
Abstract
Embryonic stem (ES) cells are useful for elucidating the molecular mechanisms of cell fate decision in the early development of mammals. It has been shown that aggregate culture of ES cells efficiently induces neuroectoderm differentiation. However, the molecular mechanism that leads to selective neural differentiation in aggregate culture is not fully understood. Here, we demonstrate that the oxygen-sensitive hypoxia-inducible transcription factor, Hif-1α, is an essential regulator for neural commitment of ES cells. We found that a hypoxic environment is spontaneously established in differentiating ES cell aggregates within 3 days, and that this time window coincides with Hif-1α activation. In ES cells in adherent culture under hypoxic conditions, Hif-1α activation was correlated with significantly greater expression of neural progenitor-specific gene Sox1 compared with ES cells in adherent culture under normoxic conditions. In contrast, Hif-1α-depleted ES cell aggregates showed severe reduction in Sox1 expression and maintained high expression of undifferentiated ES cell marker genes and epiblast marker gene Fgf5 on day 4. Notably, chromatin immune precipitation assay and luciferase assay showed that Hif-1α might directly activate Sox1 expression. Of additional importance is our finding that attenuation of Hif-1α resulted in an increase of BMP4, a potent inhibitor of neural differentiation, and led to a high level of phosphorylated Smad1. Thus, our results indicate that Hif-1α acts as a positive regulator of neural commitment by promoting the transition of ES cell differentiation from the epiblast into the neuroectoderm state via direct activation of Sox1 expression and suppressing endogenous BMP signaling.
Introduction
E
Previous studies have explored methods for producing neural cells from mouse ES cells in vitro [4 –9]. Initially, differentiation of mouse ES cells into neuroectoderm was achieved through formation of an embryoid body in the presence of retinoic acid [4] or through coculture with a particular stromal cell line, PA6 [5]. Ying et al. successfully generated neuroectoderm using adherent monoculture of mouse ES cells in serum-free medium supplemented with N2B27 [9]. This adherent culture method leads to highly efficient neural conversion via an autocrine induction mechanism in which fibroblast growth factor (FGF) signaling plays a pivotal role [9 –11].
The simplest way to recreate neural differentiation in vitro, proposed by Sasai et al., is aggregate culture of mouse and human ES cells in growth-factor-minimized medium [serum-free culture of embryoid-body-like aggregates (SFEBq)] [12 –14]. In SFEBq culture, efficient neural conversion occurs in a tissue-autonomous manner without the induction of mesodermal or endodermal differentiation and without any extrinsic inducers [12 –14]. This neural differentiation is quite different from mesodermal or definitive endodermal commitment of ES cells, in which several extrinsic signals, such as Activin/Nodal, BMP, and Wnt/β-catenin, play critical roles [15 –17].
In amphibian development, acquisition of a neuroectodermal cell identity occurs via a cell-intrinsic mechanism that requires only the absence of signals that inhibit neural differentiation [18 –21]. For example, BMP4 signals can suppress gene expression required for neuroectodermal differentiation. Correspondingly, addition of recombinant BMP4 leads to inhibition of neural differentiation of SFEBq-cultured mouse ES cells [12,22]. Intriguingly, it has also been reported that long-term SFEBq-cultured ES cells recapitulate the process of early brain development [14]. These findings strongly suggest that the microenvironment formed in SFEBq culture is optimum for the commitment of neuroectoderm and similar to that of differentiating neural tissues in vivo. However, the molecular mechanism that leads efficient neural conversion in SFEBq culture remains obscure.
Oxygen concentration is one of the important environmental factors in early development. Early embryogenesis must proceed in a low-oxygen environment (∼3% oxygen) until the vascular system is developed [23 –26]. Oxygen tension is also a critical factor for neural stem cell niche in the central nervous system (CNS), regulating the survival and differentiation of these cells [27 –29]. Hypoxia-inducible factor 1 alpha (Hif-1α) is one important transcription factor induced by low oxygen [30]. It has been reported that Hif-1α-deficient mice show retarded development and failure of neural tube closure [31]. Further, brain-specific Hif-1α-knockout mice show a reduction of neural cells [32]. These findings indicate that Hif-1α is one of the key molecules that control early neural development in mammals.
In this study, we used the SFEBq culture method as an in vitro neural development system, Hif-1α-knockdown ES cells, chromatin immunoprecipitation (ChIP) assay, luciferase assay, and forced expression of Hif-1α to investigate whether Hif-1α acts to regulate neural commitment of mouse ES cells. In SFEBq culture, a hypoxic environment is spontaneously established in ES cell aggregates. Additionally, Hif-1α-knockdown ES cells do not undergo neural conversion but tend to pause at the epiblast stage. ChIP assay, luciferase assay, and forced expression of Hif-1α indicated that Hif-1α specifically binds to the loci of the earliest neuroectodermal marker gene Sox1 and regulates its expression. Notably, in Hif-1α-knockdown cells in SFEBq culture, endogenous expression of BMP4 and the amount of phosphorylated Smad1/5/8 are significantly elevated compared with control.
Our findings indicate a dual role of Hif-1α in regulating neural differentiation; Hif-1α suppresses endogenous BMP signaling during neural commitment and promotes the transition of ES cell differentiation from the epiblast state into the neural progenitor state via the direct activation of Sox1 neural progenitor gene expression.
Materials and Methods
ES cell culture
Mouse ES cells (EB5, a kind gift from Dr. Niwa) and Sox1-GFP ES cells (46C, a kind gift from Dr. Smith) were maintained as described in previous reports [5,12,13] in the presence of 2,000 U/mL leukemia inhibitory factor (LIF). Differentiation medium was prepared as follows: GMEM was supplemented with 10% knockout serum replacement (KSR; Invitrogen, Carlsbad, CA), 2 mM glutamine, 1 mM pyruvate, 0.1 mM nonessential amino acids, and 0.1 mM 2-mercaptoethanol (2-ME). For floating culture using SFEBq method [14], ES cells were dissociated to single cells by treatment with 0.25% trypsin-EDTA and seeded in differentiation medium using 96-well low-cell-adhesion plates (3×103 cells/100 μL per well). For adherent culture, dissociated ES cells were plated on 35-mm tissue culture dishes coated with 0.1% gelatin at a density of 1×105 cells. The day on which ES cells were seeded for differentiation was defined as differentiation day 0. For hypoxic exposure, ES cells were subjected to 3% oxygen tension balanced with nitrogen gas in an isolated incubator (ASTEC, Tokyo, Japan).
