
Abstract
White adipose tissue (WAT) is a very attractive source of mesenchymal stem cells (MSCs) because of its availability and ease of harvest. However, the current method of isolating adipose tissue-derived MSCs often relies on the adhesiveness of the cultured stromal-vascular fraction (SVF). Unfortunately, the SVF is a heterogeneous cell population containing many cell types, including adipocyte precursor cells, endothelial cells, pericytes, multipotent MSCs, erythrocytes, and hematopoietic cells. Here we systematically characterized the adipose tissue-derived lineage-negative (Lin−) cell population using various surface markers and a set of cell proliferation and differentiation assays. We demonstrate clearly that the Lin− cell population represents enriched MSCs, which were identified by their high expression of MSC surface markers, and that these cells are a robust population with a vigorous growth capability and delayed aging. This cell population also demonstrated a much higher capacity for differentiation into osteogenic, chondrogenic and adipogenic cell lineages related to MSCs than did the SVF. These cells promoted recovery from limb ischemia, likely via production of vascular endothelial growth factor, an angiogenic factor. Our study demonstrates that Lin− cells are enriched in MSCs and provides a reliable method for isolating purer MSCs than SVF cells from the WAT, especially for obtaining fresh MSCs for clinical applications. In summary, this study identified a new, reliable method for enrichment of WAT MSCs with regenerative repairing features.
Introduction
M
In addition to mature adipocytes within WAT, there are other cell populations, in particular the stromal-vascular fraction (SVF). Until now, many studies considered cultured adherent SVF cells to be MSCs because they behave like bone marrow MSCs (BMSCs) and can differentiate into osteogenic, chondrogenic, and adipogenic cells in vitro [5,9 –14]. However, SVF cells are a heterogeneous cell population, containing adipocyte precursor cells, endothelial cells, pericytes, multipotent MSCs, erythrocytes, and hematopoietic cells, among others [7,15 –17]. Although studies have reported that adipose tissue-derived stromal cells promote repair after limb ischemia [18 –20], the cells used are unsorted SVF cells, and the mechanism for such an effect is unclear. In fact, owing to the heterogeneity of cell types in the SVF, it is unknown whether MSCs, endothelial cells or both contribute to the recovery from limb ischemia. On the other hand, Sugii et al. recently reported that the adipose tissue-derived lineage-negative (Lin−) cell population can be reprogrammed to become induced pluripotent stem (iPS) cells with substantially higher efficiencies than fibroblasts [7]. Recently, our work is also the first to use Lin− cells to obtain cloned mice via nuclear transfer [21]. However, the Lin− population has not been characterized, and it is unclear what cell type they belong to. It is also unknown whether Lin− cells possess the same regenerative repair features as MSCs.
In the present study, we systematically characterized the adipose tissue-derived Lin− cell population using various types of surface markers. In addition, we performed a set of cell proliferation and differentiation assays, including induction of osteogenic, chondrogenic, and adipogenic cell lineages in vitro under different culture conditions. Our studies indicated that adipose-derived Lin− cells not only selectively expressed high levels of MSC surface markers but also promoted recovery of hindlimb ischemia, likely via production of vascular endothelial growth factor (VEGF), an important angiogenic factor. Lin− cells displayed no endothelial phenotype, either in vitro or in vivo. Therefore, our study identified a new, reliable method for enriching WAT MSCs with regenerative repairing features.
Materials and Methods
Animal use and care
Adult (8–12 weeks of age) male C57B6/L and transgenic RFP mice (FVB) were used. All animal procedures were performed under the ethical guidelines of the Institute of Biochemistry and Cell Biology, Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences and Shanghai Jiaotong University.
Labeling and sorting of SVF
As described previously [15,21], WAT from mouse inguinal fat pads was harvested, washed several times with phosphate-buffered saline (PBS), and excised into very small pieces. The tissues were then digested with 0.1% collagenase (Sigma) on a shaker for 50 min at 37°C. Dulbecco's modified Eagle's medium (DMEM; Invitrogen) with 10% fetal bovine serum (FBS; Gibco) was used to neutralize the enzyme activity. The cell suspensions were centrifuged at 400 g for 8 min, and the supernatant was removed. The pelleted cells, so-called SVF cells, were suspended in 200 μL PBS containing 2% FBS and were incubated on ice for 20 min with flow cytometry antibodies: CD45 Percp-Cy5.5 (eBioscience; Clone: 30-F11), Ter119 PE-Cy7 (eBioscience; Clone: TER-119) and CD31 APC (BD Biosciences; Clone: MEC 13.3) antibodies. Samples were sorted on a BD FACSAria II cell sorter (BD Biosciences) equipped with BD FACSDiva Software. For sorting, cells were initially selected by size, on the basis of forward scatter (FSC) and side scatter (SSC). Then, cells were gated on both SSC and FSC singlets, ensuring that the individual cells were analyzed. Next, the cells were separated on the basis of the indicated cell-surface markers.
