
Abstract
The goal of this study was to determine whether a pro-survival cocktail (PSC, consisting of IGF-1, Bcl-XL, and Caspase-I Inhibitor) and long-term hypoxia (LTH) enhance survival and functional properties of bone marrow-derived stromal stem cells (BMSCs), in response to stress conditions. PSC-treated cells retained BMSC surface markers and protected cells from apoptosis under serum starvation and ischemic (1% O2 and 100 μM H2O2) conditions. LTH promoted osteogenesis, while suppressing adipogenesis. LTH alone did not result in an improvement in the apoptosis rate; however, PSC conferred significant protection regardless of the oxygenation status. One of the possible mechanisms of PSC protection was due to the elevated phospho-AKT in treated groups. PSC treatment or LTH did not alter migration toward stem cell-derived factor-1 alpha (SDF-1α) or fetal bovine serum, nor did they enhance cell motility during wound healing. There was no difference in the secreted cytokine profiles of BMSCs treated with PSC after stress when grown in normoxic or LTH. However, LTH did upregulate the vascular endothelial growth factor, hepatocyte growth factor, and SDF-1α, while it downregulated other anti- and proinflammatory cytokines and chemokines. We also observed a high degree of interdonor BMSC variability in response to pretreatment with PSC and LTH, confounding the functional results, underscoring the observation that not all donor-derived BMSCs will respond similarly.
Introduction
S
To improve cell survival, various approaches have been employed. Studies using microRNA overexpression and genetic modification methods prolonged cell survival, but these methods require viral transduction and manipulation of cells [15]. To overcome cell death by anoikis, cell spheres formed with the exogenous matrix have been surgically implanted in the peri-ischemic injury site and shown to improve engraftment, but this method requires invasive surgery for cell delivery [4].
Many studies employed pharmacological preconditioning with mitochondrial potassium-channel agonists such as diazoxide and pinacidil to suppress apoptosis [10]. Soluble growth factors such as stem cell-derived factor-1 alpha (SDF-1α) [16], vascular endothelial growth factor (VEGF) [17], insulin-like growth factor (IGF)-1 [18], and hepatocyte growth factor (HGF) [19] have been used to precondition cells. Combining these effects, a cocktail consisting of a pro-survival growth factor (IGF-I) and antiapoptotic drugs (caspase I inhibitor, Bcl-XL, cyclosporin A, and pinacidil) was used to improve cardiomyocyte survival and retention at the peri-infarct area [20].
Hypoxia has been implicated in increasing the proliferation rate and paracrine effect of BMSCs [10,21]. Culturing BMSCs in low oxygen tension (ie, <5%) simulates their physiological state and improves their genetic stability by lowering free radicals generated by oxidative stress [22]. Since ischemic tissue oxygen tension is typically 0.4%–2.3% [21], studies have used short-term (≤24 h) preconditioning of BMSCs in hypoxia to improve survival after transplantation. However, the effectiveness of hypoxic culture and preconditioning remains inconclusive due to conflicting results using different oxygen tensions, different hypoxic culture duration, and lack of long-term culture studies.
In this study, we preconditioned BMSCs with long-term hypoxia (LTH, defined as growth in hypoxic conditions for >7 days) and a modified pro-survival cocktail (PSC) [20] to investigate BMSC survival when challenged with ischemic conditions [ie, defined as 1% O2, presence of hydrogen peroxide (H2O2), and serum-free media]. To accommodate BMSC variability in our results, we used early passaged cells from several different donors in all experiments. Preconditioned BMSC survival, growth rate, and migration ability were characterized in response to ischemic conditions.
Materials and Methods
Cell culture and stress conditions
Human BMSCs were obtained from normal volunteers enrolled in an IRB approved clinical protocol in the NIH Bone Marrow Stromal Cell Transplantation Center at our institution and expanded in culture. Bone marrow donor information is documented in Table 1. BMSC cultures were used at passages 3–5, and were maintained in alpha-MEM medium (Invitrogen, Life Technologies) supplemented with 20% lot selected fetal bovine serum (FBS; Gemini Bio Products). Incubator conditions were as follows: hypoxia (Hpx, 37°C, 94% N2, 5% CO2, 1% O2) or normoxia (Nmx, 37°C, 74% N2, 5% CO2, 21% O2). Three or more different donor BMSCs were evaluated in triplicate for all experiments.
PSC was composed of insulin-like growth factor I (IGF-I, 50 nM; Peprotech), caspase inhibitor I (ZVAD-Fmk, 100 μM; EMD Millipore), and Bcl-XL (50 nM; EMD Millipore). Cells were incubated in a PSC-containing medium for 18–24 h, unless otherwise indicated. Figure 1 contains a graphic summary of the precondition schema used in this study.
Graphic depiction of the preconditioning scheme and stress conditions.
PSC-treated BMSCs were compared to untreated controls in both hypoxic and normoxic environments and experimental serum conditions were varied as necessary for the different types of assays. Analyses of apoptosis and cytokines were performed on cells grown in full serum (FS, 20% FBS) and in no serum (NS, 0% FBS) media. Because the BMSCs failed to proliferate in serum-free conditions, the cell proliferation assay was carried out in FS and low serum (LS, 2% FBS). The protective effects of hypoxia and PSC in various stress conditions, including serum starvation and H2O2 (100 μM) treatment were analyzed based on apoptosis and LDH assays.
All flow cytometry experiments were run on an Accuri C6 flow cytometer (BD Accuri, BD Biosciences), with results analyzed on the corresponding CFlow Plus software (BD Accuri, BD Biosciences).
BMSC characterization and differentiation
Characterization of cell surface markers was performed on BMSCs incubated with and without the PSC under LTH and normoxia in FS for 24 h. Cells were harvested, fixed with the Cytofix Buffer (BD Biosciences), and immunostained for the following known BMSC surface markers: CD29, CD44, CD73 (all PE conjugated; BD Biosciences), and CD105 (APC conjugated; R&D Systems). About 5×105 cells from each treatment condition were stained with each antibody and analyzed by flow cytometry, with a minimum of 104 events collected per sample.
