
Abstract
Putative cardiac progenitor cells (CPCs) have been identified in the myocardium and are regarded as promising candidates for cardiac cell-based therapies. Although two distinct populations of CPCs reached the clinical setting, more detailed studies are required to portray the optimal cell type and therapeutic setting to drive robust cell engraftment and cardiomyogenesis after injury. Owing to the scarcity of the CPCs and the need for reproducibility, the generation of faithful cellular models would facilitate this scrutiny. Here, we evaluate whether immortalized Lin−Sca-1+ CPCs (iCPCSca-1) represent their native-cell counterpart, thereby constituting a robust in vitro model system for standardized investigation in the cardiac field. iCPCSca-1 were established in vitro as plastic adherent cells endowed with robust self-renewal capacity while preserving a stable phenotype in long-term culture. iCPCSca-1 differentiated into cardiomyocytic-, endothelial-, and smooth muscle-like cells when subjected to appropriate stimuli. The cell line consistently displayed features of Lin−Sca-1+ CPCs in vitro, as well as in vivo after intramyocardial delivery in the onset of myocardial infarction (MI). Transplanted iCPCSca-1 significantly attenuated the functional and anatomical alterations caused by MI while promoting neovascularization. iCPCSca-1 are further shown to engraft, establish functional connections, and differentiate in loco into cardiomyocyte- and vasculature-like cells. These data validate iCPCSca-1 as an in vitro model system for Lin−Sca-1+ progenitors and for systematic dissection of mechanisms underlying CPC subsets engraftment/differentiation in vivo. Moreover, iCPCSca-1 can be regarded as a ready-to-use CPCs source for pre-clinical bioengineering studies toward the development of novel strategies for restoration of the damaged myocardium.
Introduction
C
A clear understanding on CPCs biology is impaired by the scarcity of progenitors present within the myocardium as well as by time-consuming isolation procedures, and, importantly, by their phenotypic variability when isolated and expanded in different laboratories [23]. Consistently, progenitor cells that apparently partake in the same population can display non-comparable features. To circumvent these limitations, the aim of this study was to create a benchmark to be used as a reference for progenitor cell populations isolated from the myocardium in different laboratories on the basis of a plethora of non-specific cell-surface markers [6 –15,17,18]. For this purpose, lineage negative/Sca-1+ (Lin−Sca-1+) CPCs have been isolated from murine hearts and immortalized via telomerase catalytic subunit [murine telomerase catalytic subunit (mTERT)] overexpression. Recently, the role of telomerase activity in maintaining CPC viability and regenerative potential was demonstrated [24]. Aging leads to telomeric shortening in CPCs, thus leading to a senescent phenotype, as shown by the expression of p16INK4a. Such events have been correlated to cardiac function impairment, suggesting that CPC loss could be a main determinant in heart failure. Moreover, the overexpression of the human telomerase reverse transcriptase (TERT) catalytic subunit in various cell lines resulted in extended cellular lifespan [25 –27] without detectable changes characteristic of malignant transformation [28].
In the present study, a cell line of immortalized Sca-1+ cardiac progenitor cells (iCPCSca-1) has been specifically generated for the first time. iCPCSca-1 were extensively characterized to assess whether after immortalization these cells had preserved the hallmarks of their native-cell counterpart and the recognized capability to engraft and differentiate when transplanted in an MI murine model. Cells grew in vitro as adherent cells with a typical spindle-shape morphology and displayed robust self-renewal capacity, while preserving a CPC tri-lineage potential (ie, differentiated in CM-, endothelial-, and smooth muscle like-cells). iCPCSca-1 transplanted into the MI border zone significantly reduced the MI-induced left ventricle (LV) anatomical and functional alterations. Importantly, the cells engrafted, established functional connections, and differentiated in loco into cardiomyocyte- and vasculature-like cells, as previously shown for their native counterparts. Thus, the iCPCSca-1 line constitutes an unlimited source of Sca-1+ CPC replicates and can be used as a model system for in vitro high-throughput studies and for the dissection of the in vivo role of adult heart-resident progenitors.
Methods
Animals
C57BL/6 mice aged 6–12 weeks were used. Procedures were approved by Instituto de Biologia Molecular e Celular–Instituto de Engenharia Biomédica Animal Ethics Committee and National Direção Geral de Veterinária (permit no: 022793), and are in conformity with the European Parliament Directive 2010/63/EU. Humane endpoints were followed according to the Organization for Economic Co-operation and Development Guidance Document on the Recognition, Assessment, and Use of Clinical Signs as Humane Endpoints for Experimental Animals Used in Safety Evaluation (2000).
Mice were anesthetized by an intraperitoneal injection (ip) of medetomidine (1 mg/kg; Sededorm) and ketamine (75 mg/kg; Clorketam), and its adequacy was monitored by the pedal withdrawal reflex.
Isolation and culture of Sca-1+ CPCs
CPCs were isolated as previously described [29]: 6 week male C57BL/6 mice hearts were minced and digested using 0.25% Trypsin/ethylenediaminetetraaceticacid (EDTA) and collagenase II (1,500 U; Worthington) in phosphate-buffered saline (PBS). Tissue fragments were cultured in Dulbecco's Modified Eagle Medium (DMEM; Lonza) that was supplemented with 10% fetal calf serum (Lonza), 100 IU/mL penicillin and 100 μg/mL streptomycin, insulin-transferrin-selenium 1×, 300 ng/mL retinoic acid, 0.8 μg/mL linoleic acid, 2 mM
Establishment of iCPCSca-1
Murine TERT coding sequence, obtained from pGRN190 (kind gift from Geron Corporation), was inserted into EcoRI restriction site in pcINeo plasmid (Promega). Lin−Sca-1+ cells were transfected with pCINeo-TERT or control vector using the Calcium Phosphate Method (Promega), according to the manufacturer's instructions and incubated in G418-containing medium (Sigma-Aldrich) for at least 3 weeks. For clonal selection, immortalized cells were plated at single-cell density by limiting dilution and cultured in Sca-1+-conditioned medium. Clones derived from a single cell were expanded under G418 selection and characterized for Sca-1 expression by flow cytometry. Proliferative capacity of the established line was calculated using the formula log10 (total no./start no.)/log2.
In vivo tumorigenicity assay
The tumorigenicity of iCPCSca-1 was assayed by a heterotopic injection into syngeneic animals [30,31]. Eight-week-old C57BL/6 female mice were subjected to a subcutaneous injection of 1×106 cells [in 0.2 mL of 2% fetal bovine serum (FBS) in α-minimum essential medium (α-MEM)] of iCPCSca-1 at each shoulder pad. Animals were monitored daily for the appearance of palpable tumors. Mice were necropsied after a 5-month surveillance and the injection region as well as the heart, spleen, lung, and liver were harvested, fixed in 10% formalin neutral buffer (VWR BDH and Prolabo), and paraffin embedded.
Flow cytometry
Before flow-cytometry analysis, single-cell suspensions were labeled with the following antibodies: APC—anti-PDGFRα (Biolegend), c-Kit and Flk-1 (eBiosciences); PE—anti-CD105 (Biolegend), CD106 (Biolegend), CD44 (Immunotools), CD45 (Immunotools), CD34 (Biolegend), CD31 (BD Pharmingen), CD62L (Immunotools), CD40L (BD Pharmingen), CD90.2 (BD Pharmingen), and Stro-1 (Santa Cruz Biotech); and FITC—anti Sca-1 (eBiosciences) and CD40 (BD Pharmingen). Samples were acquired on an FACS Canto II and analyzed using FlowJo software.