Derivation of mouse embryonic fibroblasts
Mouse embryonic fibroblasts (MEFs) were isolated from E13.5 embryos of C57B6/L mice. Gonads and internal organs were removed when the embryos were processed for MEFs isolation. Isolated MEFs in early passages were used for further experiments.
Surface marker analyses by flow cytometry
Fresh SVF cells, fresh Lin− cells, cultured SVF cells at passage 1 (SVF P1), cultured Lin− cells at passage 1 (Lin− P1), MEFs and 3T3-L1 cells were used for surface marker analyses. At first, each set of cells was divided into 13 aliquots. Each aliquot contained ∼500,000 cells and was labeled with one of the following fluorochrome-conjugated anti-mouse antibodies: CD140a PE (eBioscience; Clone: APA5), CD140b PE (eBioscience; Clone: APB5), CD24 FITC (eBioscience; Clone: 30-F1), CD146 PE (eBioscience; Clone: P1H12), CD105 PE (eBioscience; Clone: MJ7/18), CD13 FITC (BD Pharmingen; Clone: R3-242), CD73 PE (eBioscience; Clone: eBioTY/11.8), CD44 FITC (eBioscience; Clone: IM7), CD34 APC (BioLegend; Clone: MEC14.7), Sca1 PE (eBioscience; Clone: D7), MHC-1 PE (eBioscience; Clone: 28-14-8), MHC-2 FITC (eBioscience; Clone: NIMR-4) and CD45 Percp-Cy5.5 (eBioscience; Clone: 30-F11), according to the manufacturer's instructions. Gating was performed with matched isotype control antibodies. Flow cytometry was conducted on a Becton Dickinson FACSCalibur analyzer.
Culture of Lin− cells in different media
First, to detect their cell proliferation ability in vitro, 2,000 Lin− cells were plated into one well of six-well plate and cultured in normal medium (NM) and low-serum medium (LSM) [22], simultaneously with SVF cells as control. NM is DMEM-LG with 10% FBS. LSM contains 60% DMEM-LG, 40% MCDB-201 (Sigma), 2% FBS, 20 ng/mL platelet-derived growth factor (R&D), 20 ng/mL epidermal growth factor (R&D), and 20 ng/mL leukemia inhibitory factor (Chemicon). In LSM, cells were plated into 6-well plates coated with fibronectin (BD Biosciences). After the cells grew to a confluence of 70%–80%, they were passaged. The experiment was repeated three times at the same time point. At each of time point, the cells in each group were counted using a hemocytometer.
To determine whether Lin− cells could differentiate into endothelial cells in vitro, these cells were cultured in EGM-2 (LONZA) and NMV medium (NM supplemented with 50 ng/mL VEGF). After 7 days, the cells were used for reverse transcription–polymerase chain reaction (RT-PCR).
Adipogenesis, osteogenesis, and chondrogenesis in culture
The methods followed previous reports [6,21]. In brief, freshly isolated Lin− cells were plated into 24-well plates in DMEM supplemented with 10% FBS, allowed to grow to confluence, and then changed to a related inducing medium. All staining was performed on day 21 after the initial induction.
Osteogenic differentiation medium contained 10% FBS DMEM, 0.1 μM dexamethasone (Sigma), 10 mM β-glycerophosphate (Sigma), and 50 μg/mL ascorbic acid-2-phosphate (Sigma). Differentiated osteoblasts were stained with Alizarin red. The cells were rinsed with PBS and fixed in 4% paraformaldehyde for 15 min at room temperature. They were then stained with 0.1% Alizarin red in Tris-HCl (pH 8.3) for 30 min at 37°C. The plates were washed several times with distilled water.
Adipogenic differentiation medium contained 10% FBS DMEM, 1 μM dexamethasone (Amresco), 10 μM insulin (Sigma), 200 μM indomethacin (Amresco), and 0.5 mM isobutyl-methylxanthine (Amresco). The medium was replaced every 3 days. Differentiated adipocytes were assessed using oil red O staining. The cells were fixed for 15 min in 4% paraformaldehyde solution and incubated with 1% oil red O (Sigma) in 60% isopropanol for 10 min at room temperature. Excess stain was removed by washing with 60% isopropanol. Then, the plates were washed several times with distilled water.
Chondrogenic differentiation medium contained 10% FBS DMEM, 10 ng/mL transforming growth factor-β1 (TGF-β1; Sigma), 6.25 μg/mL insulin (Sigma), and 6.25 μg/mL transferrin (Sigma). Differentiated chondrocytes were assessed using Alcian blue staining. The cells were washed with PBS and fixed in 4% paraformaldehyde solution for 15 min at room temperature. Induced cells were incubated for 30 min with 1% Alcian blue in 0.1 N HCl (pH 1.0) and washed twice with 0.1 N HCl for 5 min to remove any nonspecific staining. The plates were then rinsed with distilled water for 5 min.
