
Abstract
MicroRNAs (miRNAs) regulate myriad biological processes; however, their role in cell fate choice is relatively unexplored. Pluripotent NT2/D1 embryonal carcinoma cells differentiate into an epithelial/smooth muscle phenotype when treated with bone morphogenetic protein-2 (BMP-2). To identify miRNAs involved in epithelial cell development, we performed miRNA profiling of NT2/D1 cells treated with BMP-2 at 6, 12, and 24 h, and on days 6 and 10. Integration of the miRNA profiling data with previously obtained gene expression profiling (GEP) data of NT2/D1 cells treated with BMP-2 at the same time points identified miR-18b and miR-518b as the top two miRNAs with the highest number of up-regulated predicted targets with known functions in epithelial lineage development. Silencing of miR-18b and miR-518b in NT2/D1 cells revealed several up-regulated TFs with functions in epithelial lineage development; among these, target prediction programs identified FOXN1 as the only direct target of both miRNAs. FOXN1 has previously been shown to play an important role in keratinocyte differentiation and epithelial cell proliferation. NT2/D1 and H9 human embryonic stem cells with silenced miR-18b and miR-518b showed up-regulation of FOXN1 and the epithelial markers CDH1, EPCAM, KRT19, and KRT7. A 3′UTR luciferase assay confirmed FOXN1 to be a target of the two miRNAs, and up-regulation of FOXN1 in NT2/D1 cells led to the expression of epithelial markers. Overexpression of the two miRNAs in BMP-2-treated NT2/D1 cells led to down-regulation of FOXN1 and epithelial lineage markers. These results show that miR-18b and miR-518b are upstream controllers of FOXN1-directed epithelial lineage development.
Introduction
M
Transcription factors (TFs), Polycomb group proteins, and miRNAs play key roles in regulating decisions with regard to lineage development in pluripotent mammalian cells [16 –20]. MiRNA-initiated regulatory effects can be transmitted downstream by their target TFs; however, the mechanisms involved in controlling the miRNA-directed TF function in lineage development are not well understood. In the mouse, loss of Foxn1 function results in the nude phenotype, which is characterized by abnormal skin, thymus, and mammary gland morphogenesis [21 –24]. Recent studies in mice have shown that Foxn1 is a target of the homeobox gene Hoxc13, a TF that is involved in hair follicle and nail differentiation [25]. Foxn1 is known to be induced during early stages of the epithelial program in the skin and during terminal differentiation of keratinocytes in culture [26,27]. However, upstream regulation of Foxn1 in promoting lineage development remains uncharacterized. Here, we show that FOXN1 is a target of miR-18b and miR-518b, and their silencing leads to FOXN1 up-regulation and epithelial lineage development in NT2/D1 and H9 human embryonic stem (hES) cells.
Materials and Methods
Cell culture and immunofluorescence
NT2/D1 cells were cultured and treated with BMP-2 as previously described [12 –14]. For immunofluorescence (IF) analysis, cells were fixed in 4% paraformaldehyde. Fixed cells were permeabilized in 0.3% Triton X-100 in phosphate-buffered saline containing 5% normal goat serum and subjected to IF analyses as previously described [13]. The primary antibodies used were monoclonal antibodies against mouse antigens Oct3/4 (1:100; Santa Cruz Biotechnology, Santa Cruz, CA), Keratin7, and Keratin19 (1:100; Abcam, Cambridge, MA) and a polyclonal rabbit antibody against FOXN1 (1:200; Sigma, St. Louis, MO). Appropriate fluorescence-labeled secondary antibodies were used for visualization.
