
Abstract
Mesenchymal stem cells (MSCs) have potent immunosuppressive effects in vitro and are considered as a therapeutic option for autoimmune disease and organ transplantation. While MSCs show beneficial effects on immune disease progression and transplant survival in animal models, the immunomodulatory mechanisms involved are largely unknown. In the present study, we show that intravenously infused C57BL/6- green fluorescent protein (GFP) MSCs home to the lungs in C57BL/6 recipient mice and induce an inflammatory response. This response was characterized by increased mRNA expression of monocyte chemoattractant protein-1 (MCP1), IL1-β, and TNF-α and an increase in macrophages in lung tissue 2 h after MSC infusion. Simultaneously, serum levels of proinflammatory IL6, CXCL1, and MCP1 protein increased, demonstrating systemic immune activation after MSC infusion. In liver tissue, no C57BL/6-GFP MSCs were detected, but MCP1 and TNF-α mRNA levels peaked 4 h after MSC infusion. The expression of the anti-inflammatory cytokines TGF-β, IL4, and IL10 was only marginally affected. Nevertheless, 3 days after MSC infusion, animals developed a milder inflammatory response to lipopolysaccharides. Our results suggest that the in vivo immunomodulatory effects of MSCs originate from an inflammatory response that is induced by the infusion of MSCs, which is followed by a phase of reduced immune reactivity.
Introduction
The immunosuppressive capacity of MSCs has been a subject of extensive research over the last decade. In vitro experiments have demonstrated that MSCs inhibit the proliferation and cytokine secretion of T cells [9], B cells [10], and NK cells [11]. MSCs target lymphocytes via a variety of mechanisms, which depend on both the secretion of soluble anti-inflammatory factors, such as TGF-β, hepatocyte growth factor [3], HLA-G [12], and on cell contact interactions via the inhibitory costimulatory programmed death ligand 1 (PD-L1) pathway [13]. The expression by the human MSC of the tryptophan-depleting enzyme indolamine 2,3-dioxygenase (IDO) is crucial for the inhibition of lymphocyte proliferation [14]. The expression of IDO and other anti-inflammatory factors by MSCs are strongly induced under inflammatory conditions [15 –17], suggesting that the immunomodulatory effects of MSCs act as a feedback mechanism to control ongoing immune responses.
Studies in animal models and early clinical trials have attempted to translate the potent in vitro immunosuppressive effects of MSCs in effective immune therapy. The most widely used route of administration of MSCs in these studies is via intravenous (IV) infusion. In contrast to the in vitro immunomodulatory effects of MSCs, however, little is known about the effects of MSCs after IV infusion. IV-infused MSCs home to the lungs [18 –21] and it is believed that they migrate from there to the sites of injury and inflammation [22,23] and exert their immunosuppressive function. However, recent evidence suggests that this is not the case. It has been shown that living MSCs are only detectable in the lungs after IV infusion, and not in any other tissue or at sites of injury [24,25]. Furthermore, the presence of living MSCs in the lungs is temporary and within 24 h, the majority of MSCs in the lungs die and their cell debris is distributed to other sites, in particular, the liver.
Even though MSCs appear to be short-lived after IV infusion, they induce potent immunomodulatory effects in various disease models. For instance, infusion of MSCs has been shown to alleviate experimental colitis [26], ameliorate experimental encephalomyelitis [27], and inhibit allograft rejection [28]. In these models, infused MSCs are unlikely to be present at the site of inflammation or live long enough to become activated by inflammatory conditions and exert their immunosuppressive effect. Therefore, it is unlikely that the in vitro established mechanisms of immunomodulation by MSCs are operational after infusion of MSCs. Other mechanisms are likely to mediate the effect of MSCs, probably involving other cell types. The purpose of the present study was to map the type, origin, and duration of the immunomodulatory response that is induced upon IV infusion of the MSC.
Materials and Methods
Isolation and culture of MSCs
MSCs were isolated from the adipose tissue of C57BL/6 mice expressing enhanced green fluorescent protein (GFP) under the human ubiquitin C promoter (The Jackson Laboratory). The adipose tissue was a kind gift from E. Koning en Dr. R. Mebius of the Free University Amsterdam. Animals were kept in accordance with the institutional guidelines. The adipose tissue of five animals was pooled, minced with a scalpel knife, and enzymatically digested with 0.5 mg/mL collagenase type IV (Life Technologies) in a minimal essential medium eagle-alpha (MEM-α) supplemented with 2 mM L-glutamine, 100 U/mL penicillin, and 100 mg/mL streptomycin (1% p/s) (all Life Technologies) for 30 min at 37°C under continuous shaking. After centrifugation at 1,200 g for 10 min, the cell pellet was resuspended in a red blood cell lysis buffer (eBioscience) and incubated for 10 min at room temperature. The cells were then washed, resuspended in the MEM-α supplemented with 2 mM L-glutamine, 1% p/s, and 15% fetal bovine serum (FBS; Lonza), and filtered through a 70-μm cell strainer (BD Biosciences). The cells were then transferred to a 175-cm2 cell culture flask (Greiner Bio-One) and expanded in a humidified atmosphere with 5% CO2 at 37°C. Nonadherent cells were removed after 3–4 days. The culture medium was refreshed twice weekly. Cells were removed from culture flasks at subconfluency using 0.05% trypsin-EDTA (Life Technologies) and reseeded at 1,000 cells/cm2 to ensure optimal proliferation. MSCs were used for experiments between passages 4 to 10.
Characterization of MSCs
MSCs were examined for GFP expression by fluorescence microscopy. For immunophenotyping, MSCs were harvested, washed in FACSFlow (BD Biosciences), and stained with Sca-1-PE-Cy7, CD11b-APC, CD44-PE, and CD45-PERCP (all BD Biosciences) and measured on a FACSCanto II flow cytometer (BD Biosciences) using FACSDiva software.
