
Abstract
Curculigoside, a phenolic glycoside, is the main active compound of Curculigo orchioides (Amaryllidaceae, rhizome). C. orchioides is a traditional Chinese herbal medicine and has been commonly used to treat orthopedic disorders and bone healing in Asia. This study evaluated the effect of curculigoside on osteogenic differentiation of human amniotic fluid-derived stem cells (hAFSCs). The results showed that curculigoside stimulated alkaline phosphatase activity and calcium deposition of hAFSCs during osteogenic differentiation in a dose-dependent manner (1–100 μg/mL), while the effects were reduced at the higher concentration of 200 μg/mL. From reverse transcriptase–polymerase chain reaction analysis, the osteogenic genes osteopontin (OPN) and Collagen I were upregulated with curculigoside treatment (1–100 μg/mL). Concurrently, the ratio of osteoprotegerin (OPG) to receptor activator of nuclear factor kappa-B ligand (RANKL) was increased, indicating the inhibition of osteoclastogenesis by curculigoside. Moreover, the role of Wnt/β-catenin signaling during curculigoside treatment was revealed by the upregulation of β-catenin and Cyclin D1. In summary, curculigoside improved osteogenesis and inhibited osteoclastogenesis of hAFSCs, suggesting its potential use to regulate hAFSC osteogenic differentiation for treating bone disorders.
Introduction
O
Amniotic fluid-derived stem cells (AFSCs) are reported to have strong potential to differentiate into osteoblasts and arising as a novel cell source for treating bone diseases [9,10]. AFSCs possess a phenotype of mesenchymal stem cells (MSCs), which have the ability to migrate and engraft into multiple musculoskeletal tissues, especially sites of injury, and undergo site-specific osteogenic differentiation [11]. Amniocentesis is a widely accepted procedure in prenatal testing and presents a low risk for both the mother and the fetus, enabling the easy derivation of AFSCs [12]. Compared to MSCs derived from bone marrow, AFSCs have fetal tissue origin and an extensive self-renewal capacity [13]. Compared to pluripotent embryonic stem cells, AFSCs have no ethical concerns and no tumorigenic risk for their usage in clinics [14]. Therefore, AFSCs have become ideal candidates for cell-based therapies to improve bone formation in patients who suffer from diverse metabolic and genetic bone diseases, including osteoporosis.
Curculigo orchioides (Amaryllidaceae, rhizome) is a traditional Chinese herbal medicine, which has been commonly used to treat orthopedic disorders and bone healing for thousands of years [15,16]. Modern pharmacological study indicates that curculigoside, a phenolic glycoside (Fig. 1), is the main active compound of C. orchioides. Curculigoside has been found to enhance the proliferation of mouse preosteoblastic cells and stimulate the secretion of vascular endothelial growth factor and bone morphogenetic protein (BMP)-2 [17,18]. It is also reported that curculigoside prevented oxidative damage and inhibited osteoclastogenesis of rat bone marrow cells [16,19]. However, to date, the effects of curculigoside on MSC osteogenic differentiation have not been well studied. There is only one report that investigated the use of curculigoside to promote osteogenic differentiation of rat bone marrow stromal cells [20]. In addition, no detailed signal transduction pathways have been elucidated for curculigoside-treated stem cells. Given the advantages of human amniotic fluid-derived stem cells (hAFSCs), the present study focused on the effects of curculigoside on osteogenic differentiation of hAFSCs and related signaling pathways. To our knowledge, this is the first study to use curculigoside to enhance hAFSC lineage commitment.
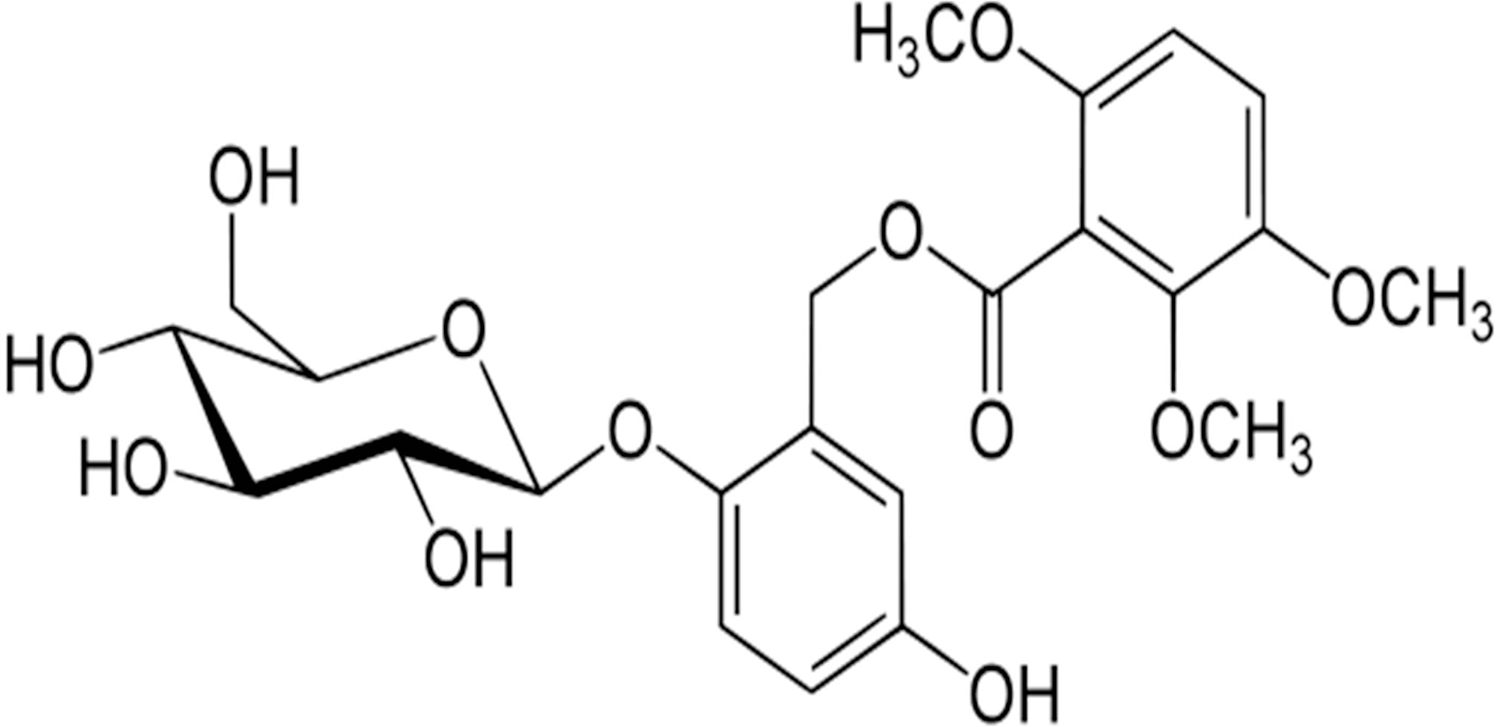
Chemical structure of curculigoside.
