
Abstract
The most widely used technique for isolation of human bone marrow stromal cells (hBMSCs) from bone marrow includes density gradient centrifugation, recovery of the mononuclear cell population, and subsequent isolation of hBMSCs by virtue of their plastic adherence. During subsequent in vitro cultivation, they may lose their original characteristics since in vitro the stem cell niche cannot yet be properly mimicked. To further characterize these culture-induced changes in regard to mRNA and extra- and intracellular protein expression, as well as potential differences between hBMSCs from different donors, we investigated a panel of CD antigens for their presence on in vitro cultured hBMSCs. Interestingly, after culture-induced downregulation of their extracellular expression, both CD146 and CD271 persist intracellularly, which hints at the possibility that culture-induced changes may be reversed by appropriate stimuli. Further, CD34—a protein whose expression on hBMSCs is still controversial—is expressed at the intracellular level in hBMSCs of all donors independently of passage number. CD34 mRNA levels are significantly higher in female than in male donors. In summary, we further elucidate phenotypical changes induced by in vitro culture of hBMSCs, highlight interindividual differences in the phenotype of these cells and for the first time show the intracellular expression of CD34.
Introduction
I
Due to the enormous importance of cell surface antigens for the characterization and isolation of specific cell populations, the so-called CD (cluster of differentiation) nomenclature has evolved. The biological functions of CD antigens are diverse; they can act as receptors or ligands, play a role in cell adhesion, function as costimulatory molecules, or work as antibody Fc receptors.
Mesenchymal stem cells (MSCs) reside in the perivascular region of bone marrow and can be propagated in vitro resulting in the generation of a heterogeneous population of more or less undifferentiated precursor cells called bone marrow stromal cells (BMSCs) [1] that can differentiate to adipocytes, chondrocytes, fibroblasts, osteoblasts, and/or stroma cells. In regard to the heterogeneous population of human bone marrow stromal cells (hBMSCs), a set of surface proteins has been defined over the years that has become the minimal standard to identify hBMSC cultures (International Society for Cell Therapy [2]: CD45−, CD34−, CD14− or CD11b−, CD79alpha− or CD19− and HLA-DR−, as well as CD73+, CD90+, and CD105+). Since then, efforts have been made to define a more concise phenotype of hBMSCs and to further characterize the changes in surface protein expression induced by their propagation in vitro. In 2011, a review by Mafi et al. was dedicated to update all available published information on the cell surface characterization of hBMSCs [3]. According to these authors, the cell surface markers that have been reported as being expressed on the cell surface of BMSCs in more than one study include CD13, CD29, CD44, CD54, CD73, CD90, CD105, CD106, CD146, and CD166. Those being reported as negative in more than one study are CD10, CD11b, CD14, CD31, CD34, CD45, CD49d, and CD106, which controversially, was found to be positive in other studies and therefore, also a part of the “positive list.” Contradictory evidence also exists for the expression of CD34 on BMSCs, as this molecule has been found to be expressed by stromal precursors in human adult bone marrow [4].
Lately, CD146 and CD271 have emerged as CD markers by the use of which an especially efficient enrichment of colony-forming unit-fibroblast (CFU-F) can be achieved [5 –7]. With the help of these markers, the in vivo phenotype of human MSCs is now becoming more defined. As MSCs usually have to be propagated in vitro to generate sufficient numbers for prospective in vivo applications, as well as to gain insight into the mechanisms of their biology, it is nevertheless important to characterize the phenotypical changes induced by in vitro culture and inter-individual differences.
The “classical” method of isolation for hBMSCs involves a density gradient (1.073 or 1.077 mg/mL) to isolate the mononuclear cells and subsequent selection of the plastic-adherent cell population [8 –10]. The plating of these mononuclear cells can be done either at clonal or at nonclonal density, with the former method allowing for the generation of a cell population derived from a single clone. Independent of the plating method, the resulting cell population is intrinsically heterogeneous with respect to morphology, cell size, proliferation, and differentiation abilities [11,12]. These are reflected by heterogeneous molecular properties like varying alkaline phosphatase expression in different clones of BMSCs or varying secretion of growth factors [11].
To assess cell surface markers quite a number of cells is usually required to perform FCM. This necessitates in vitro expansion before analysis and may confound the cell surface phenotype that the cells originally exhibited in vivo where they reside in stem cell niches and are influenced by soluble factors, cell–cell adhesions and many more parameters, which are still weakly defined especially in the case of MSCs. Searching for a continuous passage-dependent change in the expression of a certain CD antigen may help clarify its presence or absence in vivo.
