
Abstract
Cardiac cell replacement therapy by using human embryonic stem cell (hESC) derivatives remains a potential approach to regenerate myocardium. The major hurdles to clinical application of this technology are immunogenicity and post-transplantation cell death. Here we examined the effects of calcineurin-targeting immunosuppressants cyclosporine A (CsA) and FK506, as well as rapamycin and a selective inhibitor of calcineurin-binding downstream nuclear factor of activated T-cell (NFAT) transcription factor VIVIT on the proliferative activity, function, and survival of hESC-derived cardiomyocytes (hESC-CM) and endothelial cells (hESC-EC) in culture. As shown by automated microscopy, treatments with CsA, FK506, and rapamycin all decreased proliferation, reducing the percentage of hESC-CM and hESC-EC with the mitotic marker Ki67+ by as much as 60% and 74%, respectively. Administration of the cell permeable analogue 11R-VIVIT protein did not modulate their proliferative activity. All immunosuppressants reversed the proapoptotic effect of chelerythrine in hESC-CM demonstrating an inhibitory role of calcineurin/NFAT and mammalian target of rapamycin (mTOR) pathways in hESC-CM survival (using apoptotic marker caspase-3), whereas the protection was less obvious in hESC-EC exposed to H2O2. Immunosuppressants did not affect cell viability in hESC-EC. Our results show that immunosuppressants reduce proliferation, while offsetting cell loss to a smaller extent by reduction in apoptosis of hESC-CM. Immunosuppressant therapy would be compatible with stem cell transplantation, but the resulting reduction in graft expansion capabilities would potentially necessitate implantation of increased cell numbers when immunosuppressants are given. The effects of NFAT-binding immunosuppressant molecules, which do not affect hESC-CM proliferation, may point the way forward for new classes of compounds better suited to cell implantation.
Introduction
D
In this study, we examined the effects of immunosuppressants on hESC-CM and hESC-EC in vitro to predict their survival and function after transplantation combined with immunosuppressive treatment. These compounds include calcineurin inhibitors—cyclosporine A (CsA) and tacrolimus (FK506), mammalian target of rapamycin (mTOR) inhibitor rapamycin and the nuclear factor of activated T-cell (NFAT) inhibitor VIVIT. Our hypothesis was that the use of these agents may modulate the growth, survival, and proliferative activity of cardiovascular derivatives of hESC because of their effect on known signaling pathways within the primary cardiomyocytes or endothelial cells.
Materials and Methods
Differentiation, isolation, and treatment of hESC-CM
As reported earlier [9], cardiomyocytes were derived from the hESC line H7, which was grown on growth factor-reduced Matrigel (BD Biosciences)-coated plates with the mouse embryonic fibroblast-conditioned medium, supplemented with 8 ng/mL recombinant basic human fibroblast growth factor. Cells were generated from dense hESC monolayers, which were treated with human recombinant Activin A (100 ng/mL) (day 0–1) and bone morphogenetic protein 4 (10 ng/mL) (day 1–5) in the serum-free RMPI-B27 medium (Sigma). At day 30, following dissociation of spontaneously beating clusters or monolayers into single cells, cells were seeded onto gelatinized dishes and subjected to treatment after overnight attachment in the differentiation medium (after 3 days in cell death experiments). Cells were treated with CsA (0.2 μM; Sigma), FK506 (122 nM; Sigma), rapamycin (10 ng/mL, 11 nM; Sigma), 11R-VIVIT (2 μM; Calbiochem) [10], or DMSO for 24 h. The selected drug concentrations were all similar to clinically adjusted ones for transplanted patients.
Differentiation and treatments of hESC-EC
Differentiation of hESC into hESC-EC was carried out as described previously [11]. Briefly, hESC colonies were dissociated into embryoid bodies and plated on ultralow attachment plates with the endothelial growth medium-2 (Lonza-EGM2), including 2% fetal bovine serum. Embryoid bodies were then dispersed into single cells and were sorted using the anti-CD31 antibody (BD Biosciences) on a FACSAria II cell sorter (BD Biosciences) and expanded in the Lonza-EGM2 medium for further use. Cells were treated with CsA, FK506, rapamycin, 11R-VIVIT, or DMSO for 24 h. HUVECs were a gift from Caroline Wheeler-Jones (Royal Veterinary College, London) and were maintained in an identical Lonza-EGM2 medium.
Plasmids and transfection
To inhibit the NFAT/calcineurin pathway with 200 nM dnNFATc1 or pGFP-VIVIT [12], hESC-CM were transfected on day 1. Results were confirmed by treatment with the cell-permeable peptide, 11R-VIVIT, or DMSO. An additional 1 mL of media was replaced and cells were seeded for transfection. 11R-VIVIT was also replenished as it has been reported to have a half-life of 30 h. For transfection, Fugene HD (Roche) was used according to the manufacturer's instructions. Where indicated, 1 μg of pGFP-VIVIT or the control vectors pGFP or pBj-stop (Clontech) was added. To determine the effect of hypertrophic G-protein coupled receptor agonist, hESC-CM were treated with 10 μM α-adrenergic phenylephrine (PE) for 48 h in the differentiation medium in the presence or absence of immunosuppressants.