Western blotting
For nuclear extract preparation, ES cells were lysed in buffer A [10 mM HEPES, (pH 7.9), 10 mM KCl, 0.1 mM EDTA, 1 mM DTT, and protease inhibitor mixture (Nacalai Tesque, Kyoto, Japan)] on ice for 10 min, followed by addition of 0.5% Nonidet P-40 and centrifugation at 15,000 rpm for 2 min at 4°C. The pellet was suspended in buffer C [20 mM HEPES (pH 7.9), 500 mM NaCl, 1 mM EDTA, and 1 mM dithiothereitol], kept on ice for 15 min, and then centrifuged at 15,000 rpm for 10 min. The supernatant was used as the nuclear extract sample for the western blot. For whole-cell protein lysate preparation, cells were lysed in RIPA buffer [50 mM Tris-HCl (pH 7.6), 150 mM NaCl, 1% Nonidet P-40, 0.5% sodium deoxycholate, 0.1% sodium dodecyl sulfate (SDS), and protease inhibitor mix] on ice for 15 min. After centrifugation at 15,000 rpm for 10 min, the supernatant was analyzed by western blot. Twenty milligram samples of nuclear extract or whole-cell lysate were fractionated by 4%–10% gradient SDS-polyacrylamide electrophoresis and transferred to PVDF membrane (Merck Millipore, Darmstadt, Germany). The membranes were incubated overnight at 4°C with primary antibodies including anti-Hif-1α mouse monoclonal (1:1,000; Abcam, Cambridge, United Kingdom), anti-Smad1/5/8 rabbit polyclonal (1:2,000; Cell Signaling Technology, Danvers, MA), anti-pSmad1/5/8 rabbit polyclonal (1:2,000; Cell Signaling Technology), and anti-pErk1/2 rabbit polyclonal (1:2,000; Cell Signaling Technology). The blots were washed with TBS-T several times and then incubated for 1 h at room temperature with horseradish peroxidase-conjugated secondary antibody (anti-mouse IgG antibody at a dilution of 1:10,000; anti-rabbit IgG antibody at a dilution of 1:10,000; Santa Cruz Biotechnology, Santa Cruz, CA). Immunoreactivity was detected with chemiluminescence reagent (Merck Millipore). After developing, blots were reproved with goat anti-Lamin B (1:3,000; Santa Cruz Biotechnology) or goat anti-Actin (1:3,000; Santa Cruz Biotechnology) to confirm that samples contained equal amount of protein.
Immunostaining
ES cell aggregates were fixed with 4% paraformaldehyde/PBS for 30 min at room temperature and cryoprotected in 15% sucrose for overnight at 4°C. Samples were then embedded in OCT compound (Sakura Finetek Japan, Tokyo, Japan) and cryosectioned (10 μm). Immunohistochemistry was performed as described in previous reports [12,13] using the following primary antibodies: anti-Sox1 rabbit polyclonal (Stemgent, Cambridge, MA) at 1:200, anti-Nanog rabbit polyclonal (ReproCell, Kanagawa, Japan) at 1:1,000, and anti-Oct3 mouse monoclonal (BD Transduction Laboratories, Franklin Lakes, NJ) at 1:200. Alexa-488 or -546 conjugated goat anti-mouse or -rabbit IgG (Invitrogen) was used as secondary antibody. Sections were mounted with SlowFade Gold antifade reagent (Molecular Probe, Carlsbad, CA) and photographed with the Olympus BX51 fluorescence microscope (Olympus, Tokyo, Japan).
Detection of hypoxic cells in ES cell aggregates
To detect the hypoxic cells in ES cell aggregates, Hypoxyprobe-1 kit (Hypoxyprobe, Inc., Burlington, MA) was used according to the manufacturer's protocol. In brief, ES cell aggregates were treated with 60 μg/mL of Hypoxyprobe-1 (pimonidazole hydrochloride) for an hour before collecting and then fixed and cryosectioned as described before. Mouse monoclonal anti-Hypoxyprobe-1 (MAb-1; 1:500) and Alexa-546 conjugated goat anti-mouse IgG (1:1,000) were used for the detection of Hypoxyprobe-1 in hypoxic cells.
Reverse transcription-polymerase chain reaction (RT-PCR) analysis
Total RNA was purified from ES cells or ES-cell-derived cell aggregates using RNeasy Micro kit (Qiagen, Hilden, Germany), and first-strand cDNA was synthesized using Super Script II (Life Technologies, Carlsbad, CA). Semiquantitative RT-PCR was performed using EX-Taq DNA polymerase (Takara Bio, Inc., Shiga, Japan) with 20–28 cycles of denaturation (94°C, 10 s), primer annealing (60°C, 30 s), and extension (72°C, 30 s). Quantitative RT-PCR (qPCR) was performed using THUNDERBIRD SYBR qPCR Mix (Toyobo, Osaka, Japan) and 7500 Fast Real Time PCR System (Applied Biosystems, Carlsbad, CA) according to the manufacturer's instructions. The data were normalized for GAPDH expression. The primers used in this study are shown in Table 1.
RT-PCR, semiquantitative RT-PCR; qPCR, quantitative RT-PCR.
Knockdown experiment of Hif-1α with short hairpin RNA
The knockdown study of Hif-1α in mouse ES cells was performed by introducing short hairpin RNA (shRNA). We used transduction-ready lentiviral particles that contain three target-specific constructs that encode shRNA designed to knock down mouse Hif-1α expression (Santa Cruz Biotechnology). Briefly, 1×104 lentiviral-transducing particles were applied to 80% confluent ES cells (24-well dish) with 5 μg/mL Polybrene (Santa Cruz Biotechnology), and stable cell lines were selected by treatment with 5 μg/mL puromycin. ES cell colonies were selected under a microscope, expanded, and screened by Hif-1α RNA and Hif-1α protein expression. Lentiviral particles that contain shRNA construct that encodes scrambled sequence were used as a control (Santa Cruz Biotechnology).
Chromatin immunoprecipitation
ChIP assay of SFEBq-cultured mouse ES cells on day 3.5 was carried out with ChIP-IT Express Enzymatic kit (Active Motif, Carlsbad, CA) according to the manufacturer's protocol. Protein-G magnetic beads and antibodies against mouse Hif-1α or RNA polymerase II were used for immunoprecipitation reaction. IgG was used as negative control. Specific PCR primers for ChIP assays were as follows: Sox1 (forward: 5′-AGAGTAAATTGGCACCTGAC-3′, reverse: 5′-GCAAAGGCCTTTACAGATTC-3′) and Zfp521 (forward: 5′-CTTGGTTCATTCCATGACTTGT-3′, reverse: 5′-AGAATTTCTGGGTAATGCTGGT-3′).
Transfection of Hif-1α
EF1α promoter is known to exhibit high expression activity in undifferentiated or differentiated mouse ES cells [33]. A full-length coding fragment for Hif-1α was cloned into a pEF1α-IRES-AcGFP1 vector (Clontech, Palo Alto, CA) and used for forced expression. Recombinant pEF1α-Hif1α (pEF1α-Hif1α-IRES-AcGFP1) plasmids were purified using a plasmid purification kit (Qiagen) and sequences were confirmed. Transfection of recombinant plasmids was performed using Xfect Stem (Clontech) according to the manufacturer's protocol. After 48 h of transfection, green fluorescent protein (GFP)–positive mouse ES cells were sorted by fluorescence-activated cell sorting (FACS) and further analyzed for qPCR.