The differentiation capacity of treated and untreated BMSCs was tested using StemPro® Adipogenesis and Osteogenesis differentiation kits (Invitrogen, Life Technologies) according to the manufacturer's instructions. Briefly, cells were grown to confluence in six-well plates, incubated with and without PSC in FS for 24 h, washed twice with HBSS, and switched to the appropriate differentiation medium. The differentiation medium was changed every 3 days for 2 weeks, with cells maintained in both hypoxia and normoxia. Adipogenic differentiation was detected with Oil Red O staining, and osteogenesis was confirmed by means of Alizarin Red staining. An Olympus BX—inverted scope, attached to an eight megapixal Olympus digital camera was used to obtain photomicrographs of differentiated BMSC populations.
Cell viability (apoptosis) and proliferation
Cell viability and proliferation after exposure to stress factors such as hypoxia, serum deprivation, and H2O2 were assessed as a measure of protection conferred by preconditioning. Viability and apoptosis were assessed in hypoxic and normoxic conditions by an annexin-V/propidium iodide (PI) assay (Vybrant Apoptosis Assay; Invitrogen, Life Technologies). About 5×104 cells per well were seeded in 24-well plates and allowed to attach overnight. The following day, cells were washed twice with HBSS and incubated with or without PSC in NS or FS media. All cells, including those from the supernatant, were collected and stained for annexin-V/PI according to the manufacturer's instructions, with results analyzed by flow cytometry. Cell proliferation and metabolic activity were measured using the MTS AqueousOne Solution (Promega). Cells were seeded in 96-well plates at 3×103 cells per well in the following conditions: FS, FS+PSC (FSP), LS, and LS+PSC (LSP). Readings were obtained in triplicate on days 1, 3, 5, and 7.
Stress conditions and cytotoxicity analysis (serum starvation, ROS)
To mimic the effects of stress on proliferation and survival, BMSCs were exposed to various stress conditions, including the following: (1) serum starvation (ie, 0% FBS, NS) for 3 days; and (2) H2O2 (100 μM) added to NS media for 18 h in 1% O2. Following each stress condition, supernatant was collected for lactate dehydrogenase (LDH) assay. Cells were harvested and either stained for apoptotic cells or lysated for protein preparation for western blots.
LDH assay
To assess stress and cytotoxicity caused by ROS and hypoxia, LDH assay was performed on the cell culture supernatant. After different cell stress conditions, 100 μL of supernatant was collected and mixed with an equal volume of LDH Assay Mixture (Sigma-Aldrich) followed by a 30-min incubation at room temperature. LDH release was measured spectrophotometrically at 490 nm and background absorbance at 690 nm was subtracted using a multiwell plate reader (PerkinElmer Lambda 25). All LDH release was normalized to the baseline LDH release by nonstressed counter parts.
Western blot analysis for protein kinase B (Akt)
To assess the survival pathway, activation of Akt was measured by western blot. Protein samples were harvested with a cell lysis buffer (Roche Applied Science) supplemented with a protease inhibitor cocktail (Santa Cruz Biotechnology) at 4°C. After removing the insoluble fraction by centrifugation at 15,000 rpm for 10 min at 4°C, total protein was quantified using the BCA Protein Assay Kit (Pierce Biotechnology). Twenty-five micrograms of protein was separated on Novex Bis-Tris gels (4%–12% acrylamide; Life Technologies) and then transferred to nitrocellulose membranes. After blocking with 5% nonfat milk in TBS containing 0.05% Tween-20 (TBS-T), membranes were probed with the following anti-human antibodies overnight at 4°C: phospho-Akt (1: 2000, Ser-473; Abcam); total Akt (1:200; Santa Cruz Biotechnology); β-actin (1:5000; Abcam). Secondary antibody incubation was for 1 h at room temperature with a horseradish peroxidase-conjugated donkey IgG (GE Healthcare) against either mouse or rabbit at a 1:10,000 dilution. Blots were incubated with an enhanced chemiluminescence detection system (Life Technologies) for 2 min and exposed on autoradiograph films. Densitometry analysis was done using ImageJ (Version 1.44; National Institutes of Health).
Cell migration
Cell migration assays were performed using a fluorescence-based 96-well plate transwell chemotaxis system (Millipore). Ninety percent confluent BMSCs cultured under LTH or normoxia were incubated in FS with or without PSC for 18–24 h, followed by an 8-h incubation in the NS medium. The cells were harvested, washed twice with HBSS, and seeded at 5×104 per well in the upper chamber in the NS medium. The bottom chambers contained the following chemokines in triplicate: no serum as negative control, SDF1α (200 ng/mL; Peprotech), PDGF-AB (50 ng/mL; Peprotech), and 20% FBS. Migration was allowed to occur overnight at 37°C under LTH or normoxia conditions, and the cells that migrated to the bottom side of chambers were quantified the following day using the CyQUANT Cell Proliferation Assay (Life Technologies) according to the manufacturer's instructions.
Cell motility (wound healing assay)
BMSCs cultured under LTH or normoxia were plated in 24-well plates to confluence, and incubated with or without PSC in FS for 18–24 h. An artificial wound was created by scratching the confluent monolayer of BMSCs with a 200 μL pipette tip. Cells were washed twice with HBSS, and incubated in the NS medium in hypoxia or normoxia for 24 h. Images of wounds at 0 and 24 h were compared and cells that migrated into the damaged area were counted. All conditions for each donor were done in quadruplicate in at least three independent experiments.
Immunoassay for human cytokines, chemokines, and growth factors
To evaluate changes in the paracrine activity of BMSCs with LTH preconditioning and PSC treatment, 27- and 21-plex human cytokine, chemokine, and growth factor panels (Bio-plex; Bio-Rad) were run on the supernatants from normoxic and hypoxic culture cells with or without PSC. For the paracrine activity of BMSCs in the ischemic tissue, 3.5×105 BMSCs were plated in 60-mm tissue culture dishes for a day, and transferred to 1% O2 with NS medium for 1 day in which the supernatant was collected. For paracrine activity in wound healing, the supernatant was collected after scratching with a 200 μL pipette tip as described in the wound healing assay (see above). Bio-plex was run according to the manufacturer's instructions with 50 μL of neat supernatant in duplicates.
Statistics
All experiments were performed at least three times in n>3 in each donor-derived BMSCs. All data are expressed as mean±SD unless otherwise described. The Student's t-test and one-way ANOVA was used for statistical analysis using GraphPad Prism Software.