Immunocytochemistry and automated high-content imaging
Cell were fixed with 4% paraformaldehyde, permeabilized with 0.2% Triton X-100, and labeled with anti-Ki67 (Abcam; 1:100), anti-caspase-3 (Abcam; 1:250), anti-NFAT1c (Abcam; 1:100), and anti-myosin heavy chain α/β (MHC α/β, clone 3–48, Abcam; 1:200) primary antibodies. Primary antibodies were detected with Alexa 488-conjugated (Invitrogen) and Alexa 546-conjugated (Invitrogen) secondary antibodies (1:400). DNA was visualized with Hoechst 33343 (Sigma; 0.5 μg/mL). Images were acquired on Zeiss Axio Observer Z1 fluorescence microscopy. Percentage of cardiomyocytes in all experiments was evaluated by parallel immunocytochemistry for MHC α/β and ANF on fixed cells using automated high-content microscopy. The average yield of cardiomyocyte differentiation was 40%; this has been further increased by manually dissecting and replating only beating areas. Only double-positive (MHC/ANF) cells were gated and analyzed for size and sarcomere alignment. Cells after 30 days of differentiation are checked and used for the study. For fixed apoptosis assay, cells were treated with BOBO-1 or TOTO-3 (1:1,000) (Invitrogen) and Hoechst 33342 (1:1,000) for 10 min before fixation and further MHC and caspase-3 staining. Live imaging of hESC-EC cultures consisted of staining the cells with Vybrant® FAM caspase-3 and -7, followed by BOBO-1 (when cells are fixed) or TOTO-3, Hoechst, and TMRM staining (all from Invitrogen). Cells deposited into 96-well plates were scanned on ArrayScan™ VTi automated microscopy and image analysis system (Cellomics, Inc.) using modified Target Activation, Cell Cycle, Morphology Explorer, Spot Detector, and Compartmental Analysis Bioapplication protocols. Using the system of automated highly sensitive fluorescence imaging microscope with 10×objective and suitable filter sets, the stained cells were identified with Hoechst in fluorescence channel 1. Each treatment was tested in triplicates or sextuplicates in cell death assay and the experiments were repeated up to three times. In each well, an average of 1,500 cells was analyzed.
Cell death assay
Cells were plated in gelatinized 96-well plates at 50,000 cells per cm2 in EGM2 or RPMI+B27 for endothelial cells and cardiomyocytes, respectively. Drugs were added 3 days after plating. Cell death inducers used for positive control or protection experiments included H2O2 (100 μM) for 24 h and chelerythrine (CHE; 10 μM) for 90 min (both from Sigma).
Colony formation assay for hESC-CM
Cultures of hESC-CM were dissected into single cells, filtered through a 40-μm cell strainer (BD Falcon), and 6,000 isolated cells were plated on gelatin into each well of 96-well plates. After 1 day recovery, cells were treated with immunosuppressants up to 10 days. Plates were fixed and stained for MHC and Hoechst at day 0, 2, 4, 6, 8, and 10. The colony formation activity and the number of nuclei per anti-MHC-positive colony using were assessed by using a Colony Formation BioApplication on Cellomics platform.
Angiogenesis assays
Fifty microliters/well of growth factor-reduced Matrigel was added to 96-well plates and allowed to solidify in a thin layer at 37°C. CD31+ cells (10,000 per well) were seeded onto the gels and tubes were photographed after 4–6 h. VEGF-A (50 ng/mL; PeproTech) was used as a positive angiogenic stimulus.
Beating activity of hESC-CM clusters
Beating hESC-CM clusters were superfused with the Krebs–Henseleit solution containing 2 mM CaCl2, continuously bubbled with 5% CO2/95% O2, at a rate of 2 mL/min. The hESC-CM were tracked as small clusters using the IonOptix video-edge detection system and a Nikon TE2000 microscope. The spontaneous beating rate was recorded. After 20-min equilibration, measurements were made in the presence of CsA, FK506, rapamycin, or 11R-VIVIT for 20 min. To assess whether drugs have a proarrhythmic effect, we compared variances of the beating rate before, during, and after treatment. The beating rate is calculated by measuring the time between two consecutive peaks.
Statistics
Results are expressed as mean±SEM. The data were analyzed by the unpaired Student's t-test or one-way analysis of variance and Tukey's post hoc test for multiple comparisons. Differences at the level of P<0.05 were considered statistically significant. For cell death assay, a cutoff point based on nuclear caspase-3 and TOTO-3 or BOBO-1 fluorescence intensity was derived in control cells, where a small subset of cells are expected to be either in the apoptotic or necrotic state. Cells with high caspase-3 and BOBO-1 or TOTO-3 intensities were defined as cells with fluorescence intensity greater than the intensity threshold that discriminates the 5% of control cells with the greatest fluorescence intensity. This threshold value was used as a reference to determine apoptotic and necrosis in other groups. An apoptotic population was defined as caspase-3 or caspase FAM positive; A necrotic population was defined as caspase-3 negative and BOBO-1 positive. Nuclear remodeling (small nuclear size or pyknosis defined as small Hoechst area) and mitochondrial dysfunction (low TMRM intensity) were determined similarly. Data were expressed relative to threshold value and maximum (100% cells being positive) using the formula: cell death (or apoptotic or necrotic) index=[(% positive-threshold)/(100%-threshold)]. Based on image observation and data published elsewhere [13,14], the percentage of apoptotic and necrotic cells in control (the threshold value) was typically 5%. Cell loss was estimated from the difference in cell number between control and conditions. For assessment of proarrhythmic effects, the Levene's test was used to compare the equality of variances between treated and baseline samples.
