
Abstract
While most studies have suggested multipotential stromal cell or mesenchymal stem cell (MSC) therapies are useful for immune-mediated diseases, MSCs' immunomodulatory effects were not entirely reproduced in some studies, indicating the necessity to determine the underlying mechanism of MSCs' effects on immune response regulation to maximize their immunomodulatory effects. We have identified the transcription factor early growth response gene-2 (EGR2) as a novel molecular switch regulating known immunomodulatory molecules in human MSCs. EGR2 binds to the promoter regions of these genes, interleukin-6 (IL6), leukemia inhibitory factor (LIF), indoleamine dioxygenase-1 (IDO1), and cyclooxygenase-2/prostaglandin-endoperoxide synthase 2 (COX2/PTGS2), and siRNA against EGR2 was shown to downregulate these genes and reduce the production of prostaglandin E2, an immunomodulatory mediator produced downstream of COX2/PTGS2. Moreover, EGR2 knockdown restores T-lymphocyte proliferation reduced by MSC coculture. Therefore, EGR2 is a potential target for the optimization of immunomodulatory properties of MSC-based therapies.
Introduction
T
With their multidifferentiation potential, cellular therapeutics with adult multipotential stromal cells or mesenchymal stem cells (MSCs) has been gaining attention in the field of reparative and regenerative medicine [4 –10]. Initially, MSC differentiation and direct incorporation into repairing or regenerating tissues was thought to be a primary mechanism of MSC action; however, it is now widely accepted that MSC's strong paracrine capability of various growth factors and cytokines such as vascular endothelial growth factor (VEGF) plays a key role in MSC-mediated wound healing and tissue regeneration [11 –14].
Moreover, MSCs have strong immunomodulatory or anti-inflammatory properties [15 –17], and MSC-based therapeutics is also promising against immune-mediated diseases such as GVHD. Interestingly, several studies suggested the feasibility of xenotransplantation of human MSCs into immune-competent mice, due to their immunoprivileges and immunomodulatory effects [18 –22]. The key intercellular molecules, such as indoleamine dioxygenase-1 (IDO1) or prostaglandin E2 (PGE2), which mediate human MSCs' strong anti-inflammatory properties, have been identified [15 –17]; however, the key intracellular molecular switches facilitating MSCs' strong immunomodulatory properties are still largely unknown.
One major issue surrounding cell-based therapeutics is that beneficial results of stem/progenitor cell therapeutics in initial small-scale clinical studies have not been reproduced by subsequent randomized-controlled trials. For example, in a large-scale, placebo-controlled phase III clinical trial with third-party MSCs as first- and second-line therapies to treat GVHD and steroid-resistant GVHD, MSCs were shown to be no more effective than placebo [16,23]. These data strongly indicate the urgent need of further optimization of cell-based therapy [15,24]. The immunomodulatory effects also depend on the culture conditions [25], and it is imperative to understand the molecular mechanism supporting MSCs' strong immunoregulatory properties to maximize them for therapeutics.
Our previous work has shown that early growth response genes-1, -2, and -3 (EGR1–3) are expressed in MSCs at much higher levels than in human dermal fibroblasts (HDFs), another type of cells akin to MSCs, but with reduced differentiation and paracrine capability [26,27]. EGRs encode a zinc finger transcription factor (TF) whose gene expression is upregulated in response to various growth factors and cytokines such as the epidermal growth factor (EGF) or interleukin-1 beta (IL1β) [26 –29]. Biological roles of EGRs in MSCs have not been addressed except in our studies [26,27].
EGR2 was identified as a key regulator of myelination in the nervous system and of hindbrain development [30 –32] and as a negative regulator of T-cell activation [33]. However, the role of EGR2 in MSCs has not been investigated. In this study, we show that MSCs upregulate immunomodulatory molecules in response to IL1β and interferon-γ (IFNγ) treatment, and EGR2 serves as an intracellular molecular switch for MSCs' immunomodulatory properties by regulating expression of these genes.
Materials and Methods
This study was approved by the IBC committee of the University of Pittsburgh for recombinant DNA research and biohazard agents in obedience to the National Institutes of Health Guidelines for Research Involving Recombinant DNA Molecules.
Cell culture
Human telomerase reverse transcriptase—immortalized human bone marrow MSCs (one donor derived, originally obtained from BioWhittaker, currently known as Lonza) were generously gifted by Dr. Junya Toguchida (Kyoto University) [34] and were cultured in the DMEM supplemented with 2 mM
Quantitative reverse transcription–polymerase chain reaction
The level of mRNA transcripts was assessed by two-step quantitative reverse transcription–polymerase chain reaction (qRT-PCR) using TaqMan® PCR. Total RNA was harvested with TRIzol® Plus RNA Purification System according to the manufacturer's instruction. After treatment with DNase and a subsequent DNase neutralization step with the TURBO DNA-free™ Kit, cDNA was synthesized with High Capacity RNA-to-DNA kit. Then, cDNA was mixed with TaqMan Gene Expression MasterMix and each TaqMan gene-specific probe/primer and the following PCR reaction was run using an Mx3005P (Stratagene) as a quantitative real-time PCR thermal cycler: 95°C for 3 min and then, 40 cycles of two temperature PCR at 95°C for 15 s for denaturing, 60°C for 30 s for annealing and extension. The TaqMan probes/primers used in this study are listed in Table 1. TaqMan probe/primers for human ribosomal protein, large, P0 (RPLP0) (4333761F) were used as an endogenous invariant control (housekeeping gene). All reagents for qRT-PCR were from Life Technologies unless otherwise stated.
All PCR primers were designed to span intron(s) to discriminate cDNA amplicons from possible genomic amplicons. Amplification efficacy of each PCR reaction was assessed initially with serial dilution of control samples; those fell into 95%–105%. The comparative Ct method was utilized to assess the levels of each mRNA transcript relative to that level of RPLP0 mRNA transcript.
Transient gene knockdown with siRNA
Anti-EGR2 siRNA (clone SI04948762) and Allstars Negative Control siRNA were purchased from Qiagen. Cells were transfected with 60 nM of siRNA using HiPerFect® Transfection Reagent (Qiagen) according to the manufacturer's instructions.
Quantitation of secreted interleukin-6, leukemia inhibitory factor, and PGE2
After rinsing with phosphate-buffered saline (PBS), MSCs were cultured in a 0.1% FBS-containing cell culture medium for 4 or 16 h during treatment with IL1β (10 ng/mL) and 20 nM IFNγ (20 ng/mL) (Peprotech). The conditioned media were collected and secreted interleukin-6 (IL6), leukemia inhibitory factor (LIF), and PGE2 were measured by quantitative ELISA kits according to each manufacturer's instructions (for IL6 and PGE2; Cayman Chemical) (for LIF; eBioscience). The readouts were adjusted per 1×106 cells seeded initially.
Flow cytometry
All flow cytometry was carried out using a BD FACSCalibur flow cytometer (BD Biosciences). The flow cytometric data were analyzed using WinMDI 2.9 (The Scripps Institute).
