
Abstract
Somatic cells can be reprogrammed to induced pluripotent stem cells (iPSCs) by defined factors. However, substantial cell numbers subjected to iPSC induction stray from the main reprogramming route and are immortalized as partial iPSCs. These partial iPSCs can become genuine iPSCs by exposure to the ground state condition. However, such conversion is only possible for mouse partial iPSCs, and it is not applicable to human cells. Moreover, the molecular basis of this conversion is completely unknown. Therefore, we performed genome-wide screening with a piggyBac vector to identify genes involved in conversion from partial to genuine iPSCs. This screening led to identification of Cnot2, one of the core components of the Ccr4-Not complex. Subsequent analyses revealed that other core components, Cnot1 and Cnot3, also contributed to the conversion. Thus, our data have uncovered a novel role of core components of the Ccr4-Not complex as regulators of transition from partial to genuine iPSCs.
Introduction
S
Here, we performed genome-wide screening with a piggyBac vector to identify genes that participate in transition from partial to genuine iPSCs. Our screening identified the gene encoding Cnot2, one of the core components of the Ccr4-Not complex [9,10]. Our knockdown studies suggested that all of the core components of the complex, that is, Cnot1, Cnot2, and Cnot3, as well as Trim28, which shares genomic binding sites with Cnot3 [11], equally contribute to this transition. Analyses of genes with altered expression as a result of the forced expression in partial iPSCs indicated that the major role of these factors is suppression of gene expression associated with developmental processes. Thus, our screening identified core components of the Ccr4-Not complex and Trim28 as new players that contribute to the transition of abnormally reprogrammed partial iPSCs to genuine iPSCs.
Materials and Methods
iPSC induction
Mouse embryonic fibroblasts (MEFs) were prepared from 13.5 dpc embryos carrying a GFP reporter gene whose expression faithfully recapitulated Nanog gene expression [12]. iPSC induction was performed as described by Takahashi and Yamanaka [1]. iPSC induction shown in Supplementary Fig. S6 was conducted with retroviruses carrying Cnot2 and/or Trim28 in addition to those carrying the four reprogramming factors (Oct3/4, Sox2, Klf4, and c-Myc).
Establishment of partial iPSC lines
To isolate partial iPSC lines, MEFs bearing a Nanog-GFP reporter gene were subjected to iPSC induction. Colonies with ESC colony-like morphology at 3 or 4 weeks post-iPSC induction were individually recovered and maintained on feeder cells. Among them, clones exhibiting strong DsRed, but not GFP, fluorescence were chosen as candidates for partial iPSC clones. After elimination of clones that showed spontaneous conversion to genuine iPSCs, two clones designated as 2B1 and 5C5 were selected based on specific criteria. These criteria included high expression of retroviral genes and almost complete lack of expression of principal pluripotency marker genes because of strong DNA methylation. Finally, these clones were subjected to single cell sorting to ensure clonality.
iPSC culture and expression of exogenous genes by retroviral infection
Partial and genuine iPSCs were cultured in the standard mouse ESC medium supplemented with fetal bovine serum and leukemia inhibitory factor (LIF) on feeder cells unless indicated otherwise. To convert the majority of partial iPSCs to genuine iPSCs, partial iPSCs were subjected to the 2i (MAPK and GSK3β inhibitors) + LIF condition with knockout serum replacement. For genome-wide screening, partial iPSCs were cultured in standard mouse ESC medium or under the 2i condition with serum. Cnot1, Cnot2, Cnot3, Trim28, and Nanog were expressed individually or in combination in partial iPSCs by retroviral infection. For individual expression of genes, G418 was used for stable selection. Even for combinatorial expression of multiple genes, only a single gene was expressed for each infection in which appropriate antibiotics, such as hygromycin and zeocin, were used for selection.