Results
Immunosuppressants block proliferation of hESC-CM and hESC-EC
Cardiomyocytes derived from hESC were identified by cardiac-specific markers, including MHC α/β and ANF. Pilot data showed that the estimated doubling time of hESC-CM in culture was low (6–8 days) compared with those in endothelial cells (2–3 days). The proliferative activity of hESC-CM was assessed by Ki67, a marker expressed in all phases of the cell cycle except G0. Control populations maintained in 20% fetal calf serum (FCS) showed 35.63%±1.40% of 1-month-old hESC-CM expressing Ki67. Immunosuppressants were used at clinically relevant concentrations, used in transplanted patients. After 24 h of treatment with CsA, the proportion of hESC-CM expressing Ki67 decreased by 56% (P<0.001 vs. control, Fig. 1A) and FK506 or rapamycin had similar effects. The cell number of hESC-CM population was decreased accordingly (Fig. 1B). Using combinatorial library, high-affinity calcineurin binding peptide VIVIT was identified as an inhibitor of NFAT activation [15]. VIVIT acts through a calcineurin docking motif site without affecting calcineurin-mediated non-NFAT-dependent expressions or without disrupting other calcineurin-dependent pathways. Cell permeable 11R-VIVIT resulted in a 40% decrease in nuclear translocation of NFAT (P<0.05, Supplementary Fig. S1; Supplementary Data are available online at
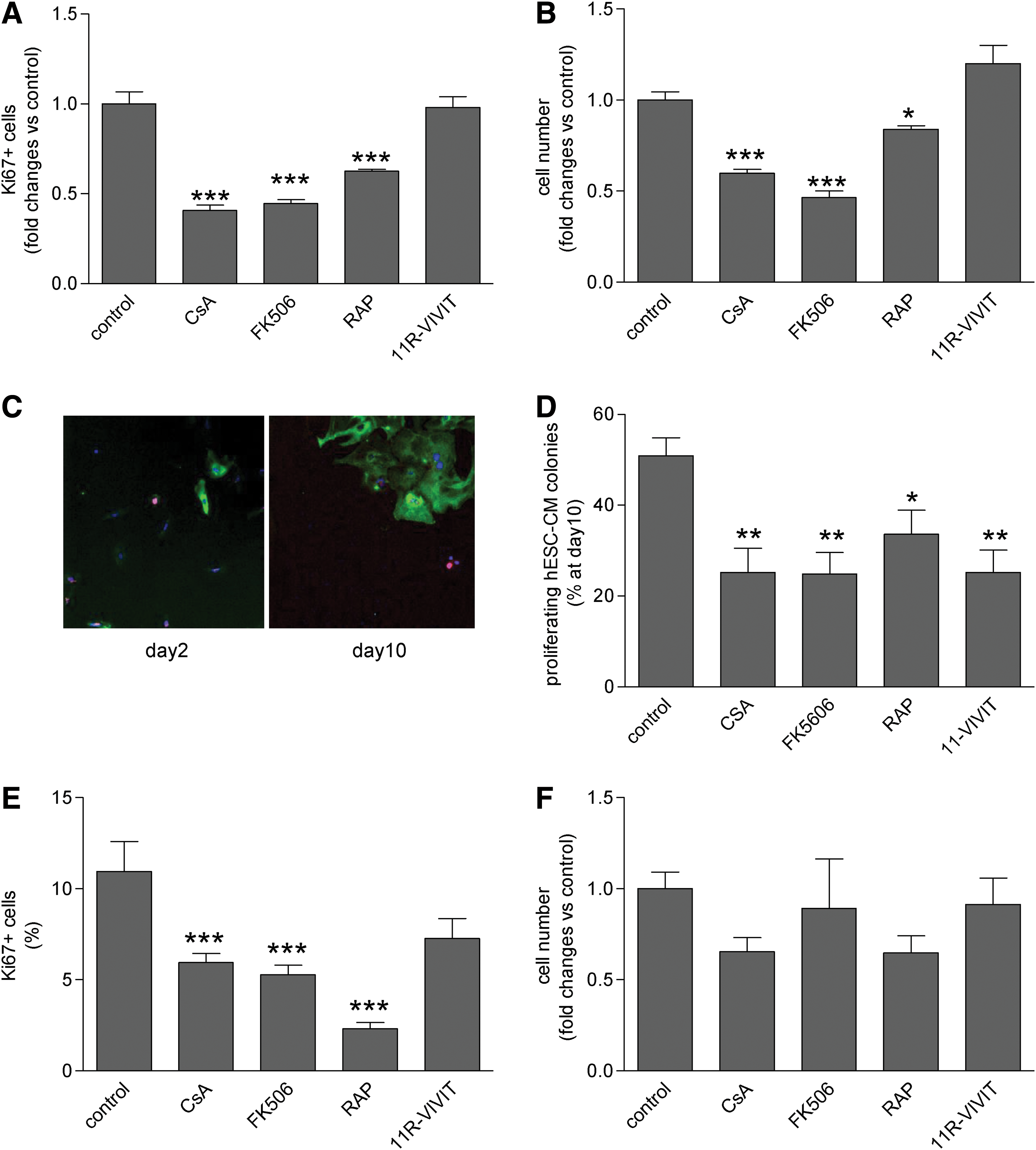
Effect of immunosuppressive agents on proliferation and cell number of human embryonic stem cell-derived cardiomyocytes (hESC-CM), human embryonic stem cell-derived endothelial cells (hESC-EC).
Immunosuppressants modulate viability and cell death of hESC derivatives
To test the cell death modulating effects of immunosuppressants, hESC-CM were incubated with CHE, a potent inducer of apoptosis in cardiomyocytes through the generation of intracellular ROS [16]. Cell death was assessed by immunodetection of active caspase-3, necrosis marker BOBO-1, and nuclear condensation (Figs. 2 and 3). In hESC-CM, all immunosuppressants reduced CHE-induced apoptosis (Fig. 3A). The necrosis index stayed low in response to CHE and was modestly increased by adding FK506 and rapamycin (Fig. 3B) (both P<0.01). FK506 and rapamycin reduced CHE-induced nuclear remodeling (Fig. 3C). To induce cell death in endothelial cells, hESC-EC were incubated with hydrogen peroxide (H2O2), since using CHE as an indirect inducer of oxidative stress had no effect on apoptosis in hESC-EC (Supplementary Fig. S3). As shown by high-content automated microscopy, H2O2 resulted in a dose-dependent increase in apoptosis and necrosis of hESC-EC with a concomitant decrease in levels of mitochondrial status marker TMRM (Supplementary Fig. S4). Rapamycin and 11R-VIVIT reduced H2O2-induced apoptosis (Fig. 3D) and necrosis (Fig. 3E), but not nuclear remodeling (Fig. 3F). Rapamycin had a very modest although significant effect on baseline apoptosis and nuclear remodeling (Supplementary Fig. S5), while the other drugs did not show any toxic effects on endothelial cells.