Luciferase assay
To construct luciferase reporter plasmids, genomic DNA fragments of mouse Sox1 gene 5′-flanking region, spanning from −3575 to −3222 bp (Chromosome8 12391934–12392297), were amplified by PCR using KOD-plus Neo (Toyobo, Osaka, Japan) and ligated to pGL3-promoter vectors (Promega, Madison, WI). Point mutations were introduced into presumptive hypoxia responsible element (HRE) sites using KOD-Plus Mutagenesis kit (Toyobo) according to the manufacturer's instructions. pGL3-Sox1-mutN is equal to pGL3-Sox1 except that 5′ HRE site CGACGT (−3562/−3557) was mutated to CGAAAT. In pGL3-Sox1-mutC, the 3′ HRE site ACGT (−3240/−3237) was mutated to AAAT. In pGL3-Sox1-mutNC, both HRE sites were mutated. pEF1α-Hif1α-deleted plasmid is equal to pEF1α-Hif1α except that carboxyl-terminal domain (797–836 amino acids, necessary for the association with p300/CBP) was deleted. All constructs were purified using a plasmid purification kit (Qiagen) and sequences were confirmed. For the luciferase assay, HEK293T cells were seeded in 24-well plate, and cotransfected with 250 ng Hif-1α overexpression plasmid (pEF1α-Hif1α or pEF1α-Hif1α deleted), 250 ng Sox1 luciferase reporter plasmids, and constitutively expressed Renilla luciferase construct (100 pg; Promega) using Xfect reagent (Clontech). After 48 h, luciferase activity was determined using Dual-Luciferase Reporter Assay System (Promega).
FACS analysis
Single cells were obtained by treatment with 0.25% trypsin-EDTA. Cells were analyzed for GFP expression following transfection with plasmids that carry GFP gene. For FACS analysis, MoFlo™ XDP (Beckman Coulter, Inc., Brea, CA) was used.
Statistical analysis
For statistics, three experiments were performed. P values for statistical significance (t-test) are described in the figure legends. Experimental data are expressed as the mean±SD.
Results
Hypoxic response in floating ES cell aggregates at critical time period of neural commitment
In SFEBq culture, dissociated mouse ES cells efficiently formed cell aggregates and undergo selective neural differentiation in a tissue-autonomous manner (Fig. 1A) [12
–14]. In this process, epiblast marker gene Fgf5 was expressed in differentiation days 2–3 (Supplementary Fig. S1A; Supplementary Data are available online at
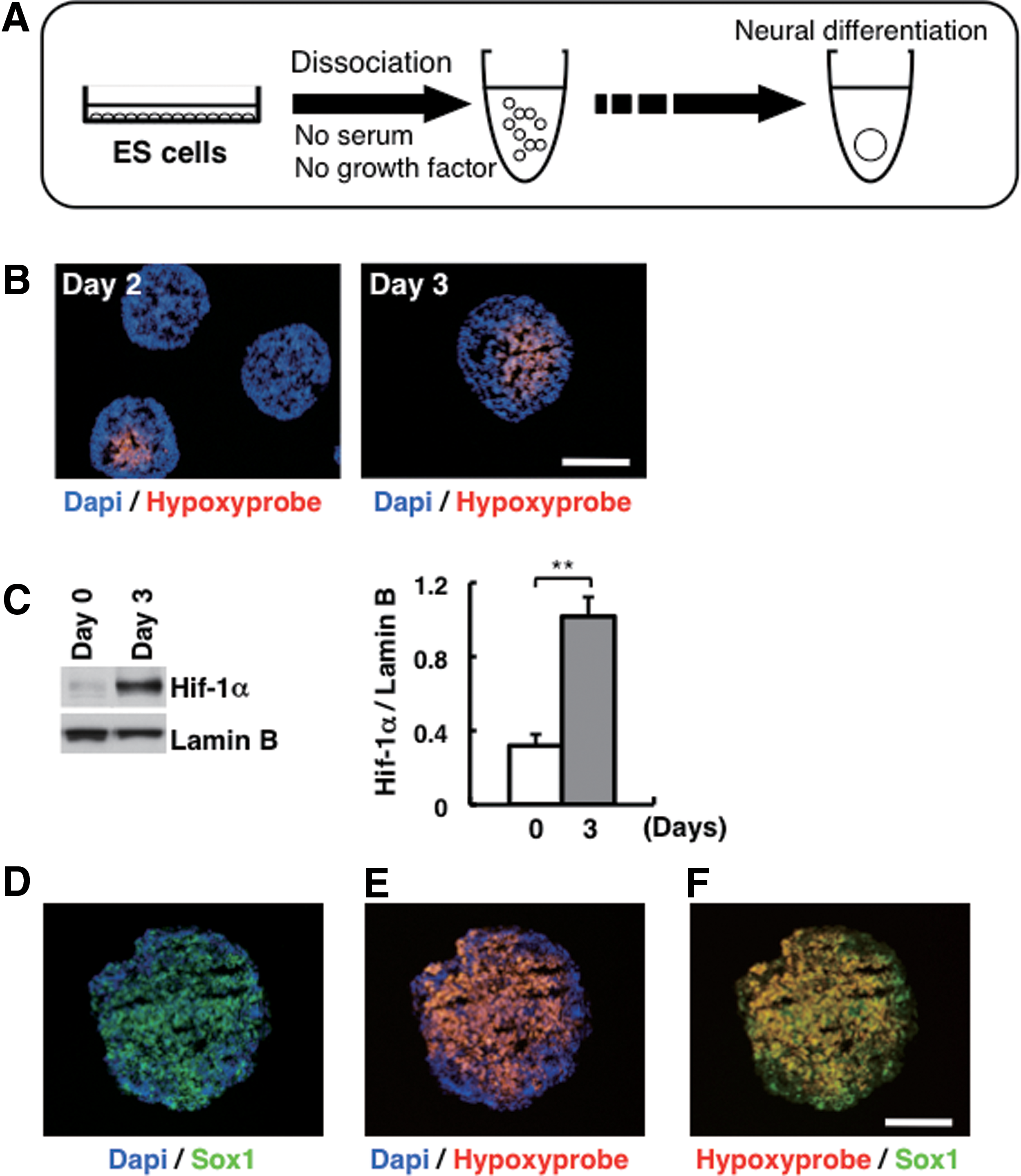
Hypoxic response in ES cell aggregates during neuroectoderm differentiation.
Improved differentiation of neural progenitor cells in hypoxia
To investigate how neural differentiation is regulated in SFEBq culture, we first compared the neuroectoderm differentiation between ES cells in SFEBq culture and in adherent culture under normoxic conditions (Fig. 2A). ES cells in adherent culture showed extremely low expression of Sox1 compared with ES cells in SFEBq culture under normoxic conditions (Fig. 2B). On the other hand, undifferentiated ES cell markers, such as Nanog and Oct3/4, were highly expressed in ES cells in adherent culture (Fig. 2B). In addition, expression of Hif-1α was not observed in ES cells in adherent culture under normoxic conditions, whereas ES cells in SFEBq culture showed high Hif-1α expression (Fig. 2C).

Improvement of neuroectoderm differentiation of mouse ES cells in hypoxia.
Next, in order to investigate the effect of low oxygen on neuroectoderm differentiation, ES cells in adherent culture were cultured under normoxic or hypoxic condition and analyzed (Fig. 2D). Adherent culture of ES cells under hypoxic conditions resulted in significant increase of Sox1 expression and reduction of Nanog and Oct3/4 expression as compared with adherent culture of ES cells under normoxic conditions (Fig. 2E). In addition, adherent culture of ES cells under hypoxic conditions produced significantly greater expression of Hif-1α (Fig. 2F). We also assessed the number of Sox1-GFP-positive cells in adherent and SFEBq cultures of ES cells with GFP knocked-in at the Sox1 locus (46C, gift from A. Smith) [38]. The number of Sox1-GFP-positive cells significantly increased when ES cells were cultured under hypoxic conditions (data not shown), as also reported by other research groups using same Sox1-GFP cell line [39,40].