Surface marker evaluation of T lymphocytes after in vitro expansion
After isolation and in vitro expansion of T lymphocytes, the cells were immunostained using PE anti-human CD3, FITC anti-human CD4, and Alexa Fluor® 647 anti-human CD8 antibodies (Biolegend) to confirm surface marker expression.
Primary T-lymphocyte proliferation assays
Effects of MSC coculture on primary T-lymphocyte proliferation were assessed through flow cytometry using the Click-iT® EdU Alexa Fluor 488 kit (Life Technologies). Following experimental treatment, cells were incubated for 2 h with the Click-iT EdU reagent (10 μM). Cells undergoing active DNA synthesis incorporated thymidine analog 5-ethynyl-2′-deoxyuridine (EdU) during this pulse labeling. After sample fixation and permeabilization, the ethynyl moiety of incorporated EdU was covalently attached to the Alexa Fluor 488 azide in the presence of copper ions (Click-iT EdU detection reaction), and the fraction of EdU incorporated cells were quantitated by flow cytometry.
IDO1 quantitation within MSCs
After sample fixation and permeabilization, intracellular IDO1 was stained by using the Human IDO Alexa Fluor 488 Monoclonal antibody (R&D Systems) and quantitated by flow cytometry.
In silico analysis of putative EGR2 binding sites
Within −10 and +3 Kb of the transcription start site of the reference sequence of the target genes, putative promoter regions were defined as H3K4me3 chromatin immunoprecipitation (ChIP)-sequencing enriched regions (layered from seven different cell lines; GM12878, H1-hESC, HSMM, HUVEC, K562, NHEK, and NHLF), which were released by the Broad/MGH Encyclopedia of DNA Elements (ENCODE) group on the UCSC Genome Browser (GRCH37/hg19) [36,37]. We interrogated those regions by using the MATCH function of the TRANSFAC database released in April 2012 (BIOBASE Biological Databases) for the possible presence of transcription factor binding site matrices (V$EGR1_01 and/or V$KROX_Q6) with cutoff settings minimizing false negatives.
Chromatin immunoprecipitation qRT-PCR
Direct binding of EGR2 to putative promoter regions was evaluated using ChIP-qPCR. Samples were prepared as per the EpiTect® ChIP One Day Kit (Qiagen), with some modifications. In brief, after indicated treatment, the cell monolayer was crosslinked by using 1% formaldehyde for 10 min. Cells were harvested using PBS supplemented with a protease inhibitor cocktail, and the cell membrane was disrupted by the lysis buffer provided with the kit. Chromatin was sheared using seven rounds of eight 2 s on–15 s off cycles with a W-225R model Sonicator Cell Disrupter with 1/16 inch tapered microtip (QSonica, formerly Heat Systems Ultrasonics/Misonix) to produce sheared chromatin DNA of 1–3 Kb. The lysates were precleared using PureProteome™ Protein G Magnetic Beads (EMD Millipore) and incubated with rabbit control IgG (ab46540; Abcam) or anti-Egr2 (PRB-236P; Covance) antibodies at 0.5 μg/mL for immunoprecipitation. After washing the immunoprecipitated samples, a 10% Instagene™ Matrix (BioRad) chelex bead slurry and 20 μg of Proteinase K were used for DNA isolation [38].
Among predesigned SYBR green-based PCR primers covering between +10 and −20 Kb of TSS of genes (EpiTect ChIP-qPCR Primers; Qiagen), we chose the primers covering predicted EGR binding sequences, as described in detail in the Results section. These primers were predesigned to cover approximately every 1 Kb from the transcription start site of each gene. PCR reactions were thermocycled as follows: 95°C for 10 min followed by 40 cycles of two temperature PCR at 95°C for 15 s for denaturing at 60°C–67°C for 1 min for annealing and extension. Melting curve analysis was conducted to determine the optimal annealing temperature of these primers to minimize the production of nonspecific amplicons. Amplification efficacy of each PCR reaction was confirmed to fall into 95%–105%.
ChIP-qPCR data were normalized by the fold enrichment method and given as the fold increase in a signal relative to the background signal with nonspecific IgG.
Immunoblotting
After the indicated treatments, cell lysates were generated by means of collection with the RIPA buffer. The protein content of resultant supernatants was determined by using BCA protein reagents (Thermo Fisher Scientific). Protein electrophoresis was performed by using the NuPAGE electrophoresis system (Life Technologies). In brief, after adding the NuPAGE LDS Sample Buffer and NuPAGE Sample Reducing Agent, protein lysates were incubated at 70°C for 10 min, and then 10 μg of whole cell lysates was loaded, separated by NuPAGE® Novex Bis-Tris Gels with the MOPS SDS Running Buffer, and transferred to a polyvinylidene difluoride (PVDF) membrane. Blots were probed by anti-alpha tubulin (DM1A) (EMD Millipore) or anti-EGR2 (ab43020) (Abcam) antibodies with HRP-conjugated secondary antibodies included in GE Healthcare Amershame ECL Western Blotting kit (Thermo Fisher Scientific) and detected using Immobilon Western Chemiluminescent HRP substrate (EMD Millipore).
Statistical analysis
All experiments were performed in triplicate. Data were analyzed using the Student's t-test. Significance was set at P<0.05 or more stringent as noted in the Figures and Figure legends.
Results
Various growth factors and cytokines induce EGR2 expression
Priming with proinflammatory cytokines produced by lymphocytes and antigen presenting cells is needed to elicit anti-inflammatory properties of MSCs [16,39]. At the initial stage of the project, we evaluated the induction of EGR2 by growth factors and cytokines known to play pivotal roles in tissue repair and immunoregulation. Among proinflammatory cytokines, both IL1β and IFNγ upregulate EGR2 expression (Fig. 1).