Representative immunofluorescence images of human embryonic stem-derived cardiomyocytes (hESC-CM). hESC-CM were analyzed after 24 h of treatment with cyclosporine A (CsA, 0.2 μM), FK-506 (122 nM), rapamycin (RAP, 10 ng/mL), or 11R-VIVIT (0.2 μM) with or without chelerythrine (CHE), as described in Methods. Cells stained for MHC α/β (with secondary antibody Alexa568), Hoechst 33342, anti-caspase-3 (with secondary antibody Alexa 647), and BOBO-1. Gray arrows show representative necrotic (BOBO-1-positive) or apoptotic (caspase-3-positive) cells. Color images available online at
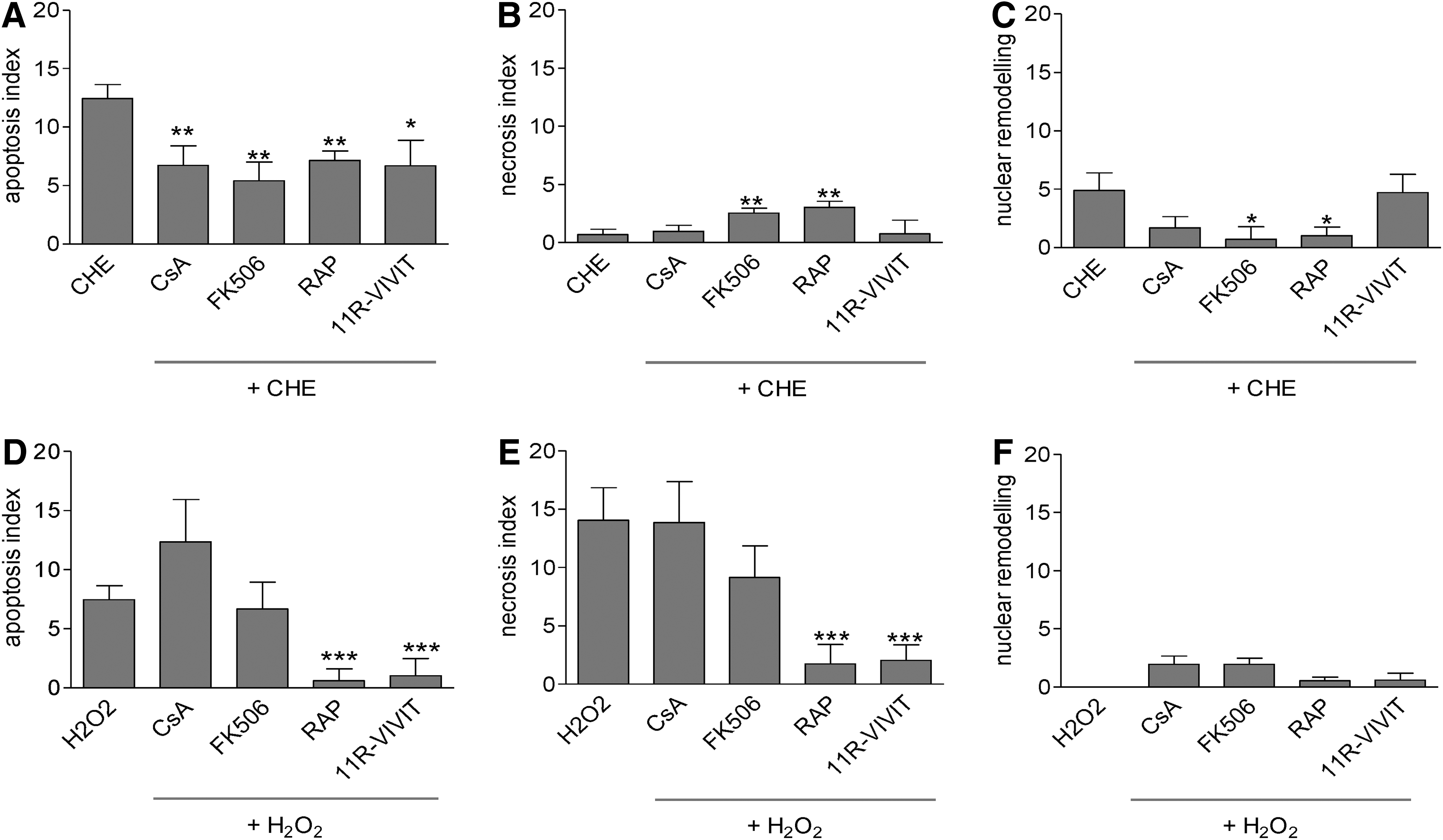
Effect of immunosuppressant pretreatment on apoptosis, necrosis, and nuclear remodeling of hESC-CM and hESC-EC challenged with pro-death stimuli. Effects of pretreatment with CsA, FK506, RAP, and 11R-VIVIT on CHE-induced apoptosis
Immunosuppressants increase hESC-EC angiogenic activity in vitro
As assessed by tube formation assay, CsA, FK506, and 11R-VIVIT increased the complexity of endothelial tubes formed as seen by the number of nodes with effects similar to those with the positive control, VEGF, although the total tube length was not significantly increased (Fig. 4A–C). Rapamycin had no effect on tube formation activity in vitro.
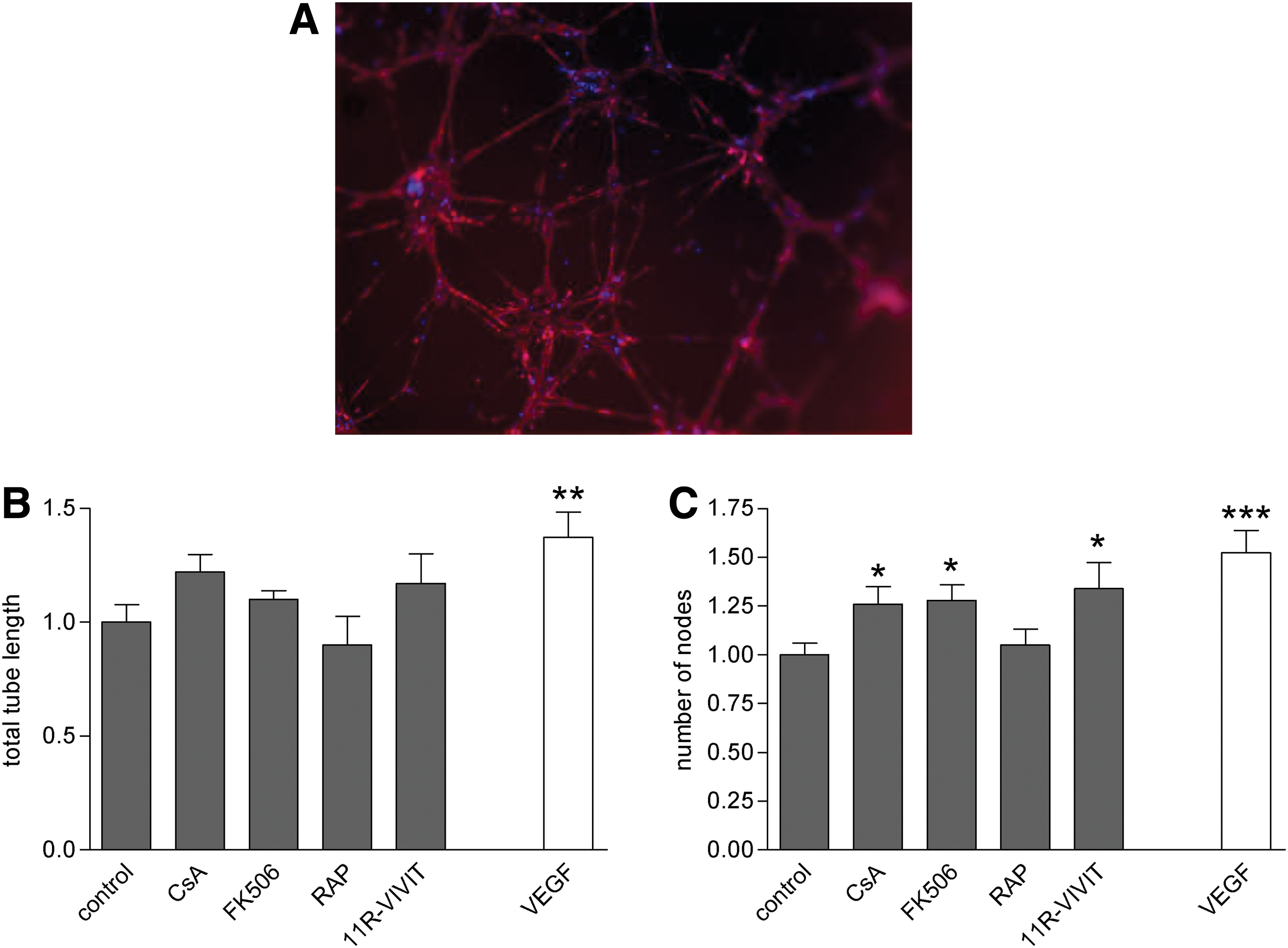
Matrigel tube formation assays in immunosuppressant-treated hESC-EC.