Gene expression
For the reverse transcription polymerase chain reaction (RT-PCR), total RNA was extracted using Trizol Reagent (Sigma-Aldrich), reverse transcribed according to the manufacturer's instructions (Bioline) and PCR was performed using BIOTAQ DNA polymerase (Bioline) and gene-specific primers. For quantitative RT-PCR (qRT-PCR) analysis, cDNAs were synthesized using PrimeScript™ RT reagent kit following the manufacturer's instructions (Takara Bio, Inc.). qRT-PCR was performed using iQ™ Sybr® Green Supermix (Bio-Rad) and gene-specific primers. Reactions were carried out in triplicate on the iCycler iQ5 Real-Time PCR system (Bio-Rad). Values were normalized to glyceraldehyde-3-phosphate dehydrogenase housekeeping gene. Primer sequences are available on request.
Immunocytochemistry
iCPCSca-1 were fixed with 4% paraformaldehyde (PFA) or cold methanol for GATA-4 detection, permeabilized with 0.2% (cytoplasmic) or 1% Triton X-100/PBS (nuclear epitopes), and blocked for 1 h with 1% goat serum, 1% bovine serum albumin (BSA), or M.O.M.™ Immunodetection Kit (Vector Lab). Incubation with anti-Nestin (Abcam), anti-GATA-4 (Santa Cruz Biotech.), anti-Sarcomeric α-actinin (α-actinin; Sigma-Aldrich), anti-α smooth muscle actin (α-SMA; Sigma-Aldrich), or anti-von Willebrand Factor (vWF; Sigma-Aldrich) was carried out for 1–2 h at room temperature (RT). Incubation with secondary antibodies was for 1 h at RT, with the exception of M.O.M.™ Biotinylated Anti-Mouse IgG Reagent (Vector Lab) by which the manufacturer's instructions were followed. Slides were mounted in Vectashield with 4,6-diamino-2-phenylindole (DAPI; Vector Lab) and observed on the inverted fluorescence microscope Axiovert 200 (Zeiss).
iCPCSca-1 differentiation
For cardiomyogenic differentiation, cells were (i) co-cultured with neonatal CM for 7 days, (ii) cultured in the presence of transforming growth factor-β (TGF-β) for 21 days [32], or (iii) maintained using a commercial cardiomyocyte differentiation medium (Millipore) for 21 days.
For the co-culture set-up, CM were isolated from neonatal (postnatal day 1–3) C57BL/6 as previously described [33]. iCPCSca-1 were pre-stained with viable red fluorescent dye Vybrant™ DiI (Molecular Probes) before seeding onto CMs for 7 days.
For endothelial differentiation, iCPCSca-1 were pre-conditioned for 10 days in Endothelial Growth Medium with EGM BulletKit (EGM; Lonza) or in α-MEM (Gibco) with 10% FBS as control. After this period, cells were plated for immunocytochemistry (ICC), processed for CD31 flow-cytometry staining, or seeded onto Matrigel Growth Factor-Reduced Matrix (BD Biosciences). After overnight culture, cells were fixed and stained for vWF (PBS/0.05% Tween-20/1% BSA for 4 h at RT) or for calcein AM (BD Biosciences). Briefly, cells were incubated for 30 min at 37°C in calcein/PBS and imaged using the inverted fluorescence microscope Axiovert 200 (Zeiss). To evaluate tube formation, number of branch points and tube length were assessed using ImageJ1.42.
For smooth muscle cell differentiation, iCPCSca-1 were cultured in DMEM high glucose containing 2% FBS and 50 ng/mL of platelet-derived growth factor BB (PDGF-BB; R&D Systems) for 10 days, as described earlier [7].
MI, iCPCSca-1 delivery, and echocardiography
MI was induced by permanent ligation of left anterior descending (LAD) coronary artery as described elsewhere [34] with minor alterations. After anesthesia, female C57BL/6 were intubated and ventilated using a small-animal respirator (Minivent 845; Harvard Apparatus). The heart was exposed (Ø 5–7 mm) via left thoracotomy on the third intercostal space, and the pericardial sac was gently disrupted. A non-absorbable 7-0 suture (Silkam®; B. Braun) was used for ligation. Immediately afterward, 5×105 iCPCSca-1 (n=8) or vehicle (0.5% BSA/PBS) only (n=9) were delivered by four intramyocardial injections (5 μL each) with a 30-gauge syringe (Hamilton company). Thoracic incision was closed using an absorbable 6-0 suture (Safil®; B. Braun), and surgical staples were used for skin closure. Anesthesia was reverted by atipamezole (ip, 5 mg/kg; Revertor). Analgesia and fluid therapy were performed by ip delivery of butorphanol (1 mg/kg; Butador) and a subcutaneous injection of 5% glucose physiological saline, respectively. This procedure was repeated every 12 h for approximately 72 h post-surgery or until full recovery.
Echocardiographic assessment
Transthoracic echocardiography was performed at 2 weeks after LAD coronary artery ligation using a portable ultrasound apparatus (GE Vivid I; General Electric) that was equipped with a 12 MHz linear probe (GE 12L-RS Linear Array Transducer; General Electric). To evaluate LV structural changes, several parameters from M-mode were measured, that is, the LV internal diameter at diastole (LVIDd) and at systole (LVIDs), the interventricular septum at diastole and at systole, the LV posterior wall at diastole and at systole (LVPWs), and heart rate. Left ventricular ejection fraction (EF) and fraction shortening (FS) were calculated as an index of systolic function: FS (%)=[(LVIDd−LVIDs)/LVIDd] ×100 and EF (%)=[(LVIDd3–LVIDs3)/LVIDd3] ×100.
Histological procedures
Hearts were harvested at 2 weeks post-surgery after an injection with potassium chloride and fixed in 10% Formalin neutral buffer (VWR BDH and Prolabo) for approximately 24 h before paraffin embedding. Representative LV sampling (∼12 sections at 3 μm) was obtained by transverse sectioning from apex to base with 300 μm intervals.
Masson's Trichrome (MT) staining was performed using the Trichrome (Masson) Stain kit (Sigma-Aldrich) with the following modifications: Nuclei were pre-stained with Celestine Blue solution following Gill's Hematoxylin staining, and tissue was incubated for 1 h in Bouin's solution before muscle staining with Biebrich Scarlet-Acid Fuchsin. Sections were photographed with a stereomicroscope (Leica) before morphometric analysis.
MI size calculation and morphometric analysis
Infarct size measurement was based on collagen deposition in the ischemic LV wall 2 weeks post-infarction and calculated by the area method using the semi-automated program MIQuant [35]. The thickness of LV-free wall was measured on the infarction region using ImageJ1.42 on MT-stained sections.
Fluorescence in situ hybridization
Sections were subjected to antigen retrieval using 10 mM sodium citrate Buffer (pH 6; Sigma-Aldrich) at 98°C for 10 min followed by 0.01% acid pepsin (Sigma-Aldrich) at 37°C for 10 min. For fluorescence in situ hybridization (FISH), samples were dehydrated and incubated with a specific probe to whole mouse Y-chromosome (Y-Chr; Cambio) for 5 min at 82°C and overnight at 37°C. Samples were incubated with Streptavidin (Invitrogen) for 30 min, mounted in Vectashield with DAPI (Vector Lab), and observed on the inverted fluorescence microscope Axiovert 200 (Zeiss).
For α-actinin and CD31 (Santa Cruz Biotech.) immunostaining combined with FISH (immuno-FISH), antigen retrieval was achieved by pronase treatment for 30 min at 37°C or with 10 mM Tris/1 mM EDTA (pH 9.0) at 98°C for 30 min. Samples were permeabilized with 0.2% Triton X-100 and blocked using 4% FBS and 1% BSA or M.O.M Immunodetection Kit (Vector Lab). Incubation with anti-Connexin43 (Cx43; Abcam), anti-α-SMA(Sigma-Aldrich) and anti-CD31 was performed at RT for 1–2 h and overnight at 4°C for anti-α-actinin (Sigma-Aldrich). After a 1 h incubation with the secondary antibody (Invitrogen), slides were rinsed in 50 mM MgCl2/PBS for 5 min and post-fixed with 4% PFA and 50 mM MgCl2/PBS for 10 minutes. Samples were dehydrated and incubated with Y-Chr probe as described earlier.
