
Abstract
Cellular interactions were investigated between human mesenchymal stem cells (MSC) and human breast cancer cells. Co-culture of the two cell populations was associated with an MSC-mediated growth stimulation of MDA-MB-231 breast cancer cells. A continuous expansion of tumor cell colonies was progressively surrounded by MSCGFP displaying elongated cell bodies. Moreover, some MSCGFP and MDA-MB-231cherry cells spontaneously generated hybrid/chimeric cell populations, demonstrating a dual (green fluorescent protein+cherry) fluorescence. During a co-culture of 5–6 days, MSC also induced expression of the GPI-anchored CD90 molecule in breast cancer cells, which could not be observed in a transwell assay, suggesting the requirement of direct cellular interactions. Indeed, MSC-mediated CD90 induction in the breast cancer cells could be partially blocked by a gap junction inhibitor and by inhibition of the notch signaling pathway, respectively. Similar findings were observed in vivo by which a subcutaneous injection of a co-culture of primary MSC with MDA-MB-231GFP cells into NOD/scid mice exhibited an about 10-fold increased tumor size and enhanced metastatic capacity as compared with the MDA-MB-231GFP mono-culture. Flow cytometric evaluation of the co-culture tumors revealed more than 90% of breast cancer cells with about 3% of CD90-positive cells, also suggesting an MSC-mediated in vivo induction of CD90 in MDA-MB-231 cells. Furthermore, immunohistochemical analysis demonstrated an elevated neovascularization and viability in the MSC/MDA-MB-231GFP-derived tumors. Together, these data suggested an MSC-mediated growth stimulation of breast cancer cells in vitro and in vivo by which the altered MSC morphology and the appearance of hybrid/chimeric cells and breast cancer-expressing CD90+ cells indicate mutual cellular alterations.
Introduction
MSC preferentially reside in perivascular niches of nearly all kinds of human tissues [13,14]. Despite functional alterations according to tissue-specific origins, these heterogenic MSC populations share a variety of common properties such as surface markers, continued proliferative capacity, and the capability to differentiate at least along certain phenotypes of the mesodermal lineage [15 –18]. Moreover, MSC contribute to regulate stem cell homeostasis, modulate immune cell functions [19], support angiogenesis [20,21], and migrate toward damaged or injured tissues to support repair processes [22]. According to this multi-functional plasticity, MSC are considered cellular all-round supporters, by which the MSC exhibit a significant sensitivity to extracellular and intracellular signals [23]. Since intracellular expression levels of several microRNAs contribute to alter the MSC state of activation and susceptibility [24], distinct functions within this unique panel of MSC biodiversity can also be triggered by alterations of the microenvironment such as the threshold of cytokines/chemokines to induce MSC adherence [25], changes in the extracellular matrix composition, and by a direct cell-to-cell contact.
Some events within a neoplastic process of breast epithelial cells may be attributed to the plasticity of MSC to undergo differentiation or to change to a different functional phenotype by transformation. Such an alteration of functional properties provides a specific and continuous support for progressive tumor cell growth. Although MSC and their multi-functional plasticity play an important role in combination with breast cancer cells, data arising from such studies are controversial. A previous work has demonstrated that MSC promote the formation of mammospheres in HMEC and breast cancer cells and support proliferation and survival [26]. In contrast, MSC extracts derived from either MSC lysates or MSC supernatants inhibit cell growth of a variety of cancer cell lines, including breast cancer, ovarian carcinoma, and osteosarcoma cells [27]. Other studies have also documented that MSC can undergo spontaneous malignant transformation in long-term cultures displaying an altered morphology and proliferation rate [28 –30], although data from the last reference [30] have been relativized [31]. In contrast, further investigations have suggested that long-term cultured MSC do not exhibit tumorigenic potential by themselves [32 –34]. Regardless of these controversial studies, bone marrow-derived MSC can be induced to acquire functional properties of carcinoma-associated fibroblasts after culture in tumor-conditioned medium for approximately 30 days [35]. Furthermore, MSC co-culture with breast cancer cells was reported with an up-regulation of EMT-associated genes [36]. In this context, MSC isolated from primary breast cancer tissues significantly promote tumor growth after co-transplantation with breast cancer cells [37]. In addition, MSC display some similarities to aged normal mammary epithelial cells and tumorigenic human breast epithelial cells, particularly after EMT, with regard to the gene expression pattern [38] and some surface receptor levels supporting an association of these stem cells with breast cancer [39 –41]. Furthermore, the close vicinity between MSC and breast cancer cells via adhesion receptors or the formation of gap junctions favors the exchange of exosomes and microRNAs that may contribute toward transforming MSC [23]. Together, culture of MSC and cellular interactions of MSC with breast cancer cells can develop a variety of opposite effects in neoplastic breast tissue, by which the underlying mechanisms remain unclear.
To address this controversy, a cellular interaction model is presented in this study that discriminates between the MSC and human breast cancer cell populations during co-culture in vitro and in vivo and follows their development in the course of tumor progression. Moreover, the involvement of certain signaling pathways, including gap junctional intercellular communication (GJIC) and notch signaling, were investigated during MSC and breast cancer cell interactions.
Materials and Methods
Mesenchymal stroma/stem cells
Human umbilical cords were obtained from three different patients after received written consent after delivery of full-term (38–40 weeks) infants either spontaneously or by Cesarean section, respectively. The use of this material has been approved by the ethics committee of the Hannover Medical School (chairman Prof. Dr. H.D. Tröger), project #3037 on 17th June, 2006 and in an extended permission project #443 on 26th June, 2009.