We have analyzed and compared a panel of 18 hBMSC preparations isolated from bone marrow aspirates that were obtained by iliac crest aspiration during routine orthopedic procedures with respect to expression of different CD antigens. Certain proteins are stored intracellularly and may be presented at the cell surface upon stimulation with appropriate ligands or cytokines. In the context of hBMSCs, the most famous examples are the exposure of HLA-DR antigens on the cell surface after exposure to interferon gamma [13] and the enhanced cell surface presentation of CD54 induced by inflammatory cytokines [14]. Because of this, we have evaluated both extra- and intracellular protein levels, as well as mRNA levels.
Materials and Methods
Materials
Ethanol and paraformaldehyde (PFA) were purchased from Carl Roth GmbH. Cell culture flasks were from Greiner. Dulbecco's modified Eagle's medium (DMEM) with 1 g/L
Human MSC isolation and culture
For studies involving human tissues we obtained ethical approval from the ethical committee of Medical School Hannover. Written informed consent was obtained from all donors. All personal information, including age and gender was made anonymous. Bone marrow aspirates were harvested by iliac crest aspiration during routine orthopedic procedures from 18 otherwise healthy donors (Table 1). Human MSCs were isolated from fresh bone marrow aspirates by density gradient centrifugation and subsequent plastic adhesion of mononuclear cells as described elsewhere [15]. Cells were cultured at 37°C with 5% CO2 at 85% humidity. The cells were passaged at a density of around 70% by the use of 0.025% trypsin-EDTA solution and seeded at a density of 2×103 cells per cm2 in MSC growth medium [DMEM supplemented with 10% (v/v) FCS (not heat-inactivated), 20 mM HEPES, 1% (100 U/mL/100 μg/mL) penicillin/streptomycin, and 2 ng/mL human recombinant FGF-2].
hBMSCs, human bone marrow stromal cells.
Flow cytometry
Cells were detached using 0.025% trypsin-EDTA solution and washed twice with FACS buffer [2% (v/v) FCS in PBS]. All centrifugation steps were performed at 300g and 4°C for 5 min. For each sample 1×105 cells were used. To verify that the enzymatic cell detachment with Trypsin does not influence the surface expression of the proteins analyzed in this study, we compared the surface expression by FCM following cell detachment with Trypsin to the surface expression after nonenzymatic cell dissociation (Cell Dissociation Solution Non-enzymatic; Sigma). The results are presented in Supplementary Fig. S1 (Supplementary Data are available online at
For intracellular analysis, cells were fixed and permeabilized: 1×105 cells per sample were centrifuged and resuspended in 100 μL PBS supplemented with 0.5% (w/v) PFA in PBS and 0.01% (w/v) Tween-20. After incubation (20 min, 37°C and shaking at 400 rpm) and two washing steps with FACS buffer, cells were resuspended in 100 μL PBS supplemented with 0.5% (w/v) saponin and 1% (w/v) bovine serum albumin (BSA) and incubated for 10 min at room temperature with shaking at 400 rpm.
For both intra- and extracellular analysis, cells were then centrifuged and incubated with appropriate fluorochrome-conjugated antibodies, which were purchased from BioLegend (Table 2) for 30 min at 4°C in the dark. After two washing steps, with FACS buffer cells were analyzed on a FACS Canto (BD Biosciences). Each flow cytometric analysis consisted of a record of 3×104 cells. Cells were measured in the FL2 (PE) and FL1 (FITC) channel. Dead cells were excluded by using scatter parameters using BD FACSDiva Software and Flowing Software version 2.5.0.
Immunocytochemistry
Cells were grown on poly-
RNA preparation and cDNA synthesis
RNA preparation and cDNA synthesis were performed as described before [15]. RNA concentration and purity was determined by measuring the absorption at 260/280 nm. 1 μg of RNA was used for cDNA synthesis, which was carried out after DNase treatment to remove genomic DNA according to the protocol of the supplier using random hexamer primers.
Reverse transcription-polymerase chain reaction
Quantitative real time PCR analysis was used to quantify the transcripts of CD31 (Hs00169777_m1), CD34 (Hs00990732_m1), CD44 (Hs01075861_m1), CD73 (Hs00159686), CD105 (Hs00923996_m1), CD106 (Hs01003372_m1), CD146 (Hs00174838), and CD271 (Hs00609977). 18 S-rRNA (Hs9999901_s1) was used as housekeeping gene. The gene specific assays and the TaqMan Fast Advanced Master Mix were purchased from Life Technologies. The PCR reaction was accomplished in a duplex run using the StepOnePlus Real-Time PCR System (Life Technologies) with the following conditions: 20s at 95°C and 40 cycles 1s at 95°C and 20s at 60°C. Differences between samples and controls were calculated based on 2−ΔΔCT method.