Blood vessel density quantification
After antigen retrieval using 10 mM Tris/1 mM EDTA (pH 9.0; Sigma-Aldrich) at 98°C for 30 min, samples were permeabilized with 0.2% Triton X-100 and blocked for 1 h with 4% FBS and 1% BSA. Slides were incubated with anti-CD31 for 1–2 h at RT, 1 h incubated with secondary antibody and mounted in Vectashield with DAPI. Images were captured using MosaiX (AxioVision modules; Carl Zeiss). CD31+ cells were counted in 30 fields per heart over 3–5 sections at infarcted and border zone regions along the heart's long axis (n=9 control group, n=8 iCPCSca-1) using ImageJ1.42. Density was calculated as CD31-positive cells per square millimeter (mm2).
Data and statistical analysis
Statistical significance was evaluated with SPSS v.19.0 using Mann–Whitney or one-way analysis of variance with post hoc Tukey's test. Values are presented as mean±standard error of the mean. P<0.05 was considered statistically significant. *P<0.05, **P<0.01, and ***P<0.001.
Results
Generation of an immortalized line representative of Lin−Sca-1+ adult CPCs
To immortalize Lin−Sca-1+ CPCs by inducing ectopic expression of mTERT, cells expressing Sca-1 were isolated from murine hearts and transfected with pCINeo vector encoding the mTERT.
Further analysis was restricted to one transfected clone of myocardium-resident Sca-1+ progenitors, that is, the iCPCSca-1. The remaining G418-resistent isolated clones were subjected to transcriptional profiling (Supplementary Table S1; Supplementary Data are available online at
iCPCSca-1 grew to confluence as a monolayer of spindle-shaped adherent cells similar to their native counterparts (Fig. 1A) and presented a doubling time of ∼21.6 h. iCPCSca-1 were continuously sub-cultured for more than 24 months with no signs of differentiation and maintained high levels of Sca-1 expression. Conversely, primary (non-transfected) Lin−Sca-1+ CPC decreased Sca-1 expression after 1 month in culture and senesced after 15–20 passages (Supplementary Fig. S1).
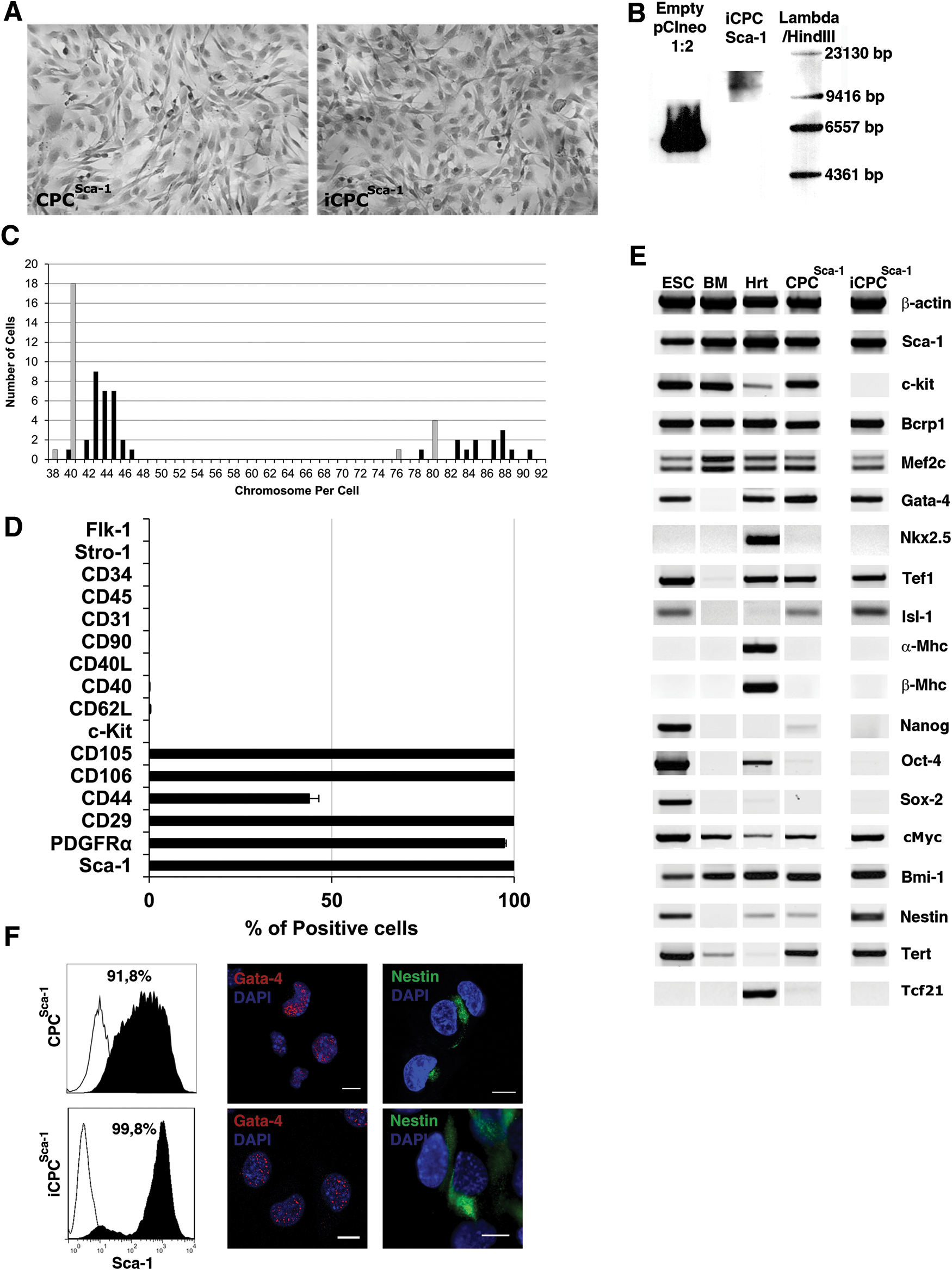
Generation and characterization of immortalized Lin−Sca-1+ cardiac progenitors (iCPCSca-1) cell line.
Genome integration of mTERT was confirmed by Southern blotting using a specific probe for the neomycin phosphotransferase gene (Fig. 1B). Chromosome number per metaphase plate of iCPCSca-1 was compared with Sca-1+ primary cells; the majority of the cells were diploid and displayed a median chromosome number of 44 and 40 in iCPCSca-1 and primary culture, respectively (Fig. 1C). In addition, tetraploid cells were present in both conditions. The tumorigenic potential of iCPCSca-1 was evaluated by means of a classical tumorigenesis assay in which syngeneic animals are used as recipients for heterotopic delivery of the cell inocula [30]. Hence, iCPCSca-1 were subcutaneously delivered to the shoulder pads of syngeneic animals that were daily monitored for tumor progression. No local mass formation was observed after a 5-month inspection period and normal histological appearance was verified for the heart, spleen, lungs, and liver (Supplementary Fig. S2).
iCPCSca-1 display a typical tri-lineage potential and a stable phenotype after long-term in vitro culture
Prospective identification of bona fide CPCs based on the unique expression of surface markers is at the present not possible. Hence, it is generally accepted that an accredited CPC will meet the following criteria: (i) derive from the adult heart; (ii) display either Sca-1 or c-Kit at their surface and lack markers of hematopoietic and endothelial commitment/differentiation; (iii) express cardiac-affiliated transcription factors while lacking mature proteins; and (iv) differentiate both in vitro and in vivo into CM, endothelial cells, and smooth muscle cells. iCPCSca-1 were, thus, characterized with regard to the cell surface phenotype, transcriptional profile, and functional properties in order to verify whether this cell line meets the premises mentioned earlier.