Materials and Methods
Culture of hAFSCs
The hAFSCs were isolated and cultured as previously described [9]. hAFSCs at passages 16–18 were used in this study. All culture regents were from Life Technologies unless otherwise noted. The cells were maintained in an alpha-minimum essential medium (α-MEM) supplemented with 15% embryonic stem cell qualified fetal bovine serum (ES-FBS), 100 U/mL penicillin, 100 μg/mL streptomycin, 2 mM
hAFSC treatment with curculigoside
Curculigoside (≥99.6% purity) was purchased from the Chinese National Institute for the Control of Pharmaceutical and Biological Products. The cells (1×104 cells/mL) were seeded in 48-, 24-, and 6-well plates and cultured in the growth medium until 70%–80% confluence. Then, the cells were treated with the differentiation medium containing α-MEM, 17% FBS (Atlanta Biologicals), 2 mM
Cell proliferation analysis
hAFSCs (1×104/well) were seeded in a 48-well plate. After 24 h of incubation, the growth medium was changed to the medium containing 0 (Control), 1, 10, 100, or 200 μg/mL of curculigoside to study the dose effects. Cells were incubated at 37°C in a humidified 5% CO2 incubator for 1, 2, 3, or 4 days. After that, the medium was replaced with 500 μL of 10% Alamar Blue (AbD Serotec) solution at 37°C for 3 h. The fluorescence of the medium was then monitored in triplicate at 535 nm excitation wavelength and 590 nm emission wavelength using a GENios Pro plate reader (Tecan). The fluorescence intensity was correlated to the cell number, using a standard calibration curve.
ALP activity assay
Osteogenesis of hAFSCs was induced by the differentiation medium containing different concentrations of curculigoside (0–200 μg/mL). At day 7, cells were washed with phosphate-buffered saline (PBS) twice and lysed with the lysis buffer consisting 20 mM Tris-HCl (pH 7.5), 150 mM NaCl, and 1% Triton X-100 for 5 min. The chromogenic substrate for ALP was p-nitrophenyl phosphate (pNPP; Sigma-Aldrich). A 50-μL supernatant of lysate was mixed with a 50 μL pNPP (1 mg/mL) substrate solution containing 1.0 mg/mL pNPP, 0.2 M Tris buffer, and 5 mM MgCl2 and incubated at 37°C for 15 min on a Belly Button Shaker (MidSci). The reaction was stopped by the addition of 25 μL of 3 N NaOH. Absorbance of p-nitrophenol released by the samples was measured at 405 nm using a SpectraMAX 250 microplate reader (Molecular Devices). The protein concentration of cell lysates was determined using the Bradford assay at 595 nm on the SpectraMAX 250 microplate reader. The ALP activity was normalized to the total protein content of cell lysates and expressed as nmol (p-nitrophenyl)/min/mg protein.
Alizarin Red S staining
Alizarin Red S (ARS) staining was performed to evaluate the calcium deposition in osteogenic cells obtained from hAFSCs after 28 days of treatment with curculigoside. Briefly, cells cultured in 24-well plates were rinsed with PBS twice, fixed with 10% (v/v) formalin, and then stained with a 1% (w/v) ARS solution. Orange red staining indicated the location and intensity of the calcium deposition. The presence of calcium was observed with a light microscope Olympus IX71 (Olympus Corporation).
Assay of calcium deposition
To quantify mineralization, the calcium deposited by hAFSCs after 14 days was measured using the Calcium Assay (Genzyme Diagnostics). Briefly, samples were added with 1 M acetic acid and placed on a vortex overnight at 4°C to extract the calcium from the mineralized matrix. In a 96-well clear polycarbonate plate, 15 μL of cell extract was mixed with 150 μL of the Calcium Assay reagent and incubated for 30 s at room temperature. The absorbance was determined at 650 nm on a SpectraMAX 250 microplate reader. The samples were measured in triplicate and compared to the calcium calibration curve.
Reverse transcriptase–polymerase chain reaction
Total RNA was isolated using the TRIZOL reagent (Invitrogen) from hAFSCs treated with different concentrations of curculigoside after 21 days. RNA concentrations were measured using a ND-1000 spectrophotometer (NanoDrop Technologies). After that, 1 μg of RNA was initially reverse transcribed into cDNA using the SuperScript™ III First-Strand Synthesis System (Invitrogen). Then, 200 ng of the cDNA was used as a template for the amplification of target genes using the Quick-Load® Taq 2×Master Mix Kit (BioLabs). The primer sequences of the analyzed genes and polymerase chain reaction (PCR) conditions are listed in Table 1. For osteogenic differentiation, expressions of osteopontin (OPN) and Collagen I were measured. For osteoclast differentiation, osteoprotegerin (OPG) and receptor activator of nuclear factor kappa-B ligand (RANKL) were assessed. For the Wnt pathway, β-catenin, cyclin D1, and runt-related transcription factor 2 (RUNX2) were analyzed. The gene of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as an endogenous reference. Amplified products were fractionated in a 2% agarose (Fisher Scientific) gel at 70 V for 80 min and visualized and photographed with a Gel Doc 2000 Gel Documentation System (Bio-Rad). The expression level of each gene was analyzed using ImageJ Software and normalized to the GAPDH expression.
Sequences are depicted in 5′-3′ direction.