Immunosuppressants block PE-induced hESC-CM growth through NFAT pathway
Our earlier data showed that immunosuppressive agents CsA, FK506, and rapamycin significantly reduced PE-induced cellular growth and hypertrophy of hESC-CM [9]. To test the role of NFAT-mediated signals of immunosuppressants in this process, hESC-CM were transfected with a GFP-VIVIT construct, dominant negative form of NFAT1c, or administration of cell permeable 11R-VIVIT protein, selective inhibitors of calcineurin-NFAT interaction or NFAT, without affecting calcineurin phosphatase activity. Inhibition of NFAT signaling by 11R-VIVIT, overexpression of GFP-VIVIT, and the dominant negative form of NFAT1c all blocked PE-induced hypertrophy (P<0.001) (Fig. 5C). Of interest, PE-induced ANF mRNA levels remained high in response to 11R-VIVIT (Fig. 5D). Direct inhibition of NFAT signaling did not affect the proliferative activity of hESC-CM (expressed as Ki67+ and cell number) (Fig. 5A, B).
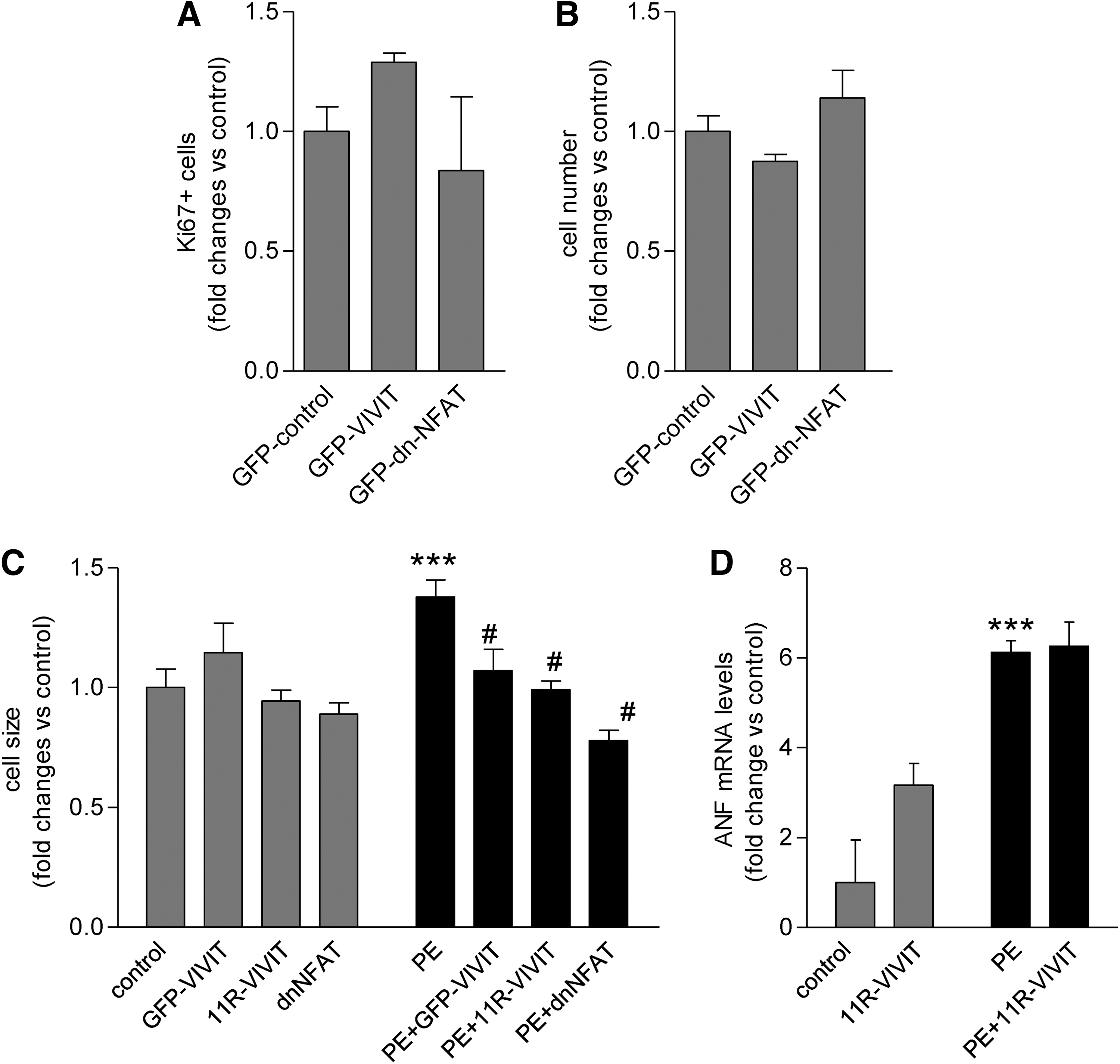
Inhibition of NFAT transcription factor modulates phenylephrine (PE)-induced cell growth and hypertrophy of hESC-CM.