Flow cytometry analysis indicated a phenotype consistent with adult Lin−Sca-1+ CPCs, that is, a population constituted of cells with high Sca-1 levels (99.8%±0.21%), lacking hematopoietic (Stro-1, CD45, CD34) and endothelial (Flk-1, CD31 and CD34) markers, and expressing mesenchymal-associated proteins (eg, CD29, CD44, CD105, CD106, and PDGFRα), several of which are critical to cell migration, adhesion, and communication (Fig. 1D). Furthermore, the characterized clone (Clone No. 3, Supplementary Table S1) did not express c-Kit, a protein that singly or together with Sca-1 has been used to isolate CPCs. A detailed transcriptional profile was carried out in parallel with primary Sca-1+ CPC (CPCSca-1), embryonic stem cells, bone marrow, and heart as controls. The side-by-side transcriptional analysis demonstrated that iCPCSca-1 and primary Sca-1+ cells consistently expressed stemness-related genes (Bcrp1, Bmi-1, and Nestin) and early cardiac transcription factors (Gata-4, Isl-1, Mef2c, and Tef-1) while lacking transcripts for mature contractile myofilaments (α- and β-Mhc), fibroblasts [transcription factor 21 (Tcf21)], and regulators of pluripotency (Sox2, Nanog, Oct4), thus corroborating that iCPCSca-1 preserves the phenotype of cardiac-affiliated Sca-1+ multipotent progenitors (Fig. 1E). It should be noted that both primary and immortalized CPCSca-1 expressed c-Myc, a transcription factor known to block differentiation in distinct model systems [36] and particularly shown to synergize with Pim-1 to induce the proliferation and survival of CPCs [37]. Overall, the data demonstrated that the immortalization of the cells did not alter the transcriptional profile of Sca-1+ CPCs. The only obvious difference between the established line and the native counterparts is the lack of c-Kit in the immortalized cells, as a result of clone selection. Flow cytometry analysis clearly exposed the enrichment for the Sca-1-expressing fraction in CPCSca-1 (91.8%) and iCPCSca-1 (99.8%). Moreover, likely as a result of clonal selection, the iCPCSca-1 displayed a stronger and less disperse pattern of Sca-1 expression as compared with primary cells. Proper sub-cellular allocation of stemness-associated proteins GATA-4 and Nestin was further confirmed by ICC for both primary and immortalized cells (Fig. 1F).
To test iCPCSca-1 multipotency, distinct sub-passages were cultured under cell differentiation conditions. Cardiomyocytic differentiation potential was evaluated by co-culture with mouse neonatal CM. iCPCSca-1 were pre-stained with viable red fluorescent dye Vybrant DiI to enable detection in the co-culture system. After a week, iCPCSca-1 up-regulated GATA-4, displayed a functional pattern of Cx43 and a typical myofibrillar assembly of sarcomeric α-actinin (α-actinin) (Fig. 2A). Although the gold-standard protocol to trigger CPCs differentiation is the direct co-culture with neonatal CM [33], this assay is prone to misleading interpretations due to spontaneous cell fusion events [38,39]. Hence, we further addressed iCPCSca-1 differentiation in CM by culturing the cell line for 21 days in cardiomyocytic differentiation medium (Millipore) or in the presence of TGF-β. In this condition, iCPCSca-1 were also able to up-regulate α-actinin expression when compared with cells subjected to basal, that is, cell-maintenance, conditions for the same time period (Fig. 2B). Furthermore, when iCPCSca-1 were transduced with a green fluorescent protein (GFP) reporter under the cardiac troponin T (cTnT) promoter and subjected to TGF-β [32] for 45 days, approximately 97% of cells displayed GFP expression (Supplementary Fig. S3). Since cTnT and α-actinin are also expressed in the developing skeletal muscle, we evaluated the possibility of iCPCSca-1 having a skeletal origin. The latter was clearly excluded by demonstrating the up-regulation of cardiac myosin heavy chain beta expression after differentiation, while no detectable expression of myogenic differentiation 1(MyoD), a marker of skeletal commitment, was found (Fig. 2B). Endothelial differentiation potential was assessed by priming iCPCSca-1 with EGM medium for 10 days followed by a classical Matrigel assay to evaluate the capacity of the cells to form tubular-like structures. Primed iCPCSca-1 seeded onto Matrigel exhibited tube-like interconnected structures, similar to the capillary assembly of human umbilical vein endothelial cells (HUVECs) (Fig. 2C, n=6), whereas unprimed iCPCSca-1 (iCPCSca-1 pre-cultured in α-MEM) were unable to assemble a fully organized capillary network (n=6). In fact, the tubular complex formed by unprimed cells presented a significantly smaller number of branch points (P<0.001) and tube length (P<0.05 and P<0.01) as compared with that assembled by EGM-primed iCPCSca-1 and HUVECs (Fig. 2C). An endothelial phenotype was further addressed by analyzing vWF and CD31 protein expression. vWF was only detected in iCPCSca-1 cells after EGM priming (Fig. 2C). Accordingly, CD31 was increased by 1.75% on primed cells when compared with iCPCSca-1 maintained in α-MEM for the same period (Fig. 2C).
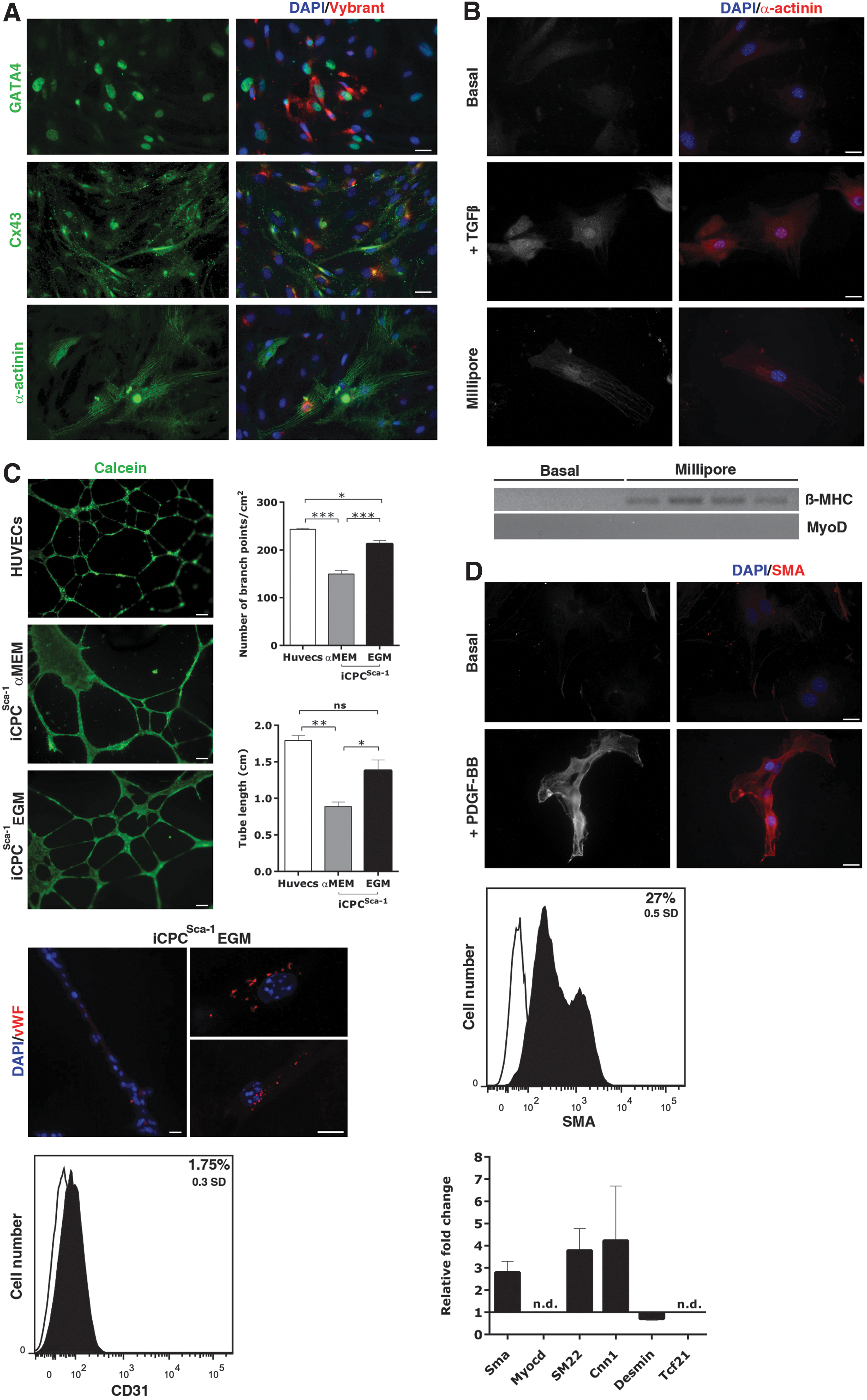
iCPCSca-1 in vitro differentiation potential.