Adipogenic differentiation was induced by culturing confluent MSCs in the MEM-α supplemented with 2 mM L-glutamine, 1% p/s, 15% heat-inactivated FBS (FBS-HI), 50 μg/mL L-ascorbic acid (Sigma-Aldrich), 500 μM 3-isobutyl-1-methylxanthine (IBMX; Sigma-Aldrich), 60 μM indomethacin, 2 μM insulin (Sigma-Aldrich), and 10 nM dexamethasone (Sigma-Aldrich) for 14 days. Lipid-filled vesicles were detected by Oil Red O staining. Cells were washed with phosphate-buffered saline (PBS), fixed with 60% isopropanol for 1 min, and incubated with filtered 0.3% Oil Red O (Sigma-Aldrich; in 60% isopropanol) for 30 min. Following three washes with PBS, the cells were photographed.
Osteogenic differentiation was induced by culturing confluent MSCs in the MEM-α supplemented with 2 mM L-glutamine, 1% p/s, 15% FBS-HI, 5 mM β-glycerophosphate (Sigma-Aldrich), 50 μg/mL L-ascorbic acid, and 10 nM dexamethasone for 21 days. The deposition of calcified nodules was identified using von Kossa staining. Cells were washed with PBS and fixed with cold 4% paraformaldehyde for 5 min. Following an additional wash step with PBS and two wash steps with distilled water, cells were incubated with 1% silver nitrate (in water) on a light box until blackening occurred. Cells were washed three times with water, incubated with 2.5% sodium thiosulfate (in water) for 5 min, washed again twice with water, and photographed.
For examination of the immunosuppressive capacity of MSCs, Swiss mouse splenocytes were stimulated with 5 μg/mL Concanavalin A (ConA; Sigma Aldrich) and MSCs added at ratios of 1:5 to 1:640 in 96-well plates. After 3 days, proliferation was measured by incorporation of 3H-thymidine (0.25 μCi/well; PerkinElmer) during 8 h of incubation using a Wallac 1450 MicroBeta TriLux Liquid Scintillation Counter & Luminometer (PerkinElmer).
Infusion of MSCs
Male C57BL/6 mice, 8–10 weeks old and weighing ∼25 g, were obtained from Harlan. The experimental protocol was approved by the Animal Experiments Committee under the Dutch National Experiments on Animals Act and complied with the 1986 directive 86/609/EC of the Council of Europe. MSCs were trypsinized, washed, and resuspended in PBS. Three hundred thousand MSCs in 200 μL PBS were infused via tail vein injections. Control mice received 200 μL PBS only. After 2, 4, 20, 68, and 216 h, mice were sacrificed and blood collected in Minicollect serum separation tubes (Greiner Bio-One). Two experiments were carried out, totaling five animals per time point. Lungs, livers, and kidneys were removed and frozen at −80°C. Spleens were removed and cell suspensions prepared. After red blood cell lysis (RBC lysis buffer; eBioscience), splenocytes were frozen in 10% DMSO at −150°C.
Treatment with lipopolysaccharides
One day or 3 days after infusion of MSCs as described above, animals were injected with 5 mg/kg body weight lipopolysaccharides (LPS; Sigma-Aldrich) dissolved in PBS via the tail vein. Animals were sacrificed 4 h later and blood collected in Minicollect serum separation tubes.
PCR analysis
Frozen lung, liver, and kidney tissue were sectioned in 20-μm slices and RNA isolated using the Trizol reagent (Life Technologies). cDNA was synthesized from 500 ng RNA with random primers (Promega). Quantitative gene expression was determined using TaqMan Gene Expression Master Mix (Life Technologies) and assays-on-demand for IL1-β (mm01336189.m1), IL10 (mm00439614.m1), CCL2 [monocyte chemoattractant protein-1 (MCP1)] (mm00441242.m1), CCL3 (MIP1-α) (mm00441258.m1), TNF-α (mm00443258.m1), IFN-γ (mm01168134.m1), TGF-β (mm01178820.m1), IL4 (mm00445259.m1), CD68 (mm03047340.m1), F4/80 (mm008002529.m1), Ly6G6c (mm00458275.m1), and housekeeping gene HPRT (mm01545399.m1) (all Applied Biosystems) on a StepOnePlus (Applied Biosystems). GFP expression was determined by SYBR green-based (Applied Biosystems) RT-PCR. Primer sequences used for the detection of GFP were 5′-CCA CAT GAA GCA GCA GGA CTT-3′ and 5′-GGT GCG CTC CTG GAC GTA-3′ (BaseClear). Results were expressed as a ratio to HPRT.
Analysis of serum proteins by Milliplex
Serum concentrations of a broad panel of chemokines, cytokines, and acute-phase proteins were measured by Milliplex cytokine/chemokine panel I (Millipore) according to the manufacturer's instructions. The panel existed of the following proteins: CRP, CXCL1, CXCL5 (LIX), CCL11 (Eotaxin), G-CSF, GM-CSF, IFN-γ, IL1-α, IL1-β, IL2, IL3, IL4, IL5, IL6, IL7, IL9, IL10, IL12p40, IL12p70, IL13, IL15, IL17, IP10, LIF, CCL2 (MCP1), M-CSF, CCL3 (MIP1-α), MIP1-β, MIP2, MIG, Rantes, SAP, TNF-α, and VEGF.
Splenocyte analysis
Splenocytes were thawed and stimulated with 5 ng/mL PMA (Sigma-Aldrich) and 500 ng/mL ionomycin (Sigma-Aldrich) for 4 h at 37°C in the presence of 1 μl Golgiplug (BD Biosciences) per 1×106 cells. Controls were not stimulated with PMA/ionomycin. The cells were then washed and stained with CD3-PERCP, CD4-FITC, and CD25-PE-Cy7 antibodies (all BD Biosciences). Subsequently, cells were washed and incubated in a Fixation/Permeabilization solution (BD Biosciences) for 30 min at 4°C, followed by a wash in the Perm/Wash buffer (BD Biosciences), and stained with the IFN-γ-APC antibody (BD Biosciences). After washing, the cells were analyzed on a FACSCanto II flow cytometer using FACSDiva software.
To determine the effect of the MSC on the proliferative capacity of splenocytes after LPS treatment, splenocytes were collected 4 h after LPS treatment and seeded at 150,000 cells per well in a 96-well round-bottom plate and stimulated with 5 μg/mL ConA for 3 days. They were then incubated with 3H-thymidine (0.25 μCi/well) for 8 h. 3H-thymidine incorporation was measured using the Wallac 1450 MicroBeta TriLux Liquid Scintillation Counter & Luminometer.