Semiquantitative reverse transcription-PCR (RT-PCR) was performed using GoTaq DNA polymerase according to the instructions of the manufacturer (Promega). The thermocycler conditions consisted of 90s at 94°C followed by 28–35 cycles of 30s at 94°C, 30s at the respective annealing temperature, 60s at 72°C, and 15 min at 72°C at the end. All reactions were performed in the linear range by appropriate choice of cycling conditions. The PCR primers and the annealing temperature are listed in Table 3. The transcriptional levels of hypoxanthine phosphoribosyl transferase 1 were used for normalization.
Data analyses
Statistical analyses were performed using GraphPad Prism 5.0 (GraphPad Software Inc.). Student's t-test was used to evaluate statistically significant differences between the values with 0.01≥p≥0.001 very significant=*.
Results and Discussion
The aim of this study was to characterize the expression pattern of different surface proteins (Table 4) on human bone marrow-derived stromal cells with emphasis on donor differences and culture-induced changes. In addition to BMSCs, bone marrow contains hematopoietic stem and precursor cells and differentiated cells. To ensure that the hBMSC preparations were free of contaminations by other cell types—specifically by marrow-derived macrophages, which also grow in a plastic-adherent manner—we chose a panel of CD antigens (CD3γ, CD8α, CD8β, CD11b, CD11c, CD14, CD19, CD31, CD45, and CD163) for further characterization of our cell populations. These antigens are representative markers for certain subtypes of blood cells or endothelial cells (see Table 4), but have repeatedly been reported to be absent on hBMSCs. Their expression was analyzed on mRNA and protein level. All these antigens were found to be negative (<2% of cells positive) in regard to surface protein expression (data not shown). The same was found in regard to mRNA expression except for CD14 and CD31, which were detected at mRNA level independent of passage number (Fig. 1). We confirmed the absence of protein expression for CD14 by extra- and intracellular FCM (Fig. 2A). Interestingly, although CD31 protein is not expressed on the cell surface of hBMSCs (Fig. 2B), it can be found intracellularly and is downregulated during in vitro culture (Fig. 2C).

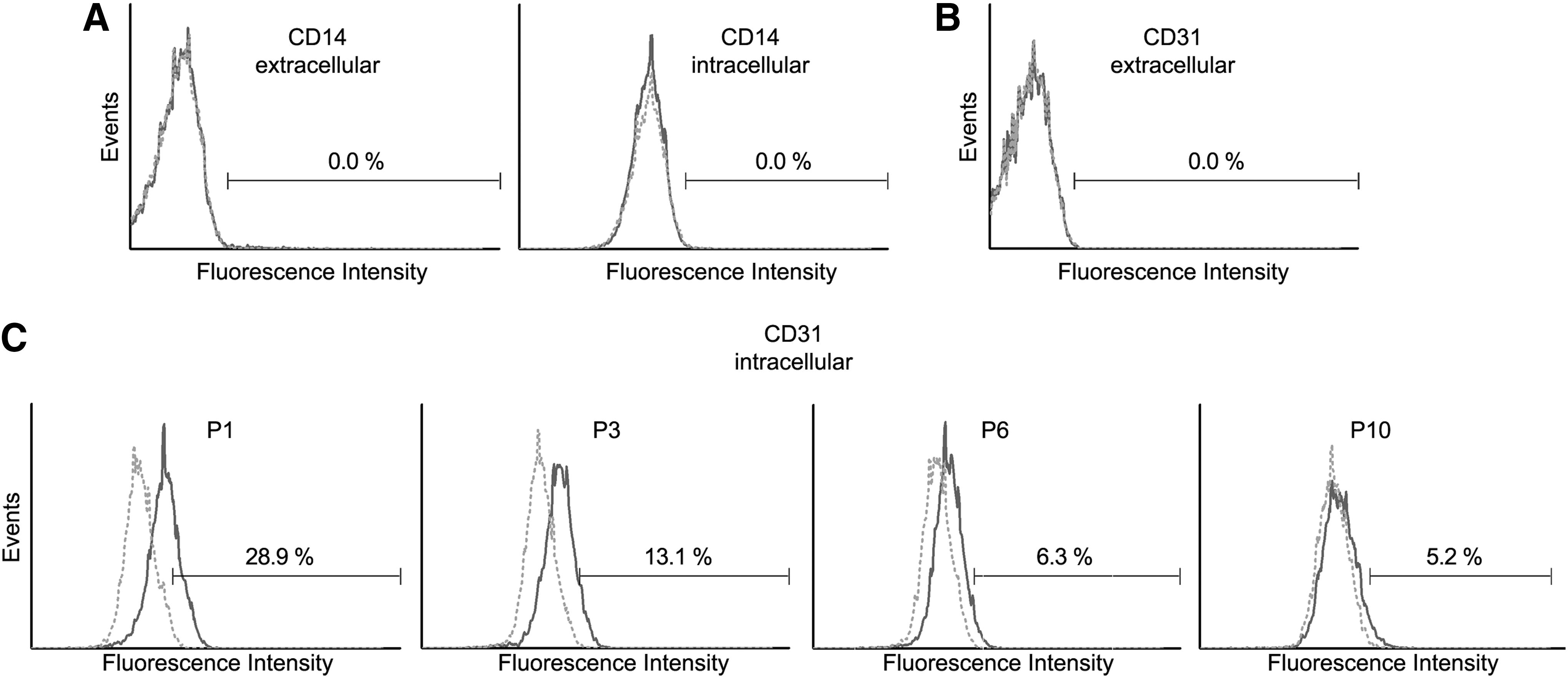
Extra- and intracellular flow cytometric analyses of CD14 and CD31 protein expression (dark gray lines). Dotted lines (light gray) are the respective isotype controls.