The capacity to differentiate into smooth muscle cells was evaluated by subjecting iCPCSca-1 to PDGF-BB for 10 days. After this period, iCPCSca-1 became elongated and displayed α-SMA protein expression, as demonstrated by ICC and flow cytometry (27%) (Fig. 2D). In addition, the expression levels of the smooth muscle-affiliated genes, α-SMA, Myocardin (Myocd), Transgelin (SM22), Calponin 1 (Cnn1), and Desmin were also assessed by qRT-PCR (Fig. 2D). After 10 days of PDGF-BB treatment, iCPCSca-1 up-regulated α-SMA, SM22, and Cnn1; while no clear alteration was observed for Desmin mRNA levels. In addition, no detectable expression of Myocd was found, even on PDGF-BB stimulation. Since some of the proteins mentioned earlier are also displayed by fibroblasts/myofibroblasts, expression of the fibroblast marker Tcf21 was evaluated in order to discard a fibroblastic origin of the iCPCSca-1. Detectable expression of Tcf21 was not found, neither in basal conditions nor after PDGF-BB treatment (Figs. 1E and 2D). Overall, these results demonstrate that iCPCSca-1 were not derived from cardiac fibroblasts and that, after PDGF-BB stimulation, cells undergo an incomplete smooth-muscle differentiation program as the levels of Myocd and Desmin have not changed after PDGF-BB treatment.
Taken together, the data demonstrate that iCPCSca-1 preserved the in vitro phenotype and functional ability to moderately differentiate as described by our team [29,33] and others [5,12,13,16] for the native Lin−Sca-1+ counterparts.
iCPCSca-1-transplanted hearts show improved systolic function and lessened LV remodeling after MI
The beneficial effect of Lin−Sca-1+ CPCs administration into the MI heart has been clearly established. Notably, CPCs are able to improve cardiac function and attenuate LV remodeling after MI. However, the in vivo differentiation of the CPCs in CM and endothelial cells is minimal, and the amelioration observed on MI is primarily attributed to paracrine effects [5]. Aiming at demonstrating that iCPCSca-1 display the hallmarks of CPCs in the myocardium in the onset of injury, a proof-of-principle experiment was performed using a commonly reported experimental injury setting [16,40]. Female C57BL/6 mice subjected to MI via LAD coronary artery ligation were immediately injected on the peri-infarct region with either vehicle medium (n=9) or male-derived iCPCSca-1 (n=8). MI size measurement (based on collagen deposition) at 14 days post-infarction showed that MI extent was comparable between both experimental groups with 52% of the LV wall being affected (Fig. 3A). Notwithstanding, echocardiography at 14 days, post-surgery in animals injected with iCPCSca-1 showed improved LV function when compared with vehicle controls, as demonstrated by significantly increased (P<0.05) EF and FS in iCPCSca-1-transplanted hearts (Fig. 3B).
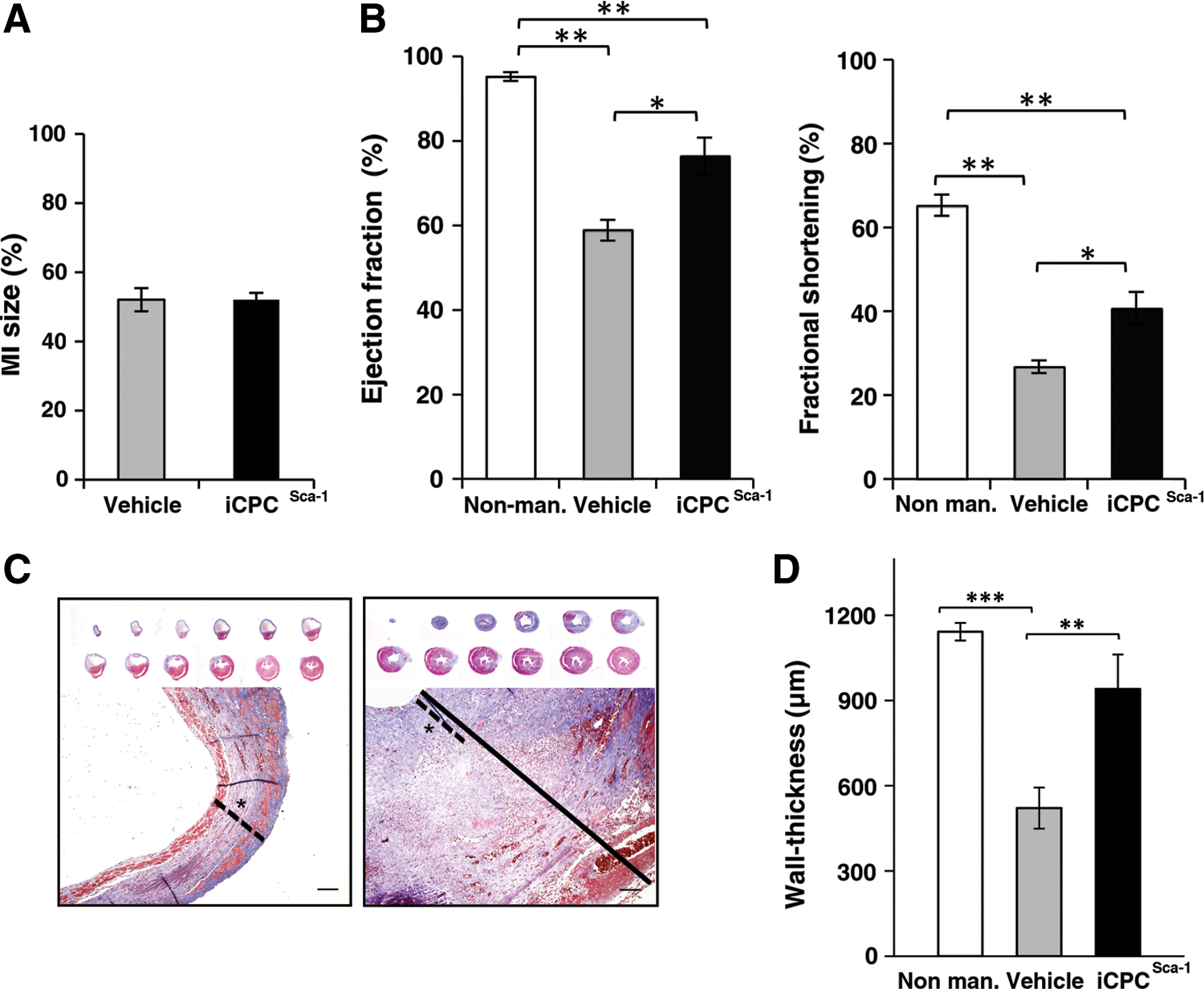
Functional and histological assessment of ischemic hearts transplanted with either iCPCSca-1 or vehicle.