Tm is the annealing temperature at which the primer binds to the RNA template during PCR. Touch down: Tm from 62°C to 52°C, decrease 0.5°C per cycle and the following cycles were run at 52°C. All the genes used 35 cycles.
Osteogenic genes: osteopontin (OPN), and Collagen I; Genes in Wnt pathway: β-catenin, Cyclin D1, and runt-related transcription factor 2 (RUNX2). Genes in osteoclast differentiation: osteoprotegerin; (OPG) and receptor activator of nuclear factor kappa-B ligand (RANKL). Housekeeping gene: glyceraldehyde-3-phosphate dehydrogenase (GAPDH).
hAFSCs, human amniotic fluid-derived stem cells; RT-PCR, reverse transcriptase–polymerase chain reaction.
Statistical analysis
Unless otherwise noted, all experiments and samples were triplicated. Experimental results are presented as mean±standard deviation (n=3) and analyzed using ANOVA followed by the paired Tukey–Kramer analysis using JMP 7.0 (SAS Institute, Inc.) with P<0.05 considered as statistically significant.
Results
Effects of curculigoside on the proliferation of hAFSCs
The stimulation effect of curculigoside on the proliferation of hAFSCs during a 4-day culture was evaluated at various concentrations (1–200 μg/mL) (Fig. 2). In the presence of curculigoside at 1 or 10 μg/mL, the growth of hAFSCs from day 1 to day 4 was significantly increased compared to the control (Fig. 2A). Specifically, 1 μg/mL curculigoside increased the proliferation by 13%, 33%, 34%, and 15% on day 1, 2, 3, and 4, respectively, and 10 μg/mL curculigoside increased the proliferation by 21%, 39%, 13%, and 19%, respectively (Fig. 2B). In the 100 μg/mL curculigoside group, the cell growth was similar to the control on day 1, but remarkably increased on day 2 (ie, 47%). Then, on day 3 and day 4, the cell number became close to the control group again. However, 200 μg/mL curculigoside inhibited the proliferation of hAFSCs over the 4-day culture (−7% to −32%), indicating that high concentrations (≥200 μg/mL) of curculigoside were harmful to cell growth. Therefore, curculigoside in the range of 1–100 μg/mL was not cytotoxic and could be used to stimulate hAFSC proliferation.
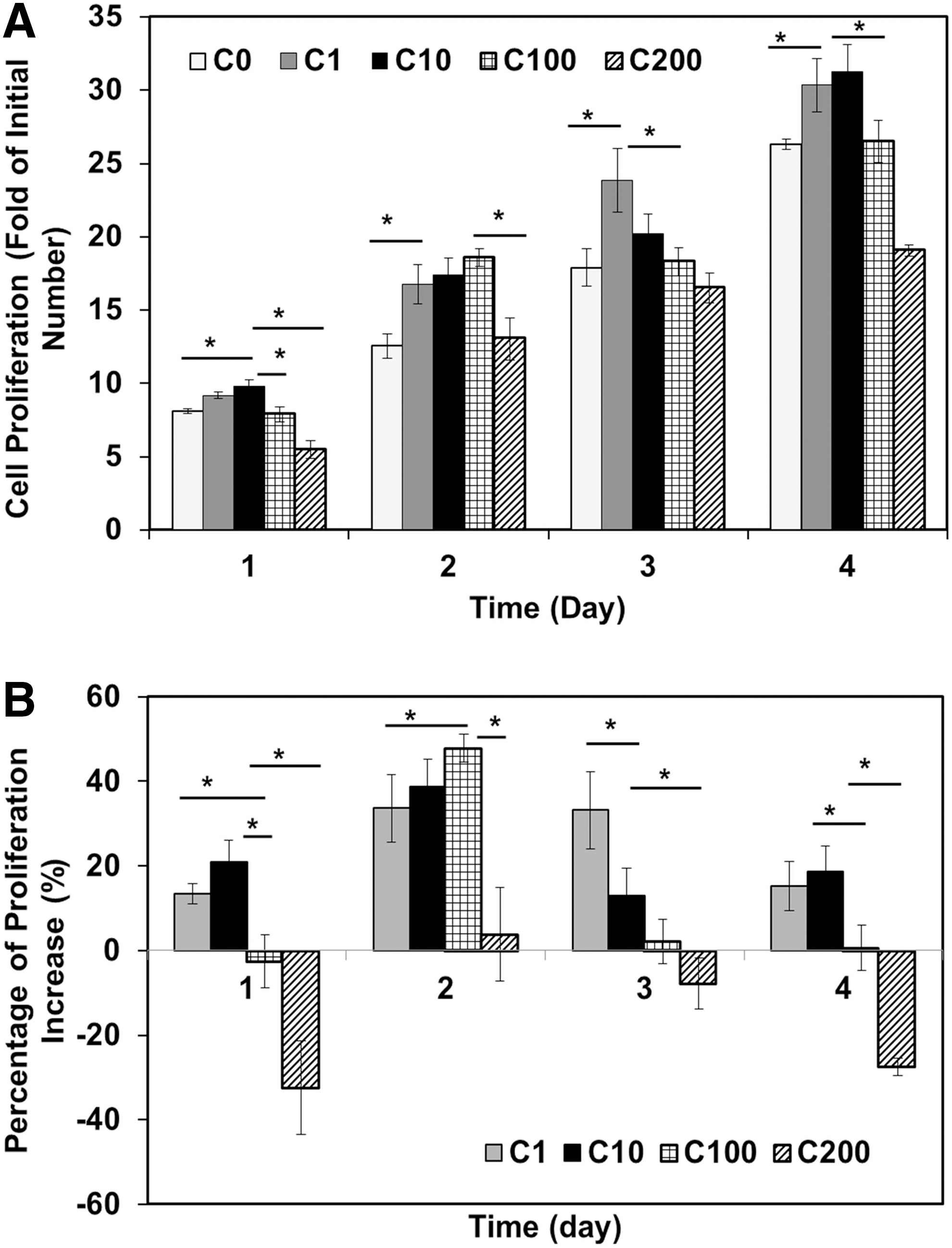
Effect of curculigoside on the proliferation of hAFSCs.
