressing groups was similar; this suggests that Notch activation promotes survival or proliferation of adult RSCs. However, a survival hypothesis seems less likely given that N1ICD-overexpressing group did not show increased numbers of clonal pigmented primary colonies (Fig. 5A).
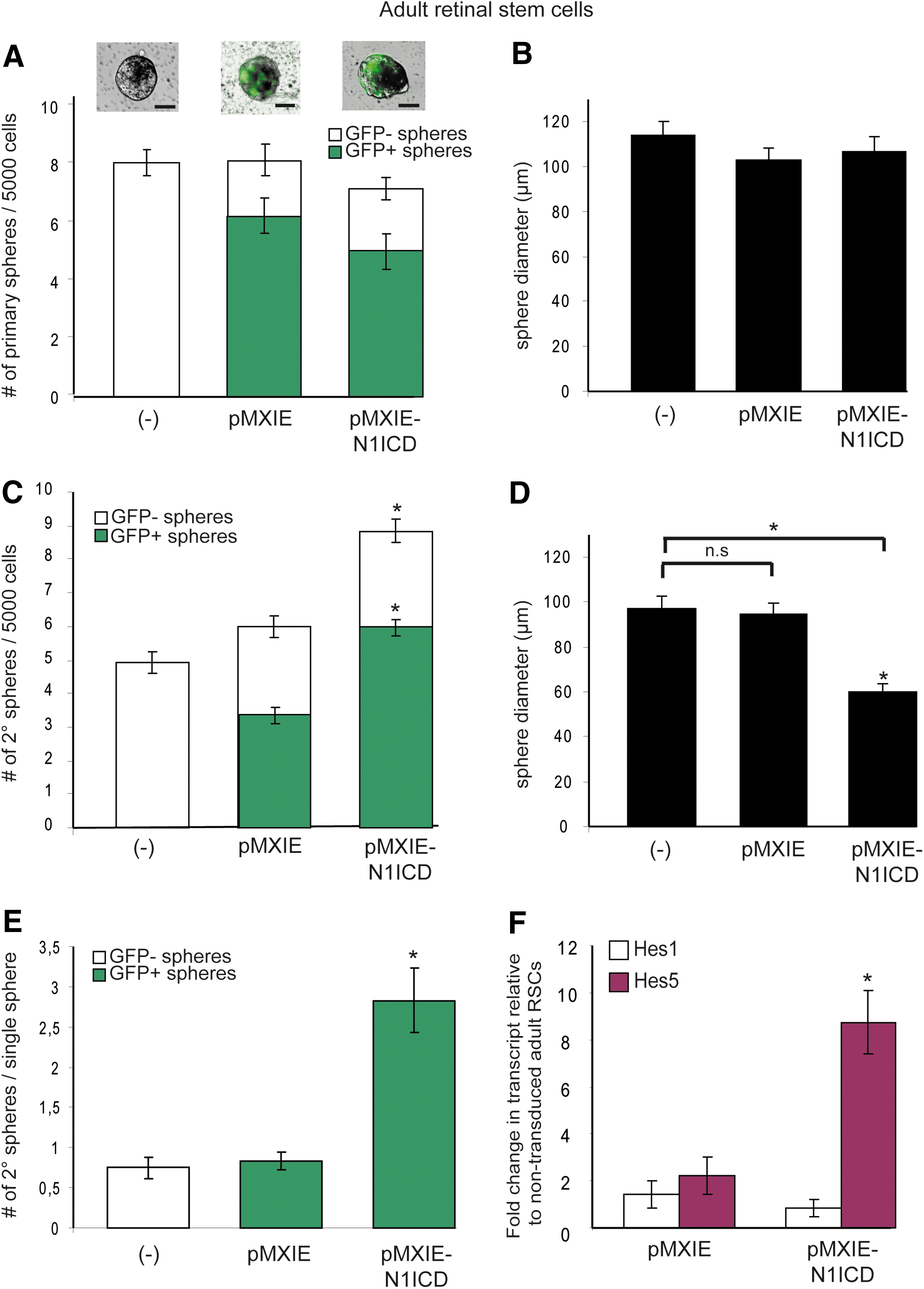
Sustained Notch signaling promotes symmetrical divisions in adult RSCs.
To examine whether the constitutive Notch activation has an effect on proliferation, individual non-infected (−) and infected (GFP+) primary spheres from control GFP- and N1ICD-overexpressing groups were passaged. Clonal adult RSC spheres overexpressing N1ICD produced three times more secondary clonal pigmented spheres compared to the non-exposed and control GFP groups (Fig. 5E). The proportion of infected GFP+ spheres that did not passage was equivalent between the control GFP- (pMXIE, 33%±4%) and N1ICD-overexpressing groups (pMXIE-N1ICD, 32%±3%), thus excluding survival effects.