NCBI, National Center for Biotechnology Information.
In general, we detected the surface expression of CD13, CD29, CD44, CD56, CD73, CD90, CD105, CD106, CD146, CD166, and CD271 on hBMSCs, which is in line with the literature (Supplementary Fig. S2). These results were confirmed by immunocytochemistry (Supplementary Fig. S3).
Donor-dependent differences in mRNA expression levels of surface antigens
The analysis of mRNA expression for the surface antigens investigated revealed a high interdonor variability for CD106 and CD271 with a more than eightfold difference between the highest and lowest mRNA expression value (see Fig. 3). This variability is independent of donor age and gender for both surface antigens (see Table 1 for donor information). All other surface antigens showed small differences in mRNA expression with less than eightfold difference between the highest and lowest mRNA expression value in qRT-PCR or very similar band intensities in semiquantitative PCR (see Supplementary Fig. S4).
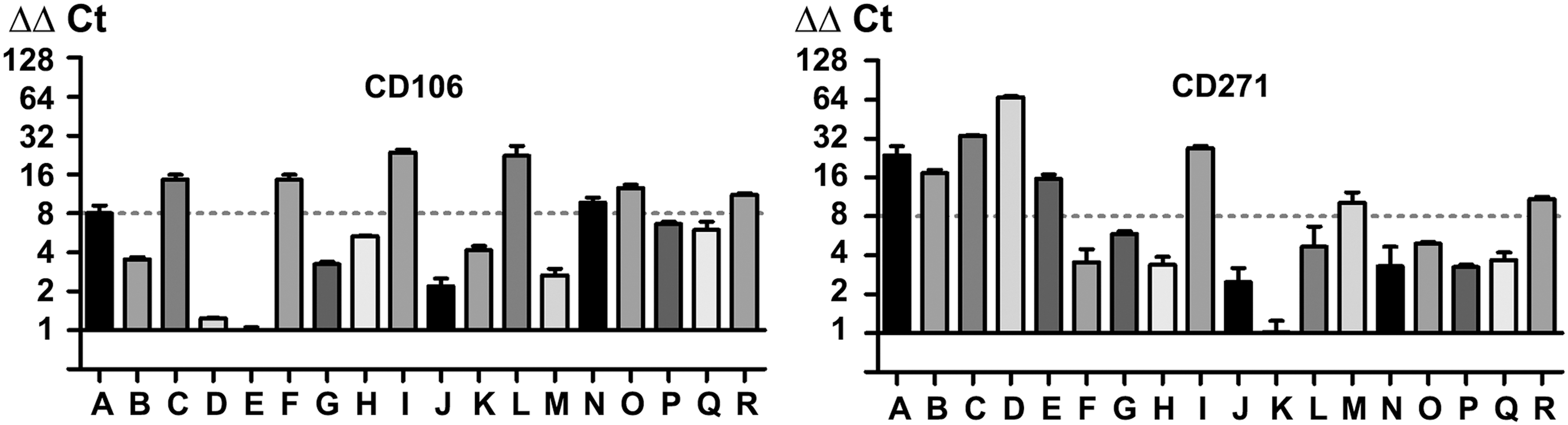
Donor-dependent mRNA (donor A–R) expression of CD106 and CD271 evaluated by qRT-PCR. Donor-dependent mRNA expression was normalized to the expression of donor with the lowest relative expression value. An eightfold difference in mRNA expression levels was used as arbitrary threshold to define notable differences in expression levels. This is marked in the graphs by a horizontal dotted line.
