
Abstract
Umbilical cord blood (UCB) is a good source of hematopoietic progenitors with increasing implementation in the clinical transplant setting. This study evaluates the molecular mechanisms of progenitor resistance to apoptosis triggered by Fas cross-linking. CD34+ and lineage-negative progenitors survive short-term ex vivo incubation and are not induced into apoptosis by Fas cross-linking. Furthermore, brief exposure of UCB cells to Fas-ligand for 24–48 h does not impair quantitative severe combine immune deficiency (SCID) reconstitution activity and appears to foster myelomonocyte reconstitution. The transcriptome of Fas receptor-positive CD34+ cells that survived an apoptotic challenge showed significant transcriptional upregulation of caspase-8, mucosa-associated lymphoid tissue lymphoma translocation gene-1 (MALT1), HtrA2, and GSK3β in addition to higher levels of c-FLICE inhibitory protein (FLIP), Bcl-2, and cytosolic inhibitor of apoptosis protein (cIAP) in all Fas-positive cells. Most prominent is the transcriptional upregulation of several key components the NFκB1 pathway including the membrane receptors TGF-β, interleukin-1 (IL-1), and TCR, the associated factor TNF receptor-associated factor-6 (TRAF6), and the converting enzymes TGF-β-activated kinase-1 (TAK1), double-stranded RNA-activated protein kinase (PKR), and α-catalytic subunit of IκB kinase (IKKα), that promote activation and nuclear translocation of this transcription factor. These data indicate that hematopoietic progenitors are not insensitive to apoptosis but are actively shielded from the extrinsic and intrinsic apoptotic pathways. This may occur through inherent transcriptional upregulation of the entire NFκB pathway in the presence of competent apoptotic signaling.
Introduction
M
On the one hand, the traditional concept has been dominated by the role of the TNF family receptors as mediators of apoptosis, one of the few irreversible events in cell development. Accordingly, the Fas/FasL interaction is considered to inhibit developing hematopoietic progenitors, similar to suppression of proliferation in the distal stages of differentiation. This contention is based on suppression of hematopoietic progenitor function in vitro [3,4,9], repression of clonogenic activity [4,10], and negative impact on human cell engraftment in immunocompromized mice and humans [10,11]. Supporting evidence is the apparent protection of hematopoietic cells by blocking antibodies and soluble FasL [12 –14], which exert antiapoptotic effects by preventing Fas trimerization [15]. All these ex vivo studies used growth factors and chemokines to support cells in culture for variable periods of time, yet such conditions induce cell cycling and generally impair engraftment [16]. Fas is also considered to be a partial mediator of the negative effects of TNF-α, due to inductive cross talk between these receptors in hematopoietic progenitors [14]. These data suggest that neutralization of the Fas/FasL interaction might be beneficial to hematopoietic cell function in the transplant setting, in particular due to involvement of this pathway in graft versus host disease [17,18].
On the other hand, Fas is ubiquitously upregulated soon after transplantation in murine and human hematopoietic reconstituting cells without causing apoptosis [10,19], but rather tansduces tropic signals [20]. Impaired engraftment of Fas and FasL-deficient bone marrow cells and enhanced clonogenic activity of progenitors induced by Fas trimerization reflect a physiological role of Fas in initial stages of murine hematopoietic progenitor engraftment and function [19]. Consistently, overexpression of FasL improves hematopoietic cell engraftment by autocrine stimulation of progenitor activity [20], activation of stromal Fas, and abrogation of alloimmune responses [21,22]. Soluble members of the TNF family also activate hematopoietic progenitors through TNF receptor-1 (TNF-R1) and consequently caspase-8 in humans and mice [23,24], and TRAIL-R1 in human cells or TRAIL-R2 (the only murine receptor) [25]. Our findings indicate that the TNF family receptors foster reconstitution under conditions of stress hematopoiesis and inhibition of mechanisms related to signaling pathways of these receptors might impair the function of hematopoietic reconstituting cells.
In this study, we assess the involvement of the Fas/FasL interaction in umbilical cord blood (UCB), a rich source of progenitors for transplantation. Characterization of apoptosis is particularly significant because thawing of UCB cells is associated with early cell death [26], which reduces dramatically the success of engraftment and hematopoietic reconstitution [6,11,27]. In addition, ex vivo manipulation of human cells is associated with extensive variations in phenotype, including Fas-dependent expression of adhesion molecules, which might impair hematopoietic cell navigation and homing [28]. Using a series of surrogate human cell assays, including SCID repopulation, long-term and semisolid cultures we show that UCB progenitors are largely insensitive to Fas-mediated apoptosis, and myeloid progenitor activity is functionally suppressed by dead cells. We hypothesized that resistance to apoptosis mediated by the TNF family receptors evolves from an imprinted molecular configuration of the progenitors. Since signaling mediated by TNF-R1 elicits dynamic changes in NFκB translocation and induction of protective genes [29 –31], we used Fas signaling to determine the intrinsic molecular configuration of hematopoietic progenitors. The transcriptional profiles of hematopoietic progenitors point to the possibility that resistance to apoptosis is actively sustained by the upregulation of antiapoptotic pathways.
Materials and Methods
Cell isolation, characterization, and staining
UCB was obtained from healthy donors pending informed consent according to the Institutional guidelines of the Beilinson Campus (IRB file 3656: 920050264). UCB samples harvested before placental delivery as previously described [24,25]. Mononuclear cells (MNCs) were washed twice, diluted twofold with the medium containing 0.5% human serum albumin and isolated over a Ficoll density gradient (1.077–1.080 g/mL) by centrifugation. The lineage−/low (lin−) subset was negatively isolated using the immunomagnetic Human Lineage Cell Depletion Kit, and CD34+ cells were positively isolated using the CD34 MicroBeads Kit by passing through magnetic cell separation column (Mini-MACS column type MS; Miltenyi Biotec, Bergisch Gladbach, Germany). The efficiency of the cell separation was reassessed by flow cytometry using labeled monoclonal antibodies.