Reagents and antibodies
The following reagents were used at the indicated concentrations unless stated otherwise: 1.5 μg/mL puromycin (Sigma); 600 μg/mL hygromycin, 300 μg/mL Zeocin (Invitrogen); 400 μg/mL G418 (Sigma); 1 μM PD0325901 (Axon Medchem); 3 μM CHIR99021 (Axon Medchem); anti-Cnot1 (14276-1-AP), anti-Cnot2 (10313-1-AP) (ProteinTech); anti-Cnot3 (H0004849-MO1) (Abnova); anti-Trim28 (KAP1) (ab10483) (Abcam); anti-β-actin (sc-47778) (Santa Cruz Biotechnology).
Western blotting
For western blotting, proteins were resolved by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE), transferred to a polyvinylidene difluoride membrane, and then probed with the indicated primary antibodies and appropriate secondary antibodies conjugated to horseradish peroxidase. Specific protein bands were detected by an enhanced chemiluminescence system (GE Healthcare).
Single cell sorting
To ensure clonality, partial iPSCs (2B1 and 5C5) and their derivative iPSCs were subjected to single cell sorting by fluorescence-activated cell sorting (FACS) of GFP−/DsRed+ and GFP+/DsRed− cells. Among the cell clones, cells exhibiting continuous expansion without a change in reporter gene expression were recovered as stable cell lines.
Production of retroviruses for shRNA-mediated knockdown
Retroviral vectors for shRNAs against the expression of Cnot1, Cnot2, Cnot3, Trim28, and luciferase were produced by subcloning the following oligonucleotides together with their complementary sequences into pLKO.1-puro vectors:
Cnot1 KD:
5′-CCGGTGGTTAGGAATGATCACATTACTCGAGTAATGTGATCATTCCTAACCATTTTTG-3′;
Cnot2 KD:
5′-CCGGCTCTTAGCTGCGGTAGAACTTCAAGAGAGTTCTACCGCAGCTAAGAGTTTTTG-3′;
Cnot3 KD:
5′-CCGGGATAAGAAGAGAGGCCGATTTCAAGAGAATCGGCCTCTCTTCTTATCTTTTTG-3′;
Trim28 KD:
5′-CCGGGGACTACAATCTGATTGTTATCTCGAGATAACAATCAGATTGTAGTCCTTTTTG-3′;
Lucifease KD:
5′-CCGGCTTACGCTGAGTACTTCGATTCAAGAGATCGAAGTACTCAGCGTAAGTTTTTG-3′.
U6-shRNA-puro fragments were recovered after NotI and NsiI double digestion and then transferred to pMX-based vectors.
To produce viruses carrying a specific shRNA expression unit, vectors bearing the above sequences were individually transfected into Plat-E cells [13] by lipofection. Partial iPSCs (2B1) were infected with the generated retroviruses in the presence of 8 μg/mL polybrene, and then selected with 1.5 μg/mL puromycin.
PiggyBac-mediated expression of Cnot3 and/or Trim28 in partial iPSCs (2B1)
Cnot3-IRES-hygromycin and Trim28-IRES-neomycin resistance genes were individually subcloned into piggyBac vectors carrying the constitutively active chicken actin gene promoter [14]. These plasmid vectors were introduced into partial iPSCs (2B1) individually or in combination together with the piggyBac transposase expression vector by lipofection. Then, stable cell lines were obtained by selection with G418 and/or hygromycin.
Bisulfite sequencing
Bisulfite sequencing of genomic DNA was performed using the same primers described by Kanatsu-Shinohara et al. [15] for the Oct3/4 gene promoter and those described by Mikkelsen et al. [7] for Nanog, Utf1, Dppa5, and Rex1 gene promoters.
Microarray and bioinformatics analyses
In vitro-transcribed biotin-labeled cRNA was synthesized according to the Affymetrix guidelines. Labeled samples were hybridized to Affymetrix GeneChip Mouse Genome 430 2.0 arrays following the manufacturer's instructions. Microarray expression data were background-subtracted and normalized using the robust multiarray analysis method [16] with R-package 2.11.1 and Bioconductor 2.6 [17]. Spotfire X.X (TIBCO) was used for hierarchical clustering. Gene ontology (GO) analysis was performed using DAVID 6.7 software [18,19].