MSC-like cells were isolated as previously reported [42]. In brief, umbilical cord tissue was washed with phosphate-buffered saline (PBS) to remove blood cells, cut into ∼0.5 cm3 large pieces, and incubated in α minimal essential medium (αMEM) (Invitrogen GmbH, Karlsruhe, Germany) supplemented with 15% of allogeneic human AB-serum [43] (kindly provided by Prof. R. Jacobs, Department of Immunology, Medical University Hannover, Hannover, Germany), 100 U/mL penicillin, 100 μg/mL streptomycin, and 2 mM
Breast cancer cells
Human MDA-MB-231 breast cancer cells were obtained from the ATCC (#HTB-26). The MDA-MB-231 cell line has been analyzed in a short tandem repeat [44]-based authentication by the Institute for Legal Medicine at the University Hospital Schleswig-Holstein as recently documented [45]. MDA-MB-231 cells were cultivated initially at about 1,500 cells/cm2 in Leibovitz's L-15-medium (Invitrogen) with 10% (v/v) fetal calf serum (FCS), 2 mM
Co-culture of human MSC and human MDA-MB-231 breast cancer cells for optimized cell ratios
Initially, 5×103, 2.5×103, 1.25×103, and 6.25×102 MSC were incubated in MSC medium for 3 days in six-well plates, respectively. Thereafter, 104 green fluorescent protein (GFP)-labeled MDA-MB-231 cells were added as co-culture target cells. In order to distinguish between the breast cancer cells and MSC within the co-culture, the MDA-MB-231 cells have been transduced with a third-generation lentiviral self in-activating (SIN) vector containing the eGFP gene according to a similar labeling technique previously described for an ovarian cancer cell line [46]. The GFP-labeled MDA-MB-231 cells cultivated in MSC culture medium as well as in MDA-MB-231 growth medium served as controls. After 4 days of co-culture, the medium was removed and the cells were lysed with 10% sodium dodecyl sulfate (SDS). Lysed cells were harvested with a rubber policeman and, subsequently, homogenized using syringes with 0.6- and 0.4-gauge needles (BD Bioscience GmbH, Heidelberg, Germany), respectively. Afterward, the fluorescence intensities of GFP that corresponded to the appropriate cell number of breast cancer cells was measured in aliquots of the cell homogenates at excitation 485 nm/emission 520 nm using the Fluoroscan Ascent Fl (Thermo Fisher Scientific, Waltham, MA).
Transwell co-culture of human MSC and human MDA-MB-231GFP breast cancer cells
About 104 of MDA-MB-231GFP breast cancer cells were cultured in six-well plates with 3 mL of MSC culture medium. Afterward, transwells carrying a sterile track-etched membrane with 0.4 μm sized-pores were applied with either MSC culture medium for single population culture of the breast cancer cells or 1.7×104 MSC for co-culture were added onto the transwell membrane. After 4 days of co-culture, the medium was removed and the MDA-MB-231GFP cells were lysed with 10% SDS. The fluorescence signal of lysed cells was measured in aliquots of the cell homogenates at 485 nm using the Fluoroscan Ascent Fl (Thermo Fisher Scientific).
Direct and long-term in vitro co-culture of human MSC and human MDA-MB-231 breast cancer cells
Based on an optimized cell ratio, a co-culture of 60% GFP-labeled human UC-derived MSCGFP and 40% cherry-labeled MDA-MB-231 (MDA-MB-231cherry) breast cancer cells were incubated in 10 mL MSC medium for approximately 11 days in cell culture plates (diameter 10 cm; Greiner BioOne GmbH, Frickenhausen, Germany) at an initial density of 500 cells/cm2 (equivalent to 300 MSCGFP/cm2 together with 200 MDA-MB-231cherry/cm2). In order to distinguish between the MSC and the breast cancer cells within the co-culture, this time, the MSC were transduced with a third-generation lentiviral SIN vector containing the eGFP gene displaying a green fluorescence; whereas MDA-MB-231 cells were transduced with a similar vector containing the m-cherry gene with a red fluorescence. Appropriate mono-cultures of MSCGFP (300 cells/cm2) and MDA-MB-231cherry (200 cells/cm2) were separately cultivated in parallel and served as corresponding controls.
In separate sets of co-culture, 10 μM of the gap junction inhibitor carbenoxolone (CBX; Sigma, St. Louis, MO) was added every 24 h without medium change. Further co-culture experiments were performed by the addition of 1 μM of the notch signaling inhibitor N-[N-(3,5-difluorophenacetyl-
For fluorescence determination, the medium of the cell cultures was removed at the indicated time points, and the cells of the co-culture and the appropriate two mono-cultures were lysed with SDS, respectively. Afterward, the fluorescence intensities for GFP that corresponded to the appropriate cell number of MSC were measured in aliquots of the cell homogenates at excitation 485 nm/emission 520 nm using the Fluoroscan Ascent Fl (Thermo Fisher Scientific). Likewise, the cherry-based fluorescence of the human breast cancer cells was quantified at excitation 584 nm/emission 612 nm in the Fluoroscan Ascent Fl. The appropriate mono-cultures of MSCGFP and MDA-MB-231cherry demonstrated no artificial cross-fluorescence (data not shown).
In an additional independent evaluation of the cell numbers, the different cell cultures were trypsinized at the appropriate time points, and the cells were counted after trypan blue staining in a fluorescence microscope (Olympus IX50) using the green and red fluorescence filters, respectively, as well as an FITC/TRIC fluorescence dual band filter.
Phenotypic analysis of MSC and human breast cancer cells by flow cytometry
Characterization of the MSC and the MDA-MB-231 cell immunophenotype was performed as previously described [47]. MSC were harvested by use of accutase for 3 min at 37°C, and MDA-MB-231 cells were collected after a 5-min trypsin treatment at 37°C, respectively. Thereafter, the cells were washed twice in ice-cold PBS supplemented with 2% FCS (Biochrom GmbH) and resuspended for flow cytometry testing after staining with a PE-labeled monoclonal mouse anti-human CD73 antibody (clone AD2) (BD Bioscience GmbH), a PE-labeled monoclonal CD90 antibody (clone 5E10) (BioLegend via Biozol GmbH, Eching, Germany), and a PE-labeled monoclonal mouse anti-human CD105 antibody (clone 43A3) (BioLegend via Biozol GmbH), respectively, for 30 min at 4°C in the dark. Control staining was performed with a PE-labeled mouse IgG1 antibody (Dako Cytomation, Hamburg, Germany). The testing of CD90 expression during the co-culture experiments was performed after fixation of MSCGFP and MDA-MB-231cherry cells in 70% (v/v) ice-cold ethanol before CD90 antibody labeling. After two washing steps of the antibody-labeled cells, 400 μL of PBS supplemented with 2% FCS were added along with 1 mL DNA staining solution from the CyStain DNA 2 step kit (Partec GmbH, Münster, Germany) and filtered through a 50-μm filter. Thereafter, the different populations were analyzed in a Galaxy flow cytometer (Partec GmbH). Cells (10,000 events per measurement) were gated in a dot plot of forward versus side scatter signals acquired on a linear scale, whereas fluorescence events were acquired on a logarithmic scale. Positive staining was obtained according to control measurements of the different populations with isotype-matching IgG control antibodies. Histograms were generated using the Flomax software (Partec GmbH).