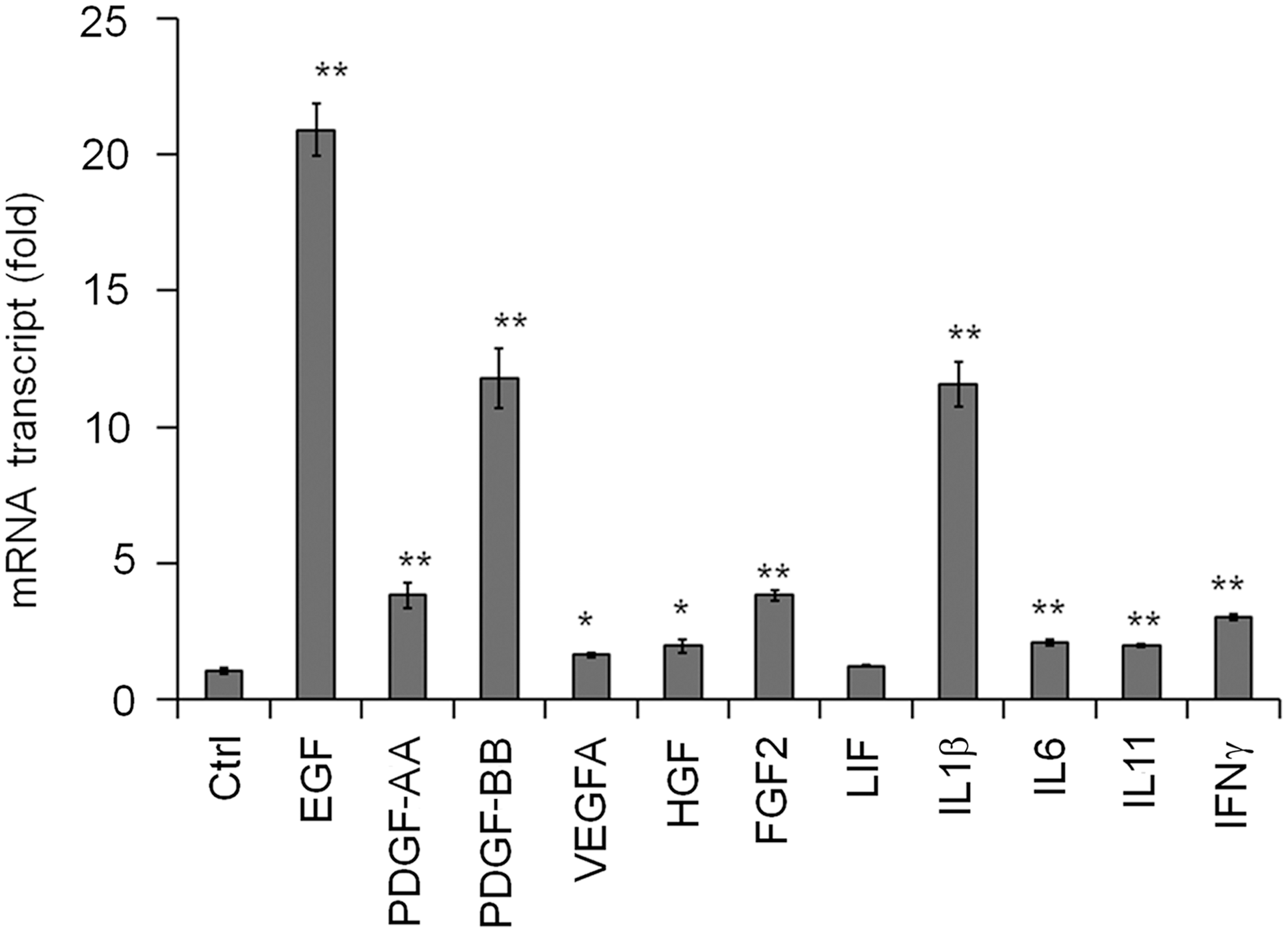
Effects of immortalized multipotential stromal cell or mesenchymal stem cell (MSC) treatments with various growth factors and cytokines on the induction of early growth response gene-2 (EGR2). MSCs were stimulated with epidermal growth factor (EGF) (10 nM), PDGF-AA (30 ng/mL), PDGF-BB (30 ng/mL), vascular endothelial growth factor A (VEGFA) (100 ng/mL), HGF (50 ng/mL), FGF2 (30 ng/mL), leukemia inhibitory factor (LIF) (10 ng/mL), interleukin-1 beta (IL1β) (10 ng/mL), interleukin-6 (IL6) (10 ng/mL), IL11 (10 ng/mL), and interferon-γ (IFNγ) (20 ng/mL) for 1 h and EGR2 induction was assessed by quantitative reverse transcription–polymerase chain reaction (qRT-PCR). The readouts are given in relative to the level of EGR2 mRNA in no treatment control (Ctrl). Shown are mean±SEM of three experiments each performed in triplicate (*P<0.05, **P<0.005 to vehicle control in each analyte).
Cotreatment with IL1β and IFNγ upregulates immunomodulatory molecules (IL6, LIF, IDO1, COX2/PTGS2) in MSCs
EGRs are known to be immediate early response genes [27 –29] and they are regulated mainly at the transcription level. Similar to EGR1, IL1β stimulation strongly and rapidly upregulates EGR2 expression within 60 min, and the level of EGR2 transcript is back to near baseline within 4 h in MSCs (Fig. 2A). IFNγ stimulation also upregulates EGR2 expression, but less strongly than IL1β stimulation (Fig. 2A). Interestingly, cotreatment with IL1β and IFNγ synergistically upregulates EGR2 expression (Fig. 2A). The EGR2 protein is also upregulated within 1 h, in parallel to its transcript (Fig. 2B). The level of EGR2 transcript in HDFs is lower than MSCs at baseline as well as under IL1β and/or IFNγ stimulation (Fig. 3A), consistent with previous microarray data (GSE9451).
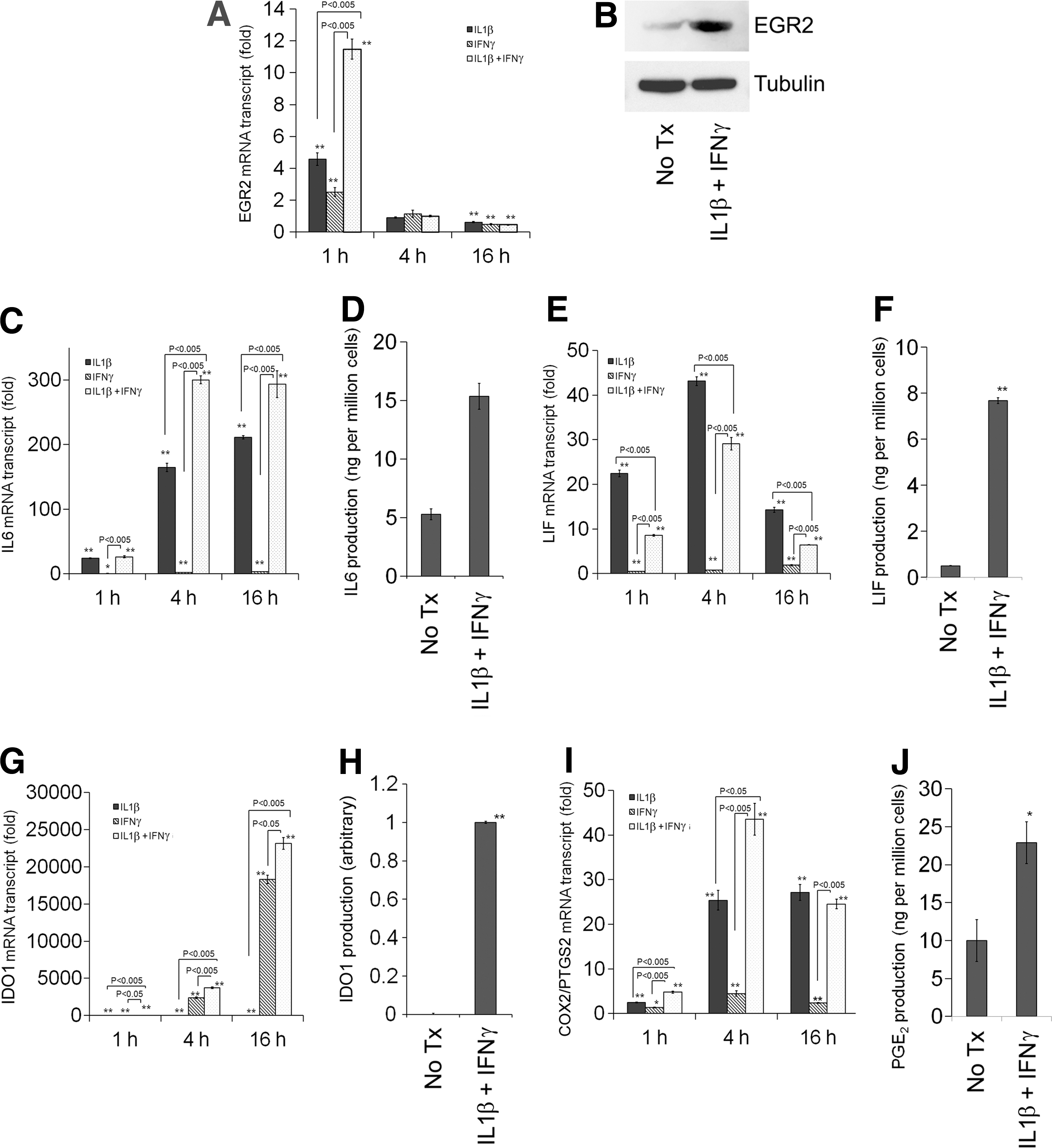
Effects of IL1β and/or IFNγ stimulation on the induction of EGR2
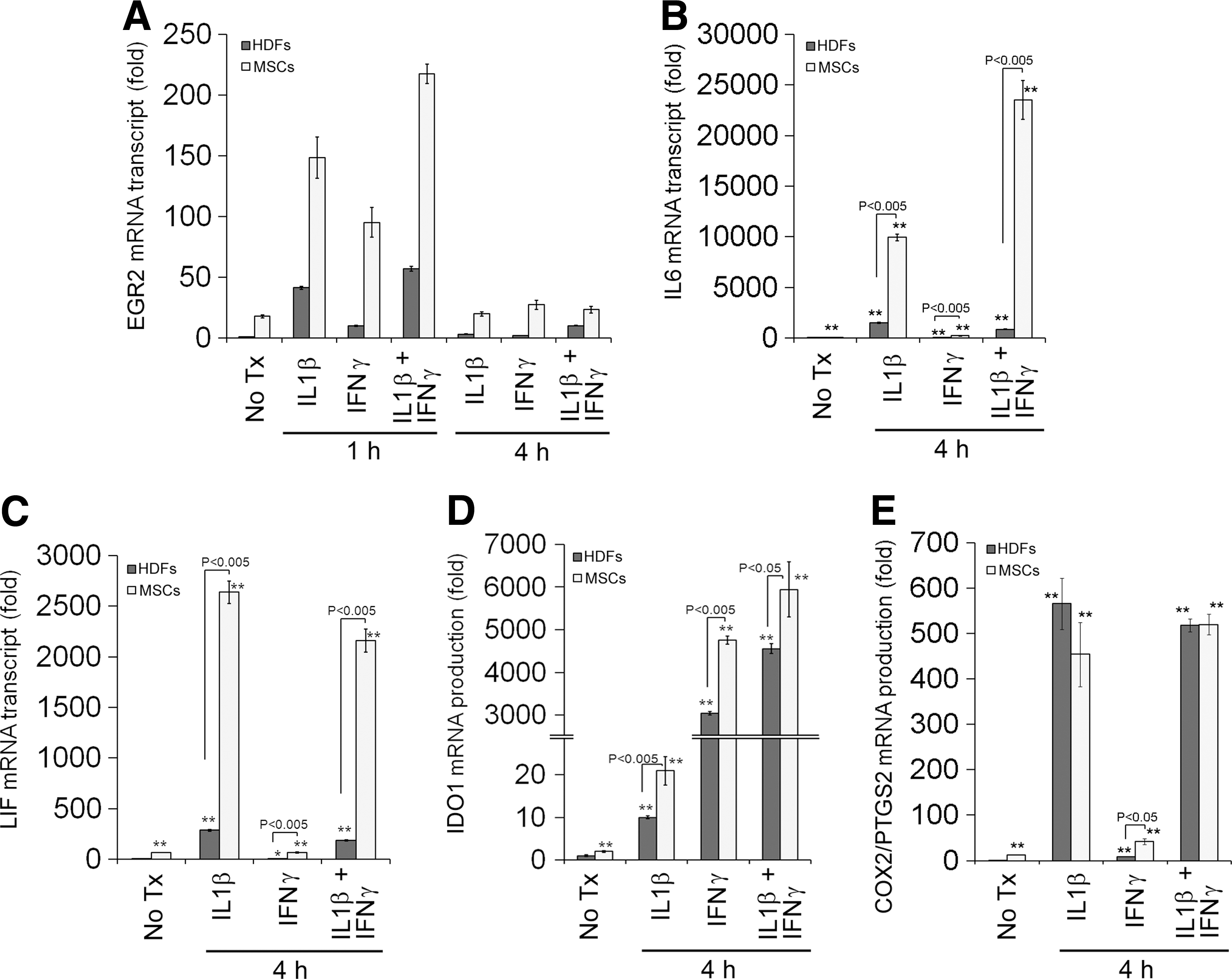
Comparison of the induction of EGR2
Previous reports identified several immunomodulatory molecules for MSCs, and stimulation with proinflammatory cytokines such as IFNγ augments the immunomodulatory properties of MSCs [15,16,39]. Consistently, IL1β treatment upregulates IL6, LIF, and cyclooxygenase-2/prostaglandin-endoperoxide synthase 2 (COX2/PTGS2), whereas IFNγ treatment strongly upregulates IDO1 (Fig. 2). Moreover, cotreatment with IL1β and IFNγ synergistically upregulates expression of these genes except LIF (Fig. 2). Treatment with IL1β and/or IFNγ also upregulates these genes in HDFs, but not as strongly as in MSCs except COX2/PTGS2 (Fig. 3), further underscoring the characteristic nature of immunomodulatory properties in MSCs.