These findings suggest that hypoxic stimuli promote neuroectoderm differentiation of mouse ES cells and that Hif-1α may be a key factor of the process.
Requirement of Hif-1α in neuroectoderm differentiation
We next examined whether Hif-1α is necessary for the neural differentiation using ablation of endogenous Hif-1α by shRNA. Introduction of Hif-1α-shRNA into ES cells successfully suppressed Hif-1α expression at mRNA and protein levels compared with the control (Fig. 3A, B). Expression of undifferentiated ES cell markers was not significantly different in Hif-1α-knockdown ES cells and control (Fig. 3A).
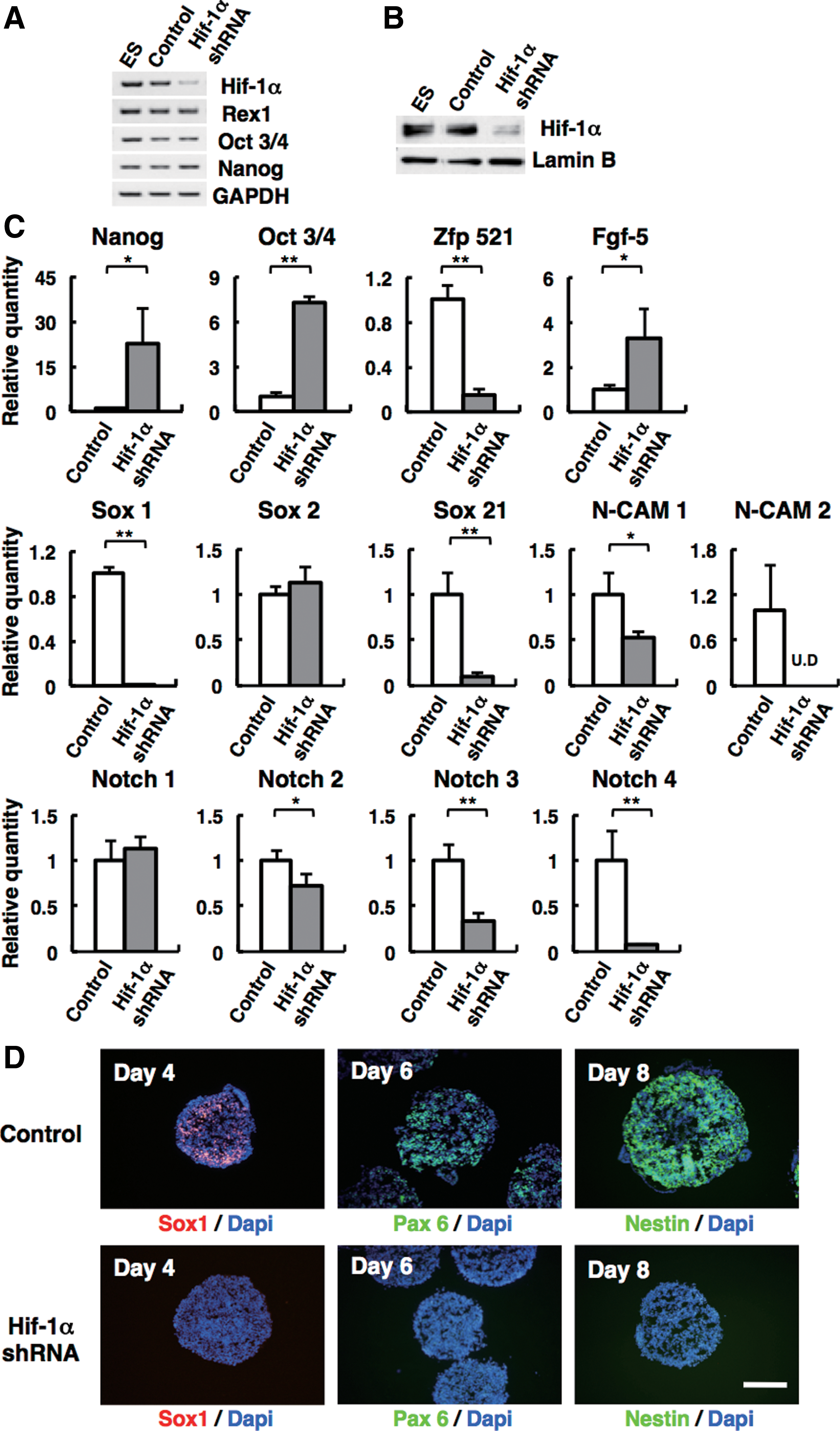
Hif-1α is required for the commitment of mouse ES cells to neuroectodermal cells.
As shown in Figure 3C, Hif-1α-knockdown ES cells in SFEBq culture manifested severely reduced expression of early neuroectoderm genes, such as Sox1 (less than 1% of control) and Zfp521 (∼15.4% of control). Other neuroectodermal marker genes, such as Ncam (Ncam1: ∼52.4% of control and Ncam2: undetected), Sox21 (∼9.4% of control), and Notch (Notch2 ∼73.1%, Notch3 ∼73.1%, and Notch4 ∼73.1%), were also downregulated in Hif-1α-knockdown ES cell aggregates (Fig. 3C). There was no significant difference on the expression of Sox2 and Notch1. At the same time, these knockdown ES cells showed high expression of undifferentiated ES cell markers, including Nanog and Oct3/4, compared with control on day 4. Immunohistochemistry showed that Hif-1α-knockdown ES cells had a markedly reduced number of Sox1-positive cells in SFEBq culture (0%, n=59; Fig. 3D, bottom), while control shRNA had no effects on Sox1 expression on day 4 (93%, n=44; Fig. 3D, top). In Hif-1α-knockdown ES cell aggregates, the cells that express neural progenitor markers, such as Pax6 and Nestin, were undetected on days 6 and 8 (0%, n=24; Fig. 3D).
Interestingly, the expression of epiblast-marker gene Fgf5 [41,42] did not decrease in Hif-1α-knockdown ES cells in SFEBq culture on day 4, whereas most of the cells treated with control shRNA showed reduced Fgf5 expression (Fig. 3C and Supplementary Fig. 1A).
Taken together, our Hif-1α-knockdown data show that Hif-1α is an indispensable molecule for the neuroectoderm differentiation of mouse ES cells; with Hif-1α knocked down, ES cells are capable of differentiating into the epiblast-like state, but do not proceed to the neural progenitor state.
Direct regulation of neural progenitor gene expression by Hif-1α
We next investigated how forced expression of Hif-1α affects downstream of Hif-1α target genes. After 48 h of transfection with pEF1α-Hif1α, GFP-positive ES cells were observed under fluorescein microscopy (Fig. 4A), and Hif-1α expression was confirmed by western blotting (Fig. 4B). GFP-positive cells were sorted by FACS (Fig. 4C) and gene expression was analyzed by qPCR.