Gene expression profiling of BMP-2 time course in NT2/D1 EC cells
The gene expression profiling (GEP) of NT2/D1 BMP-2 time course used in this analysis was previously obtained by us using Affymetrix U133 A plus B microarrays, as reported [14].
miRNA isolation and RT-PCR
RNA was isolated using the miRVANA Isolation Kit (Life Technologies, Grand Island, NY). MiRNAs were assayed with Taqman probes and primer sets in an applied Biosystems 7500 RT-PCR system (Life Technologies). For RT-PCR analysis of mRNA, total RNA was treated with Turbo DNA-free kit (Life Technologies) and reverse transcribed with superscript III first-strand synthesis system (Life Technologies). PGK1 was used as the internal normalization control, because its CT value was consistent in all experiments.
miRNA profiling and data analysis
Total RNA from three time points of the BMP-2 treatment of NT2/D1 cells [early (12 h), middle (Day 1), and late (days 6 and 10)] [14] was subjected to miRNA expression analysis using the Agilent Human V2 array. The ensuing data were quantile-normalized using the Partek® Genome Suite software, version 6.5 (Partek, Inc., St. Louis, MO), and the significantly differentially expressed miRNAs (p<0.05) were identified using the Significance Analysis of Microarrays (SAM) algorithm (supp ref 2). SAM analysis was performed on the normalized data with a two-class unpaired t-test option and 500 iterations. The miR-18b and miR-518b silenced GEPs were also analyzed similar to the time course data, and heatmaps were generated using the GENE-E software from the Broad Institute. Gene Ontology analysis was performed using the GoMiner online tool (supp ref 3). The miRNA predicted targets and GEP data were integrated using Ge-Workbench software selecting only significantly up-regulated targets (P≤0.01).
LNA transfection of NT2/D1 cells
0.4 million NT2D1 cells were plated in 6-well plates at 1 day before transfection. 80 nM LNA (Exiqon, Woburn, MA) and 14 μL siRNA transfection reagent X-tremeGENE (Roche, Indianapolis, IN) were used to transfect each well according to the manufacturer's instructions. For double transfection, 40 nM of each anti-miRNA-probe with 14 μL of the X-tremeGENE reagent (Roche) were used in each six-well plate. All the LNA oligonucleotides used for silencing had the phosphorothioate (PS) backbone, Power Inhibitor (PI), and are listed next:
oligo1:hsa-mR-18b-kd-PS/5TYE665/A*A*C*T*G*C*A*C*T*A*G*A*T*G*C*A*C*C*T*T
oligo2:hsa-miR-518b-kd-PS/5TYE563/C*T*C*T*A*A*A*G*G*G*G*A*G*C*G*C*T
oligo3:Negative_control_power_inhibitor/5TYE563/G*T*G*T*A*A*C*A*C*G*T*C*T*A*T*A*C*G*C*C*C*A
Fluorescence-activated cell sorting
NT2/D1 cells that contained 5′ labeled TYE665 or TYE 563-conjugated LNA were sorted on a BD FACS Aria cell sorter baesd on MSKCC Flow Cytometry Core facility protocols.
Luciferase reporter transfection
For the 3′UTR luciferase assay, 0.04 million 293T cells were plated in each well of 24-well plates and transfected with either 200 ng of WT FOXN1 3′UTR cloned in pEZX-MT0–1 vector or MUT1/MUT2 FOXN1 3′UTR cloned in pEZX-MT0-1 vector (GeneCopoeia, Rockville, MD) as well as 50 nM pre-miR miRNA precursors of miR-18b or miR-518b (Life Technologies
Transient overexpression of FOXN1 in NT2/D1 cells
0.5 million NT2D1 cells were seeded in six-well plates at 1 day before transfection. 1ug of the construct pReceiver-Lv156 with FOXN1 or scramble clone (GeneCopoeia) were transfected with the help of Xtreme-GENE transfection reagent (Roche) into each well. Media were changed after 5 h of transfection. Cells were collected for downstream studies at 48 h.