Mouse hindlimb ischemia
Male 8-week-old C57BL/2 mice underwent unilateral hindlimb ischemia as previously described [23,24]. The femoral artery and its branches were ligated through a skin incision with 5-0 silk. The external iliac artery and all of the above arteries were then ligated. The femoral artery was excised from its proximal origin as a branch of the external iliac artery to the distal point where it bifurcates into the saphenous and popliteal arteries.
Treatments of hindlimb ischemia
After arterial dissection, the mice were randomly divided into three experimental groups. In the first group, control animals were injected with vehicle (n=6). The treatment groups were intramuscularly injected with a microsyringe with either CD31+ cells (n=6) or Lin− cells (n=6) at three sites in the gracilis muscle in the medial thigh (1×106 cells per mouse).
Laser speckle imaging analysis
We used a laser speckle imager (LSCI-2; Dolphin Biotech) to detect hindlimb blood flow. The principle of laser speckle imaging (LSI) has been previously described [25 –27]. The LSCI-2 is a 21×18×21-cm portable instrument and is connected to a computer equipped with real-time data acquisition software (LSCI data processing system). It provides video images of blood flow in the microvasculature: up to 23 images per second at a maximum resolution of 1,700,000 pixels/cm2. The laser scan area is from 7×8.75 mm to 20×25 cm, and the working distance between the scanner and sample is 10–50 cm. After the mice were anesthetized with 2.5% Avertin, ischemic and normal hindlimb hair was removed with depilatory cream, and then the mice were imaged with the LSCI-2 to simultaneously detect the blood flow in both limbs. Blood flow recovery was measured immediately after ischemia induction and 0, 3, 6, and 9 days thereafter. The mean flow in the regions studied was calculated with the LSCI data processing system. The ratio of hindlimb blood flow is the ischemic (left)/nonischemic (right) hindlimb blood flow.
Immunohistochemistry
The ischemic hindlimb injected cells were fixed in 4% paraformaldehyde, frozen in tissue-freezing medium and sectioned at a thickness of eight microns. Sections were first permeabilized with PBS with 0.1% Triton X-100 (Sigma) and blocked with 5% donkey serum (GIBCO) in PBS and were then stained with primary antibody against CD31 (BD Biosciences; 1:50), Flk-1 (Santa Cruz; 1:50), and RFP (MBL; 1:200) overnight at 4°C. After rinsing, sections were incubated for 1 h at 37°C with a secondary antibody (Jackson; 1:500). The nuclei were counterstained with Hoechst 33342.
Reverse transcription–polymerase chain reaction
RTPCR was performed according to previous reports [6], and total RNA was extracted from 104 cells using the Absolutely RNA Nanoprep Kit (Stratagene). One microgram of total RNA was reverse transcribed using a First Strand cDNA Synthesis Kit (TOYOBO). PCR was performed for 30 cycles with an annealing temperature of 60°C with Taq polymerase (Invitrogen), and PCR products were electrophoresed on 2% agarose gels. Each set of oligonucleotides was designed to span two different exons so that genomic DNA contamination is of no concern. Primer sequences used for amplifications were as follows: CD31 Forward: 5′-CTGCCAGTCCGAAAATGGAAC-3′ Reverse: 5′-CTTCATCCACCGGGGCTATC-3′ Flk-1 Forward: 5′-GCAAAACACTCACCATTCCCA-3′ Reverse: 5′-GAGGTTTGAAATCGACCCTCG-3′ GAPDH Forward: 5′-TGCCCAGAACATCATCCCT-3′ Reverse: 5′-ATGCCTGCTTCACCACCTT-3′
Statistical analysis
Data were analyzed using GraphPad Prism software (GraphPad Software, Inc.). Data are expressed as mean±SEM and were analyzed with a two-tailed Student's t-test for two groups. A value of P<0.05 was considered significant.
Results
Sorting of the Lin− cell population from the SVF
WAT collected from inguinal fat pads was digested with collagenase, and then centrifuged. While the supernatant fractions contained mature adipocytes, the wine-colored bottom mass consisted of the SVF; and these SVF cells were incubated with fluorochrome-conjugated antibodies against various cell-surface markers, and then sorted by FACS (Fig. 1A). Lin− cells were separated by removing Lin+ cells, including endothelial cells, erythrocytes, and hematopoietic cells, on the basis of staining for CD31, Ter119, and CD45, respectively. CD31+ (endothelial) cells were isolated by depleting hematopoietic (CD45+) cells and erythrocytes (Ter119+) from the Lin+ cell population (Fig. 1B).