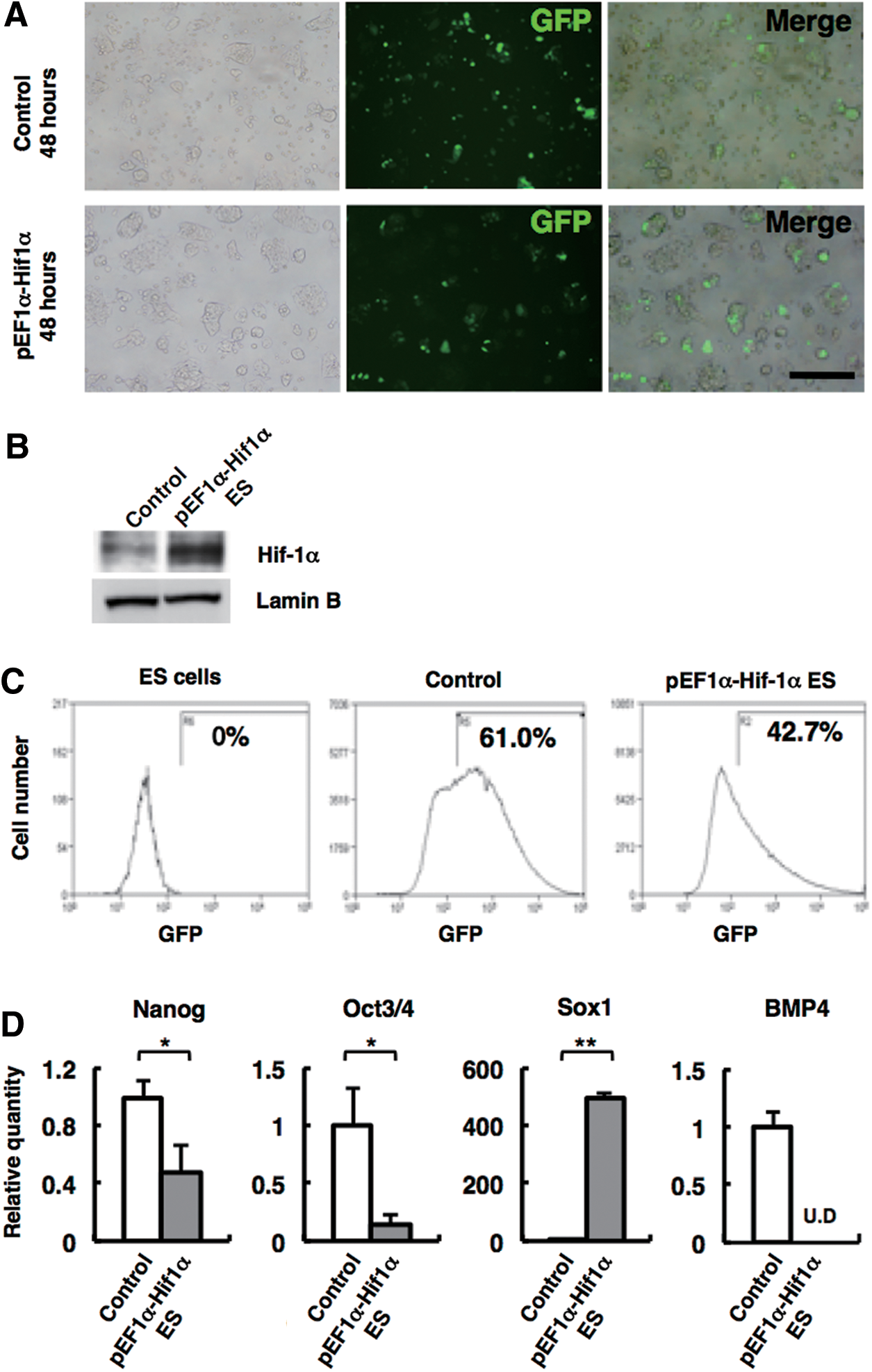
Hif-1α promotes neural progenitor gene expression.
As shown in Figure 4D, the forced expression of Hif-1α in ES cells resulted in high expression of Sox1 and reduced expression of undifferentiated markers, such as Oct3/4 and Nanog, compared with control. Bmp4 expression was undetectable in ES cells with Hif-1α overexpressed (Fig. 4D). Since, in general, Hif-1α acts as transcriptional activator [30], we hypothesized that Sox1 may be a direct target of Hif-1α. To verify this hypothesis, we performed ChIP assay using chromatin derived from SFEBq-cultured ES cells on day 3.5. As shown in Figure 5A, Hif-1α protein was specifically associated with the locus of Sox1, a neuroectodermal gene, but not with another early neuroectodermal gene locus Zfp521 [22] at the time of neural specification. The predicted Hif-1α binding site, commonly referred to as the HRE, is present in the regulatory region of Sox1 gene as described in Figure 5B. Consequently, there is a high possibility that Hif-1α directly and specifically activates Sox1 gene during neural commitment.
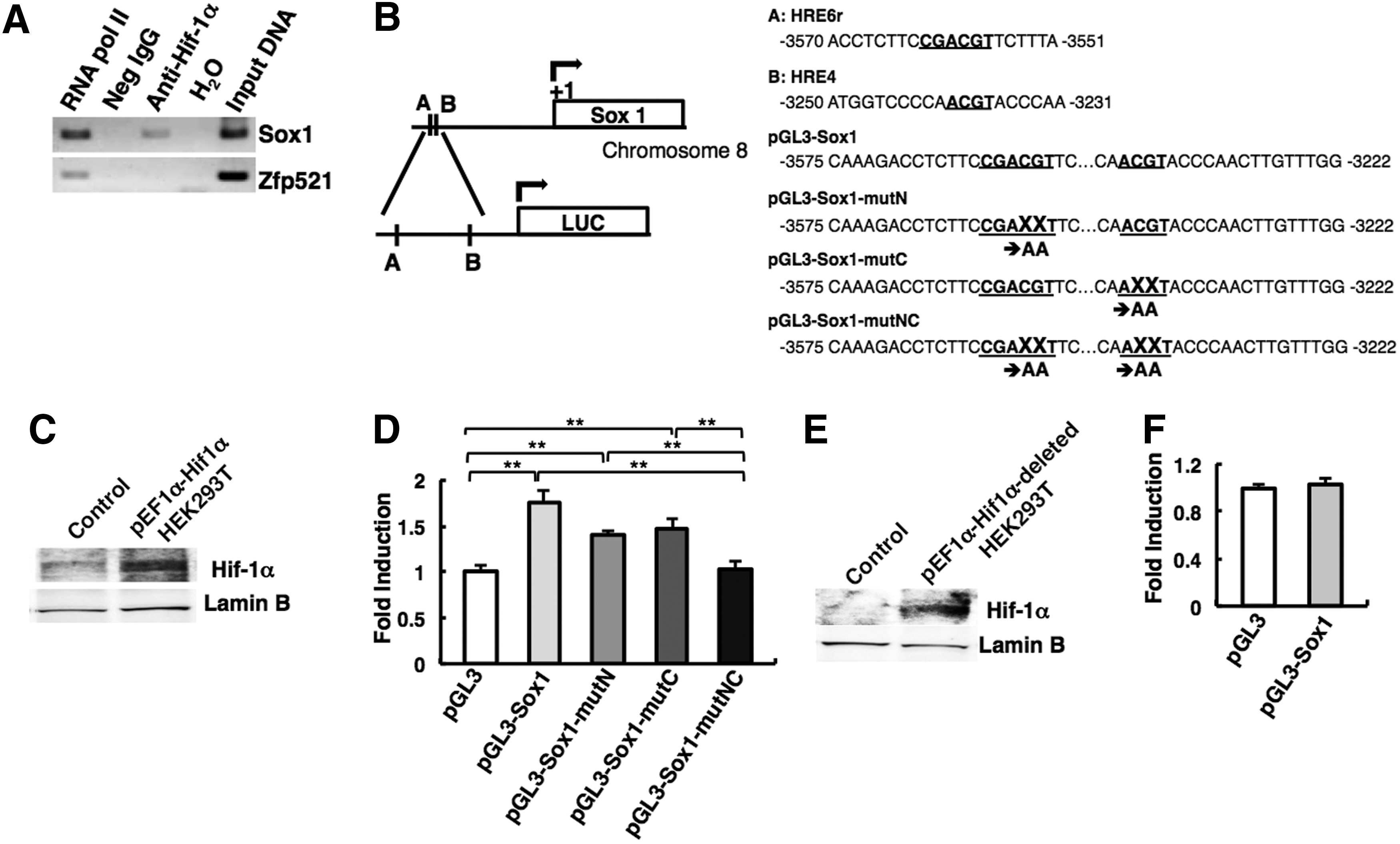
Hif-1α regulates Sox1 gene expression directly.