Passage dependence of surface antigens in hBMSCs
Throughout the 10 passages of in vitro culture analyzed in this study, the surface protein expression levels of CD13, CD29, CD44, CD73, CD105, and CD166 remain stable (positive in more than 95% of the cells) (see Supplementary Fig. S5). When taking into account that many studies have shown that the ability of these cells to form adipocytes, chondrocytes, and osteoblasts in vitro declines with the duration of in vitro culture [16], it seems that CD13, CD29, CD44, CD73, CD105, and CD166 are not directly correlated with the “stemness characteristics” of hBMSCs.
In contrast to this, CD56, CD146, and CD271 exhibit a notable downregulation of surface expression with prolonged in vitro culture (Fig. 4). Whereas the expression of both CD56 and CD271 declines dramatically, CD146 can still be found even in passage 10 in a part of the cells. Interestingly, the intracellular expression of CD146 is more stable (Fig. 5), hinting at distinct molecular mechanisms controlling protein expression and surface presentation. Regarding CD271, its surface protein expression declines more rapidly compared with CD146 during in vitro culture (Fig. 4). We found the CD271 protein to be expressed intracellularly in a slightly larger part of the cells than extracellularly (Fig. 5). Regarding the expression of CD56 in hBMSCs, contradictory data can be found in the literature. While some studies describe CD56 to be absent from the surface of hBMSCs, others find it as defining a MSC subpopulation [17]. Brooke et al. described the expression of CD56 mRNA in both hBMSCs and placenta-derived MSCs [18] and Mariotti et al. have subsequently identified the expression of CD56 on the cell surface of placenta-derived MSCs but did not detect it on hBMSCs [19]. Lastly, Battula et al. show a positive link between the expression of CD56 and an enrichment of CFU-F [20]. In light of this controversy, it is a very interesting fact that CD56 surface expression was downregulated in response to in vitro culture in this study. It seems worthwhile to investigate this in the future in respect to hBMSC differentiation potential and migratory ability.