Accession number
DNA microarray data have been deposited in the NCBI Gene Expression Omnibus under accession number GSE44339.
Results
Isolation of partial iPSC lines
As the first step for identification of genes that participate in the transition from partial to genuine iPSCs, we generated our own partial iPSC lines. MEFs from embryos carrying a Nanog-GFP reporter gene were used for iPSC induction [12]. To induce iPSCs, the MEFs were infected with a retrovirus carrying the DsRed reporter gene together with the four reprogramming factors (Oct3/4, Sox2, Klf4, and c-Myc) to monitor infection efficiency and as a marker to distinguish between partial and genuine iPSCs. At 3 or 4 weeks post-iPSC induction, colonies with ESC-like morphology were randomly picked up and transferred to 24-well plates. During expansion of the picked up colonies, two cell clones termed 2B1 (Fig. 1A) and 5C5 (Supplementary Fig. S1; Supplementary Data are available online at

Generation of partial induced pluripotent stem cell (iPSC) clones.
Identification of the Cnot2 gene as a regulator participating in the transition from partial to genuine iPSCs
To identify genes involved in the conversion from partial to genuine iPSCs, we introduced a piggyBac gene-trap vector containing the murine stem cell virus enhancer/promoter [20] into the two partial iPSC clones bearing the Nanog-GFP transgene. The cells were cultured under the conventional LIF + serum mouse ESC medium condition, and the vector was allowed to move throughout their genomes by transposase-mediated transposition. However, several trials did not produce any clones expressing the GFP reporter gene that was under the control of the Nanog gene promoter. We assumed that these failures were attributed to the requirement for the activation of more than two critical genes for the transition, which could not be achieved by the employed method. However, we considered that changing the culture condition may overcome this problem. Although kinase inhibitors (2i) are usually used together with LIF under a serum-free condition [8,21,22], an equivalent 2i + LIF condition with serum did not induce conversion of partial iPSCs to GFP-positive iPSCs (Supplementary Fig. S1A). Therefore, we used the 2i + LIF condition with serum as a starting condition for the screen (Fig. 2A). After screening, we obtained three GFP-positive iPSC clones (Fig. 2B). Next, genomic DNAs from these clones were recovered to determine the genomic integration sites of the vector. Figure 2C shows the integration site of the vector in each clone. Among the three clones, we chose clone 5 for the following reasons. First, piggyBac vector insertion in clone 5 did not disrupt the coding region of the Cnot2 gene because of the presence of the initiating methionine codon in the second exon (Fig. 2D). More importantly, reverse transcription-PCR analyses revealed that exogenous reprogramming factor genes were almost completely silenced in clone 5, but not in clones 2 or 3 (Supplementary Fig. S2B). Figure 2E shows the presence of Cnot2 mRNA fused with the sequence from the piggyBac vector in clone 5.