To prove this, we performed Luciferase assay using pGL3-Sox1 plasmid that includes the Sox1-5′ flanking sequence with two putative HRE sites. The overexpression of HIF-1α protein in HEK293T cells was confirmed by western blot (Fig. 5C). As shown in Figure 5D, significant increase of luciferase activity was observed when pGL3-Sox1 plasmid was cotransfected with pEF1α-Hif1α plasmid. Importantly, this activity was significantly diminished when the point mutations were introduced into both HRE sites (Fig. 5D). These results indicate that Hif-1α directly regulates Sox1 gene expression via the binding to both the HRE sites.
It has been reported that Hif-1α interacts with transcriptional coactivator p300 and regulates the activity of many target genes [43]. We constructed the Hif-1α carboxyl-terminal deletion mutant that lacks the domain for the association with p300, and performed luciferase assay (Fig. 5E, F). Intriguingly, this mutant failed to induce luciferase activity from pGL3-Sox1 plasmid. This result suggests that p300 is associated with HIF-1α on the HRE regions in Sox1-5′ flanking sequence.
Negative regulation of BMP signaling by Hif-1α
As Kamiya et al. have reported that extrinsic BMP4 inhibits Zfp521 expression in mouse ES cells, we examined Hif-1α expression in SFEBq culture after treatment with BMP4. There was no obvious reduction of Hif-1α expression in SFEBq culture (Supplementary Fig. S1B), indicating that Hif-1α is not a downstream target of BMP signals. The expression of Bmp4 gradually decreased with progressing differentiation in SFEBq culture; conversely, the expression of Hif-1α increased (Fig. 6A). According to these results, we predicted that Hif-1α is involved in neuroectoderm differentiation via negative regulation of endogenous BMP signaling. To prove our hypothesis, we analyzed gene expression related to BMP signaling in SFEBq culture. As shown in Figure 6B, Hif-1α-knockdown cells in SFEBq culture exhibited drastic elevation of Bmp4 expression on day 4 compared with control.
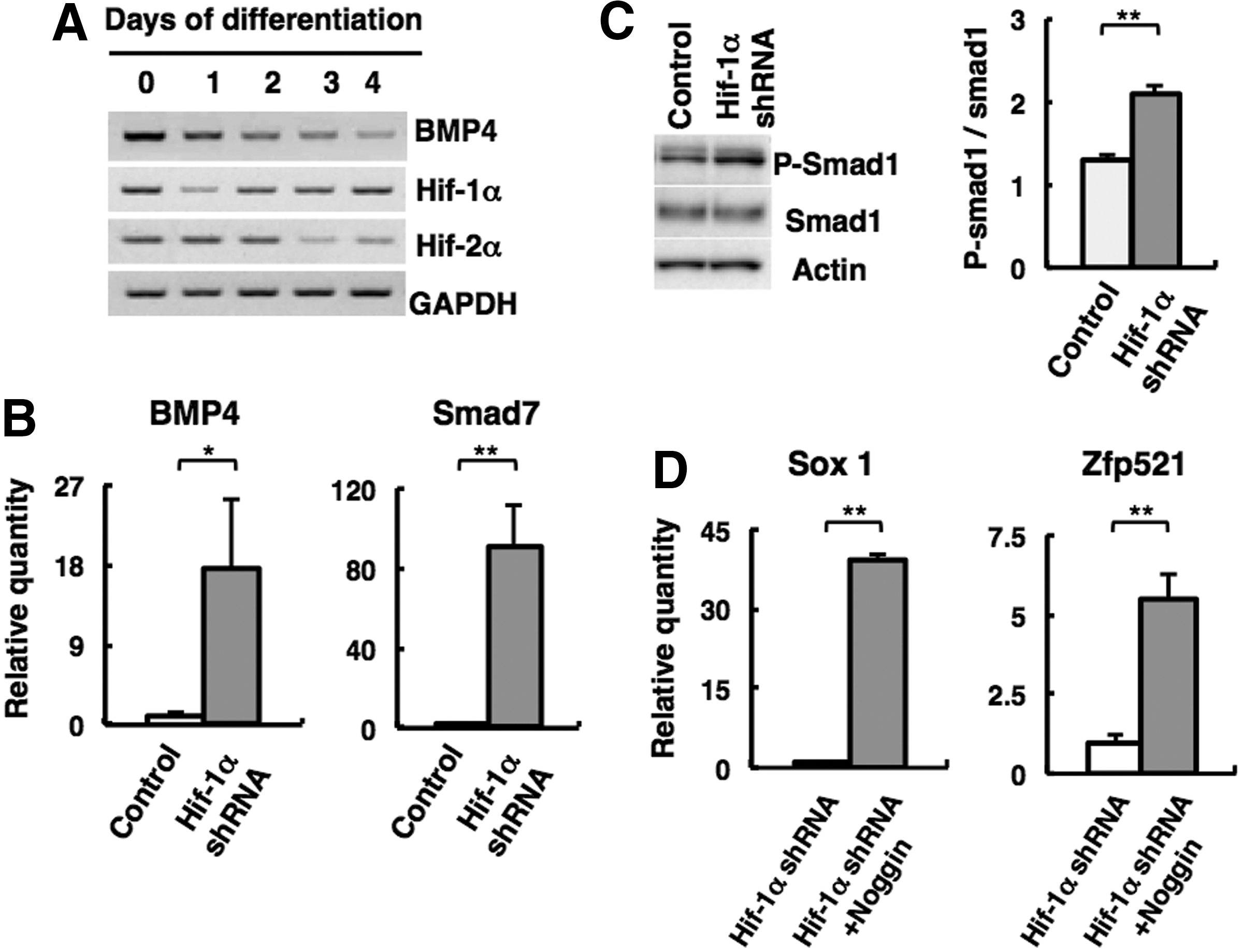
Negative regulation of BMP signaling by Hif-1α during neural commitment.