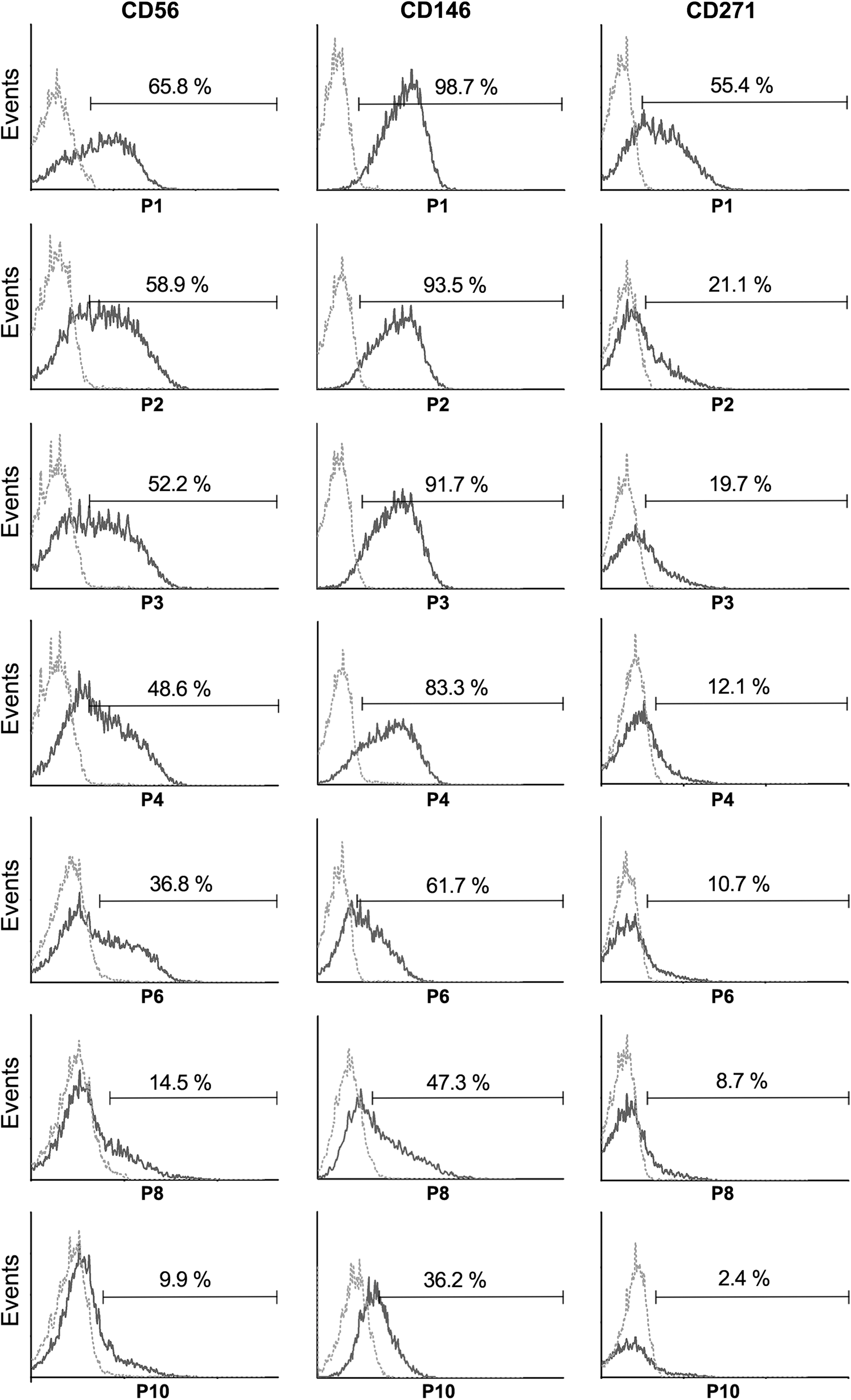
Surface expression changes of CD56, CD146, and CD271 during in vitro culture of hBMSCs from passage 1 to passage 10 (passages 1, 2, 3, 4, 6, 8, and 10 are shown) of one representative donor (donor M) as evaluated by flow cytometry (dark gray line). Dotted lines (light gray) represent the respective isotype controls. hBMSCs, human bone marrow stromal cells.

Intracellular protein expression changes of CD146 and CD271 during in vitro culture of hBMSCs shown for passages 6, 8, and 10 from one representative donor (donor M, dark gray line). Dotted lines (light gray) represent the respective isotype controls.
The expression of CD106 is highly variable between passage 1 and 10 with neither a positive or negative trend detectable (Fig. 6). Although it is thought that CD106 might be an appropriate marker for hBMSCs distinguishing them from osteoblasts [21], the plasticity in surface expression that we found in this study lets us doubt the suitability of CD106 as a marker protein and may explain the contradictory studies reviewed by Mafi et al. [3]. A recent study by Yang et al. shows that both CD106 positive and negative cell populations from human BMSCs are able to undergo adipogenic and osteogenic differentiation but differ in their immunomodulatory properties [22]. This, taken together with studies showing that CD106 expression is modulated by cell density [23], cellular senescence [24], and inflammatory cytokines [14] paints a picture of CD106 as a context-inducible protein with a highly plastic expression involved in immunomodulatory and migratory properties of hBMSCs.
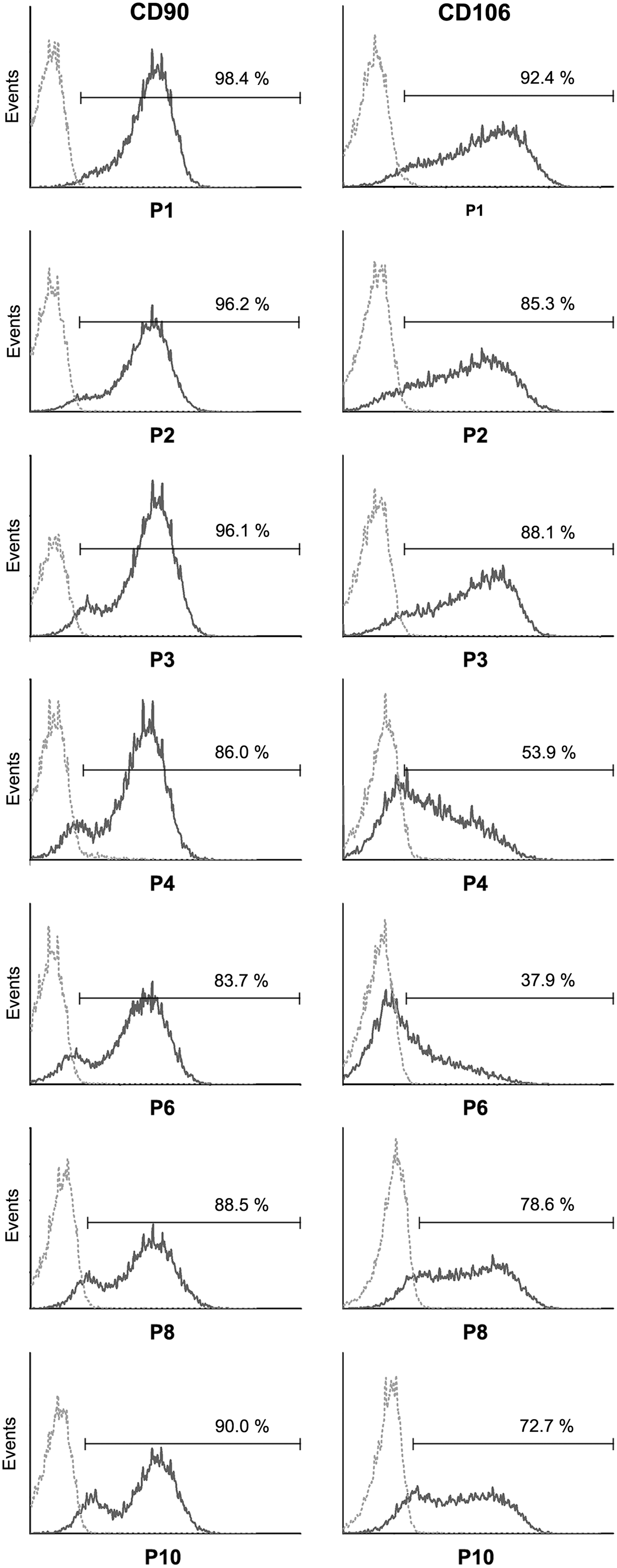
Surface expression changes of CD90 and CD106 during in vitro culture of hBMSCs from passage 1 to passage 10 (passages 1, 2, 3, 4, 6, 8, and 10 are shown) as evaluated by flow cytometry from one representative donor (donor M, dark gray line). Dotted lines (light gray) are the respective isotype controls.