EGR2 is a key regulator of MSCs' immunomodulatory effects
To test the hypothesis that EGR2 is a key regulator of MSCs' immunomodulatory effects, we used siRNA against EGR2 (siEGR2) (Fig. 4). siEGR2 reduces expression of IL6, LIF, IDO1, and COX2/PTGS2 (Fig. 4B–L and Supplementary Fig. S1; Supplementary Data are available online at
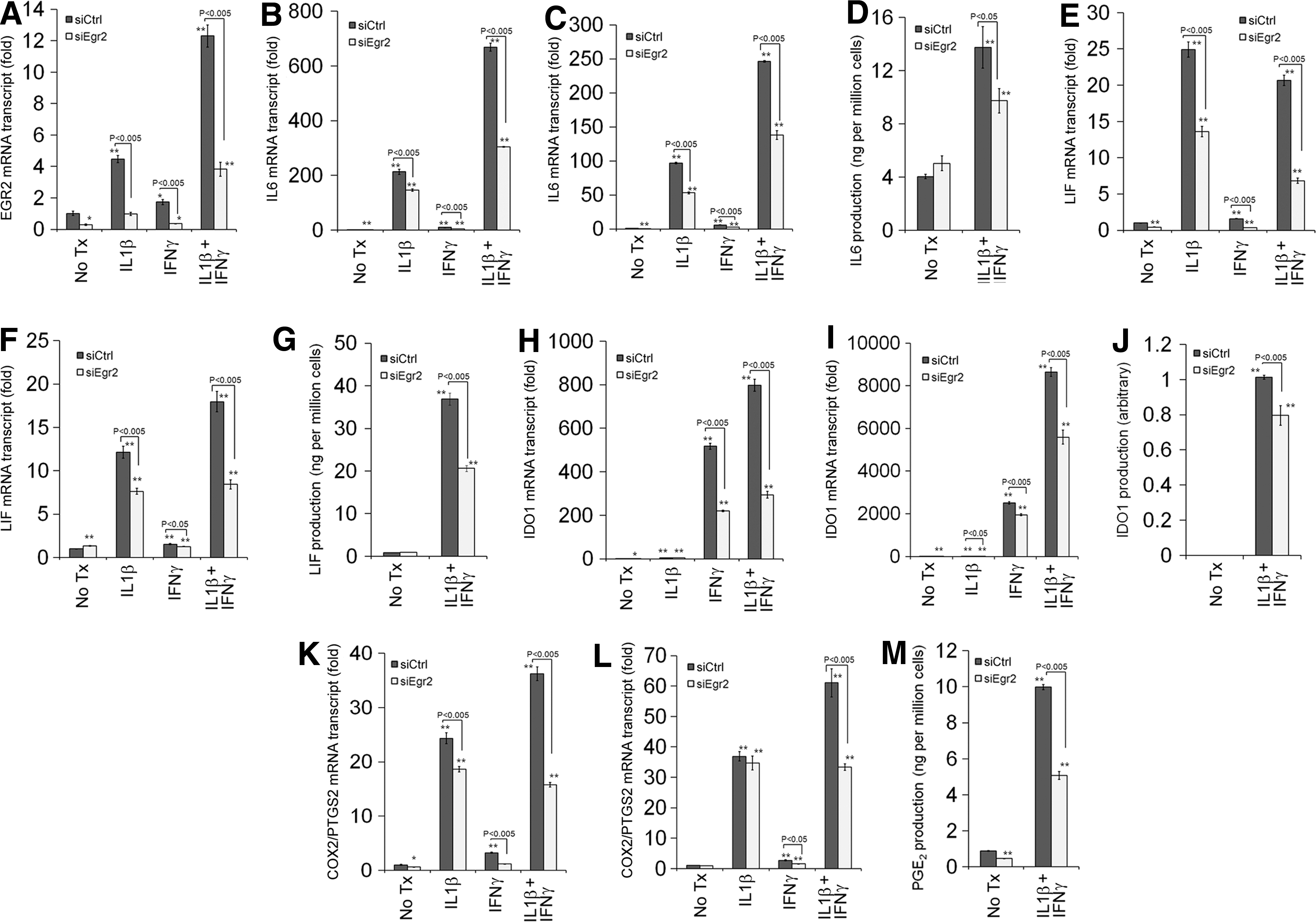
Effects of siRNA against EGR2 (siEGR2) on the induction of EGR2
Coefficient of determination was calculated through linear regression.
IFNγ, interferon-γ; siEGR2, siRNA against EGR2.
MSCs exert immunomodulatory effects in multiple mechanisms [15 –17]. One of the major mechanisms is through reduction of T-cell proliferation. In this study, we used human primary CD4+ T cells isolated from the buffy coat of one donor (Supplementary Fig. S2). In our experimental system, MSCs cocultured with T cells reduced proliferation of primary T cells stimulated by IL2 and CD3/28 activator beads (Supplementary Fig. S3). T cells under stimulation by CD3/28 activator beads and IL2 strongly express IFNG in the presence or absence of MSCs, whereas the induction of IL1B was noted in T cells only when they are cocultured with MSCs (data not shown). On the other hand, MSCs cocultured with T cells upregulate IL6, LIF, IDO1, and COX2/PTGS2 (Fig. 5B–E). Moreover, T-cell coculture strongly induces EGR2 (more than 40-fold) (Fig. 5A), much higher than the induction of EGR1 (about 1.6-fold) or EGR3 (about 6-fold) in MSCs (data not shown), further substantiating the key role of EGR2 in MSCs in T-cell coculture.
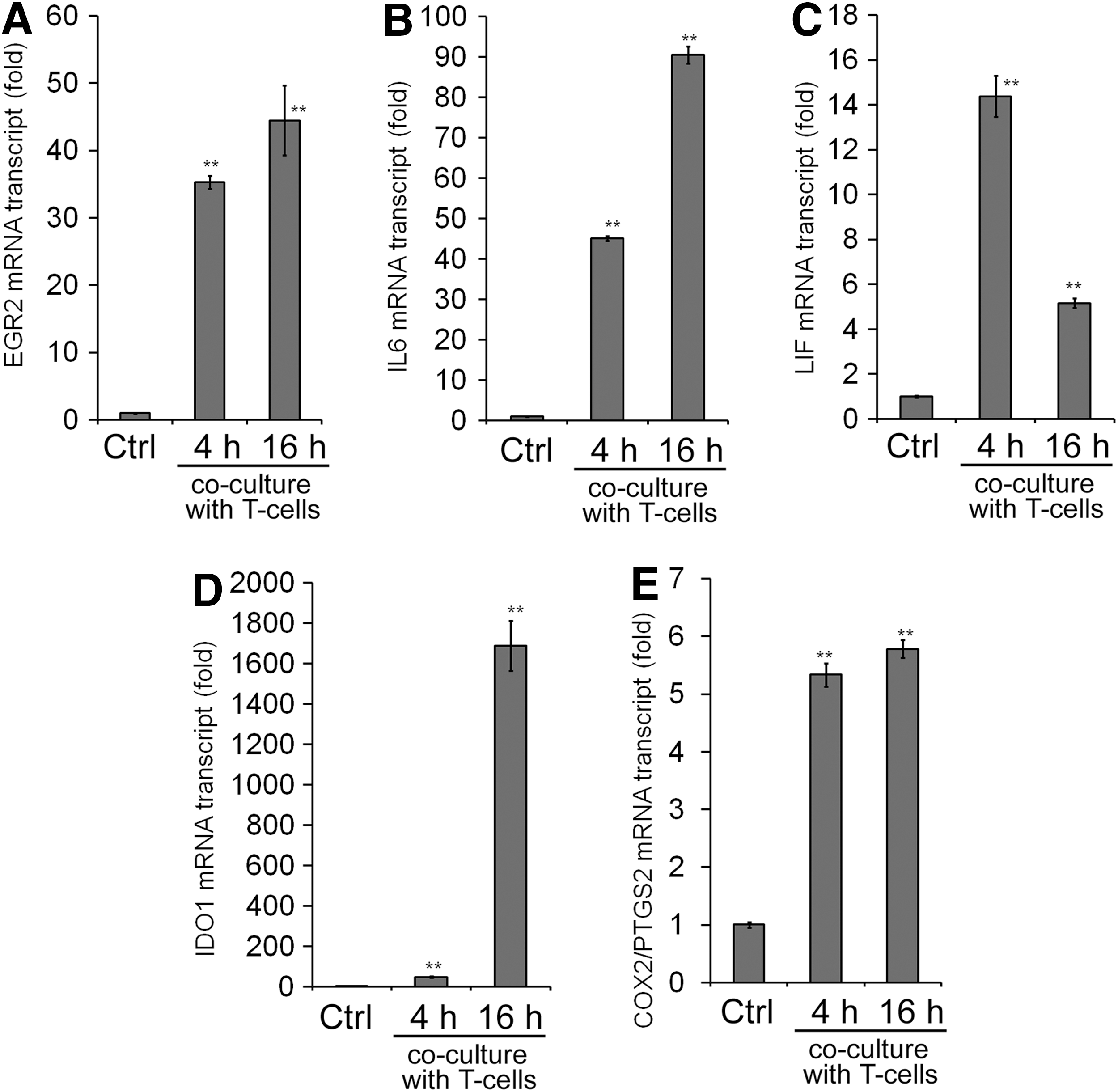
Effects of T-cell coculture on gene expression of immortalized MSCs. After coculturing with T cells for 4 or 16 h, the gene expression of EGR2
Addition of tryptophan or COX inhibitor indomethacin reversed the MSC-mediated reduction of T-cell proliferation, whereas neutralizing antibodies against IL6 or LIF did not reverse it (Supplementary Fig. S3), suggesting that MSCs reduce T-cell proliferation through IDO1 and PGE2, in agreement with previous studies [40,41].