Results
Characterization of adipose tissue-derived MSCs
MSCs were isolated from the adipose tissue of C57BL/6 mice expressing GFP under the ubiquitin promoter. In culture, MSCs displayed a characteristic fibroblast-like morphology (Fig. 1A) and showed strong GFP expression (Fig. 1B). Flow cytometric analysis confirmed GFP expression in close to 100% of MSCs and demonstrated positive expression of the MSC markers CD44 and Sca-1 (Fig. 1C). None of the cells expressed the hematopoietic markers, CD11b and CD45, indicating the absence of macrophages and dendritic cells in the cultures. Culture under osteogenic and adipogenic differentiation-inducing conditions stimulated the production of calcified nodules (Fig. 1D) and lipid-filled vesicles (Fig. 1E) by MSCs. Finally, MSCs were capable of inhibiting the proliferation of ConA-stimulated splenocytes in a dose-dependent manner (Fig. 1F). These results demonstrate that the MSCs used in this study were fully functional.
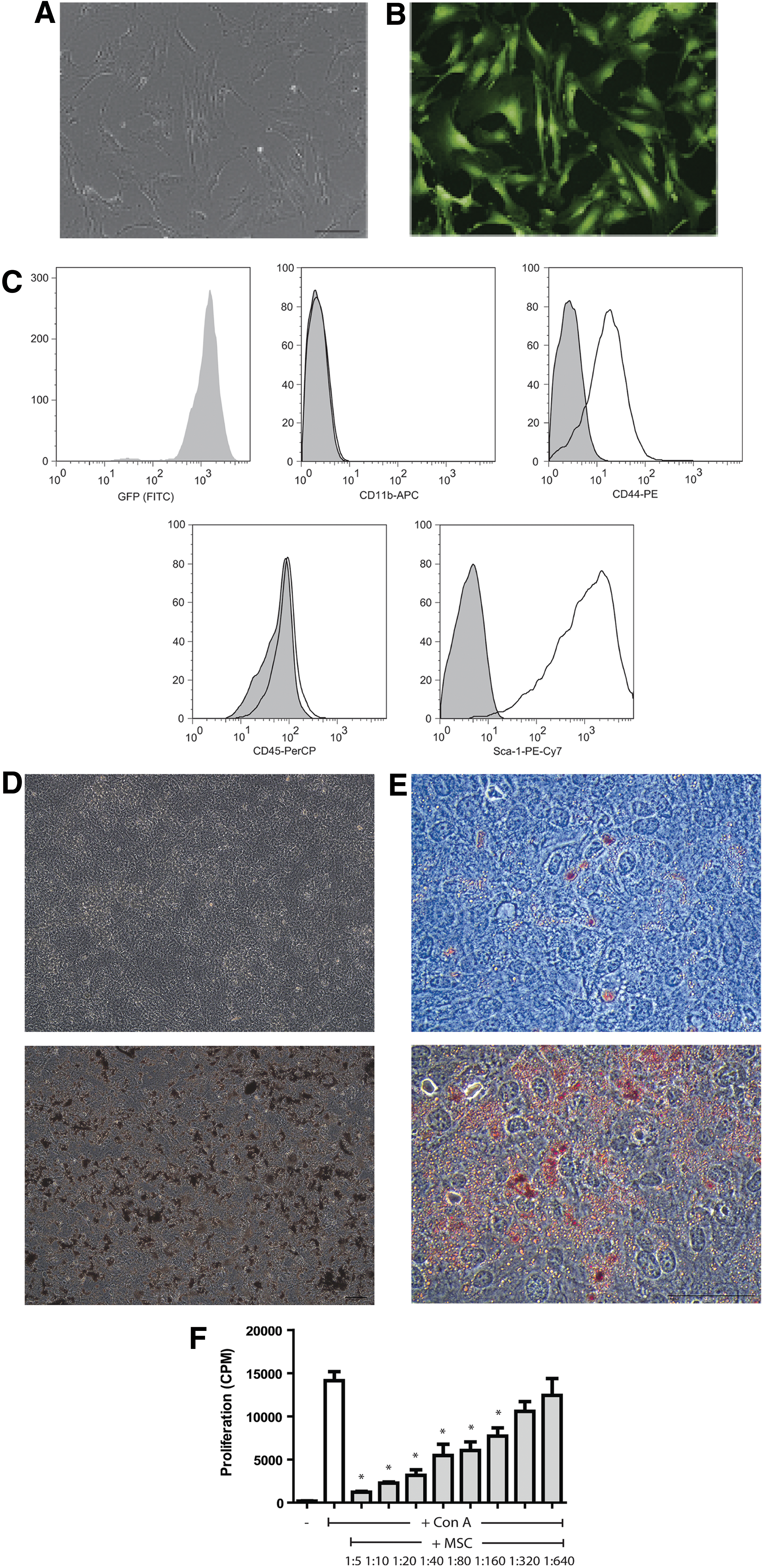
Characterization of MSCs isolated from adipose tissue of C57BL/6 mice expressing GFP under the ubiquitin promoter.
Effects of IV infusion of MSCs on lung tissue
Three hundred thousand GFP-MSCs were infused via the tail vein in syngeneic C57BL/6 mice. At 2, 4, 20, 68, and 216 h after infusion, five mice were sacrificed and lungs removed for analysis. MSCs could be detected in the lungs 2 h after infusion by means of GFP mRNA expression (Fig. 2A). GFP expression in the lungs subsequently decreased. The presence of MSCs in the lungs was accompanied by an inflammatory response, which peaked at 2 h after infusion of MSCs and was characterized by increased mRNA levels of the proinflammatory chemokines and cytokines CCL2 (MCP1; 32-fold), CCL3 (MIP1-α; 2-fold), IL1-β (5-fold), and TNF-α (2-fold) (Fig. 2B–E). Simultaneously, there was a twofold increased expression of anti-inflammatory TGF-β (Fig. 2F). All gene expression levels returned to basal levels at 4 to 20 h after MSC infusion. The expression of IL4, IL10, and IFN-γ did not significantly change upon administration of MSCs (data not shown).