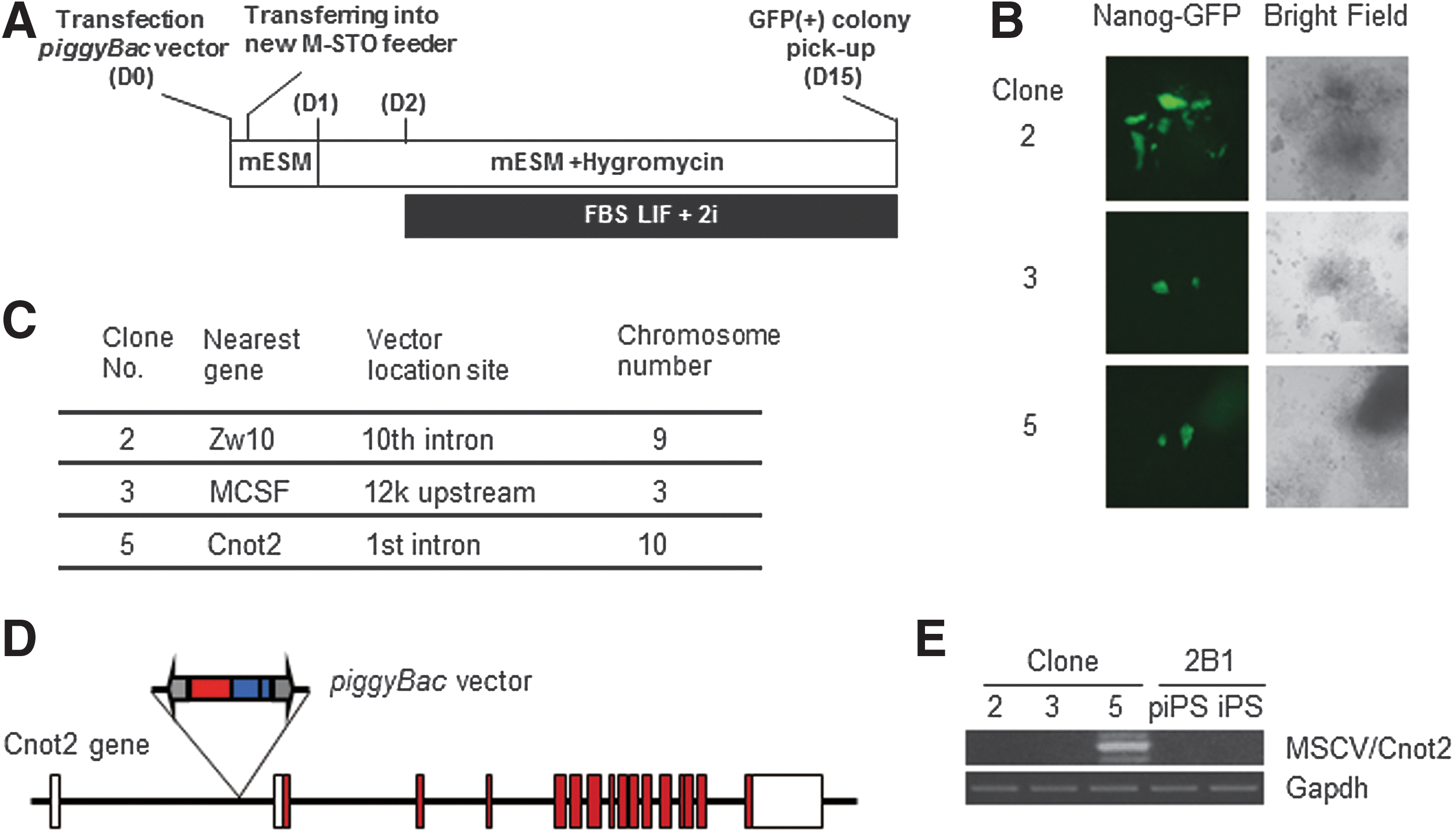
Identification of the Cnot2 gene by screening with the piggyBac vector as a positive regulator of conversion from partial to genuine iPSCs.