Cell cycle analysis
The cell cycle analysis in the breast cancer cells and MSC was performed as previously described [10]. Briefly, 5×105 cells were fixed in 70% (v/v) ice-cold ethanol at 4°C for 24 h. Thereafter, the fixed cells were stained with CyStain DNA 2 step kit (Partec GmbH) and filtered through a 50-μm filter. The samples were then analyzed in a Galaxy flow cytometer (Partec GmbH) using the MultiCycle cell cycle software (Phoenix Flow Systems, Inc., San Diego, CA).
Senescence-associated β-galactosidase assay
Cell senescence can be detected by different expression levels of senescence-associated β-galactosidase (SA-β-gal) [48]. The amount of senescent cells was determined by the use of the SA-β-gal staining kit (Cell Signaling Technology, Danvers, MA) and 4′,6-diamidin-2′-phenylindoldihydrochlorid (DAPI) (Roche Diagnostics GmbH, Mannheim, Germany) fluorescence counterstain in accordance to the manufacturer's instructions. For this purpose, 4,000 cells/cm2 were cultivated for 48 h in the corresponding media before fixation and SA-β-gal staining. After completion of the staining procedure, four representative images were taken from diverse areas of each cell culture using phase-contrast microscopy, fluorescence microscopy, and CellBImaging Software (Olympus GmbH, Hamburg, Germany). For the calculation of the percentage of senescent cells, the total number of cell nuclei and number of cell nuclei surrounded by cyan dye were enumerated.
In vivo experiments
Animal research using NOD/scid mice was carried out by following internationally recognized guidelines on animal welfare and has been approved by the institutional licensing committee ref. #33.9-42502-04-06/1178 on 22th September, 2010.
About 1×106 GFP-labeled MDA-MB-231 cells and in a separate group, a co-culture of 1×106 GFP-labeled MDA-MB-231 cells along with 1.5×106 MSC were injected subcutaneously into five animals of 5–6 week-old female NOD/scid mice, respectively. A similar injection of 1.5×106 MSC into three mice, respectively, served as a control. After about 3 weeks postinjection, all 10 MDA-MB-231GFP cells-containing mice had developed subcutaneous tumors, and the animals were sacrificed by CO2 anesthesia and cervical dislocation. After UV light examination for the detection of GFP-positive tissue, the tumors were dissected, weighted, washed in PBS, and subsequently fixed in a 4% glutardialdehyde solution. After dehydration (Shandon Hypercenter XP), the tissue was embedded in paraffin. Sections (4 μm) were deparaffinized with xylene and stained with hematoxylin/eosin for histopathological evaluations.
Results
The interaction of human MDA-MB-231 breast cancer cells with human MSC was investigated by measurement of the proliferative capacity of each cell type and cell surface marker expression. To distinguish between the two cell types in several co-cultures in vitro and in vivo, the breast cancer cells were transduced with a lentiviral expression vector containing either the eGFP gene (MDA-MB-231GFP) or the m-cherry gene (MDA-MB-231cherry); whereas primary MSC were used either unlabeled or labeled with the eGFP gene (MSCGFP).
Since MSC did not grow in MDA-MB-231 medium (data not shown), proliferative effects of a co-culture were tested in MSC medium. Thus, a constant amount of 104 MDA-MB-231GFP cells was added to MSC cultures of different cell densities, respectively, to achieve cell ratios between 10% MSC/90% MDA-MB-231GFP up to 60% MSC/40% MDA-MB-231GFP (Fig. 1A). Although the breast cancer cells alone proliferated about twice as much in MSC medium as compared with the standard breast cancer culture medium, this proliferative capacity was even further elevated in the presence of MSC. The growth rate of MDA-MB-231GFP cells was quantified in a fluorescence assay by the directly proportional GFP fluorescence intensity, which was significantly enhanced by 39.5%±9.2% (n=3) after 4 days in a co-culture mixture of 60% MSC/40% MDA-MB-231 (Fig. 1A).
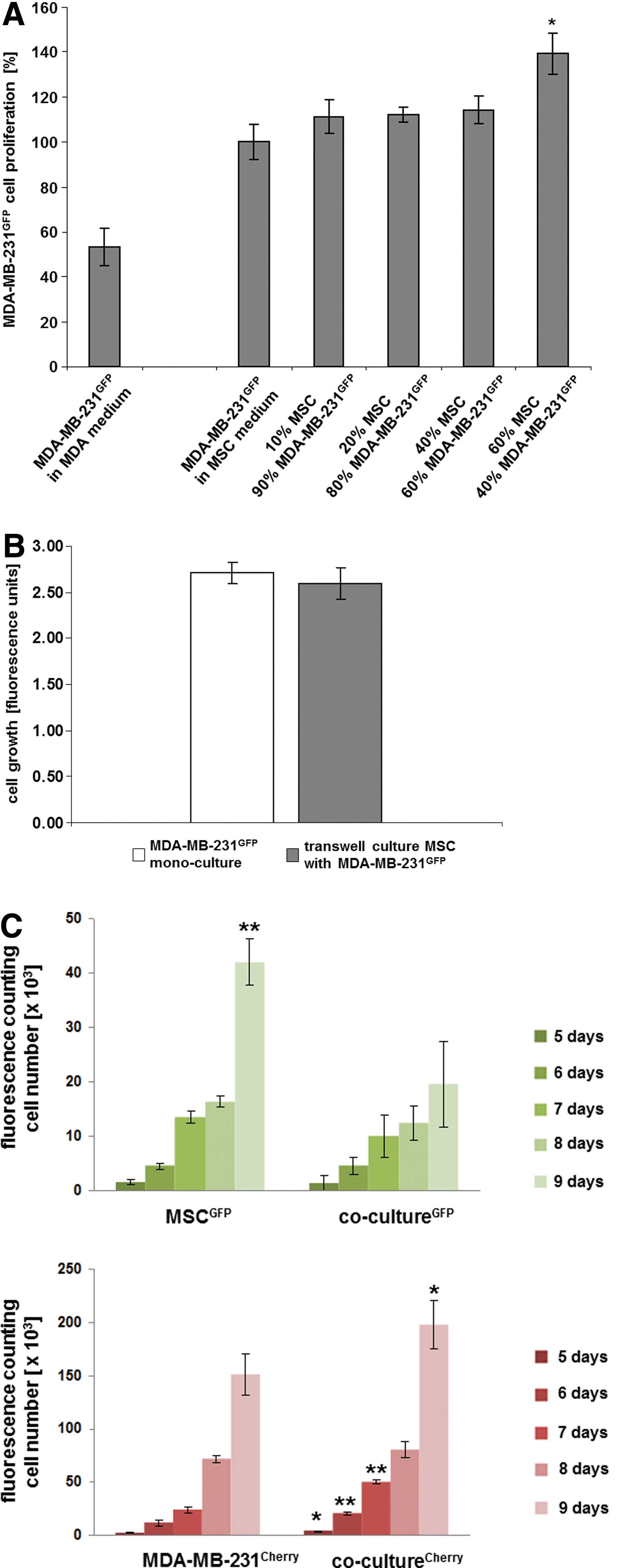
A possible requirement for a certain vicinity of the breast cancer cells to MSC to transmit growth-stimulatory signals was tested using a transwell system, by which MDA-MB-231GFP cells were cultured in a plate and primary MSC in the transwell, allowing the exchange of medium through 0.4 μm pores. After 4 days of culture, fluorescence measurements that quantitatively represent the amount of GFP-positive breast cancer cells revealed no significant differences between the fluorescence intensity of MDA-MB-231GFP cells cultured alone and in a transwell co-culture with MSC (Fig. 1B). These findings suggested that soluble factors from either cell population released into the culture medium under these conditions had little, if any, growth-stimulatory effects on the breast cancer cells.