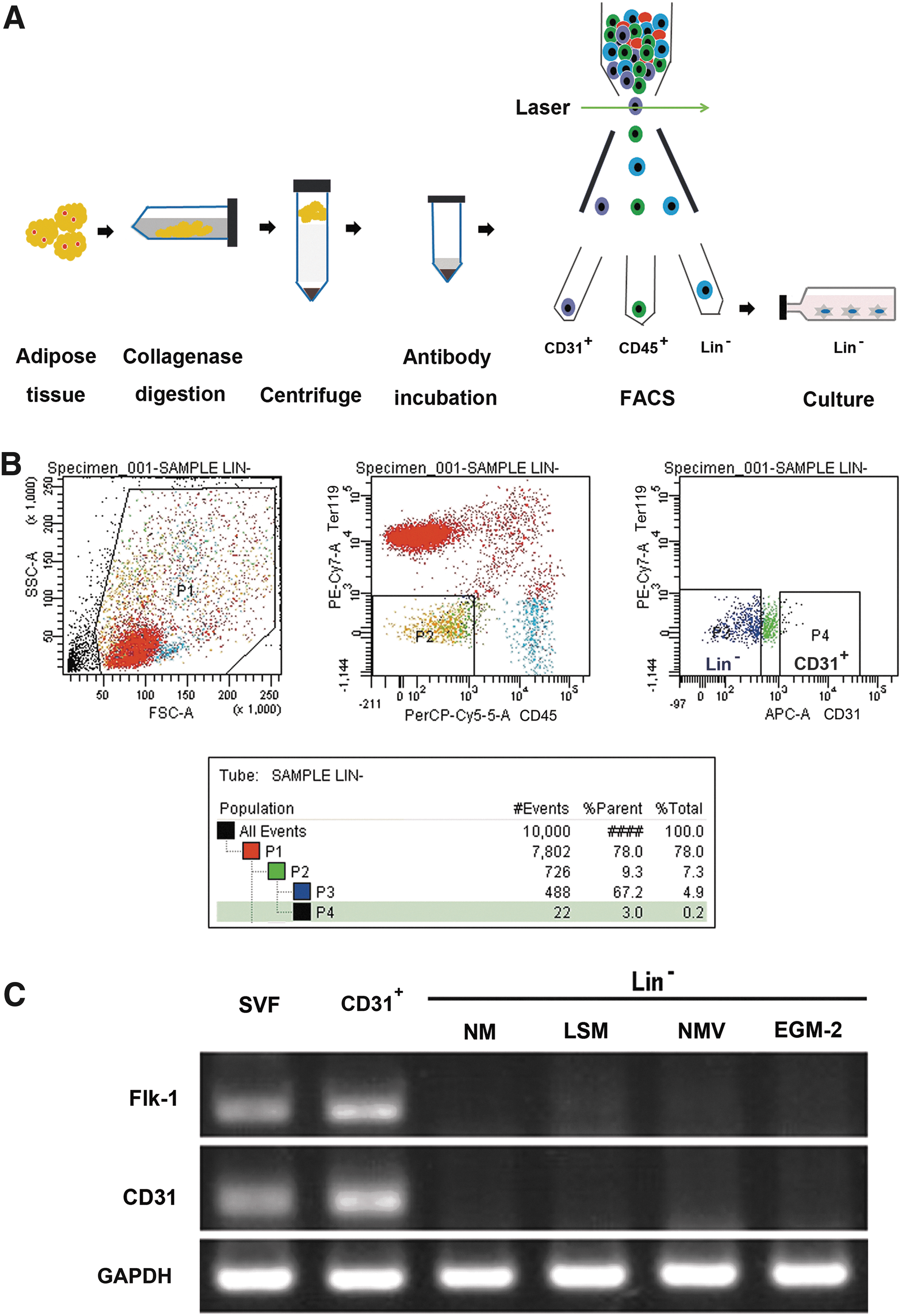
Experimental procedures for the sorting of the stromal-vascular fraction (SVF) from white adipose tissue (WAT) and RT-PCR analysis for endothelial cell markers.
SVF cells express the endothelial cell genes CD31 and Flk1
To determine whether SVF cells also contained endothelial cells, we performed RT-PCR analysis using the endothelial cell markers Flk1 and CD31. We found that SVF cells expressed both CD31 and Flk-1 (Fig. 1C).
Proliferative capacity of Lin− cells in different media in vitro
First, we used NM to culture SVF and Lin− cells. We found that SVF cells in NM appeared senescent and stopped proliferating and growing at day 9 (Fig. 2A). However, Lin− cells could grow for up to 15 days before features of senescence appeared (Fig. 2B).