One other alternative to proliferative effect might be that Notch activation, as shown in this study for PND2 NR progenitors, turns some of the pigmented RPE progenitors into retinal stem-like cells able to form clonal spheres. To test this potential conversion, PND2 RPE progenitor cells were isolated (from the central retina rather than the pCE region) and cultured in proliferating conditions (see Supplementary Fig. S5) and subsequently infected with either the control GFP virus (pMXIE) or the N1ICD-GFP-containing virus (pMXIE-N1ICD) 24 h after the initial plating. On day 7, 47%±5% in the control group and 39%±3% in the N1ICD-group of PND2 RPE progenitor cells exhibited GFP expression (Supplementary Fig. S5). However, no primary spheres emerged in either group, indicating that Notch activation did not promote proliferation nor took RPE progenitors back to RSC state. In sum, these results show that the constitutive activation of Notch signaling pathway promotes symmetrical divisions of the adult RSCs (Fig. 5D, E). Additionally, we further identified Hes5 as the principal mediator in the expansion of adult RSC population (Figs. 2E and 5F). In conclusion, N1ICD overexpression prompted adult RSCs to switch from a completely asymmetric to a more symmetric mode of division, probably at the expense of retinal progenitor proliferation since N1ICD-transduced adult RSC secondary spheres decreased in size (Fig. 5D).
Discussion
The balance between the symmetric and asymmetric divisions of stem cells is tightly controlled during development. In the developing eye, retinal precursors symmetrically divide to increase the stem/progenitor pool before switching to an asymmetric mode of division to maintain resident stem/progenitor pool while generating differentiated progeny [46]. Although we and others have shown that RSCs may continue to expand through perinatal development, potential intrinsic and extrinsic factors regulating RSC activity remain elusive [13,23,29,47]. Understanding the molecular mechanisms controlling RSC proliferation and self-renewal and whether these processes are interchangeable are crucial for therapeutic purposes. This may allow targeted therapies triggering the proliferation of quiescent endogenous RSCs and the differentiation of their progeny into appropriate retinal cells to replace lost or injured cells. Numerous studies have shown that Notch signaling plays important roles in maintaining progenitors in an undifferentiated state and in promoting glial cell fate in vertebrate developing retina [30,38,40,41,48,49]. However, little is known about Notch functions in individual retinal stem/progenitor (precursor) cells during perinatal retina development and in the adult.
The present study investigates the role of Notch signaling in perinatal RSC versus retinal progenitors and in adult RSCs (Table 1). Reports on Drosophila and vertebrates suggested that the role of Notch signaling depends on the developmental and cellular contexts of a given tissue [9,35]. We show that external and internal modulations of the Notch signaling activity acts as an important molecular component in proliferating PND2 NR progenitors and in adult self-renewing RSCs to influence their respective modes of division. Enhanced Notch signaling activity in PND2 NR progenitors induced self-renewal by producing secondary clonal, nonpigmented spheres, whereas in uninfected or in control conditions, these cells formed no secondary spheres. Increased cell survival cannot account for these secondary spheres as no difference in primary sphere formation was observed. Moreover, passages of single N1ICD-infected PND2 NR progenitor-derived spheres produced no more than one new sphere, indicating that normally proliferating perinatal NR progenitors switch their mode of division to strictly asymmetric. Our result slightly differs from a previous study where constitutive Notch activation in embryonic retinal progenitors led to the formation of very rare pigmented, self-renewing colonies [41]. Perhaps, the developmental stage and the environmental signals may help induce biochemical components required for pigmentation. The absence of pigmentation led us to propose that increased Notch activity in PND2 NR progenitors induced a “partial reprogramming” giving rise to self-renewing non-pigmented spheres. Closer analysis of the self-renewal features of N1ICD-induced retinal stem-like cells revealed properties similar but not identical to normal self-renewing adult RSCs in vitro. These reverted NR progenitors exclusively divide asymmetrically and these divisions are dependent on exogenous FGF2, as opposed to adult RSCs, which can divide without exogenous factors [23]. Further genetic analyses, however, are needed to bring insights into the molecular mechanisms involved in this reversion process and identify which signaling pathways and/or genetic programs were reactivated in response to high Notch activity. A previous report showed that overexpressing C-promoter binding factor (CBF-1), a co-factor promoting Notch transcriptional activity, failed to revert neural progenitors back to neural stem cells [32], which makes the present demonstration of a reversible state between retinal progenitors and RSCs unique. Other studies in brain and hematopoietic systems have previously correlated a high Notch activity in the stem cell population [32,50]. The fact that the extrinsic and intrinsic increase of Notch signaling activity had no effects on PND2 RSCs suggests that a substantial Notch activity already exists in such RSCs.
Table highlighting the basic proliferation and self-renewal properties of the different retinal cell types presented in this study including the PND2 NR progenitors, PND2 RSCs, and adult RSCs. The schematic summarizes the different effects observed on the different initial sphere-forming cells following N1ICD overexpression (gain of function) or DAPT treatment (loss of function).
N1ICD, Notch 1 IntraCellular Domain; NR, neural retina; PND2, postnatal day 2; RSCs, retinal stem cells.