Based on these results, a co-culture of 60% MSCGFP/40% MDA-MB-231cherry was performed for approximately 9 days, and the proliferation of the co-culture was compared with the appropriate mono-cultures. Therefore, MDA-MB-231cherry (200 cells/cm2) were seeded in a mono-culture, MSCGFP (300 cells/cm2) were seeded in a separate mono-culture, and MSCGFP (300 cells/cm2)+MDA-MB-231cherry (200 cells/cm2) (=500 cells/cm2) were seeded in the co-culture representing a cell ratio of 60%:40% (MSC:MDA-MB-231). Quantification of the appropriate cell numbers was performed by counting the fluorescing cells in a hemocytometer using a fluorescence microscope. Although the MSCGFP mono-culture demonstrated a significantly enhanced cell number as compared with the MSCGFP in the co-culture with MDA-MB-231cherry after 9 days (Fig. 1C, upper panel, n=4), opposite results were obtained for the breast cancer cells. The amount of MDA-MB-231cherry cells in co-culture with MSCGFP was significantly elevated over the MDA-MB-231cherry cell number of the corresponding mono-culture already within 5 days, respectively (Fig. 1C, lower panel, n=4).
These findings were substantiated in an independent evaluation. Quantification of GFP in a fluorescence-based microtiter plate assay revealed an enhanced growth of the MSCGFP mono-culture over the MSCGFP/MDA-MB-231cherry co-culture, which was significant after 8 and 9 days (Supplementary Fig. 1D, upper panel, n=7; Supplementary Data are available online at
Supportive results of a reduced MSC growth rate were obtained in a further control experiment by which conditioned media from MDA-MB-231 cells after 2, 4, and 6 days were incubated with MSC, respectively, and revealed a progressively reduced proliferation rate within 72 h (Supplementary Fig. S1B), which was also substantiated by continuously increased G0/G1 cell cycle arrest in MSC (Supplementary Fig. S1C) although little, if any, accumulation of SA-β-gal as a marker of senescence could be detected in MSC during co-culture (Supplementary Fig. S1D).
The progressive accumulation of a significantly increasing breast cancer cell number in contrast to MSC was also observed during morphological studies (Fig. 2). Following the randomly distributed cell types at the beginning of the co-culture that still prevailed after 3 days (Fig. 2, upper panel), there was a continuous formation of growing colonies of MDA-MB-231cherry cells after 5–9 days of co-culture, by which the MSCGFP surrounded these breast cancer cell colonies by distinct morphological changes with a long extension of some cell bodies (Fig. 2, indicated by white arrows). After 9 days of co-culture, the initial majority of MSCGFP was already overgrown by the MDA-MB-231cherry. Moreover, co-culture of MSCGFP and with MDA-MB-231cherry was also associated with the formation of certain hybrid/chimeric cells already after 3–5 days, demonstrating a yellow fluorescence in the dual fluorescence picture and a corresponding expression of both mcherry and eGFP fluorescence (Fig. 2, indicated by orange arrows).
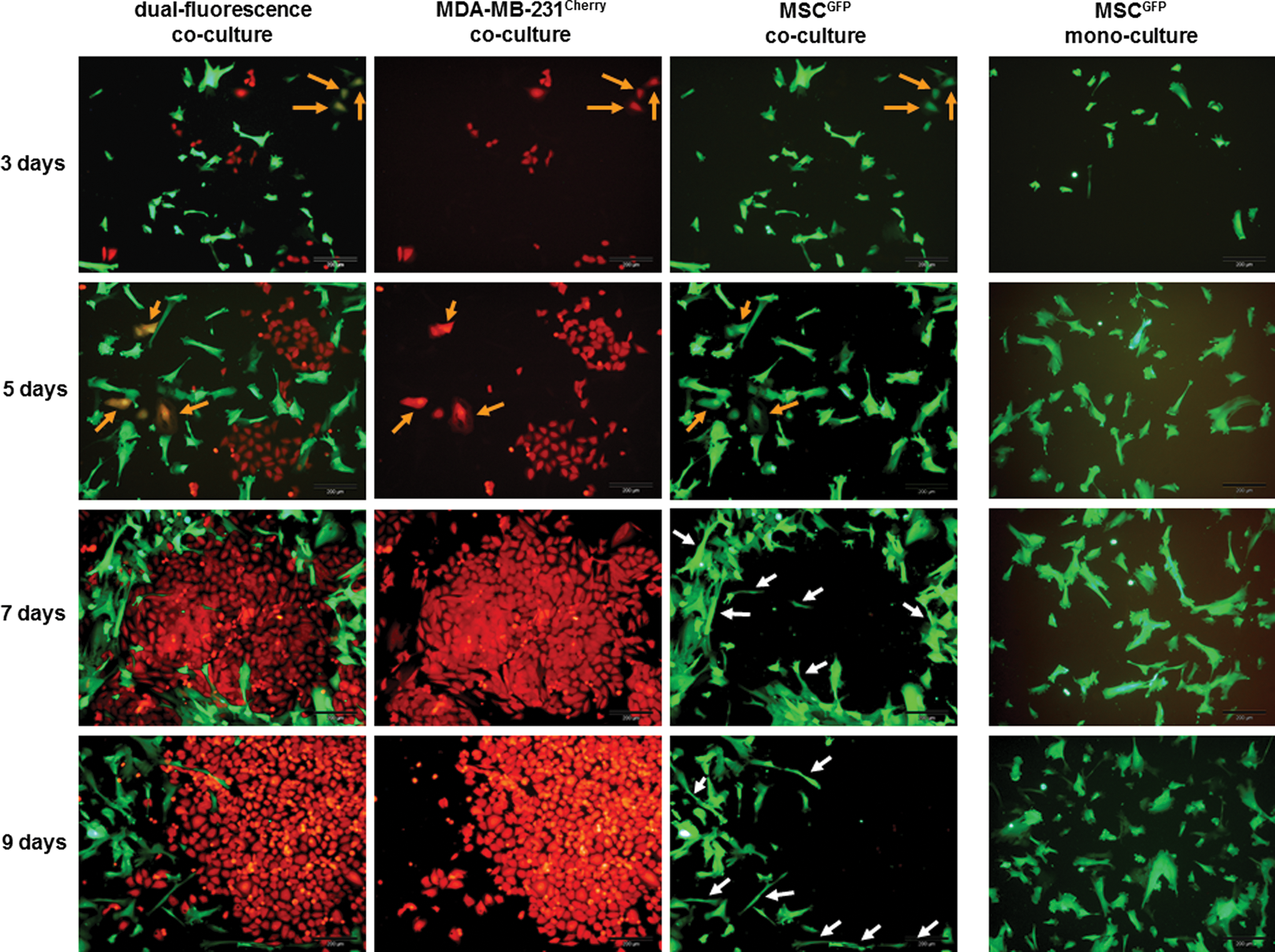
The co-culture of MSCGFP/MDA-MB-231cherry cells (ratio 60:40) was examined by fluorescence microscopy with the appropriate monochrome fluorescence filters and a FITC/TRIC dual band fluorescence filter to distinguish between the cells of the two populations. Following the initially randomly distributed cell types after 3 days, distinct and progressively increasing islands were formed by MDA-MB-231cherry cells in the course of co-culture, which were surrounded by a ring of MSC by enlarging their cell bodies. The altered morphology of some MSCGFP with elongated cell body is indicated by white arrows in the 7- and 9-days MSCGFP co-culture as compared with the appropriately cultured MSCGFP in a mono-culture. Moreover, the MSCGFP/MDA-MB-231cherry co-culture formed some hybrid/chimeric cells already within 3–5 days displaying both red and green fluorescence in the appropriate mono-cultures and a yellow fluorescence in the dual-fluorescence co-culture, which are indicated by orange arrows. Bars represent 200 μm. Color images available online at