Results
To investigate whether PSC treatment alters the proliferation of BMSCs, five different donor BMSCs were cultured under LS (2% FBS) in the presence or absence of PSC. PSC treatment had a positive effect on BMSC growth under LS conditions. Whereas LS control cells expanded <1.5-fold over 5 days, LS+PSC (LSP) cells showed a two- to threefold increase in cell numbers during the same time, whereas FS controls expanded two- to fivefold (Fig. 2). The growth advantage provided by PSC treatment was significant in all donors (P<0.05, paired t-test) under LS in normoxia. However, a growth advantage with PSC treatment was not discernable in FS (data not shown), indicating that PSC acts in a protective rather than an additive manner.
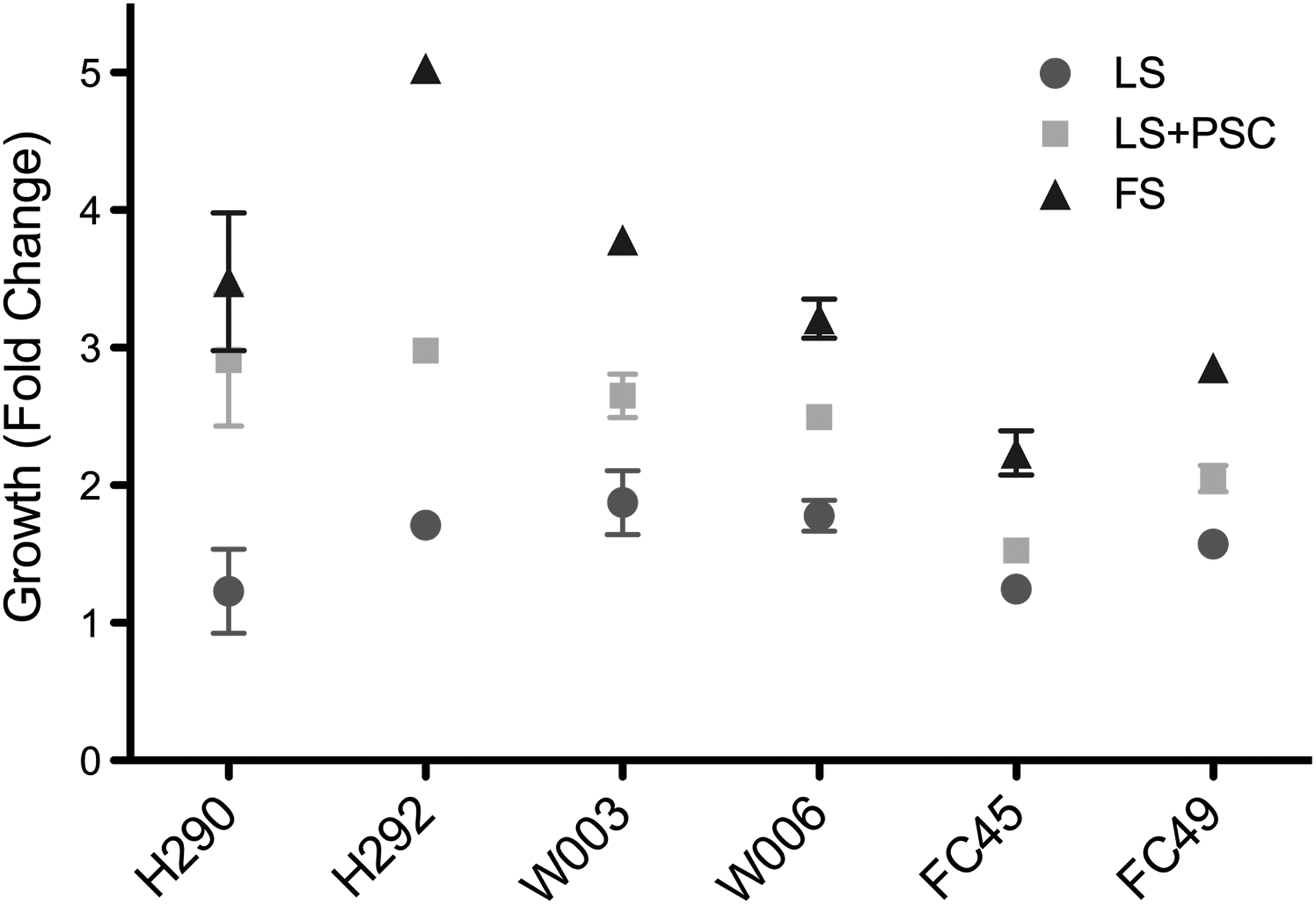
Pro-survival cocktail (PSC)-treated cells maintain metabolic activity in low serum over 5 days. Cell proliferation and metabolic activity were measured using MTS assay in triplicates. Data shown is cell growth at 5 days.
Cell characterization and differentiation
Cell characterization and differentiation potential were assessed in three of the donor BMSCs that showed proliferation advantage with PSC treatment as mentioned above. Cell morphology and phenotype were not affected by PSC treatment or LTH. Cells treated with PSC in FS for 24 h maintained characteristic cell surface markers CD29, CD44, CD73, and CD105 at the same levels as untreated controls, in both LTH and normoxia (Fig. 3A). Subsequently, the ability of PSC-treated BMSCs to differentiate into adipogenic and osteogenic cells was not altered in comparison to control cells in either LTH or normoxia (Fig. 3B). However, LTH appeared to promote osteogenesis and suppress adipogenic differentiation in the three donors tested.

PSC treatment did not alter phenotype and differentiation capability of bone marrow-derived stromal stem cells (BMSCs).
Protection against stress conditions
Serum starvation
Under serum starved stress conditions, untreated cells exhibited blebbing, poor attachment to the cell culture plastic, and anoikis (Fig. 4B). However, PSC-treated cells showed better attachment and resembled the morphology of FS control cells (Fig. 4C). Cells grown in 20% FS showed a significant increase in the rate of apoptosis when exposed to LTH, with or without PSC, compared with BMSCs grown in normoxic conditions. Since serum deprivation is known to cause apoptosis in BMSCs [10,23], apoptosis after serum deprivation was measured by AnnexinV-PI staining in different treatment groups. BMSCs cultured in NS with PSC for 72 h showed 6.1%±3.0% less apoptosis under normoxia and 19.0%±4.7% less apoptosis under LTH in three donors (Fig. 4D) compared with controls without PSC. Although there was no statistical difference between the control and PSC-treated groups in normoxia due to donor variation, the difference in LTH was significant (P<0.05). In the FS medium, LTH culture showed approximately 4% more apoptosis than normoxic control. PSC treatment in FS did not significantly affect the viability of the cells in comparison to controls in either normoxia or LTH. PSC treatment of BMSCs resulted in less apoptosis against serum starvation conditions suggesting a protective effect of the cocktail (Fig. 4D).