Flow cytometry
Measurements were performed with Vantage SE-2 and Aria flow cytometers (Becton Dickinson, Franklin Lakes, NJ). CD34+ cells were identified using APC-labeled antibodies (BD Pharmingen, Erembodegem, Belgium), and lineage-positive subsets were assessed using PE-labeled antibodies: anti-human CD3 (BD Pharmingen), CD14, CD19, and CD33 (IQ Products, Groningen, The Netherlands) [25]. Expression of the Fas receptor in UCB cells was identified and neutralized using APC-labeled anti-human Fas (Miltenyi Biotec) and anti-human Fas (Millipore, Billerica, MA) antibodies, respectively. Cell death and apoptosis were determined in cells incubated with 5 μg/mL 7-aminoactinomycin-D (7-AAD; Sigma-Aldrich, St Louis, MO) and Annexin-V (IQ Products), respectively [24]. Proliferation was determined from a dilutional assay of the intracellular dye 5-(and-6-)-carboxyfluorescein diacetate succinimidyl ester (CFSE; Molecular Probes, Eugene, OR) [24]. Cell cycle distribution was analyzed with propidium iodide (PI) and Ki-67-FITC staining in cells fixed with 100% cold ethanol for 20 min on ice and permeabilized with 0.2% Tween-20 (Sigma-Aldrich) for 15 min at 37°C. Proliferation and fractional distribution of cells in the G0/G1, S, and G2/M phases were calculated using the ModFit software (Verity Software House, Topsham, ME). Engraftment of UCB cells in NOD.SCID xenochimeras was determined in the bone marrow using anti-mouse CD45 (eBioscience, San Diego, CA) and anti-human CD45 (IQ Products).
Apoptotic challenge
UCB cells were incubated (5×106 cells/mL) for variable times in the α-MEM culture medium (Biological Industries, Kibutz Beit Haemek, Israel) supplemented with StemPro Nutrient Supplement (Stem Cell Technologies, Vancouver, BC), 2 mM
Colony-forming unit assays
Colony-forming unit (CFU) assays were performed in 1.2% methylcellulose containing Iscove-modified Dulbecco medium (IMDM) by plating 2.5×103 MNC/well or 103 CD34+ and lin− cells/well [24,25]. Cultures were supplemented with 2 mM
Long-term culture initiating cells
UCB-derived mononuclear and lineage-negative cells were seeded on feeder layers of irradiated (3,000 rad) human mesenchymal stromal cells (MSCs) derived from UCB (PromoCell, Heidelberg, Germany) [24]. The cultures were refreshed every week by replacing half the Myelocult culture medium (Stem Cell Technologies) and supplementation of 50 ng/mL SuperFasL along control cultures from the same cord unit without supplementation of ligand. After 5 weeks, the cells were harvested, washed, and plated in methylcellulose.
Animal preparation and transplantation
Mice used in this study were NOD.CB17-Prkdcscid/J (NOD.SCID), purchased from Jackson Laboratories (Bar Harbor, ME), and housed in a barrier facility. All procedures were approved by the Institutional Animal Care Committee. NOD.SCID mice were conditioned with two daily doses of 25 μg/g Busulfan (Sigma-Aldrich), administered 2 days before cell transplantation into the lateral tail vein (1–2×107 viable cells in 0.2 mL PBS). Whole UCB cell preparations (2×107 cells) were preincubated with and without 50 ng/mL FasL (5×106 cells/mL) for 24 and 48 h. Viable MNCs were collected and ∼107 cells were grafted. The numbers of UCB cells in our collections were sufficient for transplant of two groups of two to five mice with cells from the same unit following incubation with FasL and in the medium serving as an internal control for each UCB unit.
Preparation of samples
CD34+ cells isolated from fresh UCB were challenged for apoptosis using 50 ng/mL SuperFasL for 48 h. Cells were washed and stained with Annexin-V (FITC), and anti-CD34+ (APC) and anti-FAS (PE) antibodies for sorting with a FACS Aria sorter into three different groups: (i) CD34+Fas+Annexin− viable receptor-positive cells (group 2), (ii) CD34+Fas+Annexin+ apoptotic receptor-positive cells (group 3), and (iii) CD34+Fas−Annexin− viable receptor-negative cells (group 4). A control subset consisted of CD34+Fas+Annexin− viable receptor-positive cells incubated for 48 h in the medium (group 1).