All other surface antigens classically used to define hBMSCs that we have analyzed (CD13, CD29, CD44, CD73, CD105, CD166) remain unchanged positive between passage 1 and 10 (Supplementary Fig. S5) with the exception of CD90, for which all cells are highly positive at passage one but over time, a dim-positive cell fraction evolves during in vitro culture (Fig. 6). In conclusion, only CD56, CD90, CD106, CD146, and CD271 show considerable changes in their protein expression during in vitro culture.
CD34 protein is expressed intracellularly in hBMSCs independent of duration of in vitro culture
The most interesting finding relates to CD34. Although its extracellular expression is restricted to a small population of cells from some donors or even completely missing on cells from other donors [see Fig. 7A (donor P and D, respectively) for representative extracellular FCM data], the mRNA for CD34 is detected in the hBMSC samples from all donors analyzed. The level varies considerably between donors and passage (Fig. 7B and C, respectively). A statistically significant difference in expression levels is detected between male and female donors, with hBMSCs from female donors showing a higher CD34 mRNA expression (Fig. 7D: 0.001≤p≤0.01). Surprisingly, the protein is expressed intracellularly in virtually all cells independent of donor and passage (Fig. 7A: representative intracellular FCM data from donor D). This result was confirmed by immunocytochemistry with two different CD34 antibodies, one recognizing the class II epitope (clone QBEnd-10), the other recognizing the class III epitope on CD34 (clone 581) (Fig. 7E). Huge controversy exists as to the expression of CD34 on MSCs in vivo [25]. While hBMSCs are classically defined as cells lacking the expression of CD34 [2], one study has demonstrated that all CFU-F are contained in the CD34 positive cell fraction from human bone marrow [4]. Here we show for the first time that both CD34 mRNA and protein are expressed in hBMSCs during in vitro culture and that the mRNA expression level is significantly higher in hBMSCs from female donors.