To investigate whether Hif-1α affects BMP signaling at the intracellular level, western blotting was performed for activated Smad1/5/8 in cells in SFEBq culture. As shown in Figure 6C, Hif-1α-knockdown cells in SFEBq culture showed significant elevation of phosphorylated Smad1/5/8 compared with control. In addition, Smad7, a direct target of the BMP/Smad signaling pathway [44], was highly expressed in Hif-1α-knockdown cells in SFEBq culture (Fig. 6B).
If the increased BMP signaling in Hif-1α-depleted ES cells is one of the reasons for the failure of neural differentiation, then it is expected that the addition of anti-BMP molecule would lead to recovery of neural differentiation in Hif-1α-depleted ES cells. We treated SFEBq culture with anti-BMP molecule, Noggin protein [19], and checked differentiation status. As shown in Figure 6D, addition of recombinant Noggin protein could derepress early neural gene expression (Sox1 and Zfp521) in Hif-1α-knockdown ES cells. These findings further support the idea that Hif-1α functions as a negative regulator of BMP signaling in SFEBq culture and promotes neural commitment.
Discussion
In this study, we have demonstrated that Hif-1α is crucial for neural commitment of mouse ES cells. Under hypoxic conditions, adherent ES cells expressed Hif-1α (Fig. 2F) and manifested significant increase of expression of neural progenitor genes, such as Sox1 (Fig. 2E). In addition, Hif-1α attenuation led to strong inhibition of neuroectoderm differentiation (Fig. 3C, D), suggesting that Hif-1α is a specific mediator of the hypoxic response in mouse ES cell aggregates and, so, plays a critical role in neural commitment. Covello et al. have reported that Hif-2α is a direct upstream regulator of Oct4, contributing to maintaining pluripotency of mouse ES cells [45]. It has also been shown that knockdown of HIF-2α and/or HIF-3α, but not HIF-1α, leads to significant decrease in expression of pluripotency genes, such as OCT4, NANOG, and SOX2, in human ES cells [46]. We also found that Hif-2α expression in mouse ES cells quickly decreased after starting SFEBq culture (Fig. 6A). In contrast, Hif-1α expression continued during differentiation in SFEBq culture through the period of neuroectodermal fate determination (Figs. 6A and 1C).
Forced expression of Hif-1α in mouse ES cells induced high Sox1 expression (Fig. 4D), and ChIP assay showed that Hif-1α protein specifically associates with the Sox1 locus (Fig. 5A). In addition, luciferase assay showed that HRE sequences in Sox1-5′ flanking region are necessary for the transcriptional activation by Hif-1α (Fig. 5D). These results suggest that Hif-1α is capable of promoting differentiation of ES cells into neuroectoderm via direct regulation of Sox1. We have also analyzed the expression of other Sox family genes, such as Sox2, Sox3, and Sox21. Sox2-null mouse embryos die around the time of implantation, reflecting its exclusive expression in the inner cell mass (ICM) [47]. In our experiment, there was no significant difference in Sox2 expression between control and Hif-1α-depleted cells (Fig. 3C). Sox3 begins to appear at epiblast stage, and is then expressed in neuroepithelium, gut ectoderm, and prospective sensory placodes during embryogenesis [36]. Sox1- or Sox3-knockout mice display only mild abnormalities in CNS [48]; therefore, there is a possibility that functional redundancy between Sox1 and Sox3 might exist during CNS development. However, we did not see significant decrease of Sox3 expression in Hif-1α-knockdown cells in SFEBq culture (data not shown), suggesting the specific interaction between Sox1 and Hif-1α at the stage of early neuroectodermal differentiation. Sox21 is expressed in neural stem/progenitor population in embryonic mouse brain, and is reported as one of the neural stem cell marker [49]. We saw the decrease of Sox21 expression (Fig. 3C). This indicates that Hif-1α-depleted ES cell could not differentiate into neural stem cells.
Previously, it has been reported that overexpression of transcriptional coactivator p300 promotes neuroectoderm differentiation in SFEBq culture [22]. Hif-1α interacts with p300 and regulates the activity of many target genes [43], suggesting that Hif-1α may directly activate Sox1 together with transcriptional coactivator p300 in mouse ES cells. We constructed the Hif-1α carboxyl-terminal deletion mutant that lacks the domain for the association with p300, and performed luciferase assay as shown in Figure 5F. Intriguing result was that this mutant failed to induce luciferase activity from pGL3-Sox1 plasmid. It is likely that p300 is associated with HIF-1α on the HRE regions in Sox1-5′ flanking sequence in the process of neural differentiation.
BMP4 is a critical regulator of dorsoventral patterning in development and is also known to be a potent inhibitor of neural differentiation in vertebrates [18 –21]. In mouse ES cells, addition of recombinant BMP4 into differentiation medium leads to inhibition of neural differentiation [5,18,22,34,50]. This was also confirmed in our experiment using SFEBq culture (Supplementary Fig. S1B). Interestingly, the inhibitory effect of adding recombinant BMP4 on neural commitment was only observed in a limited period (Day 2–4, Supplementary Fig. S1C). This BMP4-sensitive window corresponds to the epiblast stage in development [50], which differentiates to form somatic cells of all three germ layers. Notably, BMP4 acts in the epiblast to induce primordial germ cells [51,52]. Thus, it is likely that endogenous BMP signaling in SFEBq needs to be maintained at a low level during epiblast-like stage. However, the molecular mechanism that negatively regulates BMP signaling in ES-cell-derived epiblast-like cells has not yet been elucidated. In the present study, we demonstrated that Hif-1α is expressed in the hypoxic microenvironment of SFEBq at an epiblast-like stage (Fig. 1B, C). Further, we showed that attenuation of Hif-1α results in increase of BMP signaling in ES cell aggregates and leads to inhibition of neural commitment (Figs. 3C and 6B, C). Importantly, addition of anti-BMP molecule recovered the neural gene's expression to some extent in Hif-1α-knockout ES aggregates (Fig. 6D). These results suggest that Hif-1α is a key molecule that acts to suppress endogenous BMP signaling of mouse ES cell aggregate at an epiblast-like stage and promotes neural commitment of mouse ES cells.
In contrast, Hif-1α-knockdown cells in SFEBq culture had a significantly elevated expression level of undifferentiated ES cell markers, such as Oct3/4 and Nanog, as compared with control cells in SFEBq culture on day 4; additionally, the differentiation into the epiblast-like cells seemed to be inhibited (Fig. 3C). Previously, Zhang et al. have demonstrated that BMP4 inhibits conversion of ES cell (Oct4+ cell population) to epiblast-like cells (Fgf5+ cell population) during neuroectoderm differentiation [50]. Thus, there is a possibility that the elevated BMP signaling in Hif-1α-knockdown cells in SFEBq culture may have an inhibitory effect on the differentiation process (see model in Fig. 7).