Altered cardiac loading after MI due to extensive loss of myocardial cells induces changes in the heart shape, size, and function that are commonly designated as cardiac remodeling. A stereoscopic view of representative cross-sections of infarcted hearts showed that iCPCSca-1 transplantation prevented the major LV wall thinning and LV chamber expansion observed in the vehicle control group (Fig. 3C). This was corroborated by a morphometric analysis of MT stains, evidencing an increase (P<0.01) in LV wall thickness of iCPCSca-1-transplanted hearts (Fig. 3D) to a value not statistically different from the non-manipulated animals (Fig. 3D). However and despite this increase, the LV contractile function of iCPCSca-1-injected animals was not comparable with that of non-manipulated animals (P<0.01, Fig. 3B).
Overall, the data suggest that iCPCSca-1 exerted a cardio-protective effect, translated on a reduction of the deleterious consequences of ischemia and a partial recovery of cardiac functional parameters at 2 weeks post-MI.
Transplanted iCPCSca-1 improve neovascularization of the infarcted myocardium, engraft, and differentiate into CM-, endothelial-, and smooth muscle-like cells
To address whether cardiac performance amelioration after iCPCSca-1 administration occurred at least partly via stimulation of the de novo vessel formation by paracrine mechanisms, the density of small blood vessels in the infarcted myocardium and in the border zone was quantified at 2 weeks post-surgery (Fig. 4A). Overall, iCPCSca-1 transplantation resulted in a significant increase in CD31+ cells per mm2 when compared with the vehicle control (709±19 vs. 555±24, respectively, P<0.001, Fig. 4B). iCPCSca-1, identified by Y-Chr staining were consistently observed in close proximity to vessels, suggesting contribution to either endothelial precursors mobilization/proliferation and/or differentiation into endothelial cells. The latter possibility was evaluated by FISH combined with immunostaining for CD31, and, indeed, double positive events were observed in all analyzed animals although at very low frequencies (4.1±0.7 cells per transverse heart section) (Figs. 4C and 5I). In fact, the majority of small vessels were not formed by donor cells, hinting that iCPCSca-1 contribution to neovascularization is more likely to occur by stimulating the mobilization and/or proliferation of host endothelial precursors/cells (paracrine action).
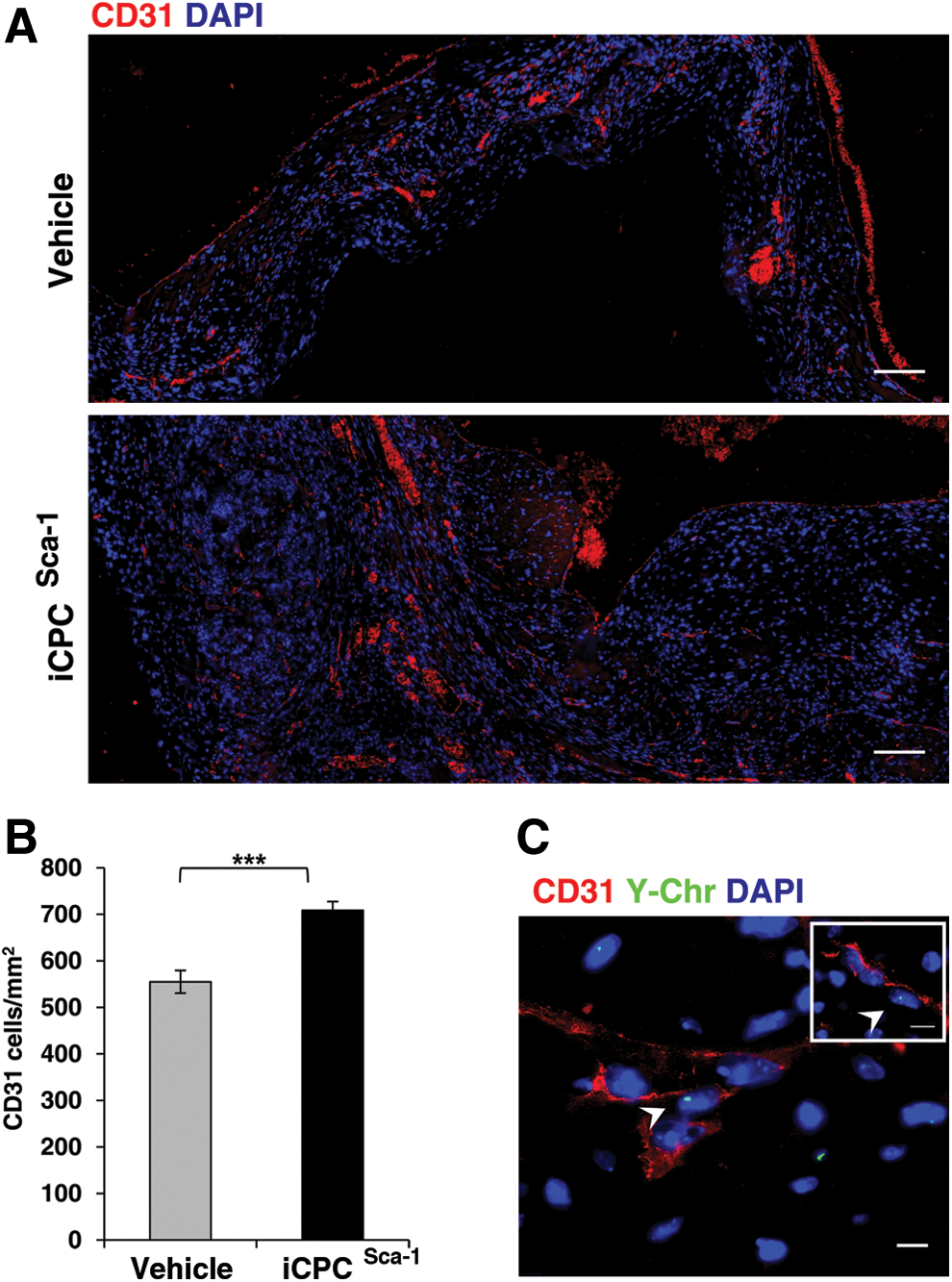
Contribution of iCPCSca-1 to neovascularization post-MI.