Results
miR-18b and miR-518b regulate epithelial lineage differentiation
We previously obtained GEP of a time course experiment of BMP-2 treatment of NT2/D1 cells that induced epithelial lineage [14]. In this study, epithelial lineage markers such as KRT19 and KRT7 were up-regulated by 24 h, and the cells were fully differentiated into an epithelial/smooth muscle lineage by day 10 [14]. To explore the role of miRNAs in this program, we performed miRNA profiling of BMP-2-treated NT2/D1 cells using the Agilent Human V2 miRNA v.10.1 array. For data analysis, we considered miRNA expression at early (12 h), mid (Day 1), and late (Day 6 and 10) points of differentiation based on our previous GEP results [14]. A set of miRNAs was identified whose expression was up- or downregulated as the cells differentiated; 53 were significantly down-regulated (P<0.01), of which 36 had predicted target genes that were known to play a role in epithelial lineage development (Supplementary Table S1; Supplementary Data are available online at
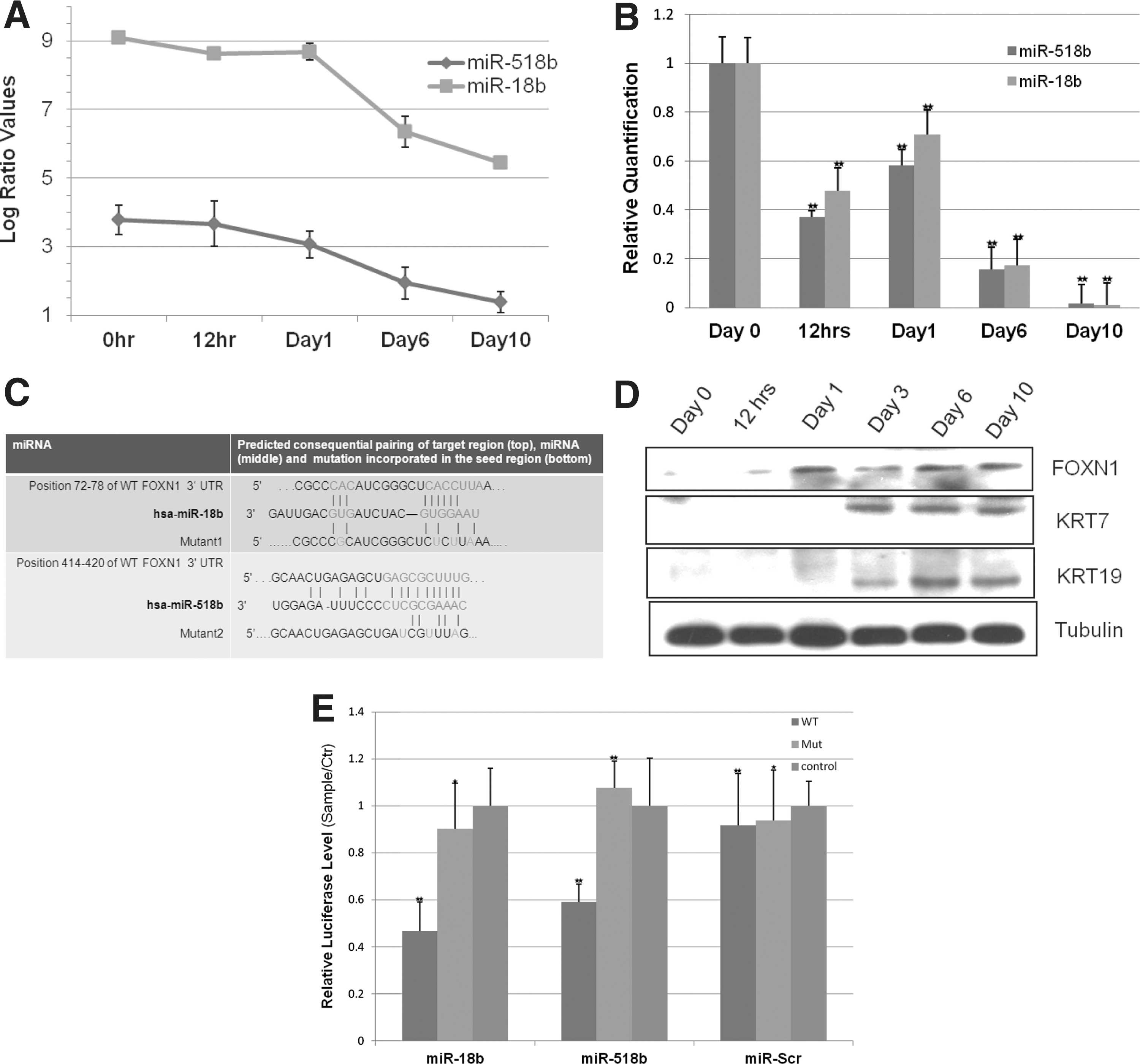
FOXN1 is a target of miR-18b and miR-518b.