Immunosuppressants have no effect on hESC-CM beating activity
Next, we assessed immunosuppressive drugs for their ability to alter contractile function on clusters of hESC-CM. After a 20-min incubation, none of the immunosuppressants modified the beating rate of hESC-CM compared with baseline (Supplementary Fig. S6). However, immunosuppressants could induce irregularities in the beating activity both during and after treatment, as evidenced by the increased variance of beating rate (for FK506 treatment, P<0.01).
Discussion
An important obstacle preceding therapeutic application of differentiated hESC progeny will be to prevent immune rejection and ensure long-term stability of the allogeneic graft. This study was carried out with a view to add information on the extent to which hESC derivatives in combination with immunosuppressive therapies are suitable for cardiac grafting in terms of their survival capabilities in a hostile environment, their proliferative activity, and maturation.
Targeting calcineurin/NFAT and mTOR pathways by immunosuppressants are the primary mechanisms for blocking immune interleukin activation. These signaling molecules are also present in nonimmune cells and are central in embryonic development, cell proliferation, growth, motility, survival, and metabolism. The outcome of the activation or inhibition of these molecules entangled in a complex signaling network is not easily predictable and seems to be cell type- and context-dependent [17]. Furthermore, immunosuppressive drugs can have multiple molecular targets, which obscure the understanding of these pathways.
The lack of regenerative capabilities of cardiac myocytes makes the heart particularly sensitive to cell stressors, in particular, oxidative stress. In contrast to earlier experimental studies in adult rat cardiomyocytes [18], we showed here that hESC-CM exposed to immunosuppressants at clinically relevant concentrations do not undergo apoptosis or necrosis. In addition to inhibiting calcineurin, CsA has been shown to elevate intracellular calcium [19] and induce oxidative stress [20], both mediating intrinsic apoptosis pathway in adult cardiomyocytes [21]. hESC-CM are relatively immature cardiomyocytes in terms of their bioenergetic circuits and morphology [22]. Their gene and protein expression profiles suggest that they are more related to their fetal rather than their adult counterparts. Given that adult cardiomyocytes are particularly sensitive to oxidative stress due to their low levels of intracellular antioxidants [23], hESC-CM may be naturally resistant to CsA due to their relatively immature phenotype. Neither FK506 nor 11R-VIVIT, a more specific inhibitor of the calcineurin/NFAT pathway induced cardiomyocyte death. Together, our data suggest that the inhibition of the calcineurin/NFAT pathway has no toxic effect on hESC-CM. The inhibition of mTOR by rapamycin has been extensively described to induce autophagy [24], a mechanism seen as a protective strategy for starved or stressed cells, but that can also lead to cell death. Under control conditions, blockade of mTOR did not affect cardiomyocyte viability. Although rapamycin did induce significant levels of cell toxicity in hESC-EC, as shown for endothelial cells in earlier reports, the effects were modest [25]. Similarly to hESC-CM, calcineurin inhibitor CsA, FK506, and 11R-VIVIT did not affect viability of hESC-EC.
After transplantation, stem cell derivatives will be exposed to a hostile environment with inflammatory cytokines, neurohormone overdrive, and low oxygenation. We further questioned whether immunosuppressive drugs would sensitize or protect cardiomyocytes and endothelial cells in an environment that promotes cell death, in particular apoptosis. CsA has been shown to protect myocytes against a range of stressors, including reoxygenation injury [26], CHE [27], and doxorubicin [28], all inducers of oxidative stress. CHE induces cardiac myocyte apoptosis through production of reactive oxygen species, possibly H2O2, rather than PKC inhibition, leading to rapid increase in caspase-3 activation [16]. We found that CsA, FK506, and 11R-VIVIT partially blocked CHE-induced caspase-3 activation in hESC-CM. Our data suggest that calcineurin inhibition is directly involved in cell protection against CHE as 11R-VIVIT has no other described mechanism than blocking NFAT activation. However, we cannot exclude additional protective effects such as the prevention of mitochondrial permeability transition pore opening by CsA [29] and mTOR inhibition by FK506 [30]. Rapamycin can also partially protect cardiomyocytes exposed to CHE, suggesting that mTOR inhibition may prevent oxidative stress-mediated cell damage, possibly through induction of protective autophagy. Our results are in accordance with other studies showing that rapamycin has a protective effect against ischemia/reperfusion-induced oxidative stress in mice and adult mouse cardiomyocytes [31]. Both FK506 and rapamycin slightly enhanced CHE-induced necrosis, while reducing apoptosis, but the increase in necrosis did not offset the global protection of these drugs. The effect of CHE was less obvious in hESC-EC, where necrosis predominated, but direct administration of H2O2 induced both apoptosis and necrosis. To obtain a comparable cell death profile to those in cardiomyocytes, we used H2O2 as a cell death inducer in endothelial cells. Protection against oxidative stress was seen with rapamycin and 11R-VIVIT in hESC-EC. In the context of transplantation of allogeneic cells in patients after infarct, where ischemia and oxidative stress may be detrimental for implanted cells, immunosuppressants would actually improve the viability of the cardiac or vascular graft. In transplantation studies, enhanced survival of transplanted hESC-CM or undifferentiated hESC has been shown using CsA [5,32,33]. The same concentration of CsA used here was part of a survival cocktail designed by Laflamme et al. [32] before transplanting hESC-CM to infarcted rat hearts to improve the survival of the cardiac graft.