Gene expression changes in lung tissue in response to IV infusion of MSCs. Lungs were removed at various time points after administration of MSCs and mRNA isolated. Expression of GFP, CCL2 (MCP1), CCL3 (MIP1-α), IL1-β, TNF-α, and TGF-β is depicted as a ratio to HPRT.
MSCs increase the number of macrophages in the lungs
To investigate whether the inflammatory response in the lungs induced by the infusion of MSCs resulted in increased numbers of macrophages and neutrophils in the lungs, PCR analysis for the macrophage markers CD68 and F4/80 and for the neutrophil marker Ly6G6c was performed. There was an increase in CD68 and F4/80 expression in the lungs 2 h after infusion of MSCs (Fig. 3A, B). At 20 h, expression levels had returned to basal levels. There was no significant change in Ly6G6c, although at 2 h, there was a trend for increased expression (Fig. 3C).
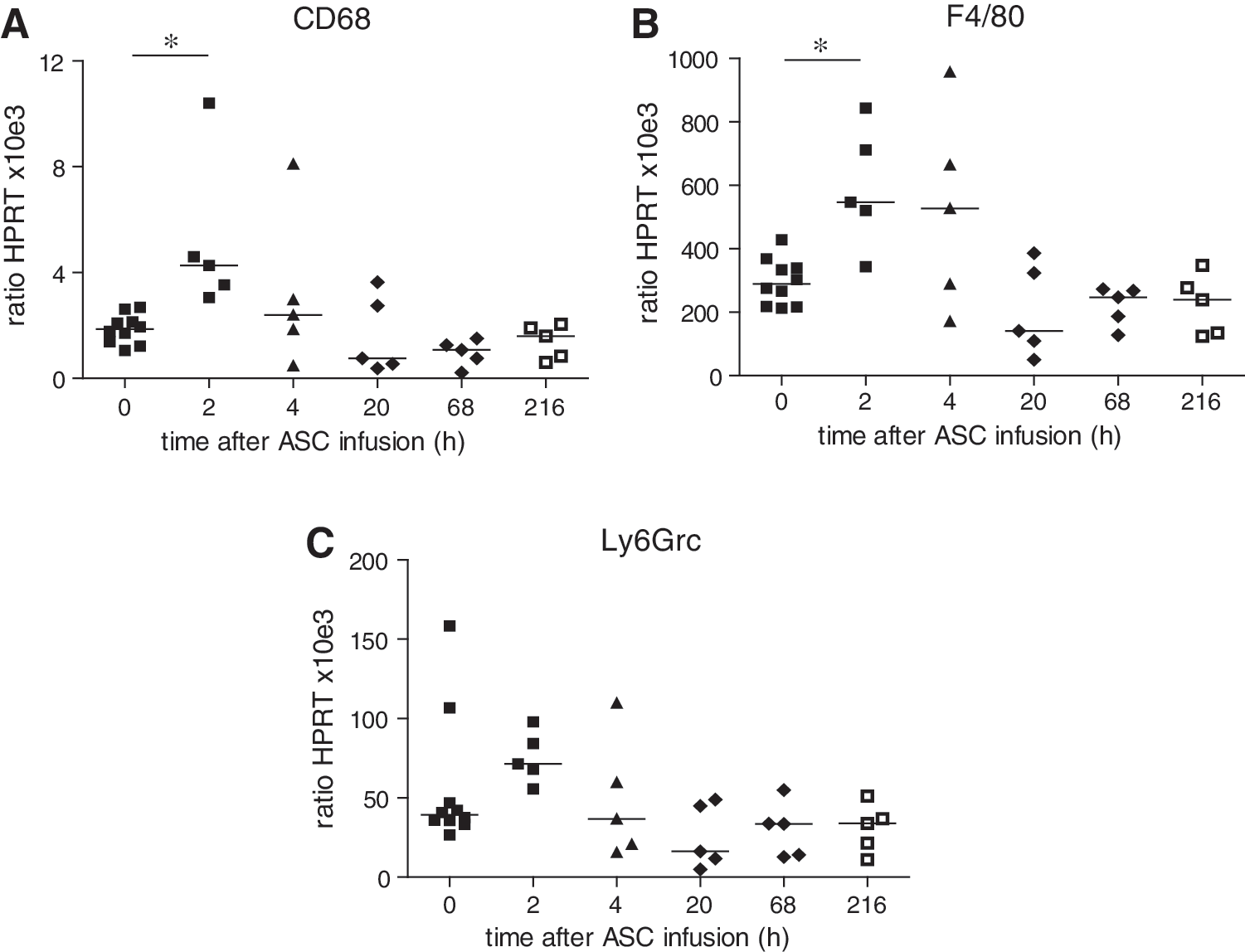
Expression of macrophage and neutrophil markers in lung tissue in response to IV infusion of MSCs.
Systemic effects of IV infusion of MSCs
To analyze whether the inflammatory response in the lungs evoked by MSC infusion was also detectable systemically, levels of 34 cytokines, chemokines, and acute-phase proteins were measured in serum at the various time points after administration of MSCs. Two hours after infusion of MSCs, there were increases in the concentrations of the neutrophil stimulator G-CSF (4-fold), proinflammatory IL6 (18-fold), the neutrophil chemoattractant CXCL1 (10-fold) and CCL2 (MCP1, 4-fold) (Fig. 4A–D). Concentrations of the proteins returned to basal levels at 20 h after administration of MSCs. In contrast, levels of the neutrophil chemoattractant, CXCL5, dropped significantly 2 h up to 20 h after infusion of MSCs, while the eosinophil chemoattractant, CCL11, was decreased at 20 h (Fig. 4E, F). No significant changes in serum concentrations of CRP, GM-CSF, IFN-γ, IL1-α, IL1-β, IL2, IL3, IL4, IL5, IL7, IL9, IL10, IL12p40, IL12p70, IL13, IL15, IL17, IP10, LIF, M-CSF, CCL3 (MIP1-α), MIP1-β, MIP2, MIG, Rantes, SAP, TNF-α, and VEGF were detected (data not shown).