FOXN1 is a target of miR-18b and miR-518b in epithelial lineage development
Silencing of miR-18b and miR-518b in NT2/D1 cells identified a number of TFs with suggested roles in epithelial lineage development. Of these, six were up-regulated in the silenced cells as assayed by RT-PCR (FOXN1, GATA3, JUN, MSX1, MSX2, and GLI2) (Supplementary Fig. S1). We used the target prediction programs TargetScan, miRanda, Pictar, and DIANAmT to identify; from among these, TFs may be direct targets of both miR-18b and miR-518b and we found FOXN1 to be the only one. As shown in Fig. 1C, nucleotide positions 72–78 and 414-420 of FOXN1 3′UTR, respectively, were aligned with miR-18b and miR-518b. Previous studies have established an important role for FOXN1 in epithelial cell growth and differentiation, along with high expression in epithelial cells [25 –27]. Furthermore, FOXN1 and the epithelial markers KRT7 and KRT19 were induced in response to BMP-2 treatment of NT2/D1 cells. Together, these results suggested that the two miRNAs target FOXN1 in promoting epithelial lineage development (Fig. 1D).
Ectopic expression of miR-18b and miR-518b targets FOXN1 3′UTR reporters
To explore the possible role of endogenous miR-18b and miR-518b in repressing FOXN1, we assayed, using 293T cells, 3′UTR luciferase reporter constructs that have either the wild-type 3′UTRs (WT) or mutant 3′UTRs of FOXN1 [a 4 bp mutation (MUT1) in the target site of miR-18b and a 3 bp mutation in the target site of miR-518b (MUT2)], along with a no UTR control vector. In comparison to MUT1 and MUT2, 24 h after transfection, there was significant repression in the WT 3′UTR luciferase reporter activity, confirming that the two miRNAs specifically target FOXN1 (Fig. 1E).
FOXN1 expression is modulated by miR-18b and miR-518b during epithelial lineage induction
To confirm that down-regulation of miR-18b and miR-518b leads to up-regulation of FOXN1 and epithelial lineage induction, we silenced the two miRNAs in NT2/D1 cells by transfection of the antisense TYE 665+ conjugated miR-18b PI and the antisense TYE 563+ miR-518b PI, followed by FACS sorting of the positive cells for downstream studies. In comparison to the LNA scramble PI, cells with the silenced miRNAs had a differentiated phenotype with morphologically flat cells. RT-PCR analysis showed that FOXN1 and the epithelial markers KRT and CDH1 were up-regulated after silencing of the two miRNAs individually or after co-silencing, whereas EPCAM was up-regulated only when both were co-silenced. (Fig. 2A–C). We confirmed the expression of FOXN1 and KRT in the silenced cells by IF (Fig. 3). Microarray analysis of the silenced cells further confirmed the up-regulation of epithelial-associated genes and the down-regulation of pluripotency-associated genes (Supplementary Figs. S2–S4). GoMiner analysis of the up-regulated genes in cells silenced for both the miRNAs identified epithelial-related developmental processes such as skin morphogenesis and epithelial cell maturation to be among the highly enriched categories (Supplementary Tables S3, S4).