Isolation and analysis of transcriptomes
Total RNA was extracted and purified using the RNeasy Plus microkit (Qiagen, Germantown, MD) and processed in RNase-free test tubes containing RNA lysis buffer. RNA purity and RNA integrity number were determined with Bioanalyzer 2100 (Agilent Technologies, Santa Clara, CA) in seven independent extracts. Three hybridization samples were prepared by pooling together equal amounts of RNA obtained from three independent RNA extractions (out of seven) to reach the detection threshold of 88–1,029 pg/μL RNA corresponding to 1,046–8,673 cells. Reverse RNA transcription and amplification were performed using the WT-Ovation Pico RNA Amplification System kit (NuGEN Technologies, San Carlos, CA) and the SPRIPlate Ring Magnet Plate (Agencourt Bioscience, Danvers, MA). In each reaction, RNA input was 0.9–36 ng and cDNA product was 3–4 mg, which was quantified and assessed for quality using a NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies, Wilmington, DE). Amplified cDNA was processed with the WT Ovation Exon Module (NuGEN) to produce purified ST-cDNA; 3.5–6 μg ST-cDNA were fragmented and labeled using the FL-Ovation™ cDNA Biotin Module V2 (NuGEN). Samples were hybridized onto GeneChip Human Exon 1.0 ST arrays (Affymetrix, Santa Clara, CA). Network analysis was performed mapping the results on the IPA knowledge database (Ingenuity Pathway Analysis). Genes exceeding the detection threshold were analyzed for differences in expression in reference to viability and Fas expression at an arbitrary ratio of 2. These values were used to calculate fold changes in expression between the samples (ie, live/apoptotic cells, Fas+/Fas− progenitors), and percent variations in ratios between anti- and proapoptotic factors (ie, caspase-8/FLIP and Bax/Bcl-2 ratios).
Statistical analysis
Data are presented as means±standard deviations for each experimental protocol. Results in each experimental group were evaluated for reproducibility by linear regression of duplicate measurements. Differences between the experimental protocols were estimated with a post hoc Scheffe t-test, and significance was considered at P<0.05.
Results
Hematopoietic progenitors are insensitive to Fas-mediated apoptosis
The primary activity of the TNF superfamily receptors is transduction of apoptotic signals in mature immunohematopoietic cells, whereas relative insensitivity of progenitors to apoptosis has been attributed primarily to low levels of receptor expression [3,4]. Our analysis of 35 UCB units confirmed relatively low levels of Fas expression in gated CD34+ progenitors (∼10%), which is sustained during 72 h of incubation in the absence of chemokines and growth factors. The excessive susceptibility of CD34+ progenitors expressing Fas to spontaneous apoptosis in the culture suggests a possible causal relationship between this receptor and cell viability (Fig. 1A). However, exposure to the cognate ligand indicates this is not the case, as Fas cross-linking had no additional apoptotic activity in CD34+ progenitors. Since negative regulation through Fas is generally more accentuated in fast cycling cells [9,11,28], we assessed the proliferation rates and cell cycle phase. Fas-positive CD34+ progenitors cycled at slower rates than receptor-negative cells without significant impact of Fas ligation (Fig. 1B). The inverse relationship between death and proliferation in unstimulated UCB progenitors suggests that cell cycling reflects rather than affects viability. Approximately 10% of CD34+ and 50% of lineage-negative progenitors were positioned in the G0/G1 phase without detectable effects of Fas cross-linking (Fig. 1C). Overall, these data indicate that expression of Fas is associated with increased susceptibility to spontaneous apoptosis, but not to receptor-mediated apoptosis.
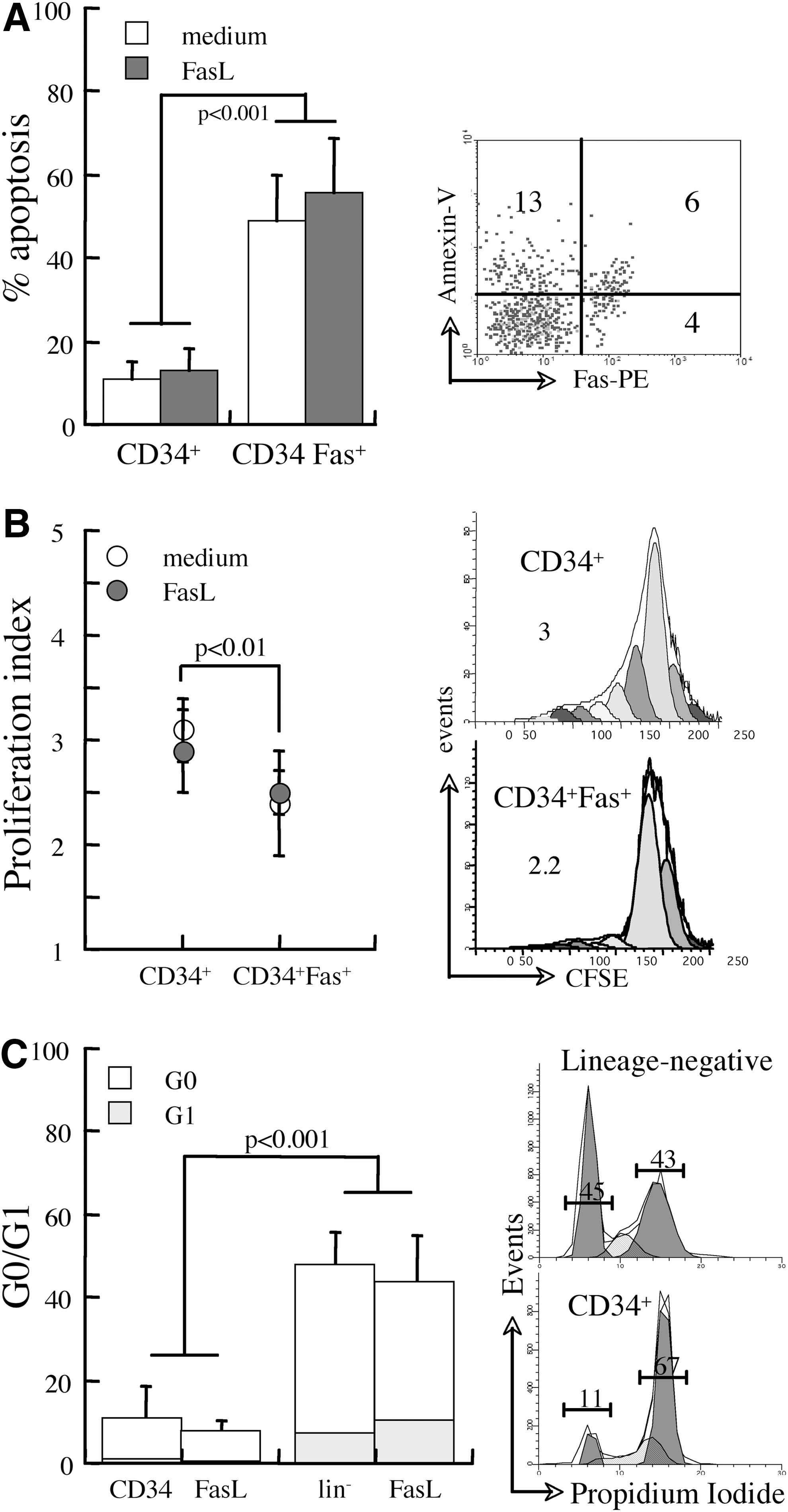
Sensitivity of umbilical cord blood (UCB) progenitors to receptor-mediated apoptosis in vitro.