PSC treatment protects BMSCs against serum starvation-induced apoptosis.
Ischemic (ROS, hypoxia, and serum deprivation) challenge
To assess protection of the cells against ischemic conditions and ROS, BMSCs cultured under normoxia and LTH (ie, >7 days hypoxia) were preconditioned with PSC as described above, and were challenged with 100 μM of H2O2 and NS media for 18 h in 1% hypoxia. PSC treatment showed significantly less apoptosis based on Annexin V/PI staining for both normoxic (19.9%±3.3% less, P<0.05) and LTH (30.5%±2.9% less, P<0.01) preconditioned groups when challenged with NS, H2O2, and 1% O2 when compared with normoxia alone (Fig. 5A). LTH preconditioned cells showed a trend in decreased apoptosis when challenged with NS, H2O2, and 1% O2 compared with BMSCs grown in normoxia alone, but the difference was not significant due to the donor variability. Three out of six individual donors showed less apoptosis with LTH preconditioning alone (Fig. 5C). The apoptosis rate of long-term hypoxia+PSC (LTHP) group was slightly lower compared with the NP group indicating potential synergistic effect of LTH and PSC in combination although it did not make a significant difference after combining all six donors. The individual donor apoptosis rate of the LTHP group is lower than the NP group in five out of six donors with two donors (H292 and W006) showing significant reduction (P<0.05) and three donors (H290, W003, and FC49) showing a trend in the reduced apoptosis rate although not statistically significant according to the Bonferroni's multiple comparison test. LDH levels after ischemic challenge (NS, H2O2, and 1% O2) was higher in the LTH preconditioned group only, and it was diminished to the normoxia level with PSC treatment (Fig. 5B) indicating that PSC not only provided protection against apoptosis, but also from necrosis caused by LTH.
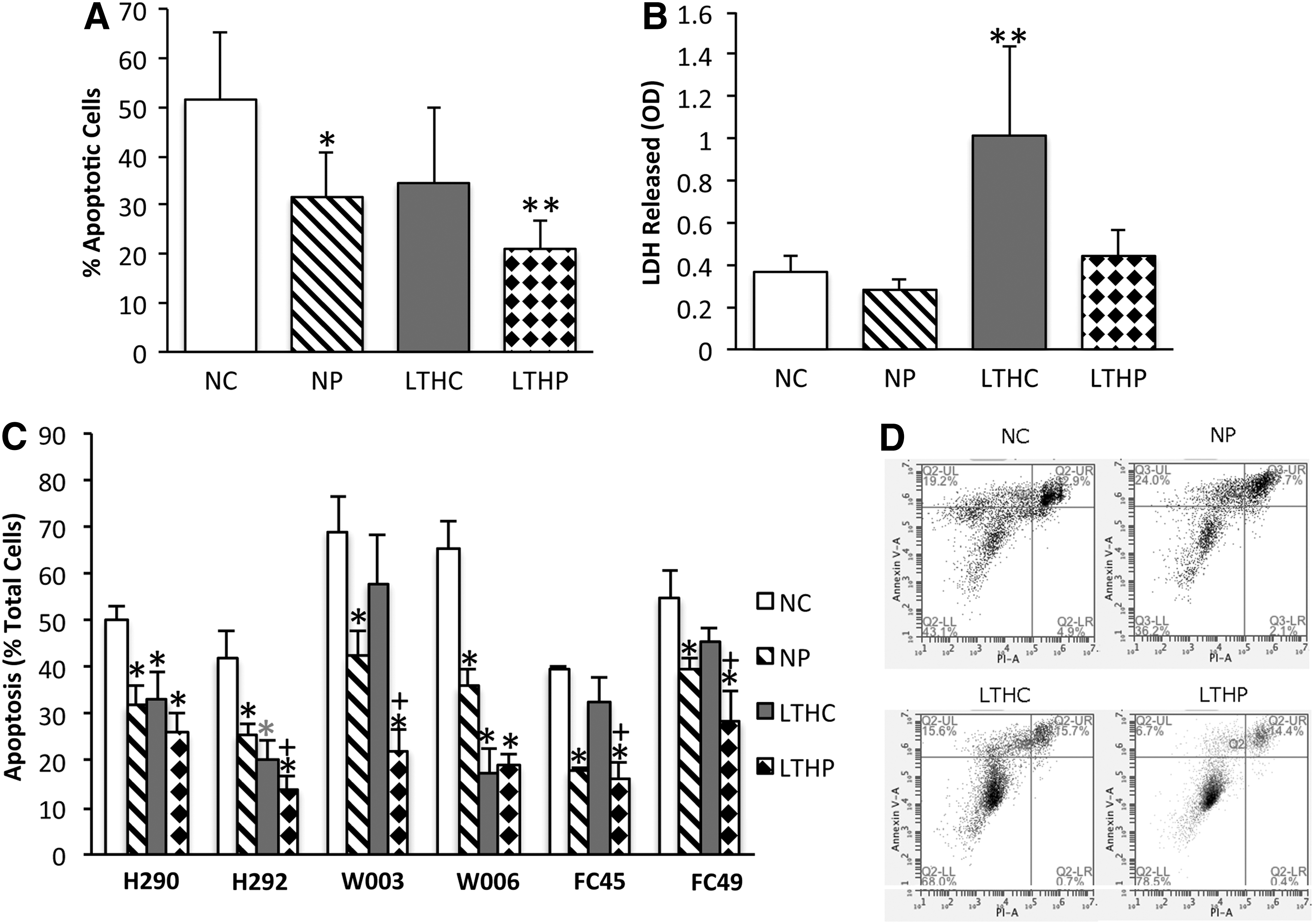
PSC treatment protects BMSCs against ROS and ischemia-induced apoptosis.