Characterization of the two different cell types was also performed by flow cytometric analysis of typical MSC surface markers, including CD73, CD90, and CD105 according to the recommendation by the International Society for Cellular Therapy [15]. Although these three surface markers were detectable in MSC to nearly 100% (Fig 3A, lower panel), the MDA-MB-231 cells expressed the CD73 and the CD105 markers to nearly 100%; whereas little, if any, CD90 was detectable in these breast cancer cells (Fig 3A, upper panel). Accordingly, CD90 expression along with the eGFP or mcherry fluorescence signal was used to monitor possible cellular changes during the co-culture. Moreover, according to different levels of autofluorescence of the two cell types, a simultaneous analysis of the appropriate cell cycles was achieved (Fig. 3B). The two overlapping cell cycles of the MSCGFP and MDA-MB-231cherry cells were assigned to the corresponding GFP and mcherry fluorescence, respectively, and quantification of the appropriate G0/G1 peak for the MSC and the MDA-MB-231 population was performed by cell cycle analysis software (Fig. 3B, upper panel), which substantiated the data obtained by fluorescence-based microtiter plate assay, fluorescence cell counting, and the morphological results. Concomitant with a progressive decline of MSCGFP in the co-culture ratio from initially 60% to about 28% after 9 days, the ratio of MDA-MB-231cherry cells continuously increased from initially 40% to about 72% after 9 days (Fig. 3B, upper panel). Little, if any, change was observed in the cell cycle distribution of the corresponding mono-cultures of MSCGFP and MDA-MB-231cherry cells within the 9-days (Fig. 3B, middle panels). CD90 expression was also quantified by flow cytometry during the co-culture experiment and revealed a significant increase in the MDA-MB-231cherry population after 5 days, which sustained until 9 days (Fig. 3B, lower panel). This induction of CD90 in the breast cancer cells was surprising, and similar results could be reproduced in appropriate co-cultures of MDA-MB-231 cells with all three different primary MSC populations used in this study, respectively.


In order to investigate the role of MSC and possible signal transduction pathways during the CD90 induction in the MDA-MB-231 cells, the MSCGFP/MDA-MB-231cherry co-culture was tested by replacing the culture medium each day. Moreover, MDA-MB-231 cells from a transwell co-culture experiment were also tested for CD90 expression for up to 8 days. Although interactions of MSC with neighboring cells may involve GJIC, 10 μM of the gap junction inhibitor CBX was added each day to a further co-culture experiment. Furthermore, cellular interactions of MSC can be mediated via the notch signaling pathway, and, therefore, 1 μM of the notch signaling inhibitor DABT was added each day to a MSCGFP/MDA-MB-231cherry co-culture. MSC expressed always nearly 100% of CD90 in the different co-culture experiments, by which a medium exchange every 24 h, a transwell culture, or the inhibitors DABT or CBX had little, if any, effect on this constitutive CD90 expression (Fig. 3C). Conversely, MDA-MB-231 cells with no detectable CD90 expression increased this level to about 33% after 4 days of co-culture; whereas DABT- and CBX-treated co-cultures demonstrated a CD90 expression in MDA-MB-231 cells of about 28% and 25%, respectively. In contrast, there was no detectable CD90 expression of MDA-MB-231 cells in a transwell co-culture at any time of the experiment (Fig. 3C). The levels of CD90 expression in MDA-MB-231 cells increased in a co-culture with or without medium exchange to about 73% after 6 days and reached more than 98% after 8 days (Fig. 3C). However, in the presence of the notch signaling inhibitor DAPT or the gap junction inhibitor CBX, the CD90 induction in MDA-MB-231 cells increased only to about 54% and 35% after 6 days, respectively, and reached 62% and 67% after 8 days of co-culture, respectively (Fig. 3C). Furthermore, DABT treatment revealed a reduced growth of the breast cancer cells by 9.9%±0.2% after 7 days and 7.5%±0.1% (n=3) after 8 days of co-culture, respectively. Likewise, breast cancer cell growth was also significantly diminished by 8.5%±0.1% after 7 days and by 8.6%±0.1% (n=3) after 8 days of co-culture after CBX treatment as quantified by cell cycle analysis.