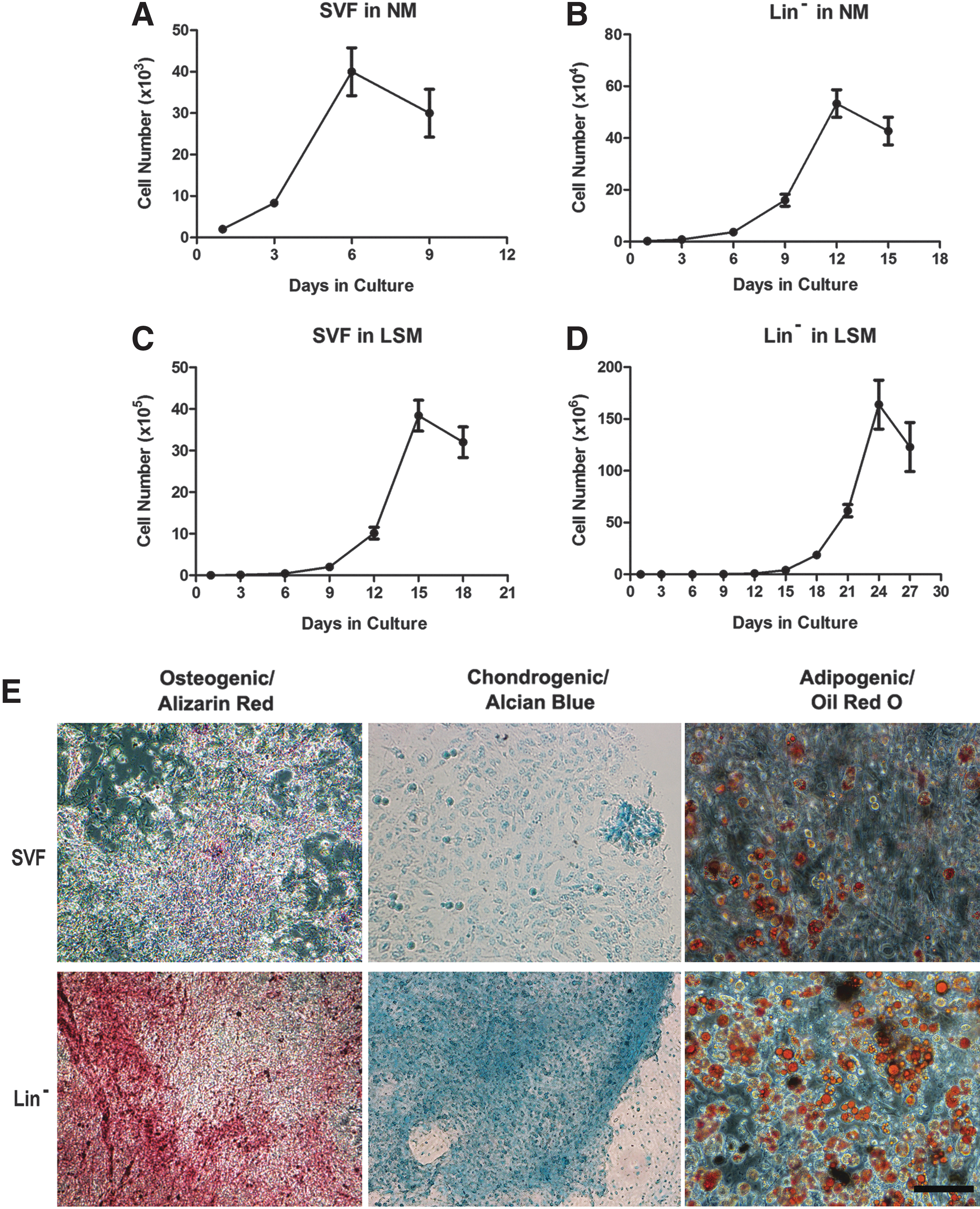
Proliferative capacity in different media and differentiation potentials of Lin− and SVF cells into osteoblasts, chondrocytes, and adipocytes in vitro.
To find a better medium to delay cellular aging, we used the LSM reported in Jiang et al. [22]. We found that SVF cells in the LSM could grow for up to 18 days before displaying aging (Fig. 2C) and that Lin− cells could grow for up to 27 days before they showed senescence (Fig. 2D), suggesting that the latter represent a much purer MSC population with a vigorous capacity for growth and a delay of aging.
Differentiation potentials of Lin− cells into osteogenic, chondrogenic, and adipogenic lineages in vitro
It is well known that MSCs have the capability to differentiate into adipogenic, osteogenic, and chondrogenic lineages in vitro. To determine and verify the multilineage differentiation potentials of Lin− cells, we induced them to differentiate toward osteoblasts, chondrocytes, and adipocytes, and then performed related histological stainings, including Alizarin red, Alcian blue, and oil red O. As shown in Fig. 2E, the Lin− cells demonstrated a much higher differentiation capability toward the three cell lineages related to MSCs (Fig. 2E, bottom row) than did the SVF (Fig. 2E, top row).
Surface marker expression of Lin− cells
To characterize the Lin− cells, we used general MSC markers, namely CD140a, CD140b, CD146, CD105, CD13, CD73, CD44, and Sca-1; a mouse MSC marker, CD34; the histocompatibility complex (MHC) markers MHC-1 and MHC-2; an adipocyte progenitor marker, CD24; and a hematopoietic cell marker, CD45. Lin− cells more highly expressed MSC markers, including CD13, CD140a, CD34, CD140b, CD44, Sca-1, CD146, CD105, and CD73, than SVF cells (Fig. 3). It is important to note that SVF cells expressed more CD24, MHC-2, and CD45 marker than did Lin− cells, suggesting that Lin− cells represent a much purer MSC population than the SVF cells.