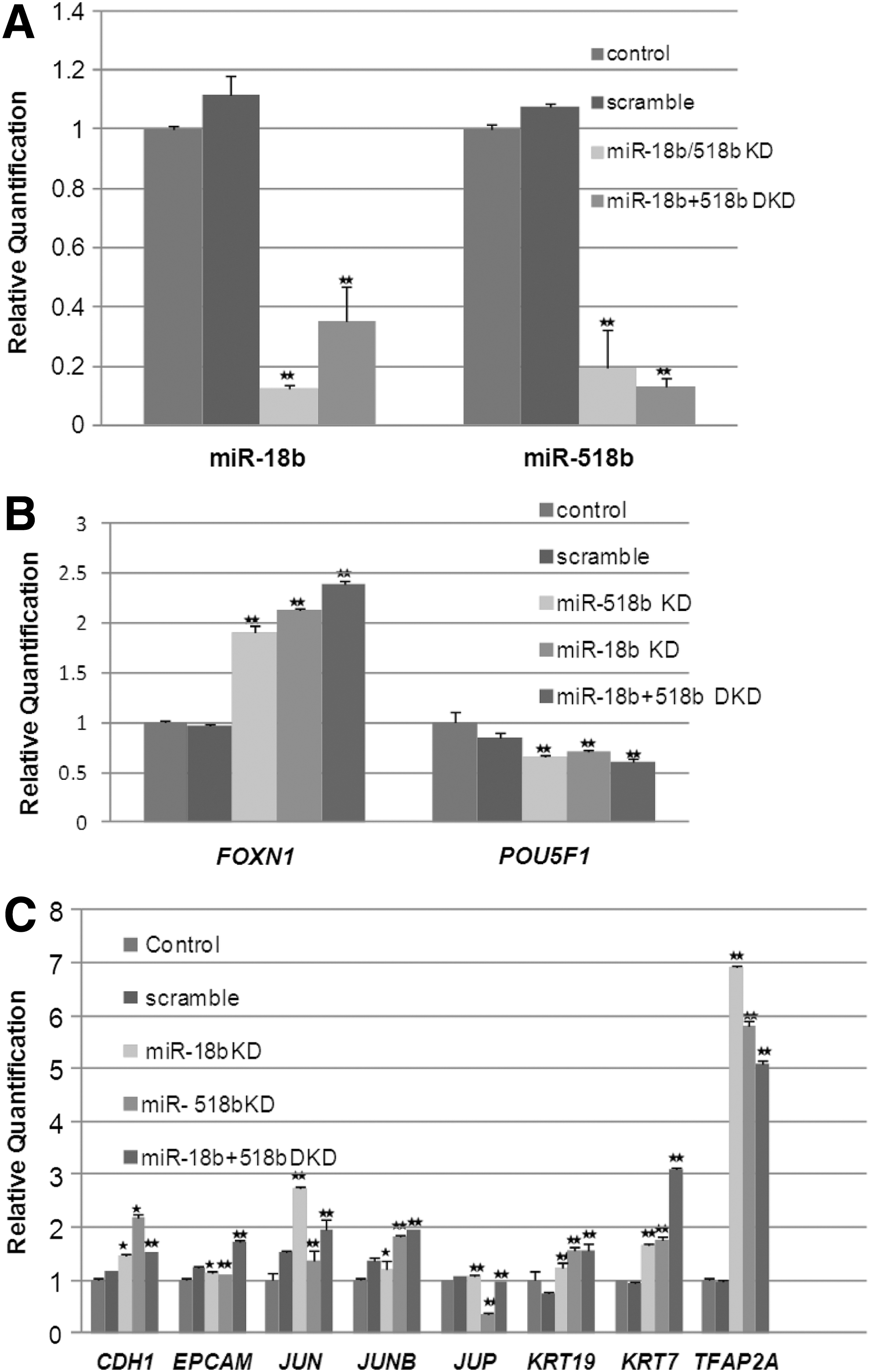
Gene expression analysis of miR-18b and miR-518 knockdown in NT2/D1 cells.