Akt activation
Akt activation is often associated with cell protection from apoptosis [24]. Akt activation has been implicated in hypoxia preconditioning [21,25,26] as well as many other growth factors [16,17,27] and small chemicals [28] providing pro-survival of BMSCs. To determine whether the protection conferred by PSC treatment or combination of LTH and PSC involved activation of Akt, western blots of phosphorylated Akt (pAkt) on Ser-473 were compared with the total Akt. Figure 6A is a representative blot showing pAkt and Akt expression levels in five donors. There were no changes in total Akt expression in all conditions tested, whereas the ratio of pAkt to total Akt increased significantly (P<0.001) in PSC-treated groups in both normoxia and LTH (Fig. 6B).
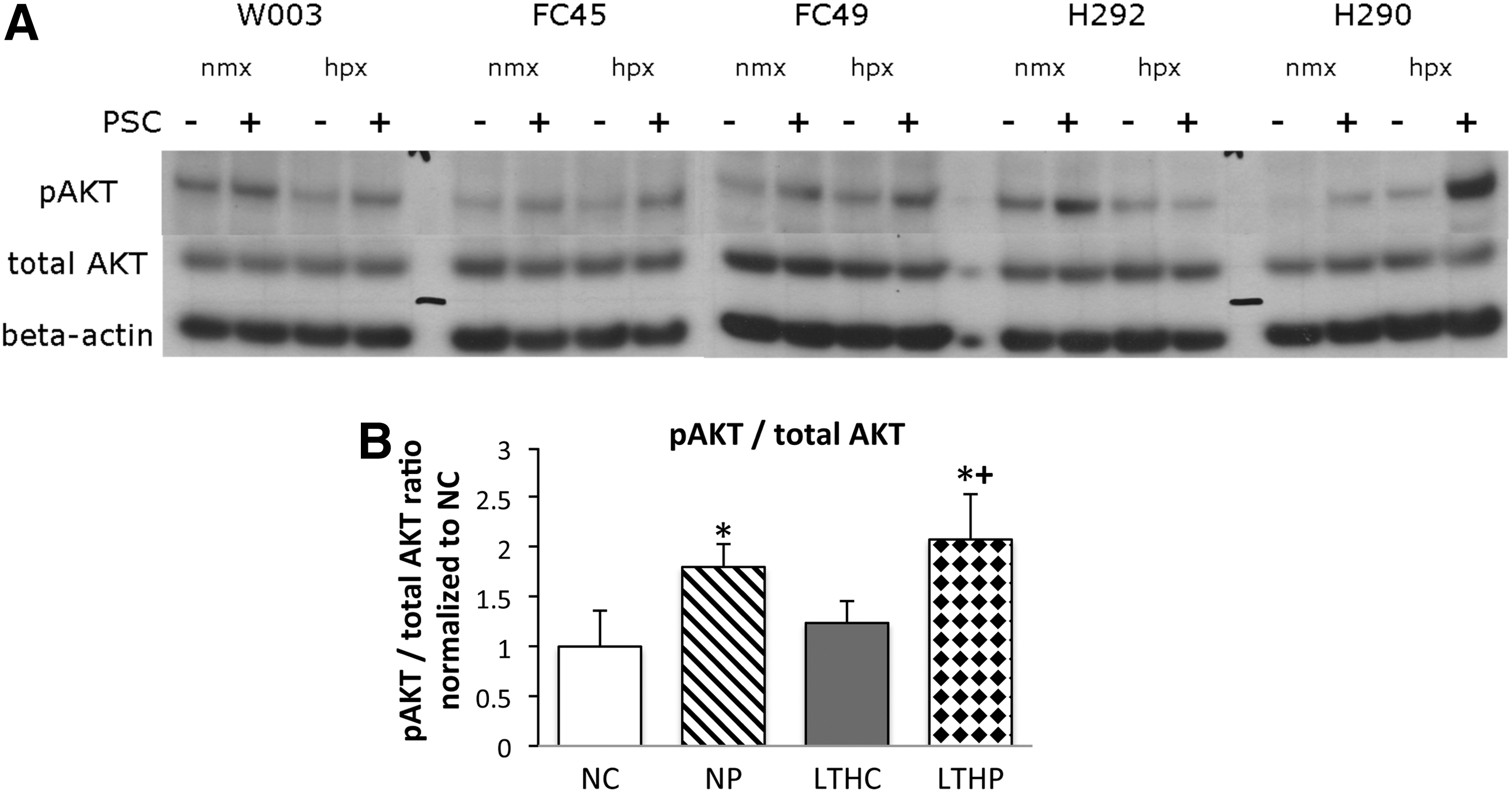
Protection conferred by PSC preconditioning is, in part, due to elevated phospho-Akt levels. Western blots were obtained from BMSCs cultured in long-term hypoxia or normoxia and pretreated with or without PSC.
Functional assays
Migration (chemotaxis)
BMSCs exposed to PSC under normoxic and LTH conditions were plated in Boyden chambers with SDF-1α, PDGF-AB, and 10% FBS gradients in the bottom wells, and the number of cells migrated for 24 h was assayed using CyQUANT. The results were normalized to a no-serum negative control gradient. There was no statistical difference in chemotaxis of the PSC-treated BMSCs toward SDF-1α (data not shown), PDGF-AB (data not shown), or FBS (Fig. 7A) by one-way ANOVA. Under normoxia or hypoxia, BMSC migration toward SDF-1α, PDGF-AB, and FBS was also unaltered with PSC treatment. Migration in response to an FBS gradient was donor dependent rather than dependent on the effects of hypoxia or PSC treatment (Fig. 7B).