Moreover, MSC-based reduction of T-cell proliferation was also reversed by siEGR2 treatment on immortalized MSCs as well as primary MSCs from two donors, further confirming the critical role of EGR2 in MSCs' immunomodulatory effects (Fig. 6).
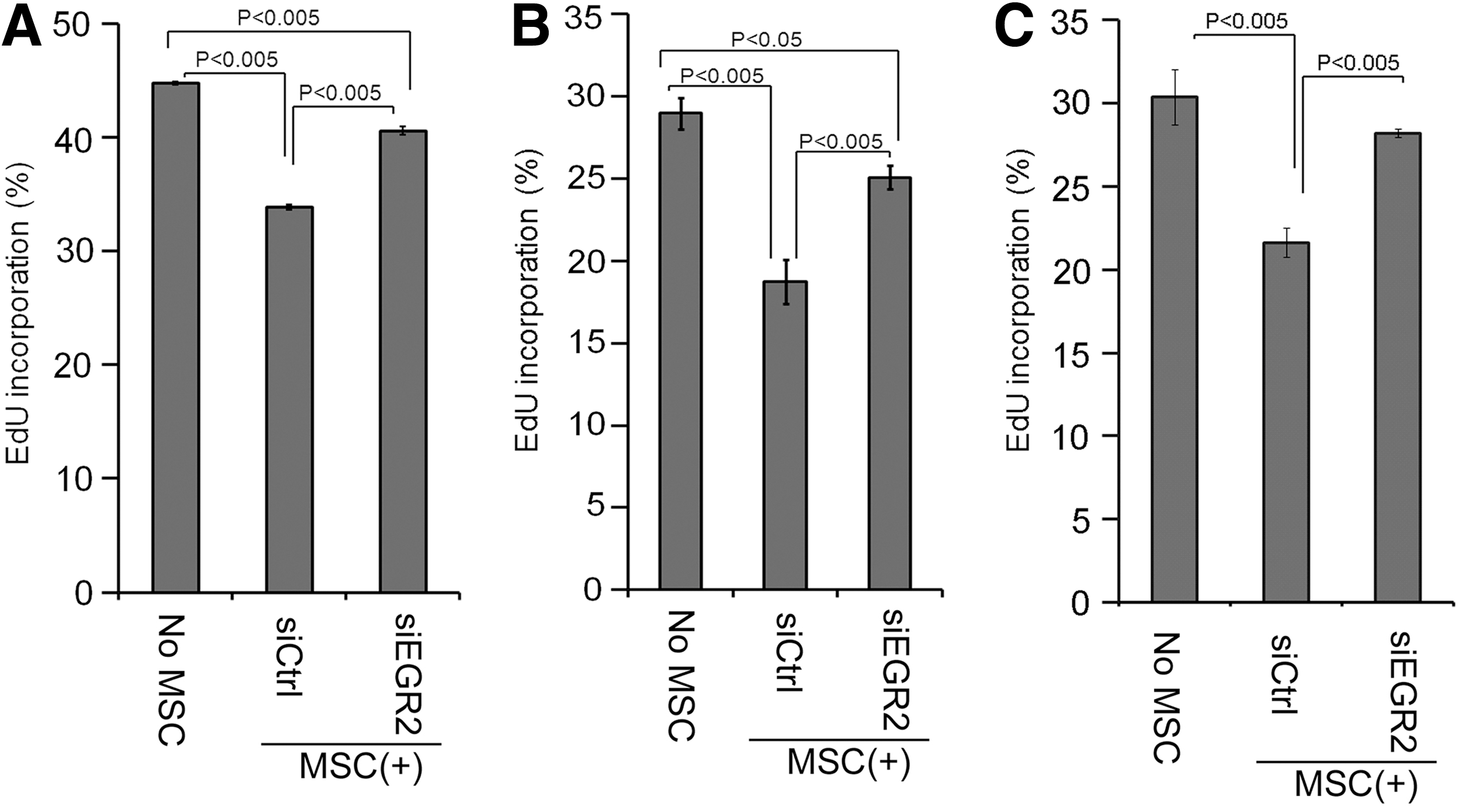
Effects of siEGR2 on the MSCs' inhibitory effect on T-cell proliferation induced by IL2 and CD3/CD28 T-activator beads
EGR2 directly regulates expression of IL6, LIF, and IDO1 genes
EGRs regulate gene expression by binding to cis-acting elements of the target genes [28,29]. We showed that gene expression of IL6, LIF, IDO1, and COX2/PTGS2 is regulated in an EGR2-dependent manner (Fig. 4). We further evaluated whether EGR2 directly regulates these genes by using ChIP-qPCR.
First, we interrogated the putative promoter regions of these genes in silico by using the TRANSFAC database. These genes contain predicted EGR binding sequences in their putative promoter regions (Table 3), and some of them were shown to bind to EGR1, a surrogate TF of EGR2 as they share the cis-binding elements [42], in the high-throughput data publicly available in the ENCODE project [36,37]. Next, we evaluated whether EGR2 directly binds to these regions by using ChIP-qPCR. Costimulation with IL1β and IFNγ increases EGR2 binding to the putative promoters of IL6, LIF, and IDO1 (Fig. 7), suggesting that EGR2 directly regulates these genes. Although siEGR2 decreased COX2/PTGS2 expression and the putative promoter region contains the predicted EGR binding sequences (Table 2), we did not observe the strong EGR2 binding to those regions (Fig. 7).