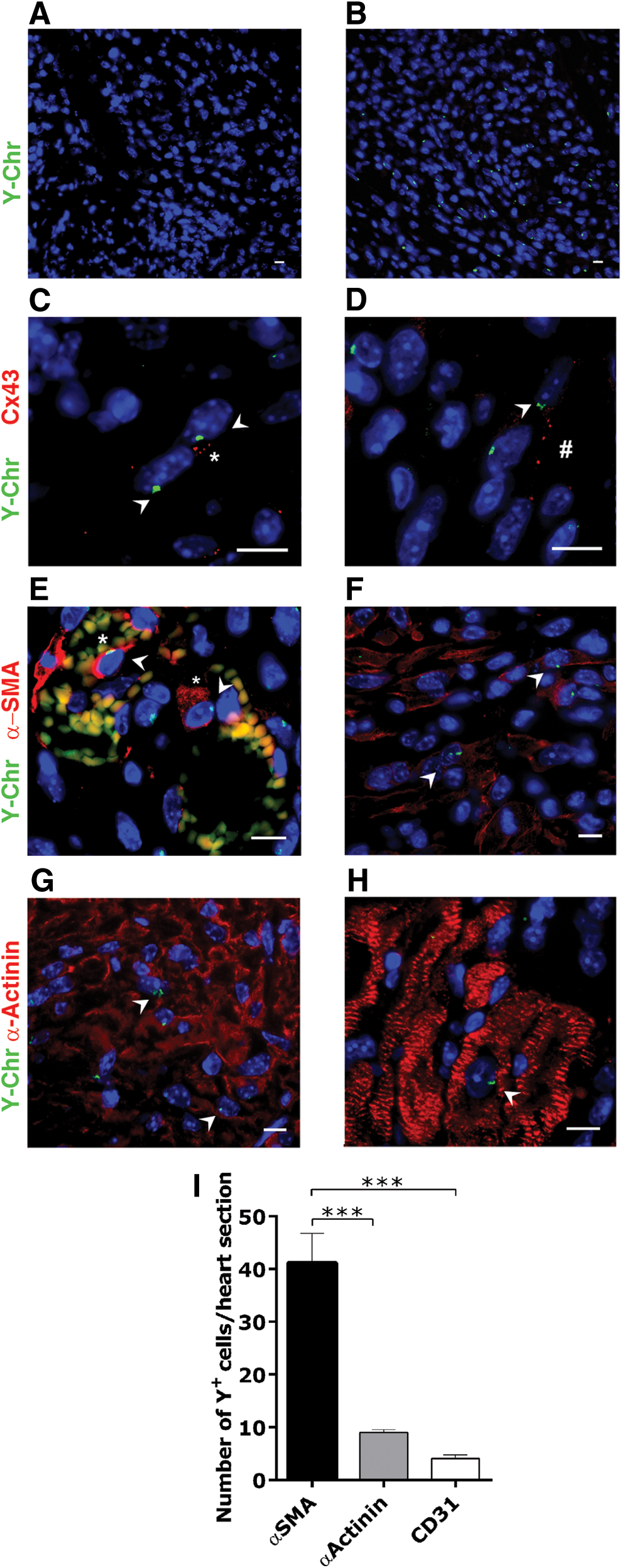
In vivo characterization of iCPCSca-1 on transplantation into MI hearts.
We next investigated iCPCSca-1 functional engraftment and contribution to the de novo cardiac cells formation. In vehicle-injected female mice, no Y-Chr+ cells were detected in the heart (Fig. 5A); whereas in transplanted animals, numerous iCPCSca-1 were observed throughout ischemic myocardium and MI border zone at 2 weeks post-infarction. This demonstrates that cells were able to survive and engraft the LV ischemic region (Fig. 5B). Moreover, iCPCSca-1 or their progeny were not found in healthy remote areas of the injury site. Ki67 staining was frequently associated with Y-Chr evidencing iCPCSca-1 proliferation within the host myocardium (data not shown). Assembly of the gap junction protein Cx43 at the cellular membrane confirmed that donor cells established functional communication to both donor and host cells (Fig. 5C, D), substantiating successful iCPCSca-1 engraftment within the host tissue.
Although the majority of grafted cells retained an undifferentiated morphology and Sca-1 expression (Supplementary Fig. S4), discrete differentiation events were observed. Co-localization of Y-Chr with α-SMA was observed in all analyzed animals (41.2±5.6 cells per transverse heart section) (Fig. 5I), either integrating small vessels (Fig. 5E) or displaying a fibroblast-like morphology (Fig. 5F). In fact, in addition to smooth muscle cells, α-SMA is also expressed by myofibroblasts, cells capable of establishing cell connections through gap junctions, and relevant players in formation of the contractile granulation tissue [41]. However, considering that Tcf21 is a marker of fibroblasts [42] and is also involved in the activation of myofibroblasts [43], and that iCPCSca-1 failed to express this marker even when specifically stimulated (Figs. 1E and 2D), there is little support for considering a myofibroblast fate. Furthermore, iCPCSca-1 contributed to the cardiomyocytic compartment as evidenced by α-actinin expression. The majority of α-actinin+ iCPCSca-1 presented an immature phenotype in three out of six analyzed animals (9.0±0.6 cells per transverse heart section) (Fig. 5I), suggesting an incomplete differentiation. Moreover, α-actinin+ iCPCSca-1 were found in close proximity with α-actinin+ host cells displaying a similar immature phenotype, suggesting an eventual iCPCSca-1—mediated recruitment/activation of endogenous precursors (Fig. 5G). Donor cells presenting well-defined cross-striations, characteristic of a more mature phenotype, were found in one animal (Fig. 5H; Supplementary Fig. S5). While we do not rule out the possibility that these scarce events resulted from cell fusion, differentiation into more mature CM seems a more consistent scenario (Supplementary Fig. S5).
Overall, these results are in agreement with the previous observations, that Lin−Sca-1+-mediated beneficial effects after MI are partially mediated by paracrine mechanisms [5,44] in detriment to a major contribution to cardiac cell types after transplantation.
Discussion
Different progenitor cell subsets have been so far suggested as candidates for the optimal cell population to be implanted in infarcted and/or failing hearts (Fig. 6), but a relationship among these populations remains undetermined. The absence of a specific molecular signature as well as the multiplicity of the isolation and characterization protocols do not enable an understanding of whether the described CPC populations constitute different subsets of a common progenitor or independent cell lineages of distinct ontogeny. Indeed, it has been suggested that the various identified CPCs partake in the same cell lineage, and the different phenotypes are a consequence of the particular culturing protocols used in different laboratories [21]. Importantly, other aspects account for an increased difficulty in the identification of CPCs and to understand their real efficiency. Namely, primary cells can easily change along time in culture as a consequence of genotypic and phenotypic drift and/or senescence [45 –47] and, thus, lead to a lack of reproducibility across studies.

Illustrative scheme of described multipotent progenitor cell populations isolated from the adult heart. Several isolation methods have been applied to separate CPCs based on the expression of specific cell-surface markers (eg, c-Kit+ and Sca-1+), in vitro functional assays (eg, cardiospheres, side population), or the expression of a specific genetic marker (Wt-1 epicardial progenitors). Regardless of the increasing number of putative CPCs reported, the functional, hierarchical, and anatomical relationship of the illustrated cell progenitor subsets is still elusive. Color images available online at
Taken together, the experience accumulated in the last decade indicates that only two strategies are used to implement a reliable cell therapy: to try and fail, that is, to implant whatever progenitor cell phenotype in the patient trusting to identify the perfect treatment, or to systematically dig the complexity of the stem/progenitor cell behavior and, based on novel data and concepts, to formulate a logically stringent procedure to transfer experimental observations to the clinical setting.
Novel tools should be developed to help understanding and, thus, governing the CPC complexity and phenotype variability and to enable the standardization of the experimental sets and the inter-laboratory comparison. Consistently, the present study has been carried out to create an immortalized CPC line retaining a constant phenotype corresponding to native features. CPC immortalization has been achieved by inducing TERT overexpression, in order to circumvent the limitations determined by the diminished mitotic capacity of somatic cells [48 –52]. The use of immortalized lines that retain the in vivo features of primary cells, by combining the advantages of a high proliferative cell source with a stable phenotype in long-term culture, is a major breakthrough in cell biology. Indeed, we have previously shown that bone marrow-derived mesenchymal stromal/stem cell (MSC) immortalized lines preserve multipotency in standard and three-dimensional conditions in vitro [49,53]. Although telomerase overexpression does not induce oncogenesis, it may provide cells with a greater opportunity to accumulate mutations, predisposing to later malignant transformation [54,55]. The iCPCSca-1 karyotype analysis showed aneuploidy, characteristic of malignant cells. However, an in vivo tumorigenicity assay [30,31], monitored for 5 months, did not reveal tumor formation either locally (subcutaneously) or in any of the analyzed organs (heart, spleen, lungs, and liver).