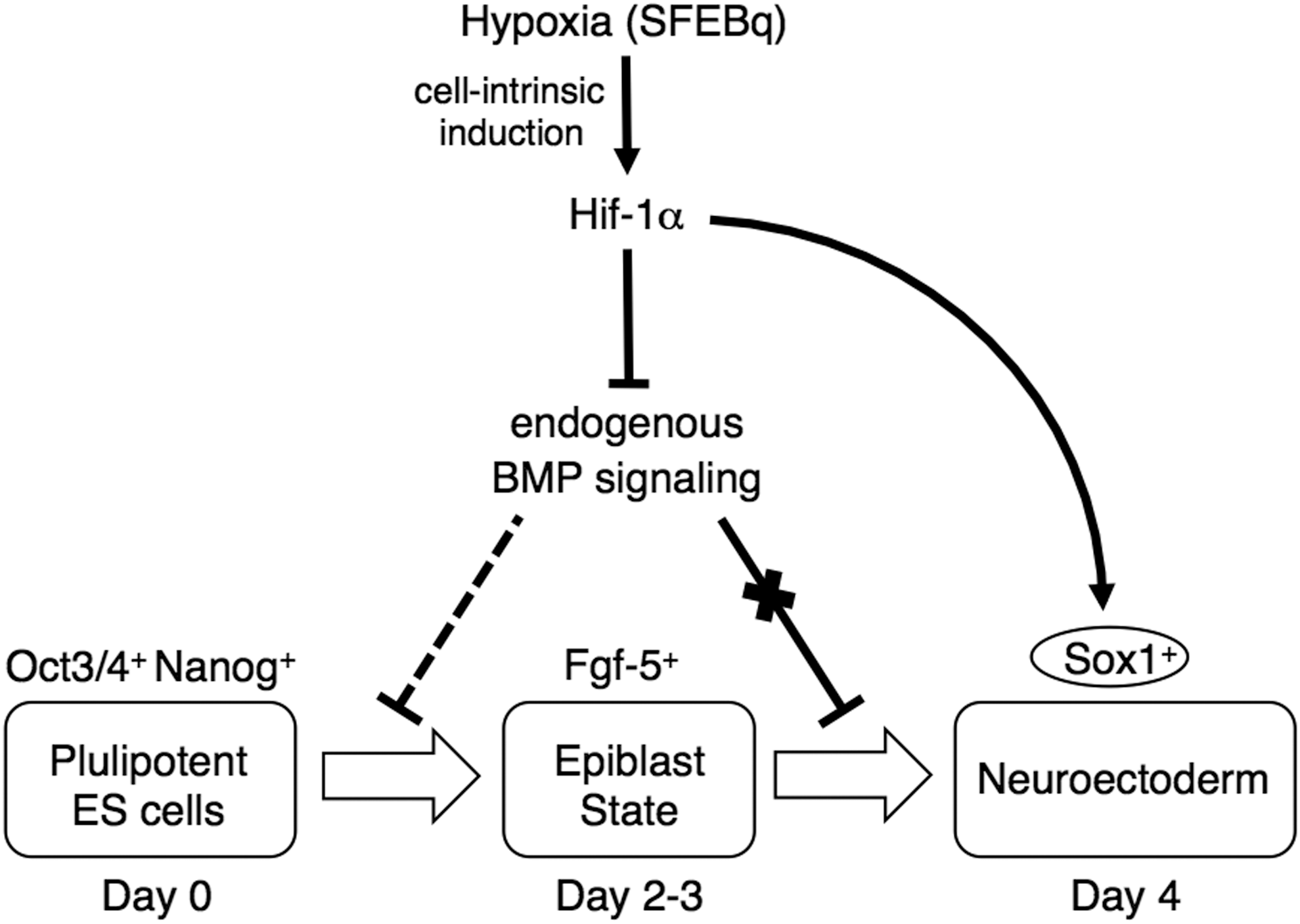
Working model for the dual roles of Hif-1α in neuroectoderm differentiation. Hypoxic microenvironment is established in SFEBq-cultured ES cells, and cell-intrinsic induction of Hif-1α occurs. Hif-1α functions as suppressor of endogenous BMP signaling at distinct site of neural commitment, and leads to neural lineage differentiation. In addition, Hif-1α directly induces neural progenitor gene Sox1. These dual activities of Hif-1α promote transition of ES cell differentiation from the undifferentiated state to the neuroectodermal state.
In mouse ES cells, it is known that BMP signaling plays a crucial role in maintaining self-renewal by suppressing neural lineage differentiation in cooperation with LIF/STAT3 signaling [53]. Intriguingly, forced expression of Hif-1α showed reduction of Bmp4 and undifferentiated ES cell markers even in the presence of LIF, and substantial induction of neural lineage was observed (Fig. 4D). This finding suggests that Hif-1α acts to suppress Bmp4 expression in mouse ES cells, and triggers the first step of differentiation of ES cells to differentiate into neural lineage. It has been also reported that HIF-1α acts as a direct suppressor of LIF-specific receptor (LIFR) expression by binding to the HRE located in the LIFR promoter [54]. Thus, Hif-1α might play a central role in the differentiation in mouse ES cells by suppressing both signaling pathways (LIF/STAT3 and BMP). Our preliminary data showed that there is no significant difference of phosphorylated STAT3 level in SFEBq aggregates under both normoxic and hypoxic conditions (data not shown). Hif-1α might be responsible for the suppression of BMP signaling but not LIF/STAD3 signaling in neural specification of mouse ES cells, although further analysis will be required to confirm this possibility.
ChIP assay did not show interaction between Hif-1α and loci of Bmp4 (data not shown); consequently, we hypothesize that Hif-1α negatively regulates BMP signaling indirectly via a downstream mediator. Further study is required to clarify precise molecular mechanisms how Hif-1α inhibits BMP signaling.
Several groups have demonstrated that FGF signaling plays a pivotal role for neural specification in mouse ES cells [10,11]. However, it has been reported that addition of exogenous FGF does not promote neural differentiation in SFEBq culture [22], indicating that the role of FGF signaling is permissive rather than instructive. In accordance with this, no obvious difference in FGF-induced Erk1/2 signaling was seen between Hif-1α-knockdown SFEBq and control (Supplementary Fig. S3), suggesting that the role of Hif-1α in neural commitment may be specific to the regulation of BMP signaling.
In Figure 7, we summarize working model for Hif-1α as a pivotal regulator of neuroectoderm differentiation of mouse ES cells. In SFEBq culture, a hypoxic environment is induced spontaneously and cell-intrinsic activation of Hif-1α occurs in parallel with neural differentiation. Hif-1α advances neural commitment and suppresses endogenous BMP signaling at distinct site of action. Hif-1α also promotes transition of the developmental status of mouse ES cells to neuroectoderm via the direct regulation of neural progenitor gene Sox1. These dual functions of Hif-1α are indispensable for neuroectodermal differentiation in SFEBq culture.
In conclusion, our study showed that the efficient neuroectoderm differentiation observed in SFEBq culture is caused by Hif-1α activation under hypoxic environment. Hif-1α plays dual roles in regulating neural differentiation, suppressing endogenous BMP signaling during neural commitment and promoting the transition of ES cell differentiation from the epiblast state into the neural progenitor state via the direct regulation of the neural progenitor gene Sox1. These results strongly suggest an indispensable role of the hypoxic response in early neural development of mammals.
Footnotes
Acknowledgments
The authors are grateful to Ms. C. Fujisawa for wonderful technical assistance and Drs. H. Niwa and A. Smith for the gifts of mouse ES cells. This work was supported by Initiative for the Promotion of Young Scientists' Independent Research program (Special Coordination Funds for Promoting Science and Technology from the Japanese Ministry of Education, Culture, Sports, Science and Technology).
Author Disclosure Statement
The authors indicate no potential conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.