Together, these findings suggested the requirement of direct cell-to-cell interactions between MSC and MDA-MB-231 cells to relay signals for induced cell growth and CD90 expression in the breast cancer cells. Moreover, this growth induction and CD90 expression was conferred, in part, by signaling pathways involving GAP junctions and the notch pathway.
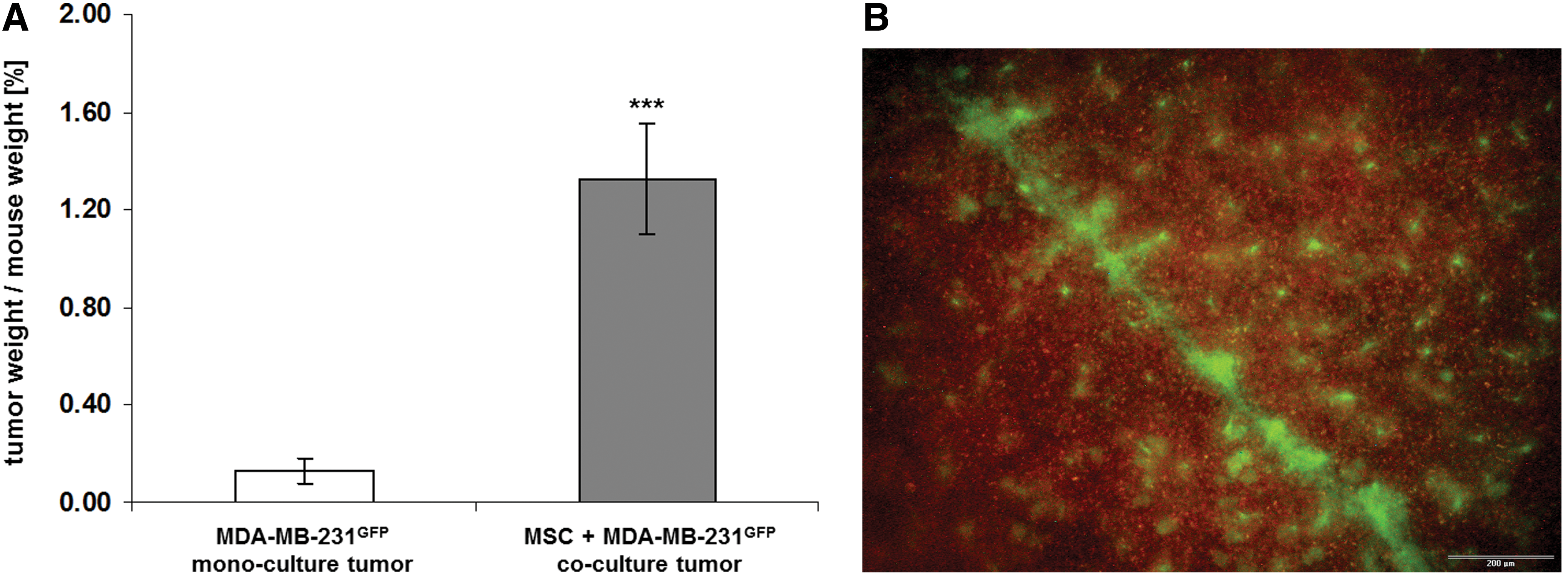
A possible MSC-mediated growth support and a concomitant CD90 induction of the breast cancer cells was also tested in an in vivo approach. A subcutaneous injection of either GFP-labeled MDA-MB-231 breast cancer cells alone or a co-culture of MSC with MDA-MB-231GFP breast cancer cells (co-culture ratio 60:40) into 5 NOD/scid mice, respectively, revealed a tumor development in all 10 animals. As a control, MSC alone were injected into three NOD/scid mice with no detectable tumor development or any significant physiological changes within 6 months. In contrast, a progressively increasing tumor development in the MDA-MB-231GFP-injected mice could be observed already after 23 days, and the comparison of the tumor sizes and weights after tumor dissection revealed a tumor weight/mouse weight relation of 0.13%±0.05% (n=5) for the MDA-MB-231GFP mono-culture (Fig. 4A). At the same time, an ∼10-fold increased tumor size was detected for the MSC/MDA-MB-231GFP co-culture reaching a tumor weight/mouse weight relation of 1.32%±0.22% (n=5). Moreover, fluorescence scanning of the mouse organs revealed several spleen metastases of green fluorescing cells derived from the MSC/MDA-MB-231GFP co-culture (Fig. 4B). In contrast, no metastases were detectable in the mono-culture MDA-MB-231GFP mouse tumors after 23 days. These findings suggested that co-culture of the breast cancer cells with MSC significantly elevated the tumor growth and simultaneously enhanced the metastatic capacity.