Effects of curculigoside on ALP activity of hAFSCs
The ALP activity was used to indicate the early stages of osteogenic differentiation of hAFSCs. ALP is a significant enzyme in the process of bone formation, which enhances the mineralization of bone matrix by increasing the phosphorous concentration [8,21]. Curculigoside was shown to increase the ALP activity of hAFSCs in a dose-dependent manner in the range of 1–100 μg/mL during a 7-day culture (Fig. 3). The ALP activity increased 26.3% and 356% in the presence of 10 and 100 μg/mL curculigoside, respectively. The ALP activity of hAFSCs treated with 200 μg/mL curculigoside increased 251%, which was lower compared with hAFSCs treated with 100 μg/mL curculigoside.

ALP activity of hAFSCs after curculigoside treatment. hAFSCs were treated with curculigoside at different concentrations (1–200 μg/mL). *P<0.05; **P<0.01. The percentage of increase was calculated as
Effects of curculigoside on calcium deposition
The ARS staining after 28 days of curculigoside treatment was performed to detect the presence of calcium. In the presence of curculigoside, more calcium deposition was observed compared to the control group especially for the 100 μg/mL group (Fig. 4A). The osteogenic differentiation of hAFSCs was also investigated by quantifying the calcium deposition on day 14 (Fig. 4B). Consistently, in the presence of curculigoside, more deposited calcium was formed than in the control group. Compared to the control group, the calcium content increased 40.5%, 92.7%, and 367% in the presence of 1, 10, and 100 μg/mL curculigoside, respectively. The calcium content in the 200 μg/mL curculigoside group was less compared with the 100 μg/mL group, increasing 243% compared to the control. Thus, calcium deposition increased in a dose-dependent manner at the concentration of 1–100 μg/mL curculigoside.
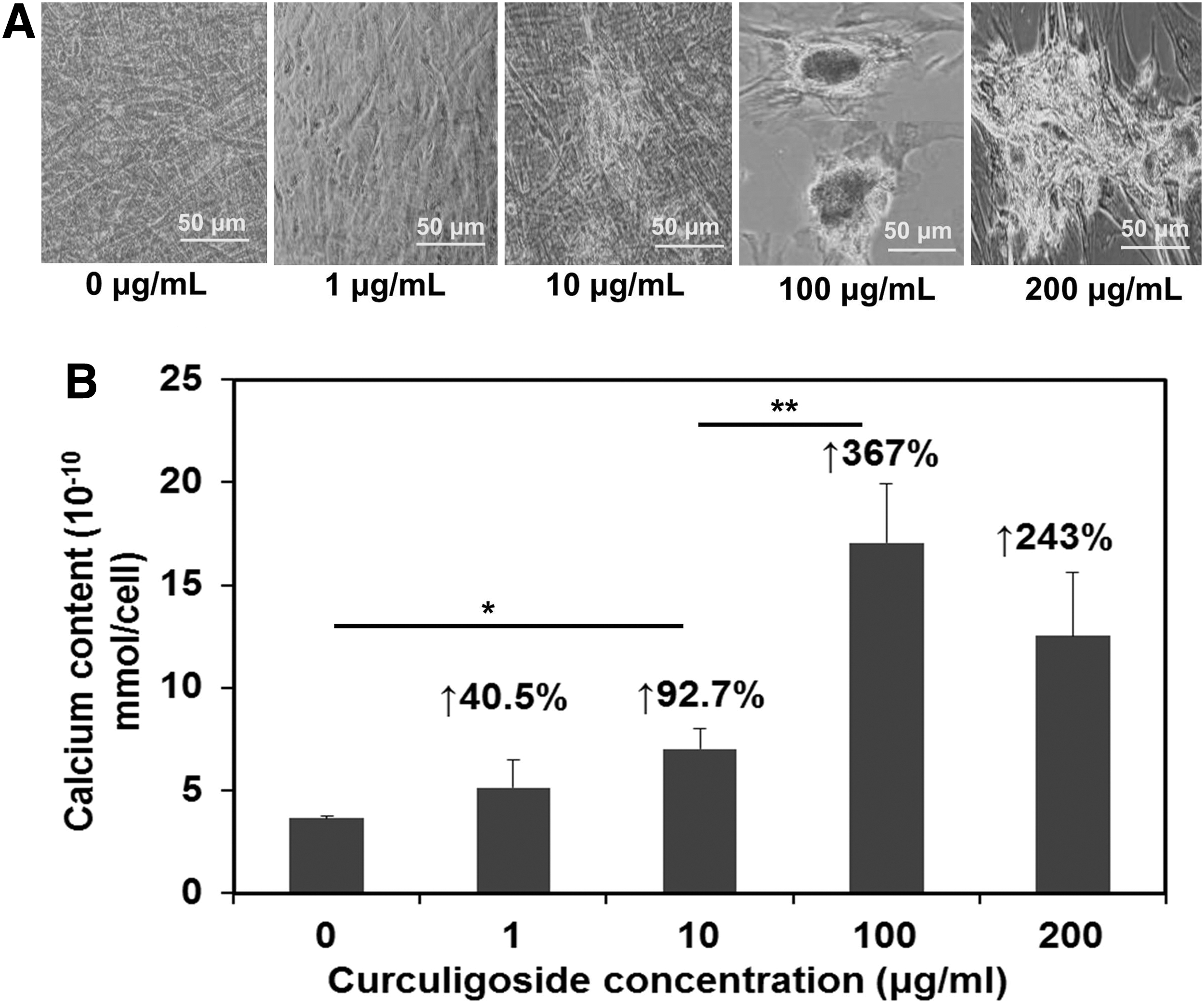
Osteogenic differentiation of hAFSCs after curculigoside treatment. hAFSCs were treated with curculigoside at different concentrations (1–200 μg/mL).
Effects of curculigoside on the expression of osteogenic genes
To further confirm the effects of curculigoside on osteogenic differentiation of hAFSCs, reverse transcriptase–PCR (RT-PCR) was performed to detect osteogenic marker genes OPN and Collagen I. The results showed that OPN and Collagen I expressions were significantly upregulated in curculigoside-containing media compared to the control group on day 21 (Fig. 5A, B). In addition, curculigoside increased the expressions of OPN and Collagen I in a dose-dependent manner at concentrations of 1, 10, and 100 μg/mL, while at 200 μg/mL the expressions of OPN and Collagen I were significantly reduced.