Evidence of the involvement of Ccr4-Not complex components in the transition from partial to genuine iPSCs
To confirm the participation of Cnot2 in the transition from partial to genuine iPSCs, we first performed knockdown experiments of the Cnot2 gene. Consequently, the reduction of Cnot2 expression significantly lowered the conversion efficiency to genuine iPSCs under the 2i + LIF condition (Fig. 3A). Cnot2 is one of the subunits of the Ccr4-Not complex in which Cnot1, 2, and 3 comprise the core of the complex. It has been demonstrated by chromatin immunoprecipitation analyses that Cnot3 shares genomic binding sites with Trim28 [11,23]. Therefore, we examined whether these factors also contributed to the transition from partial to genuine iPSCs. As a result, the knockdown of Cnot1, Cnot3, or Trim28 expression equally lowered the conversion efficiency to that of Cnot2 knockdown (Fig. 3). Next, we examined the effect of forced expression of these genes on the conversion of partial iPSCs to genuine iPSCs. None of these factors, even in combination, were able to exert a significant effect on the conversion (Fig. 4). However, we observed some positive effects using the combination of Cnot2 or Cnot3 with Trim28, although these effects were extremely subtle compared with those obtained under the regular 2i condition (comparison of GFP-positive cell numbers in Figs. 3 and 4). Although we obtained unexpected results from the above overexpression experiments, we examined whether GFP-positive cells generated by forced expression of Cnot2 and Trim28 could expand stably as an iPSC clone. To this end, we cultured partial iPSCs overexpressing Cnot2 and Trim28 for 2 weeks at a subclonal density and identified a GFP-positive/DsRed-negative clone (Supplementary Fig. S3A). After picking up the clone and transferring to a new dish, GFP-positive cells were expanded and then sorted by FACS. These sorted cells propagated robustly, while maintaining a high level of GFP expression that faithfully represented endogenous Nanog gene expression (Supplementary Fig. S3B). Quantitative PCR (Supplementary Fig. S3C) and western blot (Supplementary Fig. S3D) analyses showed that these cells were equivalent to iPSCs obtained by exposure of partial iPSCs (2B1) to the KSR-LIF + 2i condition in terms of their expression levels of endogenous pluripotency marker genes. Even with successful establishment of the iPSC clone, the extremely low efficiency of transition from partial to genuine iPSCs by forced expression of Cnots and/or Trim28 (Fig. 4) was not compatible with the fact that piggyBac gene trap vector-mediated activation of the Cnot2 gene sufficiently supported conversion from partial to genuine iPSCs (Fig. 2). A possible cause of this discrepancy may lie in the different methods used for the expression of exogenous genes between the two systems in which retroviral-mediated expression would be subjected to severe silencing, whereas piggyBac-mediated expression would persist even after conversion to genuine iPSCs. Therefore, we chose the piggyBac system to express Cnot3 and/or Trim28. However, the vast majority of partial iPSCs remained positive and negative for DsRed and GFP, respectively, even after expression of the two factors individually or in combination (Supplementary Fig. S4). Therefore, we assumed that partial iPSCs converted to genuine iPSCs by piggyBac-mediated activation of the Cnot2 gene were rather special cells that were much more competent for conversion to genuine iPSCs compared with that of the vast majority of other partial iPSCs (see Discussion for details). Another unexpected result shown in Fig. 4 was that either Cnot2 or Cnot3 together with Trim28 had a stronger effect on the conversion from partial to genuine iPSCs compared with that obtained by simultaneous expression of Cnot1, Cnot2, Cnot3, and Trim28. To gain an insight into the underlying molecular basis of this finding, we compared the expression levels of Cnot2 and Cnot3 under various conditions. As a result, we detected large amounts of these proteins under forced expression of Cnot1, Cnot2, and Cnot3 (Supplementary Fig. S5). This result implies that Cnot1, Cnot2, and Cnot3 proteins are stabilized in partial iPSCs by constituting the core of the Ccr4-Not complex. However, we do not know at present why or how construction of the core of the Ccr4-Not complex led to the decline in efficiency of conversion from partial to genuine iPSCs (see Discussion for more details). Next, we examined whether Cnot2 and/or Trim28 expression elevated the efficiency of reprogramming MEFs. To this end, we added retroviruses carrying Cnot2 or Trim28 individually or in combination together with the retrovirus derived-reprogramming factors (Oct3/4, Sox2, klf4, and c-Myc) for iPSC induction of MEFs. However, the additional factors did not elevate, but rather lowered the reprogramming efficiency as assessed by counting alkaline phosphatase-positive colonies (Supplementary Fig. S6). This observation indicated that forced expression of Cnot2 and/or Trim28 during the early stage of iPSC induction affects the reprogramming process negatively, and the promoting effect of these factors becomes evident only at the later stage of induction. An alternative possibility is that these factors promote the reprogramming process rather restrictively in the transition from partial to genuine iPSCs. These possibilities remain to be addressed at present.