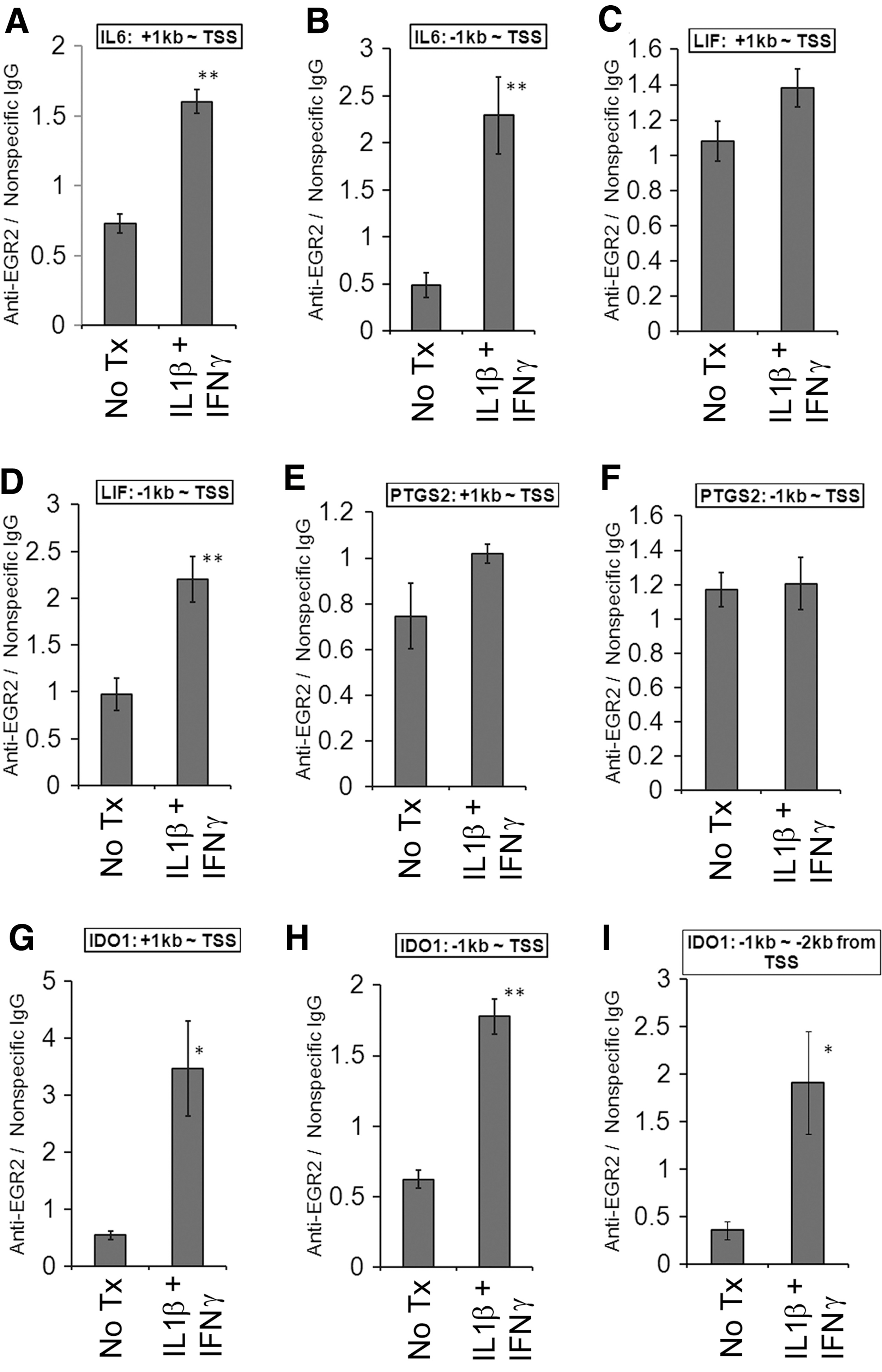
Chromatin immunoprecipitation (ChIP)-qPCR analysis of EGR2 binding. Immortalized MSCs were stimulated with IL1β (10 ng/mL) and IFNγ (20 ng/mL) for 1 h, and the cell lysates were collected for evaluation of direct EGR2 binding to the putative promoter regions of IL6
ChIP, chromatin immunoprecipitation; qPCR, quantitative polymerase chain reaction.
Discussion
Numerous studies have shown MSCs' immunomodulatory effects [43,44]. Indeed, MSC-based clinical studies for immune-mediated diseases such as inflammatory bowel disease or multiple sclerosis have been ongoing. However, MSCs' immunomodulatory effects have been challenged [16]. For example, one study showed that MSC transplantation was shown to accelerate skin allograft rejection in a rat preclinical model [45]. Moreover, MSCs were shown to be nonsuperior to placebo for GVHD in one large-scale, placebo-controlled phase III clinical trial and the authors speculate that ex vivo expanded MSCs might lose immunomodulatory effects, especially after extended serial passages [23]. These results clearly indicate the importance of mechanistic studies about immunomodulatory effects by MSCs, as these studies should lead to maximization of these immune response modifying properties [15].
MSCs exert their immunomodulatory effects through multiple mechanisms using not only T cells, but also B cells, monocytes, dendritic cells, NK cells, and neutrophils [16,17]. Even though key intercellular signaling molecules mediating human MSCs' immunomodulatory effects, such as IDO1 or PGE2, have been identified [15 –17], the entire mechanism of these effects, such as intracellular molecular switches regulating immunomodulatory properties in MSCs, remains unknown. In the current study, we showed for the first time that TF EGR2 plays a key role in MSCs immunomodulatory effects.
EGR2 is known to be a key regulator of myelination in the nervous system and of hindbrain development [30 –32]. It was also identified as a negative regulator of T-cell activation through induction of clonal anergy in CD4-positive T cells [33,46]. Whereas it is known that MSCs express EGR2, the role of this molecule has not been studied [27]. siRNA-mediated knockdown of EGR2 decreased gene expression of IL6, LIF, IDO1, and COX2/PTGS2 and the PGE2 level (Fig. 5). Moreover, EGR2 was bound to the putative promoter regions of IL6, LIF, and IDO1 (Fig. 7). Those data indicate that key immunomodulatory molecules (IL6, LIF, IDO1, and PGE2) are regulated in an EGR2-dependent manner in MSCs.
Among the various modes of immunomodulation by MSCs, we chose MSC-mediated suppression of T-cell proliferation as an in vitro model system because it is simple, robust, and widely used. We noticed that MSC-derived suppression of T-cell proliferation is more prominent with primary MSCs than immortalized MSCs (Fig. 6). One possible reason is the stronger induction of IL6 by immortalized MSCs than primary MSCs (Fig. 4 and Supplementary Fig. S1) because IL6 released from MSCs was shown to mitigate MSC-derived suppression of T-cell proliferation by stimulating T-cell proliferation [47]. It is unclear whether this strong IL6 induction is related with the immortalization process or interdonor variability of MSC itself. We chose not to add IFNγ or IL1β during coculture to make the system simpler; but regardless, MSCs' immunomodulatory properties were solely dependent on the proinflammatory cytokines produced by the MSC cocultured CD4+ T cells stimulated by IL2 and CD3/28 activator beads.