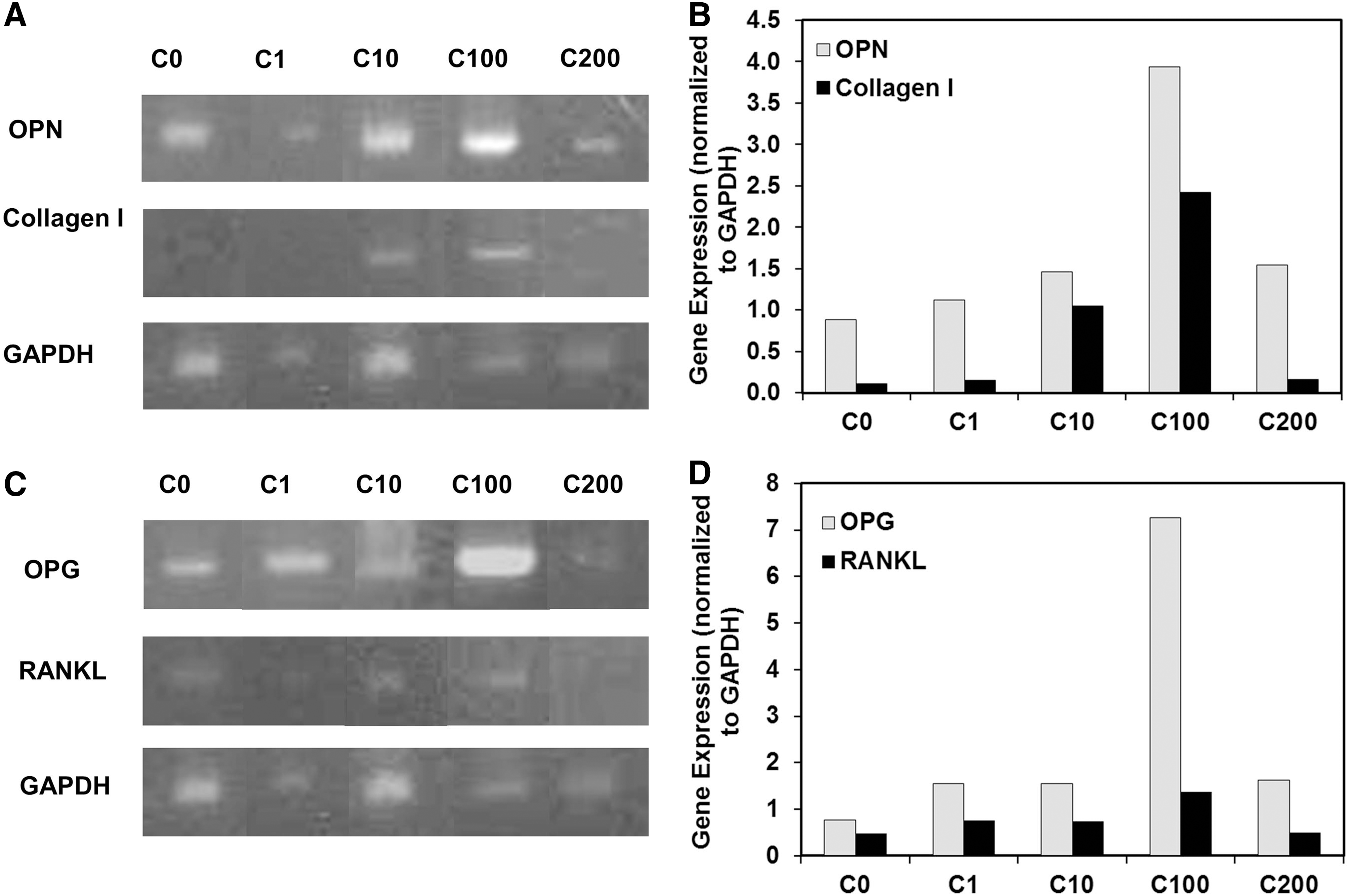
RT-PCR analysis of curculigoside-enhanced osteogenic differentiation of hAFSCs.
Effects of curculigoside on the osteoclast differentiation of hAFSCs
Expressions of OPG and RANKL were evaluated to assess the effect of curculigoside on the osteoclast differentiation of hAFSCs. The OPG expression was markedly increased with the concentration of curculigoside in the range of 1–100 μg/mL and then decreased at 200 μg/mL (Fig. 5C, D). RANKL, although weakly expressed, also showed a dose-dependent increase in the range of 1–100 μg/mL and a reduced expression at 200 μg/mL. The ratio of OPG to RANKL (OPG/RANKL) is a critical index of osteoclastogenic inhibition [22]. In this study, the OPG/RANKL ratio increased from 1.6 to 5.3 as the curculigoside concentration increased from 0 to 100 μg/mL (Table 2). Compared to the control, the OPG/RANKL ratio increased 28.9%, 30.7%, 233%, and 104% with curculigoside at 1, 10, 100, and 200 μg/mL, respectively, indicating that curculigoside inhibited osteoclastogenesis by increasing the relative portion of OPG expression.
Effects of curculigoside on Wnt/β-catenin signaling pathway
The canonical Wnt signaling pathway was reported to be involved in the osteogenic differentiation of MSCs [23 –25]. Therefore, the effects of curculigoside on the Wnt signaling pathway were studied with the cells harvested on day 21. As can be seen in Figure 6, the mRNA expression levels of Wnt-related regulators, including β-catenin and cyclin D1, were significantly upregulated in the presence of curculigoside in a dose-dependent manner at 1, 10, and 100 μg/mL. However, at 200 μg/mL, expressions of β-catenin and cyclin D1 were significantly reduced compared to 100 μg/mL. The expression of RUNX2 was weakly detected in the presence of curculigoside. To confirm the effect of Wnt signaling, Wnt-inhibitor DKK-1 was added into the curculigoside-treated hAFSC culture. As shown in Figure 7, the presence of DKK-1 reduced the ALP activity to 1.52±0.19 from 2.55±0.23 nmol/min/mg protein for the culture without DKK-1 treatment. Although BMP signaling was also reported to regulate osteogenic differentiation, BMP-related regulators, such as BMP-2 and BMP-4, were not detected in this study (data not shown).