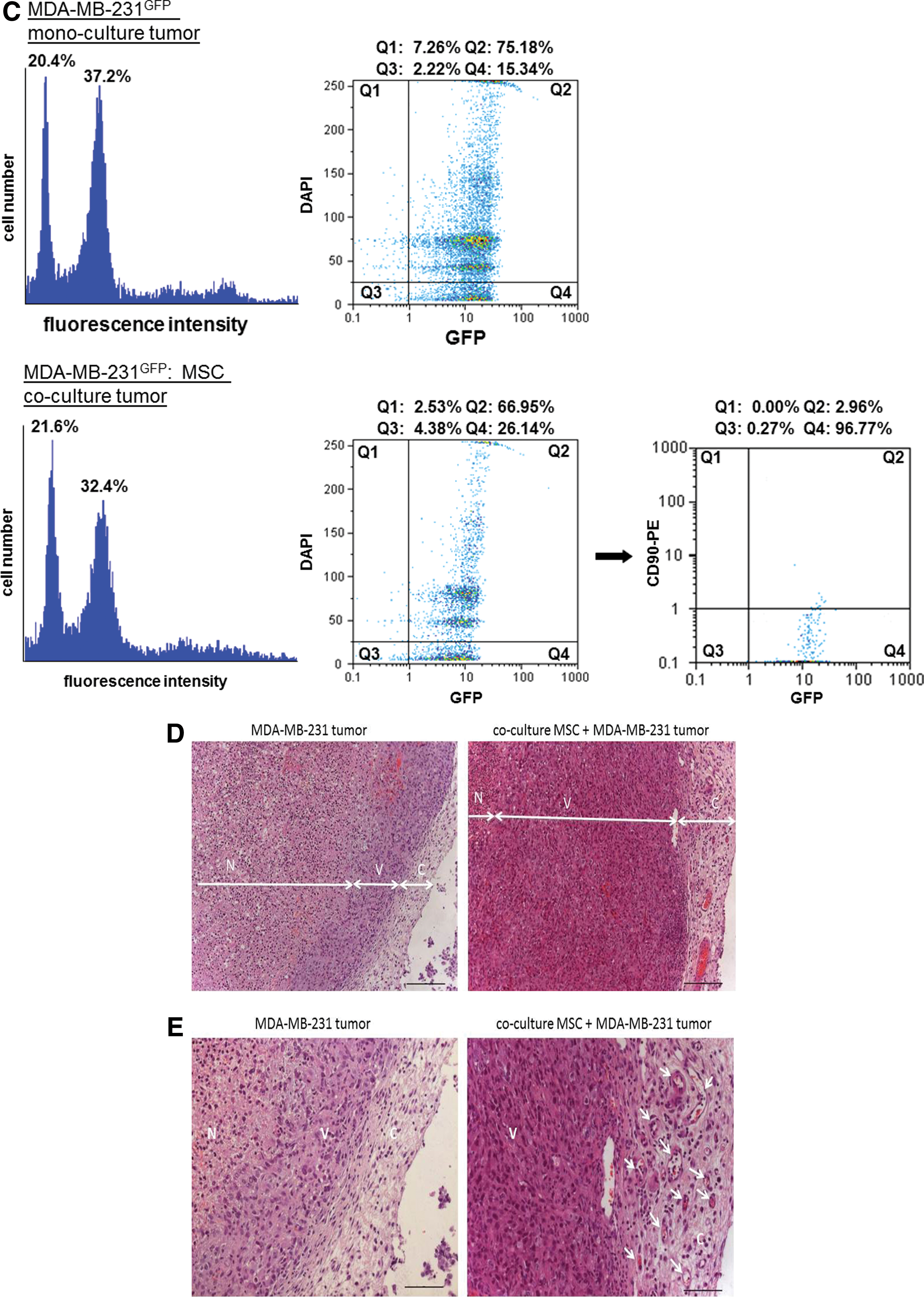
Flow cytometric analysis was performed following a mechanical preparation of a representative single cell population of the tumors from MDA-MB-231GFP as compared with the MSC/MDA-MB-231GFP co-culture-derived tumors (Fig. 4C). Thus, cell cycle analysis that is proportional to the amount of DNA revealed two prominent peaks in each tumor-derived population, by which more than 90% of each cell population included by these peaks expressed the GFP protein (MDA-MB-231GFP: 73.97%+16.10%=90.07%; MDA-MB-231GFP/MSC co-culture: 63.84%+28.60%=92.44%) [Fig. 4C, left cell cycle histograms and middle dot blot (DAPI vs. GFP) histograms]. Interestingly, flow cytometric analysis of the co-culture-derived tumor with anti-CD90 as one of the MSC markers revealed about 3% of CD90+ cells that simultaneously express GFP [Fig. 4C, right dot blot (CD90 vs. GFP, Q2) histogram], suggesting alterations of the originally CD90− breast cancer cells also in vivo. Together, these data demonstrated that the GFP-expressing tumor cells acquired certain characteristic markers of MSC during co-culture in vivo.
A histological examination of the tumors in 4 μm tissue sections was performed in four slides of each tumor representing four different adjacent tumor areas, respectively. The data revealed an enlarged region (64.1%±1.3%) of necrotic cells (N) in the MDA-MB-231GFP tumor as compared with the co-culture tumor (54.9%±0.7%) (n=4) (Fig. 4D). Conversely, the MDA-MB-231GFP/MSC co-culture tumor tissue represented a markedly larger region of viable cells (V) (45.6%±0.7%) in contrast to the breast cancer mono-culture tumor (35.9%±1.3%) (n=4) that demonstrated a tumor viability index (area ratio of viable tissue to necrotic tissue) of 0.8 for the MSC co-culture tumor and only 0.5 for the MDA-MB-231GFP mono-culture tumor. Although the width of the tumor capsule (C) displayed 278±78 μm in the MSC/MDA-MB-231GFP co-culture and 145±20 μm in the MDA-MB-231GFP mono-culture tumor, a large amount of blood vessels became detectable within the tumor capsule derived from the MSC/MDA-MB-231GFP co-culture, indicating a significantly elevated neovascularization in contrast to the MDA-MB-231GFP mono-culture tumor (Fig. 4E).
Discussion
A variety of different studies have investigated the interactions of MSC with breast cancer cells, and the data arising from these studies are controversial. Previous work revealed that MSC from bone marrow in 3D cultures promote the growth of breast cancer cells [49]. In contrast, MSC extracts derived from either human MSC lysates or MSC supernatants exhibited growth inhibitory properties in different cancer cell lines, including breast cancer [27]. Results obtained in the present study demonstrated that MSC supernatants or conditioned media in a transwell exchange system had little, if any, effect on the proliferation of breast cancer cells; however, a vice versa approach revealed that MDA-MB-231 conditioned media slowed MSC growth, which was associated with a G0/G1 cell cycle accumulation. Similar effects have been suggested in vivo, by which enhanced cytokine accumulation mediated by the MDA-MB-231 breast cancer cells in the bone marrow of athymic nude mice, including MCP-1, vascular endothelial growth factor, and eotaxins, can affect the MSC microenvironment and thus, MSC growth properties [50]. Conversely, a direct co-culture of MSC with breast cancer cells was accompanied by enhanced growth of tumor cells in vitro and in vivo and at least a transient induction of CD90 expression in the breast cancer cells. These findings may not represent a contradiction, as it appears conceivable that several mechanisms are mutually combined rather than representing controversial phenomena. Thus, direct cell-to-cell interactions between MSC and the breast cancer cells contribute toward minimizing the intercellular space for maximal interactions to exchange cytokines and microRNA-containing microvesicles within this cellular microenvironment and thereby, trigger and enhance signaling cascades that are required for tumor cell proliferation and migration [51]. Cellular interactions of MSC with breast cancer cells are also facilitated by gap junctions and subsequent GJIC [44,51 –53]. Although CBX blocks the assembly of membrane-spanning channels (connexons) on adjacent cells to connect to a functional gap junction [54], the results in the present study substantiate the important role of GJIC in MSC, demonstrating that inhibition of gap junctions by CBX could also partially block the MSC-mediated growth induction and CD90 expression in breast cancer cells.