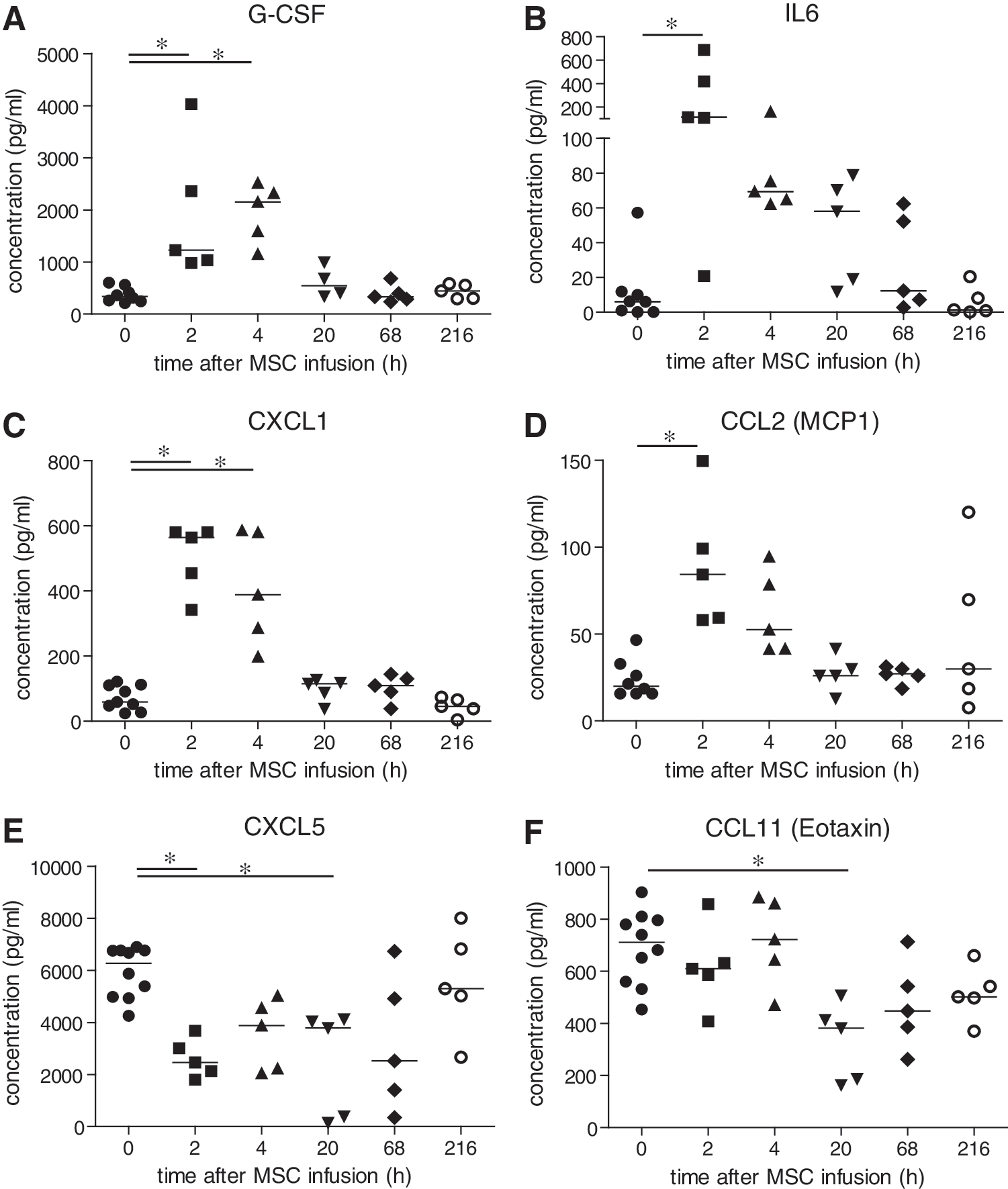
Serum concentrations of cytokines/chemokines after IV infusion of MSCs. Serum was collected at various time points after administration of MSCs. Protein concentrations of G-CSF, IL6, CXCL1, CCL2 (MCP1), CXCL5, and CCL11 (Eotaxin) were determined by Milliplex assay.
Effects of IV infusion of MSCs on liver and kidney tissue
Earlier, we found that the MSC label, but no viable MSCs, accumulated in liver tissue 1 day after infusion of MSCs [29]. To examine whether this would leave an immunological footprint in liver tissue, livers were removed for analysis 2, 4, 20, 68, and 216 h after MSC infusion and mRNA expression of a number of cytokines and chemokines analyzed. No expression of GFP was detected in the liver at any time after MSC infusion, confirming that no viable MSCs localized to the liver (data not shown). Expression of TNF-α, CCL2 (MCP1), and CCL3 (MIP1-α) increased 2 h and peaked 4 h after infusion of MSCs, showing twofold, fourfold, and twofold increases, respectively (Fig. 5A–C). IFN-γ and IL1-β levels also peaked, but at 68 and 216 h after MSC infusion, respectively (Fig. 5D, E). TGF-β showed a 1.5-fold increase after 4 h (Fig. 5F), while no changes in IL10 expression were observed (data not shown). In kidney tissue, no significant changes in cytokine and chemokine gene expression were detected at any time point after infusion of MSCs and no expression of GFP was found (data not shown).