Consistently with adult Lin−Sca-1+ CPC, the iCPCSca-1 phenotype was characterized by the expression of high levels of Sca-1, stemness-related genes (Bcrp1, Bmi-1 and Nestin), mesenchymal-associated proteins (eg, CD29, CD44, CD105, CD106, and PDGFRα), and early cardiac transcription factors (Gata-4, Isl-1, Mef2c, and Tef-1). Conversely, immortalized cells did not express c-Kit while also lacking hematopoietic, endothelial, fibroblasts, mature contractile myofilaments (α-and β-Mhc), and pluripotency (Sox2, Nanog, and Oct4) markers.
iCPCSca-1 displayed the capability to generate in vitro cardiomyocyte-, smooth muscle-, and endothelial-like cells, with the differentiation toward smooth muscle-like cells being the most efficient. This differential in vitro potential of Sca-1+ cells has been previously demonstrated by others [16]. Whether this indicates a higher commitment of CPCSca-1 to smooth muscle-like cells or to the fact that the in vitro differentiation conditions are less favorable to activate endothelial and cardiomyocytic pathways, and, thus, not suitable to demonstrate the full potential of these cells, still needs to be determined.
iCPCSca-1 is a phenotypically stable cell line that provides investigators with the same unique cell model to dissect CPC behavior and differentiation mechanisms. Furthermore, iCPCSca-1 could represent a benchmark to compare with other cells posited to bear stemness characteristics. In fact, the immortalized CPCs retain the features of their ex vivo counterparts, that is, moderate differentiation into cardiomyocyte-, endothelial-, and smooth muscle-like cells, when subjected to appropriate stimuli in vitro as well as in vivo, in a myocardial injury setting. Furthermore, the cell line combines the advantages of a high proliferative cell source with a stable phenotype in long-term culture. Indeed, murine cells are prone to undergo spontaneous immortalization after prolonged in vitro culture [46,56,57], which may result in the accumulation of unpredictable cytogenetic modifications and malignant transformations [47]. The use of uncharacterized spontaneously immortalized cell clones as in vitro models for their native cell counterparts should be circumvented due to the dissimilar phenotypes generated by unpredictable cytogenetic alterations in overextended sub-culture in vitro.
The iCPCSca-1 capability to mimic their native non-immortalized corresponding cells has also been apparent in in vivo experiments. In fact, iCPCSca-1 were able to contribute toward repairing the myocardial injuries experimentally provoked by coronary ligation in vivo, as shown by minor LV chamber expansion and increased thickness of the LV-free wall. Importantly, echocardiography follow-up revealed improved systolic function after iCPCSca-1 transplantation into MI animals. iCPCSca-1 abundance in the host infarcted and peri-infarcted myocardium was particularly surprising, and cells were able not only to survive but also to engraft. Moreover, engrafted cells established donor–donor and donor–host Cx43-mediated connections. Indeed, Cx43 promotes MSC survival in the ischemic heart [58], and it is critical for protection against ventricular tachycardia, which is a resultant from the transplantation of embryonic CM in MI [59]. Accordingly, it can be conjectured that Cx43 is involved in iCPCSca-1 resistance to the adverse scenario of myocardial ischemia.
The implantation of immortalized cells into infarcted mouse hearts also induced a significant improvement in the vascularization of the damaged region, very likely, through a paracrine action. Since an improved vascularization is associated with a “pro-regenerative” response, the formation of new vessels in the infarcted tissue was carefully assessed. iCPCSca-1 transplanted hearts exhibited a denser capillary network, when compared with the vehicle control group. In addition, iCPCSca-1 contributed directly to de novo vessel formation, as demonstrated by CD31 and α-SMA expression on Y-Chr bearing cells. However, endothelial differentiation was modest; hence, direct contribution of engrafted cells to new vessel formation is minimal, suggesting that paracrine mechanisms are involved in the recruitment and/or proliferation of endothelial cells and/or precursors.
CPCs differentiation into CM has been frequently demonstrated by the expression of proteins of the contractile machinery [5,6,12,15]. Indeed, although CPCs appear to grasp a lot of expectations with regard to cardiac cell therapies, most studies still report an immature phenotype for the differentiated cells, to a great extent resembling fetal-neonatal CM [5,6,12,15]. In our in vivo setting, iCPCSca-1 differentiated into cardiomyocyte-like cells, though the majority displayed a disorganized structure with no detectable sarcomeres, characteristic of an immature phenotype. It should be noted that both primary CPCSca-1 and iCPCSca-1 express the transcription factor c-Myc, which has been previously shown to play a role in the blockage of cell differentiation [36] and in the survival and proliferation of CPCs [37]. Whether the levels of c-Myc are regulating CPCs cell-fate decision to differentiate at the expenses of cell proliferation remains to be determined and should be subject to further investigation. Since iCPCSca-1 showed an in vitro and in vivo differentiation potential similar to their native counterparts, it is our conviction that iCPCSca-1 constitutes a reliable easy-to-use model to dissect the key molecular mechanisms governing cardiomyocytic differentiation from adult CPCs.
In this work, an immortalized cell line for modeling mouse Lin−Sca-1+ CPCs was specifically generated by means of TERT overexpression. The immortalization did not appear to impair any important cellular property, as iCPCSca-1 were phenotypically and functionally similar to their primary cell counterparts. Overall, the results indicate that iCPCSca-1 are representative of the native Sca-1+ cardiac population, thus constituting a suitable tool for functional and mechanistic studies in need in the field. Hence, with regard to the heterogeneity and low frequency in the adult heart of described CPC populations (Fig. 6), iCPCSca-1 comes forth as a uniquely validated off-the-shelf line for in vitro high-throughput and bioengineering studies. The latter combined with the remarkable capacity of iCPCSca-1 to engraft and differentiate in loco makes this cell line a valuable model system to investigate in vivo the role of the Sca-1+ stem/progenitor cells resident in the adult heart.
Last but not the least, the widespread use of this and other similarly generated cardiac progenitor-subset cell lines will enable the reduction of animal usage and will contribute in a properly standardized manner to the definition of benchmark(s) for stem/progenitor cells from distinct organ systems and from different laboratories.
Footnotes
Acknowledgments
This work was supported by Fundação para a Ciência e a Tecnologia (SFRH/BD/64715/2009 to A.G.F., SFRH/BPD/42254/2007 to D.S.N., SFRH/BD/74218/2010 to M.V., SFRH/BPD/80588/2011 to T.P.R., and Ciência2007 to P.P.O.); Fundo Europeu de Desenvolvimento Regional, Programa Operacional Factores de Competitividade-COMPETE, Quadro de Referência Estratégico Nacional, and Fundo Social Europeu (PEst-C/SAU/LA0002/2011, PTDC/SAU-OSM-68473/2006, PTDC/SAU-ORG/118297/2010 and NORTE-07-0124-FEDER-000005 to D.S.N.); and European Regional Development Fund – Project FNUSA-ICRC (no. CZ.1.05/1.1.00/02.0123). Travel funds were awarded by Boehringer Ingelheim Fonds to (T.P.R.); by American Portuguese Biomedical Research Fund to (P.P.O and D.S.N.).
The authors are indebted to Joana G. Guedes (CIBIO), Rui Fernandes (IBMC), Paula Sampaio (IBMC), Dina Leitão (IPATIMUP), and the members of MCA Lab (IBMC) for sharing scientific expertise and equipment.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.