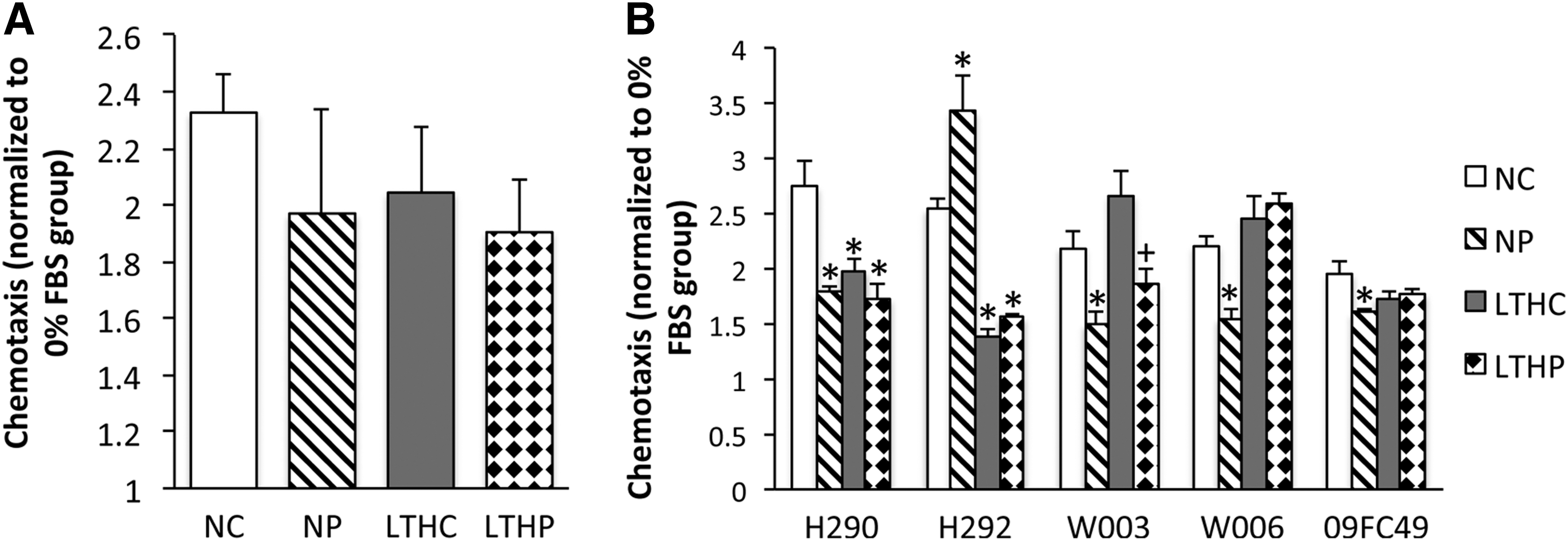
LTH or PSC treatment had no significant effect on the migratory capacity of BMSCs. Using 10% FBS as a chemotactic agent in a modified Boyden chamber, the migration of BMSCs from the upper chamber across the membrane toward the lower chamber was measured overnight by CyQUANT fluorescence. The cell migration was normalized to the control with no chemotactic agent.
Wound healing (scratch) assay
To assess the effects of PSC treatment and LTH on cell motility, confluent monolayers of BMSCs preconditioned under LTH or normoxia in the presence or absence of PSC were scratched with a 200 μL pipette tip to create artificial wounds. At the 24-h endpoint, the number of cells that had migrated into the scratched line was counted to assess cell motility. PSC treatment did not change cell motility in either oxygenation culture condition (Fig. 8A). However, the LTH condition showed a trend in increasing cell motility despite the lack of statistical differences in the combined data of all six donors. When individual donor cell motility was separately analyzed, three of the donors showed an increase in cell motility with LTH, whereas three other donors showed no difference (Fig. 8B).
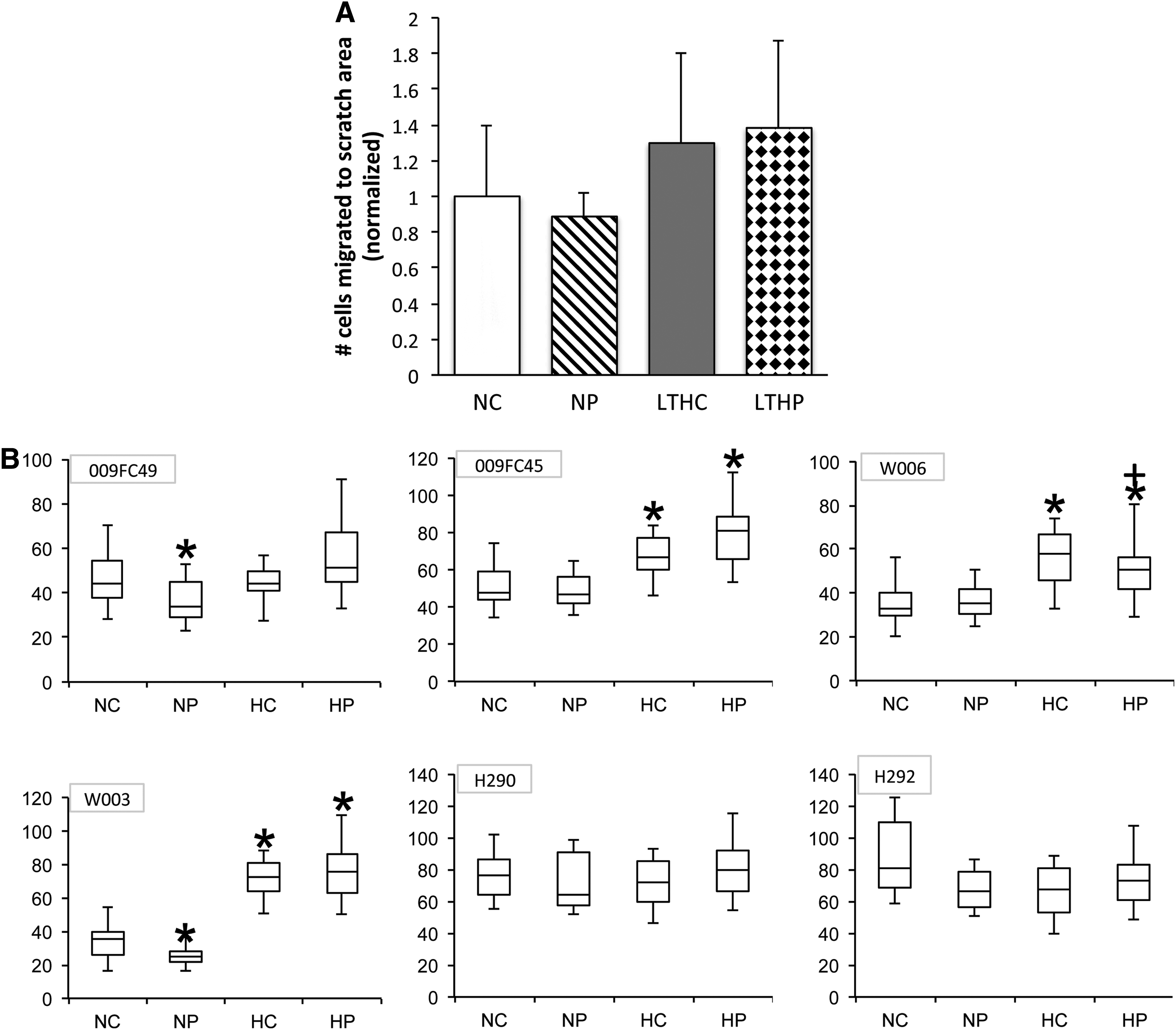
Long-term hypoxia or PSC had no effect on cell motility toward wounds. The monolayers of BMSCs grown in tissue culture plates were scratched with a pipette tip to create wound. The locations were the migration will be measured was marked with a fine-tip marker, and then photomicrographs were taken at time 0 and 24 h to count cells migrated into the scratched area.