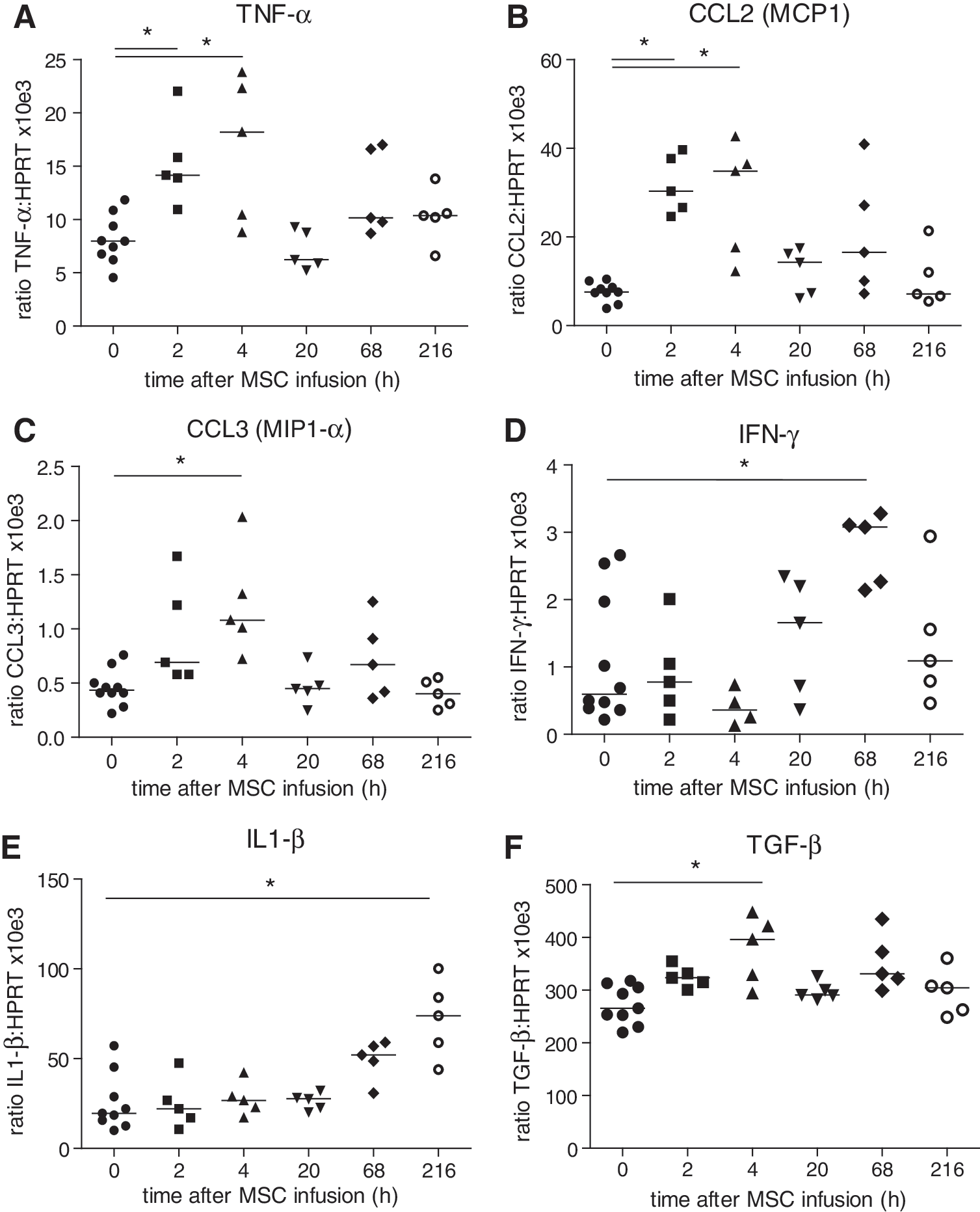
Gene expression changes in liver tissue in response to IV infusion of MSCs. Livers were removed at various time points after administration of MSCs and mRNA isolated. Expression of IFN-γ, CCL2 (MCP1), CCL3 (MIP1-α), IL1-β, TNF-α, and TGF-β is depicted as a ratio to HPRT.
Effect of IV infusion of MSCs on splenocytes
The effect of MSC infusion on the responsiveness of splenocytes was determined by in vitro stimulation of splenocytes with PMA/ionomycin and measurement of their IFN-γ production. PMA/ionomycin induced an IFN-γ response in ∼6% of CD3+ and CD3− splenocytes of control animals. The IFN-γ response in CD3+ splenocytes was not significantly different in animals that received MSCs, although there was a trend for a reduced IFN-γ response at 2, 4, 20, and 68 h after administration of MSCs (Fig. 6A). The IFN-γ response in CD3− splenocytes was not affected by MSC treatment (Fig. 6B).
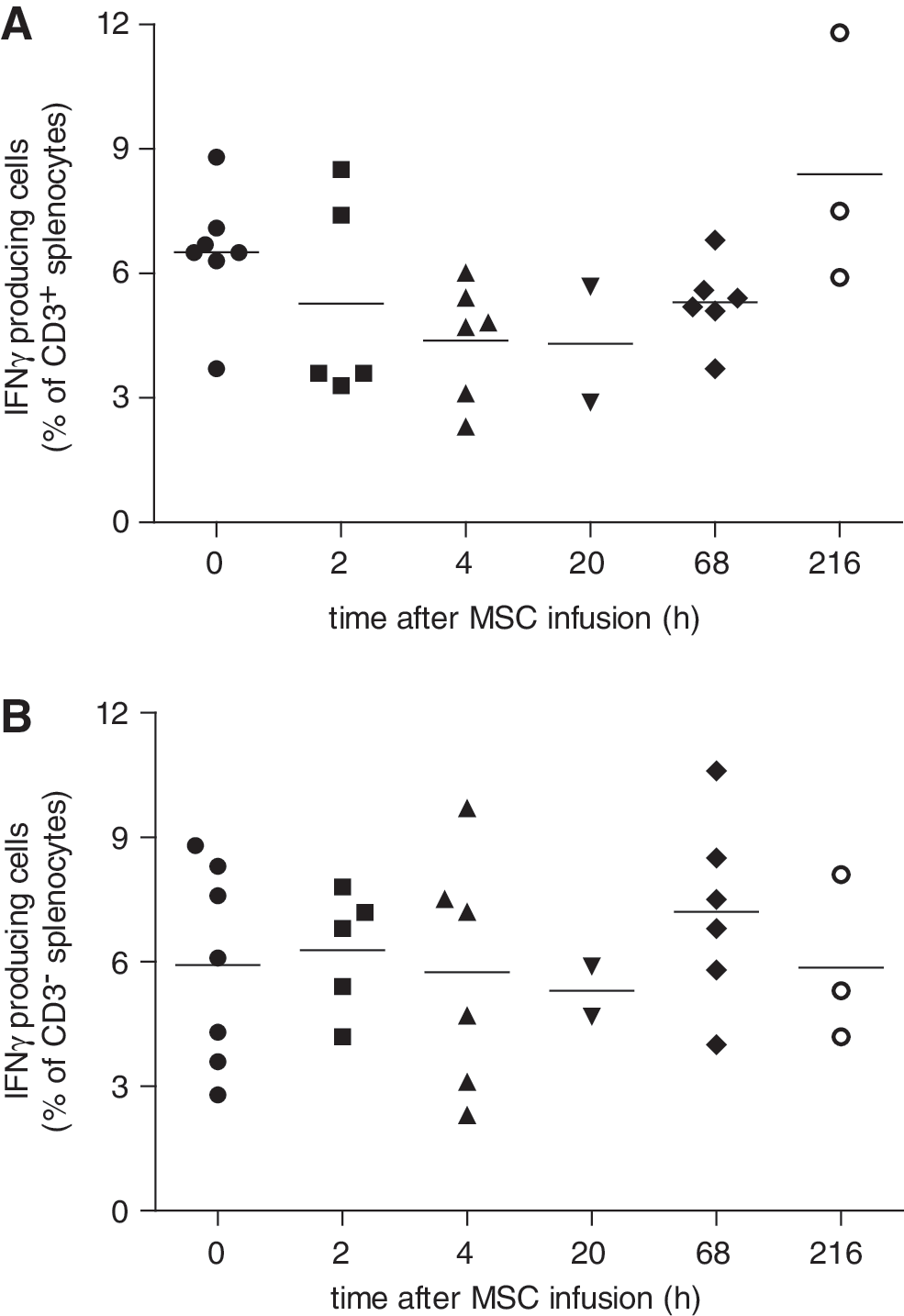
In vitro IFN-γ response of splenocytes isolated at various time points after IV infusion of MSCs. Splenocytes were stimulated for 4 h with PMA/ionomycin and intracellular IFN-γ production measured by flow cytometry. The percentage of IFN-γ producing cells of CD3+