Surface marker characterization of the Lin− cells.
Lin− cells promote recovery of hindlimb ischemia
To investigate whether Lin− cells can behave like MSCs to promote recovery from hindlimb ischemia, as previously reported [23], we established a mouse model of unilateral hindlimb ischemia, and then performed cell transplantation as previously described [23,24]. Because previous experiments had reported the recovery effects by implantation of SVFs, and considering that SVF cells contain both CD31 endothelial cells and Lin− cells, we separated these two types of cells and assessed their effects separately in the hindlimb ischemia model. Our results indicated that both Lin− and CD31+ cells could promote recovery of hindlimb ischemia at days 3, 6, and 9, but the control animals failed to show any improvement (Fig. 4).
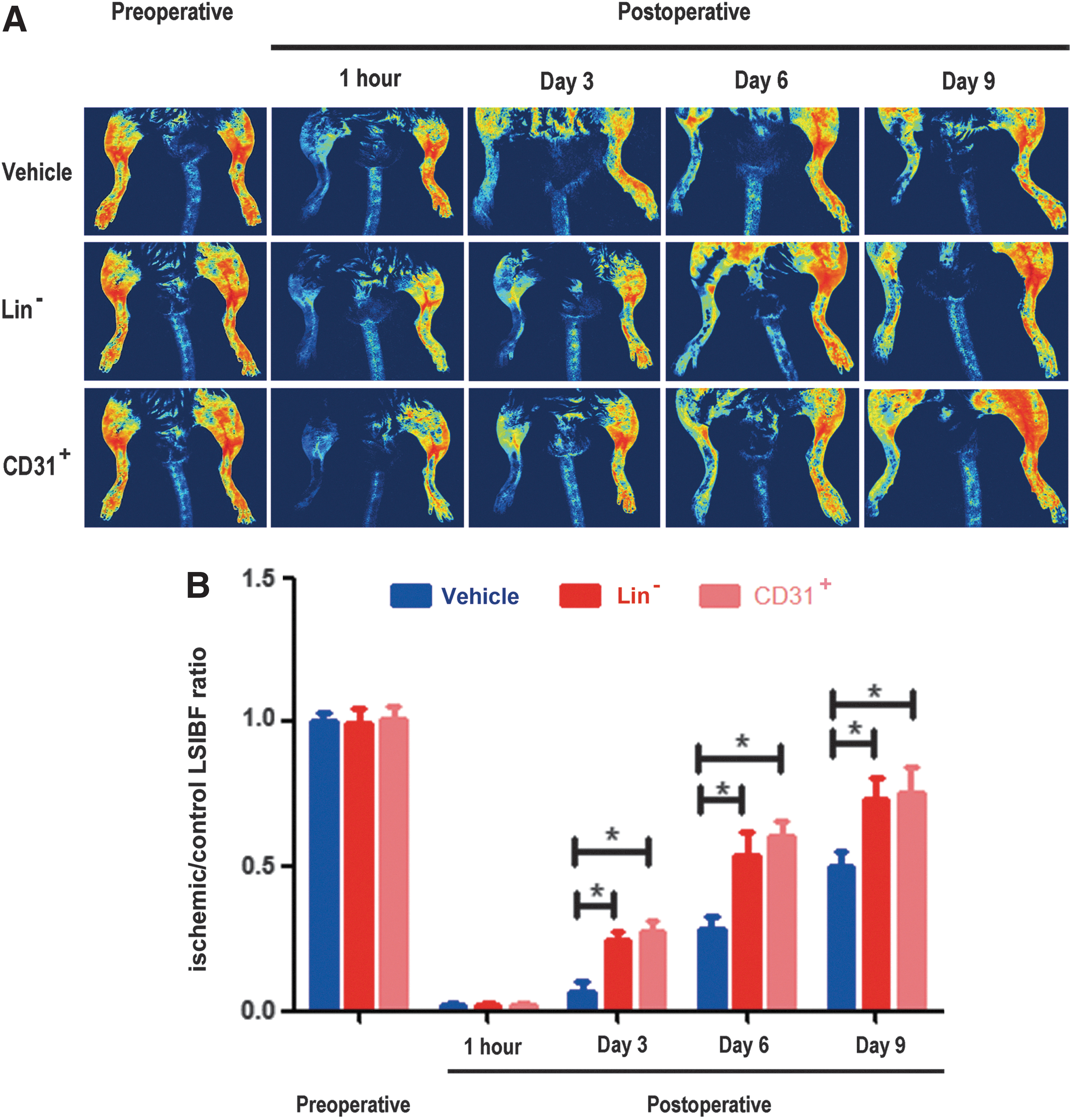
Lin− cells promote recovery from hindlimb ischemia.