Bio-plex cytokine/chemokine assay
The levels of growth factors and cytokines secreted by BMSCs were measured using the Bio-plex immune assay after long-term hypoxic preconditioning and PSC treatment. VEGF, monocyte chemotactic protein-1 (MCP-1/CCL2), interleukin (IL)-6, macrophage migration inhibiting factor (MIF), and stem cell growth factor-beta (SCGF-b) were the most abundant proteins detected in the BMSC-conditioned medium. Other cytokines and growth factors, including IL-1 receptor antagonist (IL-1ra), IL-4, IL-5, IL-7, IL-8, IL-9, IL-10, IL-12 (p70), IL-13, granulocyte-colony stimulating factor (G-CSF), interferon gamma (IFN-g), platelet-derived growth factor (PDGF-bb), regulated on activation, normal T cell expressed and secreted (RANTES), IL-16, HGF, monokine induced by gamma interferon (MIG), beta-nerve growth factor (b-NGF), stem cell factor (SCF), and SDF-1α were detected at low levels. PSC treatment alone did not change profiles of secreted cytokines and growth factors either in short-term hypoxia and serum-free stress conditions or wound healing stress conditions (data not shown). LTH preconditioning, before short-term hypoxia and serum-free stress conditions, downregulated IL-1ra, IL-4, IL-5, IL-8, IFG-g, MCP-1, and RANTES, while it upregulated VEGF, IL-16, HGF, MIG, b-NGF, and SDF-1α in at least three out of four donors (Table 2). In response to the wound healing stress condition, LTH preconditioning caused decreased levels of IL-1ra, IL-5, IL-9, IFN-g, MCP-1, and RANTES secretion, while it increased the levels of VEGF, MIF, b-NGF, SCF, and SCGF-b (Table 1). PSC treatment in combination with LTH induced increases in HGF, MIG, SCF, SCGF-b, and SDF-1α compared with preconditioning alone (data not shown). All other cytokines and growth factors showed donor-dependent conflicting changes in their expression levels.
LTH, long-term hypoxia; HPX, hypoxia; NS, no serum; BMSC, bone marrow-derived stromal stem cells.
Discussion
For cells to be used effectively for therapy and tissue regeneration, the early cell loss due to anoikis and or oxygen/nutrient deprivation needs to be minimized. Various strategies have been developed to improve or prolong survival of cell products for treatment of diseases. Laflamme et al. identified a cocktail of pro-survival factors that improved hESC-derived cardiomyocyte engraftment and differentiation in a rat MI model [20]. Since the physiological niche for BMSCs is hypoxic (2%–7% O2), the beneficial effects of short-term hypoxic culture (1%–3% O2) and long-term preconditioning on cell viability and proliferation rates have been evaluated [21]. Estrada et al. demonstrated that culturing adipose-derived human MSCs at a physiological level of hypoxia long-term improves the growth and genetic stability by limiting oxidative stress, DNA damage, and telomere shortening [22]. In the current study, we investigated the effects of hypoxia in combination with PSC (IGF-1, Bcl-XL, and caspase inhibitor I) as a strategy to improve BMSC survival under hostile conditions.
The original pro-survival factors described by Laflamme consist of IGF-1, Bcl-XL, caspase inhibitor ZVAD-fmk, cyclosporine A, and pinacidil in a growth factor reduced Matrigel™ [20]. We modified the PSC by eliminating the Matrigel because the BMSCs were preconditioned in vitro, and for most purposes of cell therapy in which cells are infused intravenously, the use of Matrigel would not be applicable. We also eliminated cyclosporine A and pinacidil because of their effects in vivo as an immunosuppressant and a vasodilator, respectively.
The interindividual variability in BMSCs in the current study are similar to what has been reported by Deskins et al. [29] in which cell potency was evaluated based on proliferation, growth rate, and metabolism. In the current study, one major finding was the significant variability of BMSCs derived from six donors in response to all experimental conditions (ie, serum starvation, ischemic exposure, chemotactic migration, and wound healing). These results confounded the interpretation of the data in BMSC responses to stress conditions or cell function, however, it underscores the importance of evaluating cells for potency from multiple donors when testing approaches to improved survival and/or function. Despite donor variability, the modified PSC supported BMSC proliferation and metabolism in LS over 5 days. PSC treatment demonstrated improved protection against apoptosis when BMSCs were grown in serum deprivation independent of oxygen concentrations. Although the percentage of apoptotic cells slightly increased in LTH preconditioning alone, the level of protection conferred by PSC was greater in LTH as compared with cells grown in 21% O2. When PSC and LTH preconditioned cells were challenged with H2O2 to mimic ROS exposure found in inflammation and injury, PSC treatment showed some improvements in survival (ie, decrease in apoptosis and necrosis rates). LTH preconditioning alone did not significantly reduce the level of apoptosis caused by ischemic stress (ie, 1% O2, H2O2, and serum deprivation) although there was a trend toward reduced apoptosis in three out of six donor cells examined. This is contrary to previous studies that have shown that exposure of murine BMSCs to low oxygen tension improves cell survival under ischemic conditions by priming the cells through adaptation [25 –27,30,31]. However, most of the hypoxic preconditioning was done short term (≤24 h) or cyclic, and the oxygen tension in these studies varies from 0% to 3%. Chacko et al. also have reported that rat BMSCs exposed to 0.5% O2 for 72 h showed no significant pro-survival or angiogenic protein expressions, supporting our observation in human BMSCs [26]. LTH alone also increased the necrosis rate reflected by increased LDH release, but the necrosis rate was reduced to the same level as all other groups with PSC treatment, indicating that PSC provided protection both through inhibition of apoptosis and necrosis.