Infusion of MSCs dampens the immune response to LPS
To examine whether the early immune activation induced by the infusion of MSCs would lead to an immunomodulatory effect in vivo later on, animals received 5 mg/kg LPS at day 1 or 3 after administration of MSCs or PBS and were sacrificed after 4 h. LPS induced major increases in serum concentrations of multiple cytokines and chemokines, including G-CSF, IL6, CXCL1, MCP1, IP10, and IL12p40. Infusion of MSCs 1 day before administration of LPS had no effect on the serum levels of these factors (data not shown). However, infusion of MSCs 3 days before LPS significantly reduced the increases in G-CSF, IL6, CXCL1, IP10, and IL12p40, suggesting a dampened immune response to LPS (Fig. 7A–F).
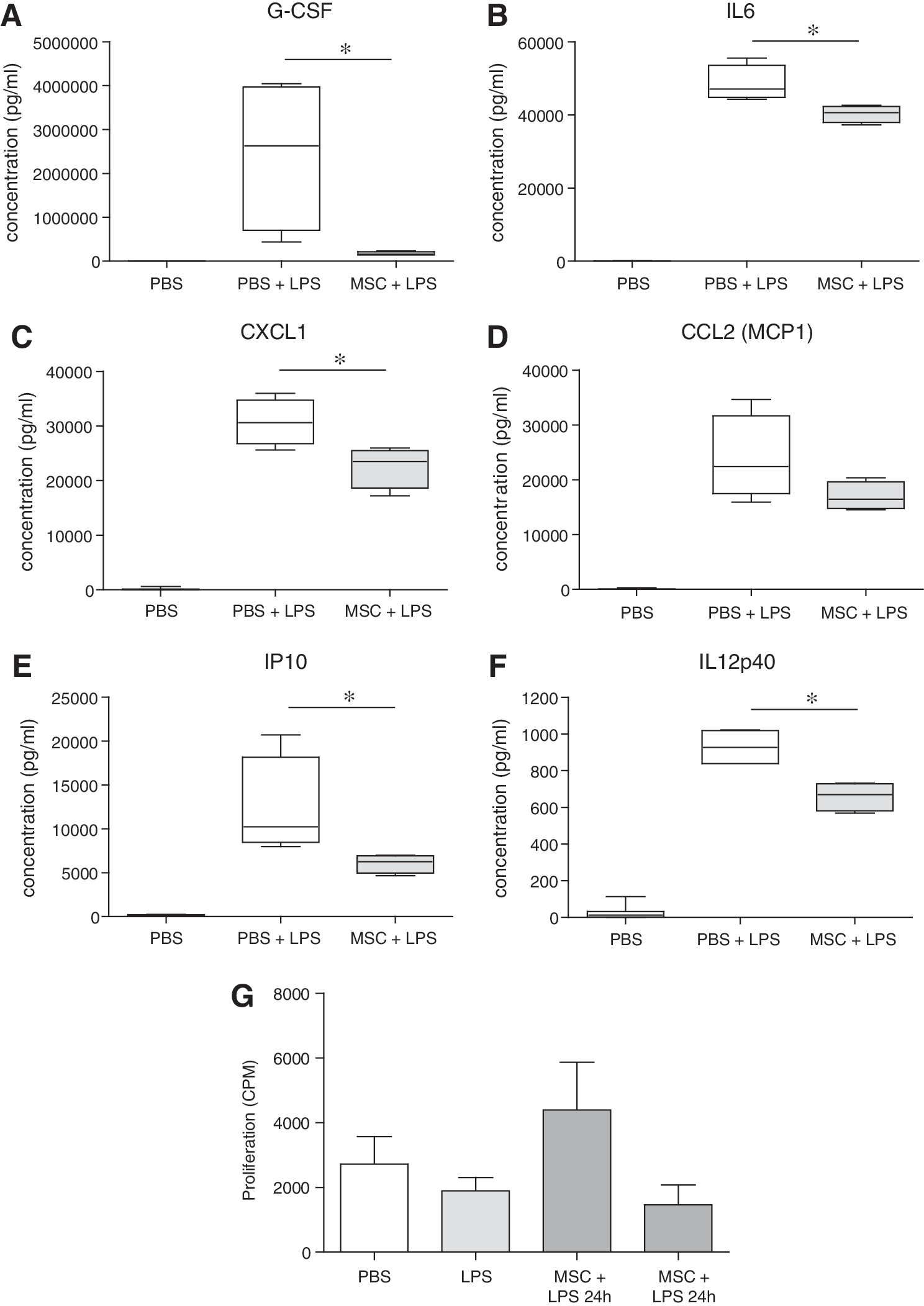
Effect of MSC infusion on the response to LPS. Mice received 5 mg/kg LPS 3 days after IV infusion of 300,000 MSCs or phosphate-buffered saline. Four hours later, mice were sacrificed and serum and splenocytes collected.