Exposure to FasL does not impair quantitative human UCB cell engraftment
Human hematopoietic progenitor function is tested in surrogate assays for cells in various stages of commitment to differentiation. The most primitive subset represents the SCID repopulating cells (SRCs), which is considered to reflect the activity of human reconstituting cells [16]. The purpose of this experiment was to determine possible detrimental impact of FasL on SRC, by comparing engraftment of equal absolute numbers of progenitors at culture onset. To determine possible influences of Fas cross-linking on SRC function, UCB cells were incubated ex vivo with and without FasL for 24–48 h before transplantation into busulfan-conditioned NOD.SCID mice (Fig. 2A). Given the similar levels of fractional apoptosis, all mice received grafts of ∼107 viable MNCs. Incubation with FasL increased the fractions of CD34+ and lineage-negative progenitors (P<0.05), with concomitant decrease in B lymphocytes and myeloid cells (Fig. 2B). Despite large variability in values of engraftment in individual recipients, the mean levels of xenochimerism were unaffected by Fas cross-linking (Fig. 2C). Similar levels of engraftment are consistent with the lack of effect of FasL on cell cycle since SRC activity is confined to mitotically quiescent cells [16]. Furthermore, analysis of human lineages in the murine bone marrow after 12 weeks revealed that the exposure to FasL for 24 h increases the fractions of myelomonocytes along a reciprocal decline in lineage-negative progenitors (Fig. 2D). Thus, the activation of the Fas receptor bfore transplantation does not impair SRC engraftment and activity but augments myeloid differentiation of lineage-negative human progenitors in vivo similar to the outcome of TNF-R1 activation [24].
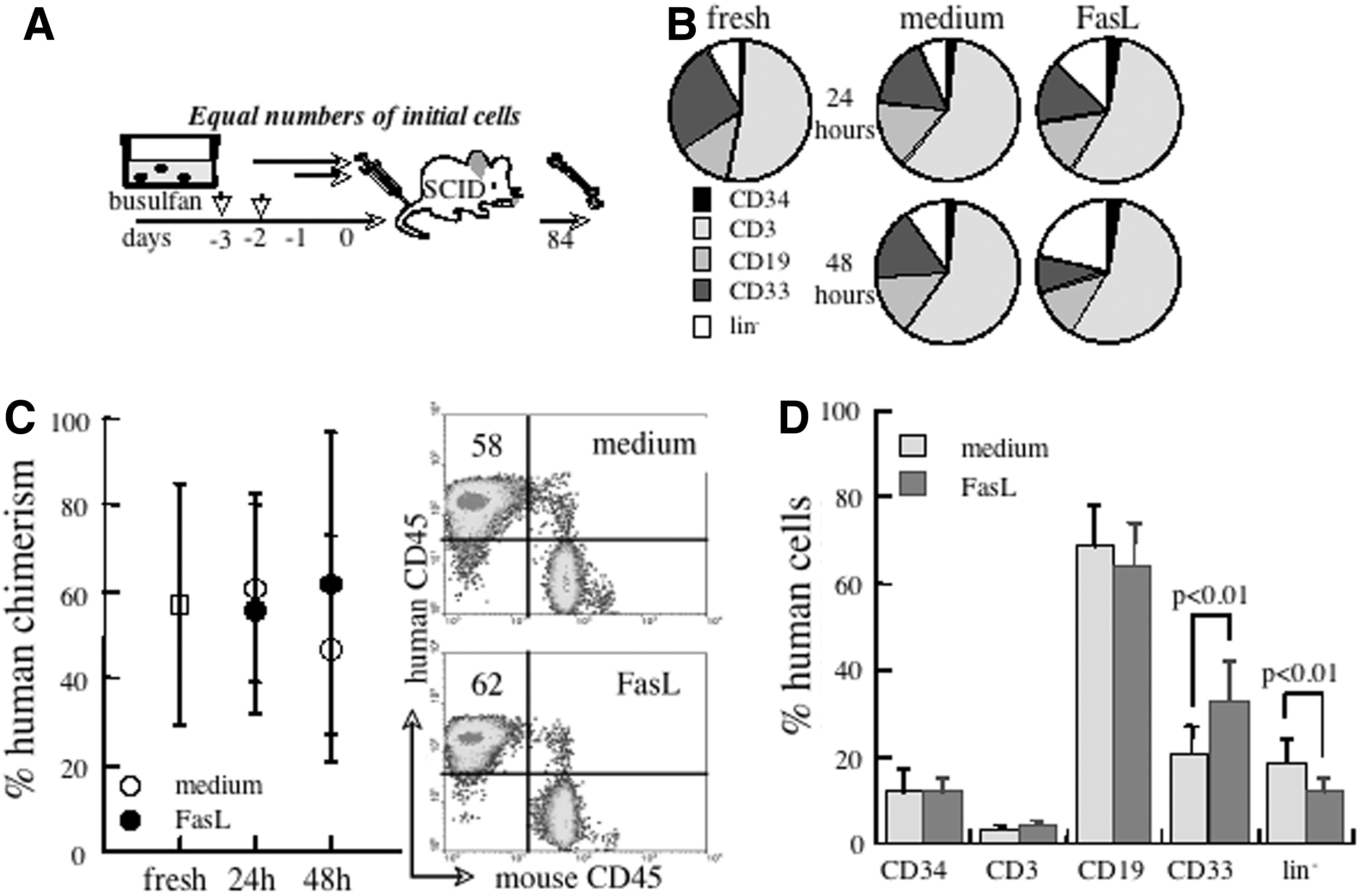
UCB progenitors with engraftment potential are resistant to receptor-mediated apoptosis.