CsA and FK506 have been shown to alter cell proliferation in different systems. In this study, we found that these agents strongly inhibited hESC-CM proliferation activity and reduced cell number. In addition, CsA and FK506 inhibited proliferation of hESC-EC, although with no change on the actual cell number, possibly because the high turnover of endothelial cells offset relatively low changes in the nonproliferating population. The effect of CsA and FK506 on cell proliferation most likely is independent of NFAT as VIVIT had no effect on cell proliferative activity of hESC-CM on in hESC-EC. Similarly, overexpression of a dominant-negative NFAT construct did not change the Ki67-positive population of hESC-CM, further suggesting that VIVIT therapy may be accompanied by non-NFAT-mediated side effects on calcineurin signaling. This may also suggest that targeting the calcineurin-NFAT interface by VIVIT and analogous approaches, but sparing other calcineurin, merits study as a means to enhance the therapeutic grafting of hESC derivatives, without confounding the benefit of donor cell proliferation. Rapamycin has also been shown to antagonize proliferation [34]; proliferative activities of hESC-CM and hESC-EC showed marked reduction. Rapamycin prevents or delays the entry of these cells into S phase after stimulation by mitogens in other proliferating cell lines. In addition, mTOR is involved in upregulating transcription factors associated with cell proliferation [35]. hESC-CM and hESC-EC responded similarly to rapamycin. A longer term exposure of endothelial cells with rapamycin should be performed to evaluate whether the proliferative activity of hESC-EC will offset the apoptotic effect of rapamycin.
We showed that immunosuppressive agents display further pleiotropic distinct non-immunologic effects on hESC-CM and hESC-EC in vitro. We found that CsA, FK506, rapamycin [9] as well as using a GFP-VIVIT fusion protein and a cell permeable 11R-VIVIT peptide blocked PE-induced hypertrophy of hESC-CM. This is in line with earlier findings showing that VIVIT-GFP inhibited adult cardiomyocyte hypertrophy to PE [36]. Similarly, direct inhibition of NFAT by overexpression of the dominant negative NFAT construct blocked hESC-CM growth. Immunosuppressants had no short-term effect on the overall beating rate of hESC-CM clusters, but the significant increase in variability of beat interval by FK506 could warrant further investigation of a potential arrhythmogenic effect given its interaction with the FK-binding protein in the sarcoplasmic reticulum Ca release channel complex [37]. In contrast to earlier findings reporting that VIVIT only marginally affects endothelial wound healing in vitro [38], our data show that VIVIT induces hESC-EC tube formation capacity, in a comparable extent with those of VEGF-A. Different responsiveness of stem cell-derived or fetal-like endothelial cells as compared with adult endothelial cells may constitute a promising lead in combination of immunosuppressive [10] and revascularization therapy.
Comparison of in vitro results with in vivo data on immunosuppressive regimens can be difficult particularly for the heart. It is well established that even with immune matching, or in immunocompromised animals, the highly kinetic environment of the heart acts against engraftment. Notably, studies that investigate immunosuppressive effects on differentiated hESC derivatives like hESC-CM chose to implant in a site other than the heart [5,33]. Added to this is the effect of ischemia and inflammation on survival of implanted cells, as described above, which may be modulated quite separately by immunosuppressants in a compound-specific manner. The host heart too will be affected by the protective effects of immunosuppressants, which may change the outcome of the experiment in terms of cardiac function. This has not been an issue during whole heart transplantation, but must be considered for cell engraftment during the early injury phase. These complications necessitate the strategy of the present study, to dissect the effects of immunosuppressants in a more reductionist manner.
Summary
Immunosuppressants modulate stem cell-derived cardiovascular cell function and survival with significant consequences. The combination of decreased proliferation and cell growth and the reduced responsiveness to endogenous hypertrophic signals would be predicted to result in a marked reduction in graft expansion capabilities post-transplantation. This would mean that a greater quantity of cells would need to be delivered for clinical benefit. However, this effect may be mitigated by protective effects in the context of cell-based therapy, where these drugs will be added at the time of transplantation. Overall, because cardiomyocyte turnover is low in hESC-CM compared with hESC-EC, we would predict that the increased survival of these cells will offset the antiproliferative properties of immunosuppressants to a greater extent. Potential proangiogenic and arrhythmogenic effects of individual immunosuppressants should be the subject of further study.
Footnotes
Acknowledgments
This work was funded by the British Heart Foundation, Leducq Foundation (G.F., S.E.H.), and Hungarian Scientific Research Fund (OTKA 105555, G.F.).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.