To examine their proliferative response, splenocytes were stimulated with ConA. Splenocytes from animals that were treated with MSCs 24 h before LPS tended to show an increased proliferative capacity, whereas splenocytes from animals that were treated with LPS 72 h earlier tended to show reduced proliferation, although the differences were not significant (Fig. 7G).
Discussion
There is increasing interest in the use of MSC therapy for immunomodulatory purposes. In contrast to pharmacological immunosuppressants, which need to be taken lifelong and which suppress specific molecular pathways, MSC therapy is thought to be able to achieve long-term effects by modulating the function of various cells of the immune system. Although beneficial effects of MSCs have been demonstrated in inflammatory disease and transplant models, the mechanisms of immune modulation by MSCs after in vivo administration remain obscured. Understanding of these mechanisms is essential for the development of efficient MSC therapy.
Surprisingly, in the present study, we found that infusion of syngeneic MSCs induced an inflammatory response, which was detectable systemically and in lung and liver tissue. No inflammatory response was evident in the kidneys. The response in lung tissue was characterized by increased expression of monocyte and neutrophil chemoattractants and proinflammatory cytokines. In addition, we observed an increase in the expression of the macrophage markers, CD68 and F4/80, suggesting that intravenous infusion of MSCs causes an influx of macrophages in the lungs. We and others have described that MSCs interact with macrophages and induce macrophages with an immunomodulatory phenotype [30]. Whether macrophages that are recruited to the lungs upon infusion of MSCs are adapting a regulatory function will need further investigation.
The question rises whether the administered MSCs themselves were the source of the proinflammatory cytokine response. Gene expression changes in lung tissue coincided with the presence of MSCs and the genes that showed significant upregulation in the lung encode proteins that are expressed by MSCs. Nevertheless, a more likely possibility is that MSCs trigger the expression of proinflammatory factors in resident cells of the lungs. MSCs are shown to accumulate in the microvasculature after infusion due to space restriction [31], where they mechanically interact with endothelial cells and leukocytes. The encounter of MSCs with lung endothelial cells and cells of the immune system may evoke the inflammatory response.
Our results strongly suggest that the increased expression of proinflammatory factors in liver tissue did not directly derive from MSCs. At the time the increases were observed, 4 h up to 216 h after MSC infusion, no MSCs were detected in the liver, indicating that liver resident cells are responsible for the gene expression changes. It is possible that the initial inflammatory response in the lungs is transferred via circulating chemokines and cytokines, which trigger a response in the liver. Another possibility is that mobile cell types like macrophages are activated by MSCs and transfer the signal from the lung to the liver via the blood stream. Some of the systemic changes in the levels of chemokines and cytokines are also unlikely to be a direct effect of MSCs. We observed a significant increase in G-CSF levels in serum after MSC infusion, but MSCs do not produce G-CSF [32]. Furthermore, MSCs decreased serum levels of CXCL5 and CCL11, showing that MSCs inhibit the production of these chemokines by other cells.
Although it seems that MSCs elicit a response of the immune system after infusion, at the moment, we can only speculate about the trigger that induces this response. As we used syngeneic MSCs, an alloresponse can be excluded. The infused MSCs, however, expressed GFP, whereas the recipients did not, but as GFP is an intracellular protein, it is unlikely to evoke an immune response as rapidly as was observed. It is possible that the immune system responds to infused MSCs as the cells are culture generated and differ in size and protein expression pattern from noncultured MSCs [33]. In addition, under normal conditions, MSCs are present in the circulation at extreme low numbers [34,35] and the high numbers present after MSC administration may activate a clearance response. It is thus likely that the immune reaction induced after infusion of MSCs is a response to cells that are seen as foreign and/or are present at an unusual location, rather than a MSC-specific response. Other cell types may well induce a similar response. We have not set out to test this in the present study.
How the immunoactivating effects of MSC administration lead to beneficial immunomodulatory effects in disease models remain unclear. The immune responses, which are evoked by MSCs, may result in an immunosuppressed status. In line with this hypothesis, we observed that the systemic inflammatory response to LPS was reduced 3 days after infusion of MSCs. We were not able to demonstrate reduced in vitro responsiveness of splenocytes from animals that received MSCs 3 days before LPS. In fact, splenocytes from animals that received LPS did not respond more potently to in vitro ConA stimulation than splenocytes from PBS control animals. This observation could relate to the fact that the ConA stimulation assay takes 3 days, after which the effect of LPS treatment of the animals may have been washed out.
In line with our data, organ transplant models have demonstrated that infusion of MSCs 7 to 4 days before transplantation was effective in prolonging transplant survival, whereas administration at later time points was not [28,36]. In contrast, in models for arthritis [37] and colitis [38], administration of MSCs after inflammatory disease onset was shown to be effective. There may be differences in the effect of MSCs under inflammatory and homeostatic conditions. First, cytokine footprints are different under these conditions and MSCs will have a different modulating effect, and, second, MSCs, which are exposed to inflammatory conditions, become more immunosuppressive in vitro [15,16] and in vivo [39] and may induce a different response upon infusion. Further research should clarify which conditions are optimal for effective MSC therapy for various applications.
In summary, intravenous infusion of syngeneic MSCs in mice triggers a systemic inflammatory response, which originates in the lungs and proceeds systemically. This response may lead to an immunosuppressed status in a few days via feedback mechanisms. Even though MSCs are present for only a few hours, their effects last for much longer and this offers possibilities to use MSCs for immunomodulatory therapy.
Footnotes
Acknowledgments
We are grateful to Eelco Koning and Dr. Reina Mebius of the Department of Molecular Cell Biology and Immunology of the Free University Amsterdam for the supply of adipose tissue from GFP-expressing mice. We kindly thank Leonie van der Zee of the Department of Internal Medicine of the Erasmus MC for her help with the Luminex measurements.
Author Disclosure Statement
No competing financial interests exist.