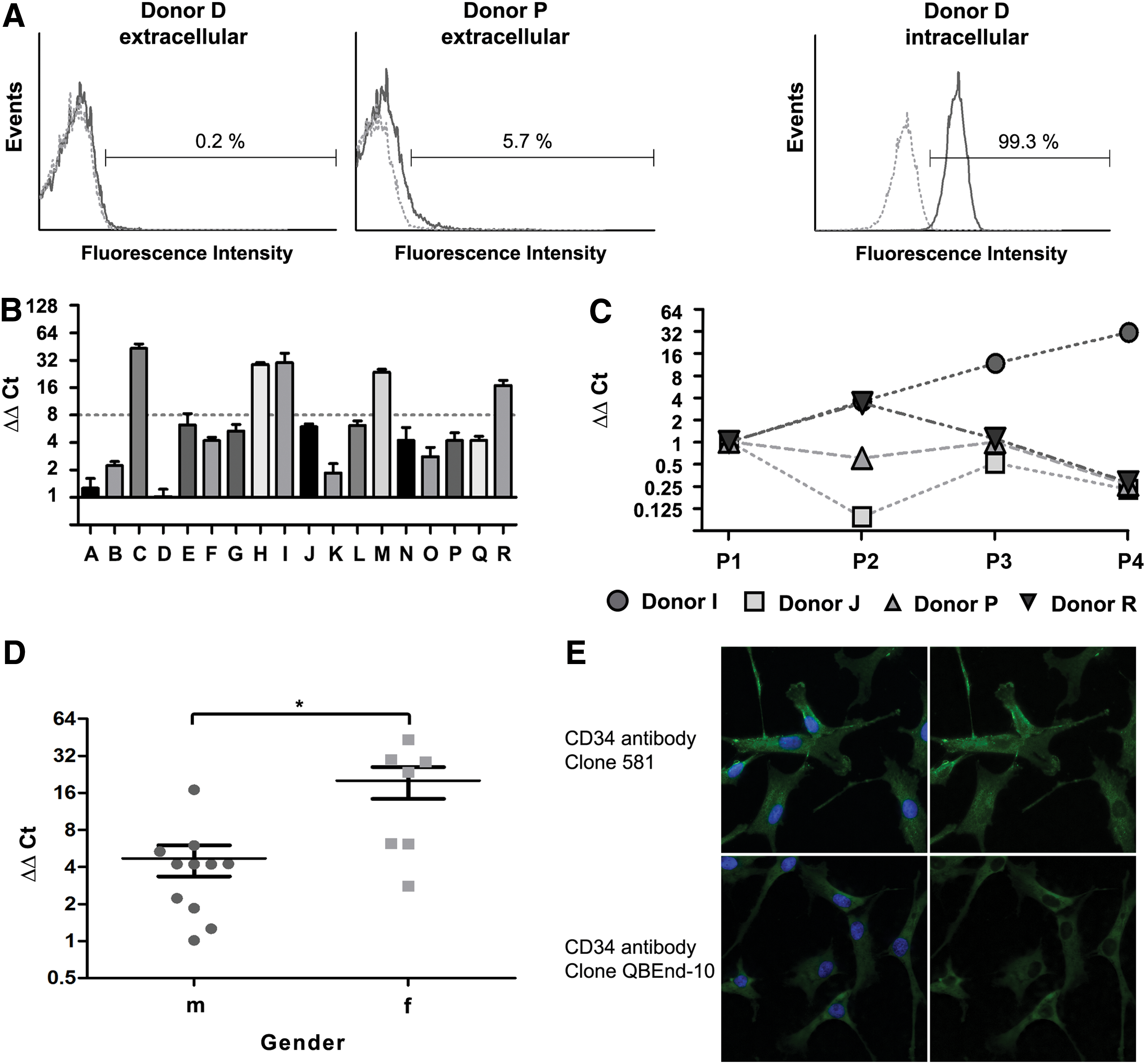
Expression of CD34 mRNA and protein in hBMSCs.
In conclusion, important findings of this study include the discovery that CD146 and to a lesser degree also CD271 persist intracellularly as proteins after their downregulation from the surface. The sustained intracellular expression of both CD146 and CD271 hints at the possibility that their expression on the cell surface might be reinducible and may implicate a reversibility of culture-induced changes on the phenotype of hBMSCs. It should be interesting to see whether these culture-induced changes can be avoided by culturing hBMSCs in the presence of ligands for these molecules.
Further, the intracellular occurrence of CD34 protein could signify that CD34 has an intracellular function previously ignored. In another scenario, CD34 is primed to allow for a quick mobilization to the cell membrane after an appropriate stimulus similar to CD146 and CD271. This theory is supported by the finding of Fackler et al. obtained from different hematopoietic cell types that CD34 can be stimulated to the cell surface by activation of protein kinase C [26]. A third possibility would be that hBMSCs produce soluble CD34 that is secreted by the cells, a process that has been shown to occur in human myeloid cells by Fernández et al. [27]. These possibilities have to be elucidated in the future and will help to better understand the role of CD34 in stem cell biology. The gender-specific difference in CD34 mRNA levels raises the question whether this is mirrored by a higher protein expression in hBMSCs from female donors and the possible functional consequences thereof.
Footnotes
Acknowledgments
The skillful technical assistance of Annika Hamm is gratefully acknowledged. Andrea Hoffmann acknowledges funding by the German Research Foundation (DFG) within SFB 599 and HO2058/4-1. Luisa Marilena Schäck gratefully acknowledges the financial support by the Studienstiftung des Deutschen Volkes.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.