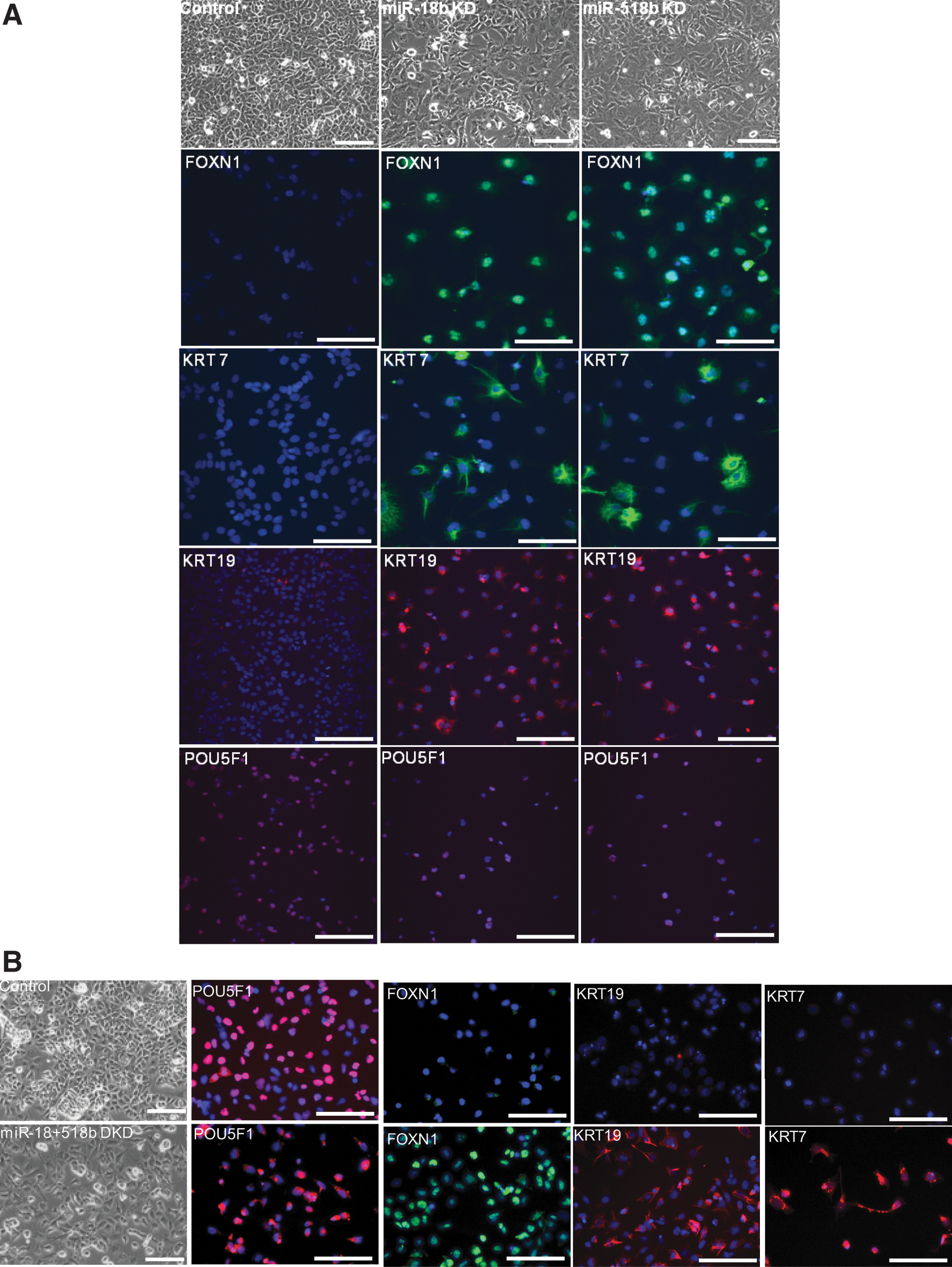
FOXN1 is up-regulated as cells differentiate toward epithelial lineage by down-regulation of miR-18b and miR-518b.
To evaluate the potential role of the two miRNAs in other pluripotent cells, we silenced miR-18b and miR-518b in H9 hES cells following the same methods as used for NT2/D1 cells (see Supplementary Methods). Silencing yielded similar results to those in NT2/D1 cells, although FOXN1 up-regulation was more pronounced in miRNA co-silenced H9 hES cells than in NT2/D1 cells (Supplementary Fig. S5). We conclude from these results that the down-regulation of miR-18b and miR-518b in EC as well as in hES cells leads to derepression of FOXN1, ultimately driving the cells into epithelial lineage.
Overexpression of FOXN1 in NT2/D1 cells and miR-18b, miR-518b in BMP-2-treated NT2/D1 cells confirms their role in epithelial lineage induction
To confirm the role of FOXN1 in epithelial lineage development, we transiently overexpressed the gene in NT2/D1 cells. As shown in Fig. 4, a number of epithelial marker genes, including CDH1, EPCAM, KRT19, and KRT17, were up-regulated. We overexpressed miR-18b and miR-518b in NT2/D1cells at 6 h of BMP treatment when an epithelial lineage program is initiated [14], and harvested cells after a further 24 h for downstream studies. We confirmed the overexpression of the two miRNAs by RT-PCR (Supplementary Fig. S6A). The overexpression of one miRNA did not affect the level of expression of the other, confirming the specificity of the probes used (Supplementary Fig. S6A). FOXN1 protein was down-regulated in miR-18b, and miR-518b overexpressed BMP-2-treated NT2/D1 cells as confirmed by western blot (Supplementary Fig. S6B). RT-PCR analysis of miRNA-overexpressed BMP-treated NT2/D1 cells showed down-regulation of the epithelial marker genes EPCAM, CDH1, and KRT, proving that overexpression of the miRNAs led to their repression (Supplementary Fig. S6C).