The impact of FasL on human cells in clonogenic assays
In next stage, we assessed the impact of Fas cross-linking on the activity of progenitors in the culture. Long-term culture initiating cells (LTC-ICs) are considered to represent an intermediate subset of uncommitted precursors that differentiate with the support of MSC. Exposure of lineage-negative UCB progenitors to toxic concentrations of FasL (50 ng/mL) throughout the entire 5-week period of the culture over mesenchymal stroma did not attenuate the LTC-IC activity (Fig. 3A), emphasizing insensitivity of this subset of precursors to Fas cross-linking.
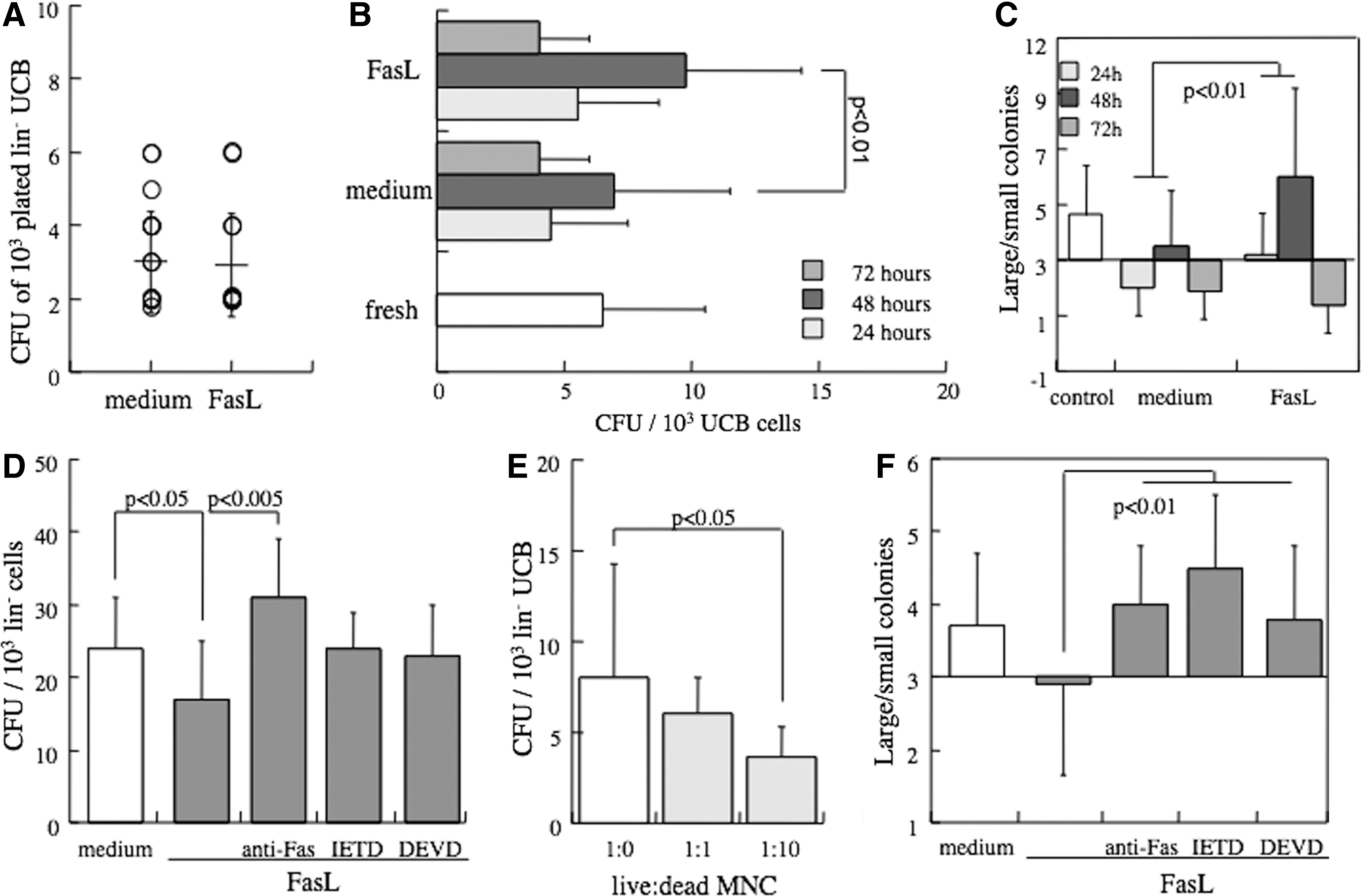
The influence of FasL on myeloid progenitor activity in the culture.