Lin− cells did not differentiate into endothelial cells in vitro or in vivo
First, we studied whether Lin− cells could differentiate into endothelial cells using special medium in vitro, including NMV and EGM-2 medium, and NM and LSM as controls. After 7 days, cells were subjected to RT-PCR. The Lin− cells never expressed the endothelial cell markers Flk1 and CD31 under any of these culture conditions. In sharp contrast, freshly sorted CD31+ cells from the SVF cells expressed high levels of these markers (Fig. 1C).
Next, we sought to determine whether Lin− cells could differentiate into endothelial cells in vivo. We obtained RFP-labeled Lin− cells from RFP transgenic mice, and then injected them into the unilateral ischemic hindlimb in wild-type mice. Fifteen days post-transplantation, we found that the Lin− cells in the ischemic hindlimb did not express the endothelial cell markers CD31 (Fig. 5A1–A4) or Flk-1 (Fig. 5B1–B4). However, immunohistochemical staining using an anti-VEGF antibody revealed that these cells produced an angiogenic factor, VEGF (Fig. 5C1–C4), which appears to be the driving force for the new blood vessel formation and improvement of ischemic injury.
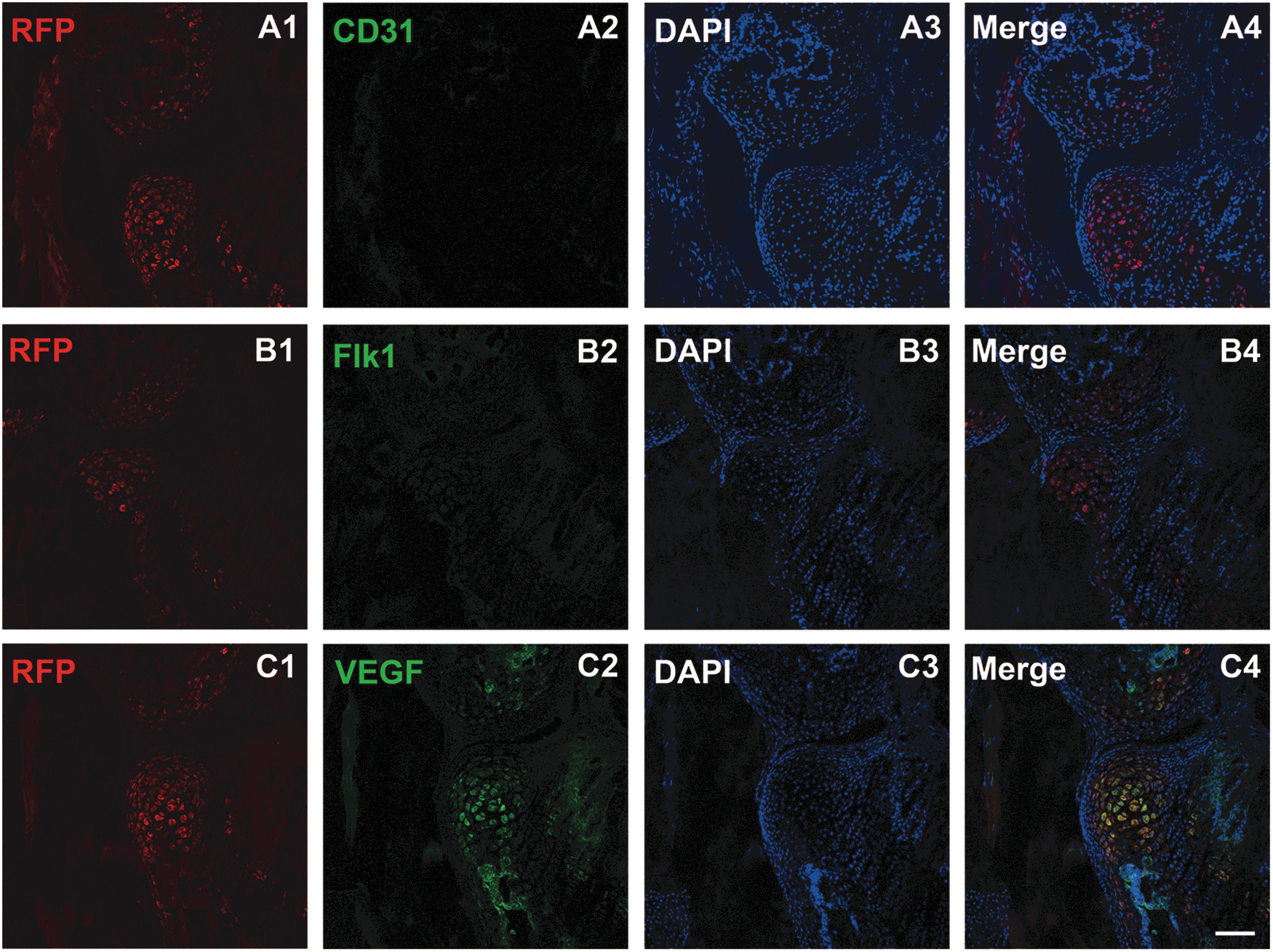
Lin− cells do not differentiate into endothelial cells after being transplanted into the ischemic hindlimb, but they express VEGF. Lin− cells derived from RFP mice did not express the endothelial cell markers CD31
Discussion
Although it is thought that adipose tissue-derived MSCs are likely an attractive cell population with potential clinical applications owing to their regenerative capability and ease of harvest, there has not been a purification method for obtaining fresh MSCs from adipose tissue. In fact, the current method of isolation of MSCs often relies on the adherence of cultured SVF cells from adipose tissue. However, it is well-known that adipose-derived SVF is composed of heterogeneous cell populations, including adipocyte precursor cells, endothelial cells, pericytes, multipotent MSCs, erythrocytes, and hematopoietic cells [7,15 –17]. The present study is the first to fully characterize the adipose-derived Lin− cells and to demonstrate clearly that these cells are enriched MSCs based on their expression of cell-type-specific markers and that they offer much purer MSCs than the SVF cells.