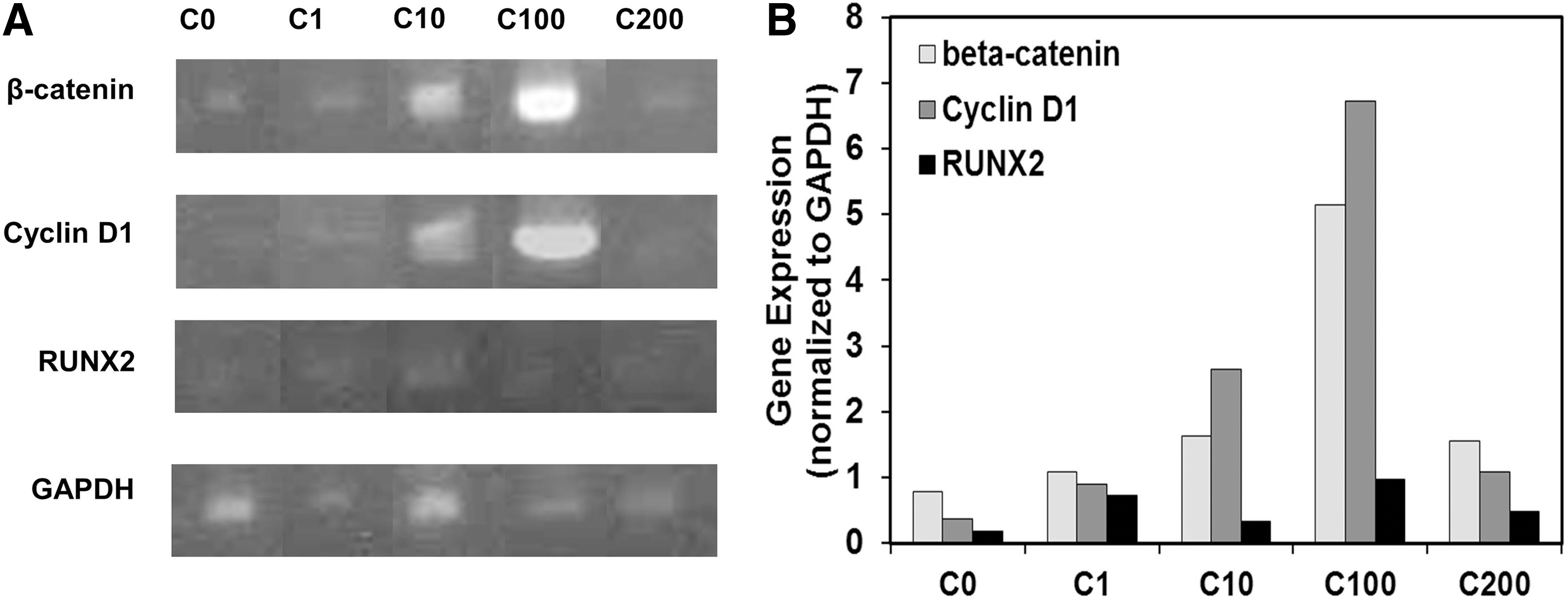
RT-PCR analysis of curculigoside-enhanced Wnt signaling of hAFSCs.
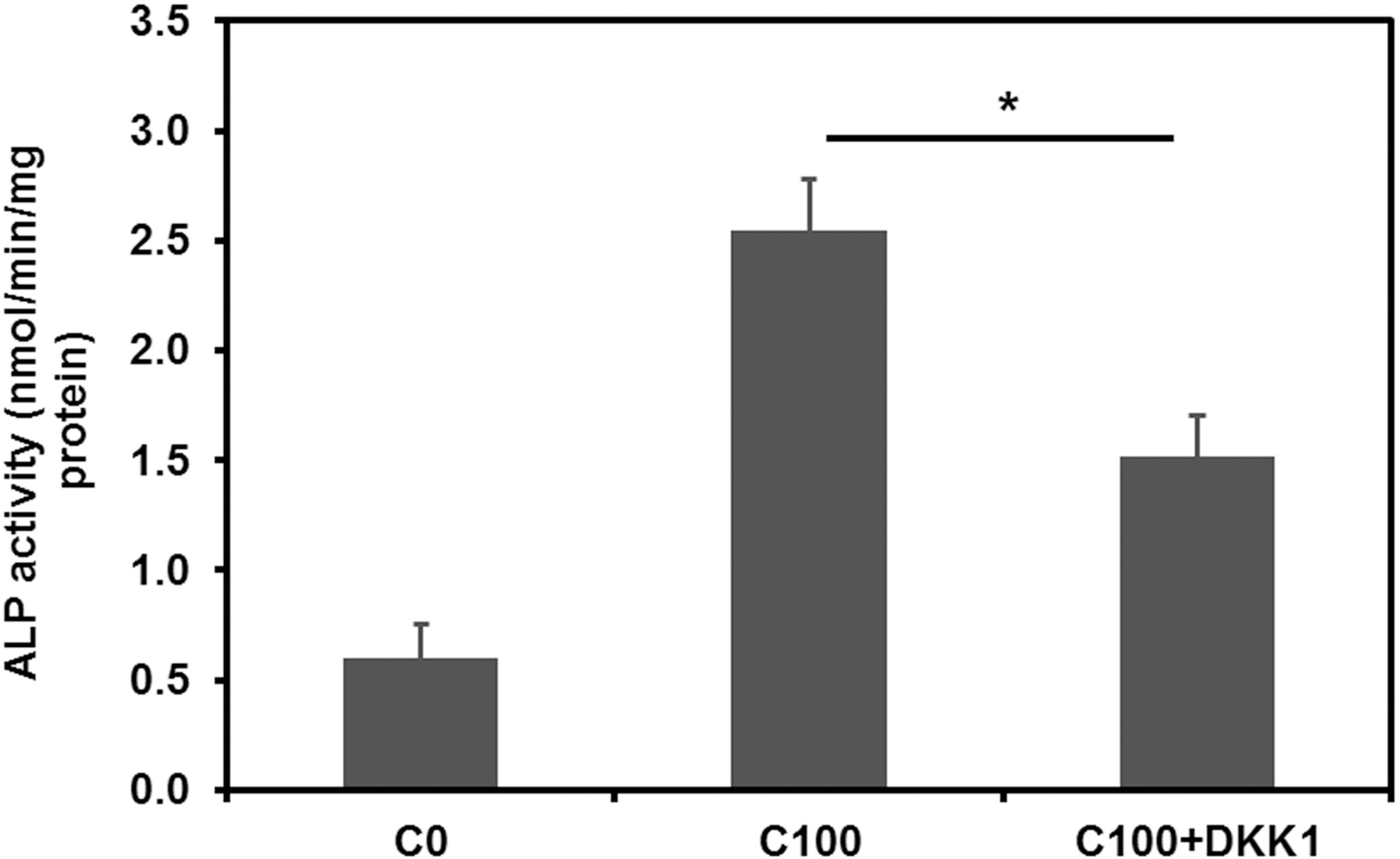
Effect of Wnt inhibitor DKK-1 on ALP activity of hAFSCs treated with curculigoside. C0, control; C100, 100 μg/mL curculigoside; C100+DKK-1, 100 μg/mL curculigoside plus 0.2 μg/mL of DKK-1. *P<0.05.