Further cellular interactions of MSC with breast cancer cells are associated with the production and release of a large panel of chemokines, cytokines, and growth factors by which factors, including IL1, TNF, and prostaglandins, are discussed as contributing to CD90 induction [55]. Vice versa, breast cancer cells respond to these compounds by the expression of distinct receptors such as CCR5 and CXCR4, which promote signals for proliferation and migration of the tumor cells on further corresponding stimuli such as CCL5 and SDF-1 [23,56 –58]. These effects also involve notch signaling, which plays an important role in MSC for supporting the repair of damaged tissues and modulating distinct immune cell functions [59]. Activation of notch signaling involves cleavage of the intracellular notch receptor domain as a downstream signal transducer by a presenilin-γ-secretase complex, which can be blocked by DABT [60] and, therefore, prevents further notch responses [61]. Consequently, inhibition of notch signaling by DABT in MSC co-culture was accompanied by a partial inhibition of growth and CD90 induction in breast cancer cells, indicating a functional involvement of the notch pathway in MSC-MDA-MB-231 cell interactions. The importance of notch signaling has also been demonstrated in developing T cells for the maintenance of CD90 expression [62].
MSC-mediated CD90 induction in the MDA-MB-231 cells was observed in vitro during direct co-culture within 6–8 days; however, the in vivo analysis was performed in a multi-population microenvironment after 23 days when MSC were largely overgrown by the tumor cells, which limits the effects of MSC and may, therefore, explain differences in the amount of CD90-expressing breast cancer cells in vitro and in vivo. Further interference of direct MSC effects on the breast cancer cells in vivo is also related to cellular interactions of the breast cancer cells with other cell types, including carcinoma-associated fibroblasts that contribute to the organization of the tumor stroma, tumor-associated macrophages and lymphocytes for the immune response, and endothelial cells for the tumor neovascularization. Although expression of the heavily N-glycosylated GPI-anchored CD90 surface molecule in MSC represents one of the typical MSC markers besides CD73 and CD105, it is interesting to note that the MDA-MB-231 breast cancer cells already constitutively express CD73 and CD105, which is also confirmed by other work [43,63]. Therefore, the MSC-mediated CD90 expression in co-culture with MDA-MB-231 cells induces a further stem cell marker that attributes more MSC-like characteristics and similarities to these breast cancer cells and raises more difficulties to distinguish between the populations and properties of MSC and MDA-MB-231 cells. Indeed, CD90 expression in tumor-associated cells has been identified as a possible marker for cancer stem cells in high-grade gliomas and other solid tumors [64].
During the cellular interactions with the breast cancer cells, some MSC changed their phenotype with cytoplasmic extensions and an elongated cell body to build a cellular rim of MSC surrounding the continuously proliferating and extending breast cancer cell colonies. While these processes of MSC appear to function as a border for the breast cancer cells, such MSC structures support to limit further expansion of the tumor cells. However, opposite effects of this kind of interaction can also promote altered and stabilized extracellular matrix structures of a tumor microenvironment that provides a certain protection for the breast cancer cells, for example, against chemotherapeutic treatment and, therefore, facilitates tumor cell expansion.
Further cellular interactions between MSC and the breast cancer cells were observed by the formation of hybrid/chimeric cells. These newly formed MDA-MB-231-MSC hybrid/chimeric cells express genomic properties from both the MDA-MB-231cherry breast cancer cells and the MSCGFP that are represented by new cell populations displaying dual fluorescence. MSC cell fusion with tumor cells has been previously described [65], and other studies have extensively characterized hybrid cell populations derived from human bone marrow cells and MDA-MB-231 or MA11 breast cancer cells with increased metastatic capacity while exhibiting a mixed breast cancer-mesenchymal expression profile [66]. Together with the present results, these findings furthermore suggest that cell fusion with tumor cells or partial exchange of the genome applies to MSC from different human tissue sources and may, thus, represent a more general property of MSC. Moreover, these genomic exchange processes between MSC and tumor cells as a further mechanism of MSC/tumor cell interaction can also occur in vivo with the formation of a variety of different hybrid/chimeric cell populations, which would be interesting to further explore with regard to their growth properties and altered cellular functionalities.
According to the biodiversity of MSC that are present in nearly all kinds of human tissues and exhibit regenerative potential, maintenance of stemness, migration and transport function, support of repair and neovascularization of tissues, immune modulation, differentiation capacity [13,14,23], and even cell fusion properties, these all-round capabilities of MSC in normal tissues are also available in concert with tumor cells. Thus, the cell-to-cell interactions in combination with the exchange of an appropriate cytokine/chemokine cocktail synergistically enhanced the breast cancer cell growth in vitro and in vivo. Moreover, these interactions also markedly increased the viability of intratumoral cancer cells in vivo and contributed to a significantly elevated vascularization of the tumor tissue and the formation of metastases in distant organs.
Conclusion
MSC performed multiple cell-to-cell interactions with breast cancer cells. These cellular interactions were accompanied by an increased island-like breast cancer growth during direct in vitro co-culture and in vivo xenografts. Moreover, MSC established a distinct microenvironment during co-culture with the breast cancer cells to facilitate cellular alterations by which new populations of hybrid/chimeric cells were generated. Furthermore, MSC induced at least transiently CD90 expression in the breast cancer cells, which partially involved notch signaling and GJIC. Based on these direct cellular interactions, MSC may be considered a potential vehicle in future studies to target tumor cells by including GJIC and notch signaling pathways.
Footnotes
Acknowledgment
This work was supported by a grant from the Niedersächsische Krebsgesellschaft to Ralf Hass.
Author Disclosure Statement
The authors disclose any commercial associations that might create a conflict of interest in connection with this article. The authors declare that competing financial interests of all authors have been appropriately disclosed according to the policy of the Journal. Moreover, the authors declare that there are no conflicts of interest, be they actual or potential.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.