The effect of PSC on BMSC migration was examined because of the possibility that in a previous study by LaFlamme in which hESC-derived cardiomyocytes transplanted with PSC and growth factor reduced Matrigel may have inhibited migration of the cells out of the infarcted area resulting in higher engraftment rates. Since the cells are expected to migrate into the injury area toward a cytokine gradient after intravenous injections [32], inhibition of migration by PSC would provide no therapeutic advantage and possibly be detrimental in cellular therapy. PSC had no effect on chemotaxis in either normoxia or LTH when cells were exposed to SDF-1α, PDGF-AB, and FBS gradients. The ability of BMSCs to migrate in response to PSC or hypoxia was highly donor specific, and the interdonor variability may have contributed to the observed lack of differences along a chemoattractant gradient.
Liu et al. have reported improved migration toward SDF-1 with 24 h of hypoxic preconditioning, which was attributed to an increase in SDF-1 receptors, CXCR4 and CXCR7 [27]. Chacko et al. reported no significant increase in CXCR4 expression after hypoxic preconditioning up to 72 h in rat BMSCs although the study did not include migration data [26]. In the current study, cells were grown under LTH (≥7 days) to simulate oxygen tensions detected in the bone marrow niche that may have contributed to the observation that there was no increased migration when exposed to chemoattractants. Moreover, differences in cell surface chemokine receptor expression levels in murine and human BMSCs may explain for the lack of increased migration [33].
Previous studies have demonstrated that hypoxia-conditioned cells have increased motility toward wounds; however, these studies were performed using either a single donor or pooled BMSCs [21,30]. BMSC motility toward wounds in response to LTH preconditioning was found to be highly donor specific and correlated with increased migration in chemotaxis assays. PSC treatment did not show any significant effect in cell motility toward wounds.
There are conflicting reports as to whether hypoxia improves stemness and differentiation potential of BMSCs. A few studies have reported decreased stemness and differentiation potential [34,35] when cells are grown in low oxygen tension, whereas others have demonstrated maintenance of stemness and unaltered differentiation capability under hypoxia [21,26,36]. It has also been suggested that BMSC stemness under various conditions may be defined as the ability to differentiate in vivo and support hematopoietic elements [37]. Superparamagnetic iron oxide nanoparticle labeling of BMSCs does not affect their stemness [38]. In the current study, it was evident that BMSCs had a significantly greater proliferation rate when grown in low oxygen tension compared with normoxia after an initial lag phase. BMSCs grown under hypoxic conditions maintained normal morphology, surface markers, and differentiation potential toward the osteogenic lineage as compared with cells grown in normoxic conditions. We observed a decreased adipogenic potential in hypoxia-grown BMSCs compared with the normoxia-grown cells in all donor populations. However, further studies on differentiation potential and stemness would require in vivo transplantation and BMSCs.
Akt is involved in one of the major survival pathways, PI3K/Akt/mTOR, and the activation of Akt promotes survival and growth [24]. The decision to investigate the pAKT pathway was because it was the common pathway indicated in both in hypoxia and PSC treatment although multiple cell survival pathways involving molecules such as HIF-1α, ERK1/2, survivin, and Bcl-2 are implicated [39]. Pharmacological activation of Akt through IGF-1, FGF-2 [18], and VEGF [17,25] has been reported to improve survival. Hypoxic preconditioning has also been reported to activate the PI3K/Akt pathway and promote survival of BMSCs [21,26,27,31]. We observed that pAkt was increased with PSC treatment, but not with LTH. The lack of increased Akt activation in LTH-cultured cells was reflected in the apoptosis assay, where the hypoxia alone group showed no statistical difference in protection against ischemic injury (ie, NS, H2O2, and 1% O2), although there was a trend toward decreased apoptosis. Kim et al. showed that anoxic exposure of rat BMSCs to 10-min cycles increased pAKT up to three cycles; however, when 30-min cycles were used, the pAKT level stayed up till cycle 2, then decreased on cycle 3 [31]. In addition, Chacko et al. demonstrated that pAKT was upregulated after 24 h of exposure to hypoxia and stayed the same level up to 72 h, but without a dose response [26]. Contrary to the BMSC survival advantage of short-term hypoxic preconditioning, LTH culture does not protect against apoptosis and cell death in response to ischemic injury.
The changes in paracrine activity of BMSCs after LTH preconditioning were highly donor dependent. Cytokines previously reported to be upregulated with hypoxia, such as VEGF [25,26,30], HGF [21], and SDF-1α [40] were upregulated in at least three out of four donors after LTH. However, the expression levels and changes in the levels of cytokines in response to stress conditions were vastly donor dependent, making analysis difficult. Increased secretion of VEGF and HGF can explain the trend of increased cell motility observed during wound healing, but no direct relationship between a specific donor's cell motility and their VEGF or HGF expression levels was found. A variety of anti- and proinflammatory cytokines and chemokines were downregulated with LTH preconditioning, but since these cytokines work together in complex relationships, it is difficult to interpret the data.
The unique design of the current study was to use three to six different human donor cell lines to assess cellular protection against a variety of stress factors. Most studies using human BMSCs only use cells derived from one donor or pooled cells, therefore ignoring donor differences. Moreover, many studies use rodent BMSCs despite the species differences in some cell surface chemokine receptor expressions [33] and immortalization problem through the culture selection of rapidly expanding cells from bone marrow [41]. Strain differences in inbred mouse BMSCs on growth characteristics and potency also have been reported [42]. It is difficult to predict outcomes in human clinical trials based on the translational work done with rodent BMSCs given these species differences. Although donor variability in these types of assays makes the analysis difficult, it is important to document the range of donor differences and acknowledge that not all cells from different donors will respond similarly to a particular treatment to enhance survival and engraftment. Almost all assays performed showed donor differences: migration, wound healing, proliferation, and survival following a variety of stress conditions, differentiation capability, and Akt activation. Based on these donor differences, development of assays that enable measurement of responsiveness that could reflect the success of cell therapy may be useful.
Footnotes
Acknowledgments
This research was supported by the intramural research program of the Clinical Center and National Institute of Biomedical Imaging and Bioengineering at the National Institutes of Health. We thank Dr. Pamela Robey for critical discussions and manuscript review. BMSCs were supplied by Dr. Pamela Robey and Dr. Marianna Sabatino at the Center for Bone Marrow Stromal Cell Transplantation at the NIH.
Author Disclosure Statement
No competing financial interests exist.