Discussion
Chinese herb C. orchioides, growing in subtropical regions in Asia, has been used to maintain healthy energy and support bone healing for thousands of years [15,16]. The hypothesis of this study is that the components in C. orchioides should affect osteogenic differentiation of hAFSCs. Our results demonstrated that curculigoside, the main effective component of C. orchioides, promoted osteogenic differentiation of hAFSCs and concurrently inhibited the osteoclastogenesis.
The process of osteogenesis from stem cells was depicted into three major stages: early stage from stem cell to osteoprogenitor, intermediate stage from osteoprogenitor to preosteoblast, and late stage from preosteoblast to mature osteoblast (Fig. 8A) [23,26]. In this study, osteogenesis of hAFSCs was found to be promoted by curculigoside at the early, intermediate, and late stages, which were proved by the increased ALP activity on day 7, the increased calcium deposition on day 14, and the upregulated expressions of OPN and Collagen I on day 21, respectively. The addition of curculigoside exhibited a biphasic effect on cell proliferation and ALP activity of hAFSCs. At 200 μg/mL, curculigoside suppressed the growth of hAFSCs, while at lower concentrations (<200 μg/mL), curculigoside enhanced cell proliferation. The ALP activity was enhanced by curculigoside in a dose-dependent manner in the range of 1–100 μg/mL. Similarly, gene expressions of OPN and Collagen I peaked at 100 μg/mL. Therefore, at the appropriate dose level, curculigoside could significantly enhance osteogenesis of hAFSCs.
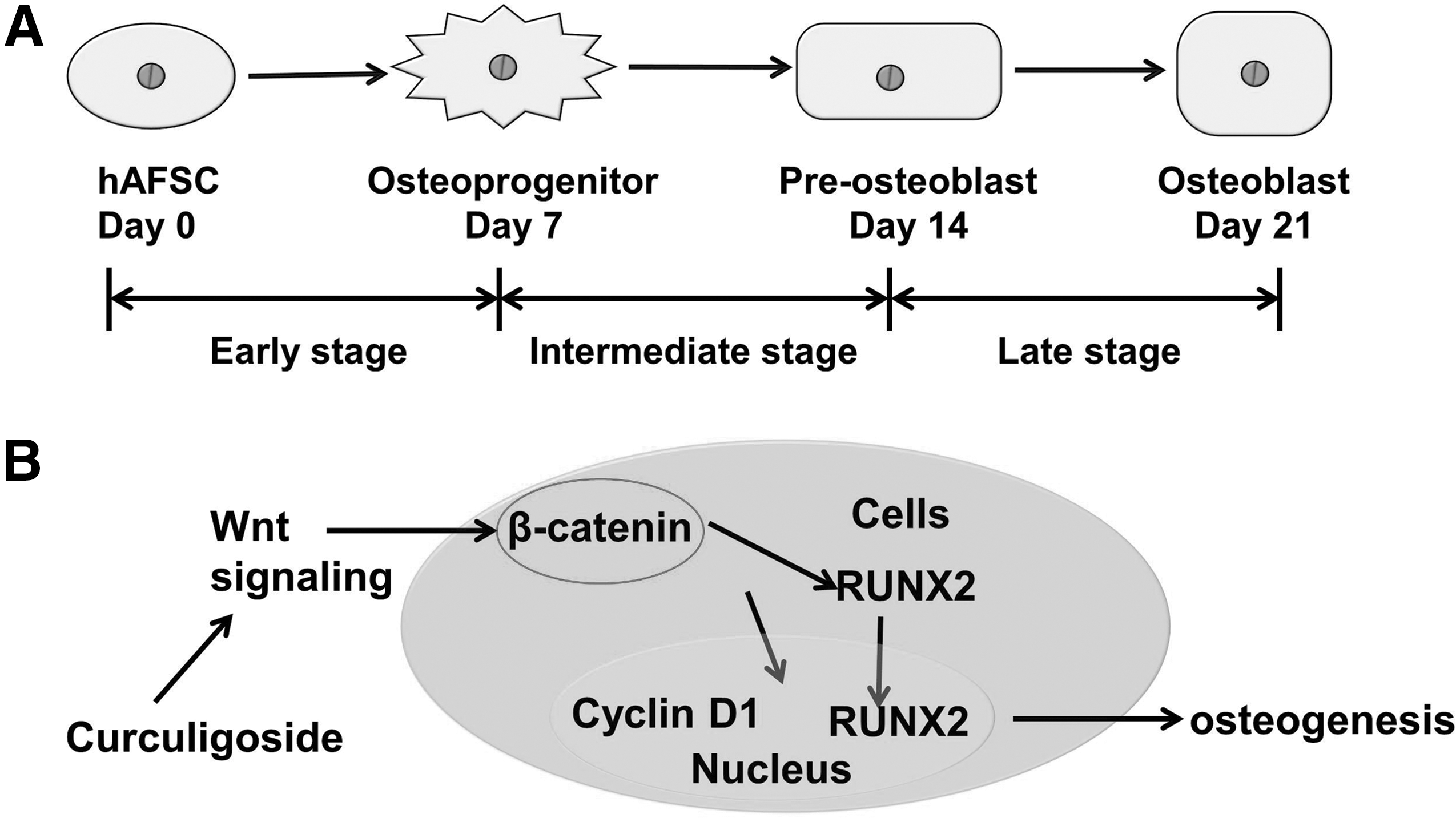
Schematic illustration of Wnt signaling pathway in osteogenic differentiation of hAFSCs.