Our serial cell proliferation and differentiation analyses support the notion that the adipose tissue-derived Lin− cells are purer MSCs than the SVF cells. First, we found that, while SVF cells in NM appeared senescent at early stages, as described in previous studies [28], the Lin− cells proliferated for much longer before they showed signs of aging. Jiang et al. reported a type of modified LSM, which was able to maintain longer-term proliferation of mouse BMSCs in vitro [22]. Here we used that medium and found that, even in the same LSM, SVF cells grew for up to 18 days before showing aging, whereas the Lin− cells could grow for up to 27 days before becoming senescent. Second, the Lin− cells showed higher expression of MSC markers, such as CD13, CD140a, CD140b, CD44, Sca-1, CD146, CD105, and CD73. Furthermore, the Lin− cell population is able to undergo differentiation into more osteoblasts, chondrocytes and adipocytes than SVF cells.
It is interesting to note that a recent report by Sugii et al. shows that an adipose-derived Lin− cell population could be reprogrammed to iPS cells with substantially higher efficiencies than from human and mouse fibroblasts. Furthermore, they found that these cells intrinsically express high levels of factors, such as basic fibroblast growth factor, TGF-β, fibronectin, and vitronectin [7]. These findings support the notion that Lin− cells are enriched for MSCs.
It is important to note that some studies have shown that the adipose-derived SVF cells could differentiate into endothelial cells in vitro when exposed to specific growth factors [29 –31]. In fact, the SVF contained endothelial cells. Here we removed existing endothelial cells from the Lin− cell population and found that Lin− cells do not differentiate into endothelial cells either in vitro under any experimental conditions examined or in vivo after the transplantation of Lin− cells prepared from RFP transgenic mice into wild-type ischemic hindlimb. Although both SVF cells and the Lin− cells are able to improve the recovery of hindlimb ischemia, the mechanisms of action are different. While the former appears to enhance angiogenesis presumably due to direct participation in the formation of new blood vessels [19 –21], the latter contributes to the improvement of hindlimb ischemia likely via production of a specific angiogenic factor, VEGF, which in turn promotes angiogenesis. On the other hand, MSCs exposed to an inflammatory microenvironment can downregulate many immune effector functions [3], so we considered Lin− cells to be enriched in MSCs.
In conclusion, adipose tissue-derived Lin− cells not only have more sustained proliferative capacity and differentiation ability into osteogenic, chondrogenic and adipogenic cells in vitro but also express greater amounts of MSC markers than SVF cells. In addition, these cells show no endothelial cell phenotypes. They promote recovery of limb ischemia via production of a specific angiogenic factor, VEGF, after cell transplantation in vivo. Thus, our study demonstrates that the Lin− cell population is enriched for MSCs and provides a reliable way of isolating purer MSCs than SVF cells from adipose tissue.
Footnotes
Acknowledgments
The study is supported by funds from the Chinese Ministry of Science and Technology (2012CB966800, 2013CB945600 and 2012CB967903 to W-Q Gao; 2009CB941101 to J-S Li), the National Natural Science Foundation of China (81130038 and 81372189 to W-Q Gao; 31225017 to J-S Li), the Science and Technology Commission of Shanghai Municipality (Pujiang program to W-Q Gao), the Shanghai Education Committee Key Disciplines and Specialties Foundation (J50208 to W-Q Gao), the Shanghai Health Bureau Key Disciplines and Specialties Foundation and the KC Wong foundation to W-Q Gao; the Chinese Academy of Sciences (XDA01010403 to J-S Li), the Shanghai Municipal Commission for Science and Technology (12JC1409600 and 13XD1404000 to J-S Li); the Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences (2010KIP502 to to Y-R Qin). We thank Professor Jiarui Wu of the Institute of Biochemistry and Cell Biology, Chinese Academy of Sciences, for providing us with 3T3-L1 cells.
Author Disclosure Statement
The authors delare that there is no conflict of interest that would prejudice the impartiality of this scientific work.