Myeloid progenitors developing in semisolid methylcellulose cultures are considered to represent more committed progenitors that respond to stimulation with SCF, IL-3, and GM-CSF [32]. UCB cells were assayed in two ways: transient exposure to FasL protein for variable periods of time before plating and continuous exposure to the ligand throughout the culture period. Transient exposure to FasL for 48 h resulted in increased number of colonies (Fig. 3B) as well as increased ratios of large to small colonies (Fig. 3C), consistent with an inductive effect of the ligands both on progenitor commitment to myeloid differentiation and development of the colonies. The decline in numbers of colonies and their size after 72 hours reflects the deterioration in hematopoietic progenitor activity during extended ex vivo culture, independent of the activation of the TNF family receptors [11,24].
Fas cross-linking throughout the entire culture period caused moderate suppression of the CFU activity, which was effectively reversed by neutralization of the receptor (Fig. 3D). Likewise, suppression of CFU activity was reversed by inhibition of both caspases-8 and −3, suggesting involvement of the apoptotic pathway in Fas-mediated inhibition of clonogenic activity. To evaluate whether decreased clonogenic activity is mediated by direct apoptosis induced by Fas in progenitors or by adverse effects related to Fas activity, cultures of lineage-negative progenitors were supplemented with dead cells. Apoptotic cells caused dose-dependent suppression of the clonogenic activity, pointing to functional inhibition of CFU activity rather than induction of Fas-mediated progenitor death (Fig. 3E). Therefore, myeloid progenitor activity is functionally suppressed by accumulation of apoptosis-sensitive cells in the culture, an indirect effect that is reversed by receptor neutralization and inhibition of the caspase cascade.
The Fas receptor serves as a negative regulator of expanding clones of differentiated hematopoietic lineages. This activity was visible in the size of the myeloid colonies, which was markedly suppressed by Fas cross-linking (Fig. 3F). As expected, repression was effectively reversed by receptor neutralization and inhibition of effector caspases. Considering the results of the three surrogate assays, our data emphasize gradual sensitization to Fas cross-linking along differentiation: SRC, LTC-IC, and committed myeloid progenitors are insensitive, whereas their differentiated progeny become progressively sensitive to apoptosis.
Molecular basis of progenitor insensitivity to Fas-mediated apoptosis
We reasoned that intrinsic insensitivity of hematopoietic progenitors to Fas receptor-mediated apoptosis is granted by a molecular configuration regulated at the transcriptional level. Isolated CD34+ progenitors were incubated for 48 h with toxic doses of FasL (50 ng/mL), and cells were subsequently sorted for analysis of the transcriptomes of live (group 2, Fas+Annexin−) and dead (group 3, Fas+Annexin+) subsets expressing the Fas receptor, and live cells lacking Fas expression (group 4, Fas−Annexin−) (Fig. 4A). A control group consisted of live progenitors expressing Fas (group 1, CD34+Fas+Annexin−) following incubation in the medium. Comparative full genomic analysis displays significant differences between viable and apoptotic CD34+Fas+ cells (Fig. 4B), including transcriptional upregulation of 10 genes and downregulation of 2 genes (Supplementary Table S1; Supplementary Data are available online at

Transcriptional profiles of apoptosis-resistant CD34+ progenitors expressing Fas.
Overexpression of mRNA encoding components of the extrinsic and intrinsic caspase-mediated cascades points to functionality of both apoptotic signaling pathways in human hematopoietic progenitors (Fig. 4C). This indicates that hematopoietic progenitors are not insensitive to receptor-mediated apoptosis due to the absence of receptor expression or absence of downstream signaling factors because cells sorted according to receptor expression possess intact apoptotic mechanisms. Active defense of these cells from apoptosis is evident from the twofold higher transcriptional levels of caspase-8 in viable cells compared to apoptotic cells (Supplementary Table S1). In variance, both physiological inhibitors FLIP and cytosolic inhibitor of apoptosis protein (cIAP) were transcriptionally upregulated in all CD34+Fas+ cells irrespective of sensitivity to apoptosis (Supplementary Table S2). Individual levels of expression were used to determine the FLIP/caspase-8 ratio, which was reduced by 20% in viable cells compared to apoptotic cells (P<0.05).
Proteomic analysis has suggested that increased Bcl-2 levels and consequently an increased Bcl-2/Bax ratio are significant in sustaining the viability of hematopoietic progenitors [5,33]. The transcriptomes showed increased Bcl-2 expression in all Fas+ progenitors (compared to Fas−) without significant difference between cells that succumbed to apoptosis (Supplementary Table S2). In variance, decreased Bax in live versus apoptotic Fas+ progenitors (Supplementary Table S1) resulted in 28% higher Bcl-2/Bax ratio in cells surviving the FasL challenge (P<0.05 vs. apoptotic cells). Thus, the Bcl-2/Bax ratio was increased by selective transcriptional downregulation of Bax at stable levels of Bcl-2 expression in CD34+ progenitors surviving Fas cross-linking. Bcl-2 inhibits and Bax promotes selective efflux from the mitochondria into the cytosol of cytochrome c, Smac/Diablo, and HtrA2/Omi, which facilitate the activation of caspases-3 and −9 [34,35]. Cells surviving Fas cross-linking displayed elevated transcription of HtrA2/Omi, which has apparently opposite functions: on the one hand proapoptotic activity is mediated by translocation to the cytosol where it binds and inactivates X-linked IAP [36], on the other hand, it has a physiological cytoprotective role within the mitochondria of viable cells [37]. Cytochrome-C was upregulated in all CD34+Fas+ cells at stable levels of Diablo, and the dominant cIAP isoform was upregulated in all CD34+Fas+ cells (Supplementary Table 2). These mitochondrial factors translocate to the cytosol to bind and inactivate inhibitory apoptosis proteins, thus removing the suppression of caspases-3 and −9. Although transcriptional analysis of these factors evaluates neither the activity of their protein products nor translocation and complex formation, transcriptional low levels of Bax and high levels of HtrA2/Omi in viable cells in the presence of high levels of Bcl-2 and cIAP in all CD34+Fas+ cells is a configuration compatible with the suppression of effector caspases.