In this system, the effect of siEGR2 on the MSC-derived suppression of T-cell proliferation was rather modest (Fig. 6). One possible reason is that siEGR2 only moderately reduces the levels of IDO1, especially in 16 h (Fig. 4H–J), and it is IDO1 that plays a pivotal role in the MSC-derived suppression of T-cell proliferation (Supplementary Fig. S3A). A higher concentration of siRNA might be an option to attain better knockdown of EGR2 and subsequent decrease of IDO1; however, it also deranges expression of EGR1 and −3 in a nonspecific manner presumably because EGRs are over-reactive to those exogenously applied siRNA molecules (data not shown), which subsequently affects EGR2 expression [48]. We might need to target additional molecules, such as upstream regulators of EGR2, for better knockdown of EGR2. Another probable reason is the possible presence of EGR2-independent mechanism of immunomodulation by MSCs, such as HLA-G5 or tumor necrosis factor-stimulated gene 6 (TSG6) (vide infra). Indeed the T-cell proliferation is different between no MSC control and MSC with siEGR2 in MSCs (Fig. 6), which is consistent with that speculation. We used immortalized MSCs for ease of handling and better consistency of results for this model system. siEGR2 produces similar effects on the expression of immunomodulatory molecules as well as MSC-mediated suppression of T-cell proliferation in human primary MSCs (Figs. 4 and 6 and Supplementary Fig. S1), confirming the validity of the roles of EGR2 in human primary MSCs in general. The effects of siEGR2 on the MSC-based suppression of T-cell proliferation were rather prominent in primary MSCs from two donors (Fig. 6C). It is unclear whether this apparent discrepancy among these cells is due to the immortalization process or interdonor variability of MSCs (vida infra).
Interdonor variability of MSCs' immunosuppressive potentials has been noted, as seen in our data with primary MSCs (Fig. 6B, C) [23]; however, little is known about the molecular mechanism of the interdonor variability [23]. According to the latest single-nucleotide polymorphism (SNP) database (dbSNP build 137) established by the National Center for Biotechnology Information (NCBI) in collaboration with the National Human Genome Research Institute (NHGRI) dbSNP build 137 [49], there are 22 common SNPs and 5 clinically associated flagged SNPs within the EGR2 gene. All the flagged SNPs are found within the zinc finger domains or NGFI-A-binding (NAB) repressor binding site, and these are associated with congential hypomyelinating neuropathies [32,50,51]. Among 22 common SNPs, 2 SNPs are found in 3′-untranslated region (3′-UTR), which can be a target of microRNA. Indeed, rs61865883 is predicted to be located within the target site of hsa-miR-320a, hsa-miR-320b, hsa-miR-320c, hsa-miR-320d, hsa-miR-320e, and hsa-miR-4429 by polymorphism in the microRNA Target Site (PolymiRTS) Database [52]. Indeed, MSCs express hsa-miR-320 (data not shown). Moreover, 18 common SNPs are found within introns or 5′ upstream regions. All of these regions are within the DNase clusters and 16 of them are within the H3K4Me3 regions, where TFs could bind. Specifically, nine of them (rs10995315, rs2297488, rs2297489, rs7893472, rs113921640, rs475489, rs7917418, rs149153286, and rs61865884) are located within the predicted TF binding sequences by TRANSFAC database. Thus, theoretically, SNPs within EGR2 promoter sequences or the 3′-UTR of EGR2 gene could possibly affect the EGR2 mRNA level by altering the efficacy of microRNA-based degradation and/or altering the efficacy of the EGR2 gene activation/repression by TFs, although there is no experimental evidence to support the speculation.
EGRs are immediate response genes whose strong induction is quickly counteracted and their mRNA levels get down quickly even in the presence of IL1β and IFNγ. Moreover, we noticed that the baseline expression of EGRs is fluctuating (data not shown). Those factors could explain the apparent decrease of EGR2 mRNA after 16 h, lower than the baseline in Figure 2A. Interestingly, EGR2 mRNA remains elevated in MSCs after 16 h of coculture with T cells (Fig. 5). The apparent discrepant response of EGR2 between IL1β/IFNγ stimulation and T-cell coculture should indicate the presence of unidentified mechanisms inducing EGR2 responses in MSCs cocultured with T cells. At this point, we do not have any data about this unknown mechanism, and this is beyond the scope of this study.
Importantly, MSC-derived suppression of T-cell proliferation only reflects a glimpse of the entire MSC-mediated immunomodulatory effects. IL6, LIF, IDO1, and COX2/PGE2 mediate MSCs' immunomodulatory effects beyond suppression of T-cell proliferation [16,17], and thus it is quite likely that MSC EGR2 signaling modifies the immune response in multiple ways. However, the overall immune response in the body is too complex to be fully reproduced in vitro; for example, the bidirectional interactions of MSCs with various types of cells (immune and nonimmune cells) could affect the immunomodulatory capabilities of MSCs. Thus, it is challenging to evaluate the role of MSC EGR2 signaling in every aspect of MSC-derived immunomodulatory effects solely in vitro. Further in vivo studies would be needed to evaluate the overall role of MSC EGR2 signaling in the MSCs' immunomodulatory effects in the body.
HLA-G5, inducible nitric oxide synthase (iNOS), and TSG6 were reported to mediate the immunomodulatory effects of MSCs [16,17]. Unlike in human MSCs, iNOS was shown to be the key immunomodulatory molecule for murine MSCs [53,54]. Thus, even though iNOS expression is regulated by EGR2 in human MSCs (data not shown), we did not cover iNOS in the current study. We do not believe that EGR2 regulates expression of HLA-G5 and TSG6, because no EGR1 (surrogate of EGR2) binding was shown in these regions in the high-throughput ChIP sequence data in the ENCODE project. Moreover, siRNA against EGR2 did not reduce TSG6 expression in MSCs (data now shown). That suggests immunomodulatory properties of human MSCs would also be regulated in an EGR2-independent manner as well.
In summary, we showed that human MSCs exert suppression of T-cell proliferation in vitro, and EGR2 serves as a molecular switch to regulate immunomodulatory molecules in MSCs. EGR2 can be a molecular target to manipulate human MSCs' immunomodulatory properties.
Footnotes
Acknowledgments
This study was supported by the University of Pittsburgh Schools of Health Sciences (Bridge Funding Category One) and the University Pittsburgh Medical Center Health System (Competitive Medical Research Fund). We would like to thank Dr. Junya Toguchida (Kyoto University, Kyoto, Japan) for kindly providing the immortalized human MSCs. We would also thank Drs. Michael R. Shurin and Nahed Ismail (University of Pittsburgh) for their technical guidance and discussion.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.