By contrast, reducing Notch activity decreased, but did not abolish, the proliferation of PND2 and adult RSCs, since, despite the DAPT treatment, some perinatal and adult RSC primary spheres were generated. Emerging RSC spheres also displayed smaller sphere diameters indicating that DAPT perturbed also retinal progenitor proliferation. The 5-day DAPT treatment transiently induced a cell cycle arrest of RSCs and retinal progenitors since these cells resumed proliferation to form equivalent numbers of primary spheres with comparable diameters to controls when cultured back in standard culture conditions. The presence of some remaining spheres in DAPT-treated cultures suggests that the off-target toxic effects at higher doses might have prevented us from observing a complete inhibition of RSC-derived sphere formation by blocking the Notch function. Alternatively, the remaining clonal RSC-derived spheres might reflect the presence of different RSC and retinal progenitor populations not dependent on Notch signaling to divide. Since the inhibition of FGF signaling impaired proliferative potential of adult RSCs while it completely inhibited proliferation of PND2 NR progenitors, the second hypothesis seems more likely.
In the developing retina, progenitors progress through a series of “competent stages” in which retinal progenitors present variable differentiation abilities [30]. This evolving competence of progenitors might parallel upstream in the cell lineage the stem cell capacity to divide symmetrically or asymmetrically, in which Notch signaling activity would play a crucial role [27,32,51 –53]. Although, constitutive activation of Notch was shown to have no or little effects on the competence of early progenitor cell differentiation [41,54], we compared the functions of activated Notch signaling in PND2 versus adult RSCs. In the developing eye within the pCE, PND2 RSCs still divide, whereas RSCs become quiescent within the adult CE and no longer proliferate even following injury [9,10]. However, once extracted from their niche, both PND2 and adult RSCs are able to generate clonal primary pigmented colonies in vitro. Subsequently, passaging of individual pigmented PND2 RSC primary colonies resulted in more than 1 new secondary pigmented sphere, whereas a clonal adult RSC sphere did not produce more than one secondary pigmented sphere. Consequently, PND2 RSCs have the ability to both symmetrically and asymmetrically divide, while adult RSCs adopt only an asymmetric mode of division. This observation might explain why the constitutive activation of Notch signaling induced no changes in PND2 RSCs, whereas it increased the numbers of adult RSC colonies on passaging.
Hes5 was reported in other studies to be the most reliable Notch target genes and a good read-out of the Notch activity [32,55,56], whereas Hes1 expression was shown to be regulated by other cell signaling pathways [57,58] (the present study). The Notch-induced competence of adult RSCs to symmetrically divide is further supported by the concomitant upregulation of Hes5 expression to reach a level similar to the one quantified in PND2 RSCs in which, we assumed, Notch activity is maximal (Supplementary Fig. S4D). A similar upregulation of Hes5 was observed upon Dll3 stimulation in adult RSCs, which also resulted in enhanced adult RSC proliferation, as attested by the augmentation in the numbers of primary and secondary pigmented spheres. Conversely, downregulating Hes5 reduced specifically the numbers, but not the diameters, of adult RSC primary spheres, thus emphasizing the effector role of HES5 in adult RSC proliferation. Levels of ligand and receptor expression were shown to be important to distinguish a signal sending cell from a signal receiving cell, respectively, between adjacent cells [59]. In chick and mouse developing retina, Dll ligands are preferentially expressed in retinal progenitors [60,61]. We propose that our culture conditions mimic in vivo environment of RSC where Dll3-expressing retinal progenitors preferentially trans-activates Notch-expressing RSC signal receivers to expand the stem cell population while the stepwise downregulation of Dll3 (Fig. 1B) induces the subsequent quiescence of adult RSCs. The fact that exogenous ligands impacts on neither the numbers of PND2 NR-derived primary spheres nor the diameters of adult RSC colonies suggests that PND2 NR progenitors and adult RSC-derived progenitors are more likely to be the Dll-signal sending cells.
A converging piece of evidence is that direct Notch over-activation in single adult RSCs led to a three-fold increase in clonal pigmented secondary colonies compared to control, thus revealing a symmetric expansion of adult RSCs. This result indicates that Notch activity effects on the RSC mode of division rather than producing changes in cell cycle length or proliferation rate. N1ICD-infected adult RSC colonies appeared indeed smaller in size compared to controls suggesting a decrease in progenitor numbers due to decreased asymmetric divisions of the N1ICD-infected RSCs. This hints at another fundamental aspect of a niche, as keeping a perfect balance between stem cells and their downstream progenitors.
The present results demonstrate that increased activation of Notch signaling reverts PND2 NR progenitors back to retinal stem-like state, and promotes symmetrical divisions in adult RSCs, thereby increasing the RSC pool.
Footnotes
Acknowledgments
We thank members of D.v.d.K laboratory for thoughtful discussion and critical reading of the article, and Brenda Coles for her excellent technical assistance. The authors also thank Lilian Doss for her help with the statistics and Stephanie Heyraud for her constructive suggestions on the article. L.B. is supported by Fondation pour la Recherche Medicale and is a recipient of the St. Georges' Society (McEwen Centre for Regenerative Medicine) fellowship. This work was supported by CIHR and the Foundation Fighting Blindness.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.