The apparent configuration that confers resistance to Fas receptor-mediated apoptosis is a remarkable transcriptional upregulation of several factors that converge to activate the NFκB pathway (Supplementary Fig. S1B). All components of the complex formed by TGF-β-activated kinase-1 (TAK1), mucosa-associated lymphoid tissue lymphoma translocation gene-1 (MALT1), and TNF receptor-associated factor-6 (TRAF6) were upregulated in viable cells [38,39]. The canonical activation pathway of TAK1 was upregulated in all Fas-positive cells (Supplementary Table S2), including factors responsible for the direct activation of NFκB through disassembly of the TAK1-TRAF6-TAK1 binding protein-2 (TAB2) complex [40,41] mediated by ubiquitin-conjugating enzyme E2N (UBE2N) [42,43]. The alternative pathway included transcriptional upregulation of TRAF6 [38 –40] and double-stranded RNA-activated protein kinase (PKR) in viable cells, supported by abundance of NFκB-inducing kinase (NIK) in all Fas-positive cells [44]. Both pathways converge to the activation of the α-catalytic subunit of IκB kinase (IKKα) [42], which was markedly upregulated in cells that survived the apoptotic challenge. This enzyme is the common determinant of the NFκB activity by phosphorylation and detachment of inhibitory IκB and conversion of NFκB precursors P100 and P105 [45,46]. Overall, the transcriptome reveals robust tonic upregulation of the NFκB pathway accompanied by multiple factors responsible for the activation and facilitation of nuclear translocation, as a mechanism of active protection from apoptosis. This is apparently a distinct baseline configuration independent of phasic NFκB activation in response to stimuli [47] with a distinct transcriptional pattern compared to progenitors induced into myeloid and erythroid differentiation [48].
Discussion
As units of regeneration, grafted hematopoietic precursors operate under conditions of severe injury and inflammation mediated by radiochemotherapy. In search for the mechanisms of hematopoietic cell survival under such harsh conditions, we assessed the role of TNF family receptors, which have been traditionally attributed a detrimental effect on hematopoietic function. As a common executioner of apoptosis within the TNF superfamily, the Fas receptor is expressed in a minute fraction of the UCB cells; however, low levels of receptor expression are not the main mechanism of defense from apoptotic death. Engrafting progenitors largely fall in the Fas-positive fraction of murine and human hematopoietic cells [14,19], and this receptor is acutely upregulated under conditions of stress hematopoiesis and transplantation [9 –11,20,28]. In fact, the most primitive human and murine hematopoietic precursors ubiquitously upregulate several TNF family receptors under stress conditions, which largely serve for transduction of tropic signals [20,23 –25]. Therefore, despite low levels of Fas expression in fresh UCB-derived progenitors, our data are relevant to the significant fraction of progenitors that upregulate this receptor early after transplantation [10].
The fraction of CD34+ progenitors expressing Fas display excessive sensitivity to spontaneous apoptosis during extended ex vivo culture associated with progressive loss of engraftment potential [11]; however, this phenomenon is independent of Fas receptor cross-linking. Surrogate assays of human hematopoietic progenitor function demonstrate insensitivity to Fas-mediated apoptosis of SRCs and LTC-ICs. This phenomenon is partly attributed to stable proliferation rates and sustained G0/G1 cell cycle phase under conditions of Fas cross-linking since SRC activity is restricted to mitotically quiescent cells [16]. More committed myeloid progenitors, assessed in semisolid methylcellulose cultures, are functionally suppressed by factors released from apoptotic cells. As these surrogate assays evaluate hematopoietic progenitors positioned in different stages along the differentiation axis, it is evident that sensitivity to Fas-mediated apoptosis is acquired in late stages of differentiation, and the receptor becomes a negative regulator of expanding clones. Fas expression may represent an intrinsic default sensitization response to hematopoietic differentiation similar to its function as the common AICD effector in the immune system [1].
In variance from Fas, TNF-R1, and TRAIL-R2 in murine bone marrow [20,23,25], and TNF-R1 and TRAIL-R1 in human UCB cells [24,25], Fas has no trophic attributes in semisolid cultures of UCB progenitors. Our data infer that repression of the CFU activity by the Fas/FasL interaction is functional rather than apoptotic, an indirect mechanism caused by accumulation of factors released from apoptosis-sensitive cells. Therefore, Fas-mediated suppression of human UCB progenitor activity in vitro was effectively reversed by caspase inhibition, which prevented the accumulation of dead cells in the semisolid cultures. It is also evident that modulation of hematopoietic cell function requires Fas cross-linking [15] and is not a consequence of receptor neutralization by soluble FasL that prevents trimerization [12,13]. In contrast, short-term exposure to FasL before transplantation increases the fractions of lineage-negative progenitors and fosters myeloid differentiation in vivo: increased myeloid cells and reduced lineage-negative fractions in the bone marrow of xenochimeras. Induction occurred at both the level of SRC and more committed myeloid progenitors, as evident from increased CFU activity in semisolid cultures following transient exposure to the ligand.