Interactions between osteoblasts and osteoclasts maintain the balance between the bone formation and bone resorption [27]. Osteoblasts are responsible for the secretion of new bone matrix (bone formation), and osteoclasts deal with breaking the bone down (bone resorption). An imbalance in the regulation of bone formation and bone resorption results in many bone diseases, including osteoporosis [28]. RANKL is known to be the major factor responsible for osteoclast differentiation by providing a signal to osteoclast progenitors through the membrane-anchored RANK [22]. OPG is a decoy receptor binding to RANKL and its expression inhibits osteoclast differentiation [22]. Therefore, the ratio of OPG/RANKL is an essential indicator in the regulation of osteoblast and osteoclast formation. An increase in the OPG/RANKL ratio favors bone formation, while a decrease in the ratio favors bone resorption. Our study showed that curculigoside significantly increased the OPG/RANKL ratio during hAFSC osteogenic differentiation especially at 100 μg/mL, indicating the inhibition effect of curculigoside on osteoclastogenesis. Thus, curculigoside could provide a new approach to treat hAFSCs and other MSCs for cell-based therapy in bone resorbing diseases such as osteoporosis.
Osteogenesis is a complicated process involving several signaling pathways especially the canonical Wnt pathway [29 –31]. In our study, the mRNA expressions of β-catenin and Cyclin D1 were upregulated by curculigoside along with OPN, Collagen I, and OPG/RANKL, while in the presence of DKK-1, a specific inhibitor for Wnt/β-catenin signaling, the ALP activity was remarkably reduced (Fig. 7). The Wnt/β-catenin signaling is critical for osteogenic differentiation and bone formation (Fig. 8B) and has been shown to participate in embryonic skeletal patterning, fetal skeletal development, and adult skeletal remodeling [23,29]. Wnt signaling represents both a cell autonomous mechanism for inducing osteoblast functions and a mechanism in fully differentiated osteoblasts for stimulating OPG production to inhibit osteoclast formation [30,32]. Therefore, the activation of canonical Wnt signaling resulted in a higher bone density and higher expression of ALP [33,34]. Wnt signaling contributes to osteoblast differentiation through β-catenin activation [31]. A high level of β-catenin enhances bone formation, whereas knockdown of the β-catenin gene at an early developmental stage causes abnormal osteoblast differentiation [30,35,36]. β-catenin also regulates osteoclastogenesis by affecting the expressions of OPG and RANKL [36]. Cyclin D1 is the target gene of the Wnt pathway and is upregulated when Wnt/β-catenin signaling is activated [37]. RUNX2 was reported to integrate Wnt signaling for mediating the process of osteoblast differentiation and play an essential role in the secretion of bone-specific extracellular matrix [38,39]. In this study, the expressions of β-catenin and Cyclin D1 were enhanced in the presence of curculigoside, indicating that Wnt/β-catenin signaling is involved in the stimulating effect of curculigoside on osteogenesis of hAFSCs and the inhibition effect on osteoclastogenesis.
Generally, osteoprotective medicines have potential adverse effects. For instance, increased risk of cardiovascular diseases was reported to be associated with hormone replacement therapy [40] and antiresorptive bisphosphonate might result in upper gastrointestinal tract complications as well as long-range effects on skeletons [41]. Curculigoside is a natural ingredient in C. orchioides and can be used as a nutrient supplement in the diet. Our results showed that 1–100 μg/mL curculigoside had no cytotoxicity to hAFSCs, while 200 μg/mL might inhibit cell growth and reduced osteogenic marker expression. It is not clear why the higher concentration of curculigoside had a significant inhibition effect on proliferation. It is also not clear why osteogenic differentiation increased more at 100 μg/mL than at 200 μg/mL curculigoside. It is likely that higher concentrations could induce more complex effects that cannot be explained by a single pathway. The dose-dependent effect of Chinese herbal medicine has also been observed in our previous studies during cell-based assay development [42,43]. Therefore, with an appropriate dose, curculigoside-enhanced hAFSC osteogenic differentiation could be a promising strategy to treat bone disorders such as osteoporosis.
Conclusions
Osteoporosis is a crucial public health problemm which can lead to fracture and deformities. Osteogenic differentiation of hAFSCs could be used for the treatment of osteoporosis and other bone-related diseases. Chinese herb C. orchioides, which is safe and cheap, has been used for fracture treatment and bone healing for thousands of years. The present study demonstrated that curculigoside, the main effective component of C. orchioides, promoted proliferation and osteogenesis and concurrently inhibited osteoclastogenesis of hAFSCs. Moreover, this study elucidated that curculigoside functions through the Wnt/β-catenin signal transduction pathway. Due to the economic and safety advantages, curculigoside-enhanced osteogenesis of hAFSCs would be a promising and attractive treatment strategy to augment bone formation in patients with osteoporosis and other bone disorders.
Footnotes
Acknowledgments
This work was supported, in part, by the Alumni Grants for Graduate Research and Scholarship (AGGRS) of The Ohio State University. We would like to acknowledge Dr. Anthony Atala and Dr. James Yoo of the Wake Forest Institute for Regenerative Medicine (Winston-Salem, NC) for kindly providing hAFSCs used in this study. We also would like to acknowledge Dr. Sebastien Sart for the assistance in ImageJ analysis.
Author Disclosure Statement
No competing financial interests exist.