Up-regulation of FOXN1 in NT2/D1 cells promotes expression of epithelial marker genes
Discussion
The regulation of TFs that mediate biological processes by miRNAs is increasingly recognized in many contexts; however, the regulation of TFs that mediates lineage development by miRNAs has been less well investigated. MiR-1 and miR-133 are cardiac and skeletal muscle-specific miRNAs, where miR-1 promotes the differentiation of cardiac progenitors and miR-133 inhibits the differentiation of skeletal myoblasts in mouse [5,28,29]. HAND2, a TF that promotes ventricular cardiomyocyte development and plays an essential role in cardiac morphogenesis, is a direct target of miR-1and helps in regulating the balance between muscle differentiation and proliferation during cardiogenesis [28,29]. In another example, the zinc finger E-box binding homeobox1 TFs ZEB1 and ZEB2 and the miR-200 family regulate the switch between epithelial and mesenchymal states through reciprocal inhibition [30].
Here we show, for the first time, that miR-18b and miR-518b constitutively repress the expression of FOXN1, a key regulator of epithelial lineage; their down-regulation releases FOXN1 repression, leading to lineage development in pluripotent NT2/D1 and hES cells. Human EC cells have been shown to be similar to ES cells in terms of their pluripotency characteristics and, hence, serve as good models to study in vitro lineage differentiation [31 –33]. We found that the silencing of miR-18b and miR-518b individually led to up-regulation of FOXN1 and epithelial marker genes in NT2/D1 and hES cells; however, their co-silencing in NT2/D1 cells did not have a further additive effect, although co-silencing in hES cells had a complementary effect, causing a further increase in the expression of FOXN1 and the epithelial marker genes KRT7, EPCAM, and CDH1. These results show that although the miRNA-directed regulatory pathway to epithelial lineage development is common to both cell lines, they exhibit cell-line-specific differences in response.
MiR-18b is a part of the genomic cluster-hsa-mir-17 and the sequence family sf-hsa-miR-17; whereas miR-518b belongs to the genomic cluster-hsa-mir-498 and the sequence family sf-hsa-miR-515-1-3p (miRBase). Recent studies have suggested that miR-18b and miR-518b may play a role in tumor suppression. MDM2 has been shown to be a target of miR-18b, and its overexpression leads to down-regulation of MDM2 and activation of P53 in melanoma [34]. Similarly, in vitro studies have shown RAP1B to be directly regulated by miR-518b in esophageal squamous cell carcinoma (ESCC), and the down-regulation of miR-518b in ESCC tissues correlates with metastasis and survival [35]. So far, these two miRNAs have not been implicated in the regulation of developmental processes directly or through TFs.
FOXN1 is a member of FOX (forkhead box) gene family, which comprises at least 43 members that share the common property of directing tissue-specific transcription and lineage choice in human cells [36]. Among these, FOXN1 is a crucial regulatory TF of developmental processes, including normal thymic and epithelial skin development [37 –41]. In the mouse, Foxn1 has been shown to control the epithelial differentiation program by the regulation of Protein Kinase C activity [42]. In this study, we show that FOXN1 is regulated upstream by miR-18b and miR-518b in the epithelial lineage development; the down-regulation of these two miRNAs derepressed the expression of FOXN1, which initiates the downstream signaling required for the promotion of lineage development. We have, thus, identified a novel miRNA-regulated pathway of early epithelial lineage differentiation in pluripotent cells.
Footnotes
Acknowledgments
This study was supported by the Byrne Fund. The authors thank the MSKCC Genomics Core Facility for help with miRNA expression profiling. They also thank the MSKCC Flow Cytometry Core Facility for cell sorting.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.