Survival of hematopoietic progenitors is an inherent characteristic regulated at the transcriptional level. In variance from the contention that progenitors are protected from apoptosis by low levels of Fas expression [3,4,9,10] and upregulation of individual antiapoptotic factors [5 –7], we found that cells responsive to Fas-mediated signaling are resistant to apoptosis in the presence of competent caspase-mediated cascades of both extrinsic and intrinsic signaling pathways. This inherent resistance is conferred by a molecular configuration unfavorable to apoptosis that includes multiple regulatory mechanisms. Constitutive resistance of progenitors to apoptosis is demonstrated by the similar transcriptomes characteristic of CD34+Fas+ cells that survived incubation with and without exposure to the cognate ligand. Likewise, there were few differences between viable cells that were positive and negative for Fas, consistent with the observation that apoptosis in culture is dissociated from the activity of this receptor. Factors that facilitate apoptosis include upregulated transcription of the orphan TNF receptor DR6, a small heat shock protein HSP27 that protects cells from stress [49] and the adaptor molecules TNF-associated factor-3 (TRAF3) and protein (TTRAP) that inhibit the noncanonical NFκB pathway through inactivation of NIK [50]. The only upregulated antiapoptotic factor related to Fas expression was phospholipase-γ2, involved in PKC-mediated activation of the canonical NFκB pathway [51]. These variations are consistent with downsized and enhanced activities of the developmental and adaptive NFκB pathways, respectively, in hematopoietic progenitors expressing the Fas receptor.
The transcriptional profiles of viable CD34+Fas+ progenitors display upregulated expression of a series of factors involved in cell survival. The diagrammatic presentation of the transcriptomes according to the location of the protein products along the signaling pathways was used to facilitate interpretation. Constituents of the NFκB pathway have been demonstrated to be active in CD34+ progenitors at the protein level [52] for protection from cytokine-mediated injury signals [8] and receptor-activated apoptosis [53] and for induction of clonogenic activity [48]. Transcriptional upregulation includes both developmental (noncanonical) NIK-mediated and adaptive (canonical) Tak1-mediated NFκB activation pathways, with a more pronounced adaptive component [46]. Nuclear translocation of NFκB is allowed by IKKα-mediated degradation of the IκBα binding protein [42], an enzyme that is the downstream target of both developmental and adaptive NFκB pathways [54,55]. IKKα was markedly upregulated in cells surviving the apoptotic challenge. It is also evident from the diagram that the two complementary NFκB activation mechanisms are redundant and might synergize under the particular molecular configuration of viable hematopoietic progenitors. Characterization of the transcriptomes in this study is consistent with the predominant role of the NFκB pathway in progenitor differentiation into erythroid and various myeloid lineages [8,52]. Consistently, we demonstrate that transient ex vivo Fas cross-linking fostered myeloid reconstitution in vivo and in semisolid cultures.
The contention that hematopoietic progenitors are actively protected rather than insensitive to apoptotic signals is based on the well-developed extrinsic apoptotic pathway related to the Fas receptor. Caspase-8 is a dominant activator [56] submitted to tight regulation under both extrinsic and intrinsic signaling pathways. Although high levels of FLIP protein expression in CD34+ hematopoietic progenitors [7] is consistent with our transcriptional analysis of CD34+Fas+ cells, the FLIP/caspase-8 mRNA ratio is remarkably low in viable cells. Likewise, transcriptional profiling confirms prior protein analysis showing elevated Bcl-2 and IAP [5,6,33], although the high Bcl-2/Bax mRNA levels were primarily caused by Bax silencing. Both apoptosis suppressors Bcl-2 and cIAP were equally transcribed at high levels in Fas-sensitive and Fas-insensitive hematopoietic progenitors, thus they are unlikely key factors responsible for cell survival. Furthermore, Bcl-2 and cIAP as well as FLIP and TRAF1/2 are target genes of the NFκB transcription factor [8,57] and may be induced by its nuclear translocation. We conclude from these data that resistance to apoptosis is not conferred by a singular factor, but by a complex molecular configuration that protects progenitors from death through redundant mechanisms that are likely required to preserve viability under various and dynamic environments. For example, transcriptional upregulation included all constituents of complexes formed by caspase-8, MALT1, TAK1, TRAF6, and TTRAP, which synchronize the apoptotic caspase cascade to restrictive and stimulatory functions of NFκB.
The molecular configuration revealed in this study attributes to the NFκB pathway, an important role in resistance to apoptosis in response to Fas cross-linking. Since Fas is a less effective activator of the NFκB pathway than TNF-R1 [29 –31], we consider the described transcriptomes to reflect an inherent pattern of hematopoietic progenitors that includes the low fractions of more primitive hematopoietic stem cells. Extension of these data to stem cells is supported by prior murine studies, where stress-induced upregulation of the receptor and ligand were found in most primitive hematopoietic precursors and fractions enriched in multipotent stem cells [20,21]. Despite differences in apoptotic signaling triggered by the Fas and TNF-R1 receptors [58], the complex mechanisms of defense have the capacity to protect from apoptosis signaled by multiple TNF family receptors.
Footnotes
Acknowledgments
This work was funded by grants from the Frankel Trust for Experimental Bone Marrow Transplantation. We thank Mrs. Ela Zuzovsky and Mrs. Ana Zemliansky for their outstanding technical support.
Author Disclosure Statement
The authors have no conflict of interest to declare.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.