Effect of knockdown of Cnot1, Cnot2, Cnot3, or Trim28 in partial iPSCs on the production of Nanog-GFP-positive iPSCs. Partial iPSCs (2B1) were infected with a retrovirus expressing a puromycin-resistance gene and shRNA targeting the indicated genes. Transfectants were enriched by puromycin selection (1.5 μg/mL) and then exposed to the 2i condition for iPSC conversion. GFP-positive cells were quantified by flow cytometry. Data are the mean±standard deviation (n=3). **P<0.01. Lower panels show western blots examining the efficiency of knockdown.

Effect of overexpression of Cnot1, Cnot2, Cnot3, and Trim28 on the production of Nanog-GFP-positive cells from partial iPSCs. Partial iPSCs (2B1) cultured in the empirical mouse ESC medium were subjected to the overexpression of Cnot1, Cnot2, Cnot3, and Trim28 individually or in the combinations as indicated. After stable integration, MAPK and GSK3β inhibitors (2i) were added to the empirical mouse ESC medium containing leukemia inhibitory factor and serum. Data are the mean±standard deviation (n=6). GFP-positive cells were quantified as described in Fig. 3. Lower panels show western blots confirming forced expression of Cnot1, Cnot2, Cnot3, and Trim28 that were overexpressed individually.
Effect of expression of the core components of the Ccr4-Not complex on the global gene expression profile of partial iPSCs
Although there was only a marginal effect of the combinatorial expression of Cnot2 and other factors on the transition from partial to genuine iPSCs, we examined global changes of gene expression caused by forced expression of these factors in partial iPSCs by microarray analyses. We performed these analyses because, although these factors were apparently insufficient to convert the majority of partial iPSCs to genuine iPSCs, they may be sufficient for the modulation of at least some aspects of partial iPSCs to make them somewhat closer to genuine iPSCs. RNA was recovered from the stable cell lines in which Cnot1, Cnot2, Cnot3, and Trim28 were overexpressed individually or in combinations. Because these cells were cultured without any kinase inhibitors (empirical mouse ESC culture condition) during antibiotic selection and expansion, no cells were positive for Nanog-GFP. First, we characterized downregulated genes. We found that genes downregulated by expression of Cnot1, Cnot2, or Cnot3, or simultaneous expression of all three in partial iPSCs were significantly overlapped (Supplementary Fig. S3A). Indeed, 551 genes were commonly downregulated by more than twofold by these factors. The 551 genes were then divided into three groups consisting of genes commonly regulated by Trim28 and core components of the Ccr4-Not complex, and those regulated specifically by either one (Fig. 5A and Supplementary Table S1). We first examined whether these genes showed relatively lower levels of expression in iPSCs compared with those in partial iPSCs by gene set enrichment analyses (GSEA). These analyses revealed that, with all three sets, most, if not all, genes showed a strong tendency of lower expression in genuine iPSCs compared with that in partial iPSCs (Fig. 5B). These results supported the idea that downregulation of these genes by forced expression of the core components of the Ccr4-Not complex and/or Trim28 represents a step in the process required for transition from partial to genuine iPSCs. Analyses of the expression of these genes with box plots also provided results consistent with this notion (Fig. 5C). Notably, the three gene sets did not alter their average expression levels significantly by forced expression of Nanog that is essential for conversion from partial to genuine iPSCs [24]. These results imply that the roles of Ccr4-Not/Trim28 and Nanog are rather distinct in the conversion. To correlate the downregulation of these genes with overall molecular function, we next conducted GO classification analyses. GO terms related to development and morphogenesis were significantly enriched in genes downregulated specifically by core components of the Ccr4-Not complex and those downregulated commonly with Trim28 (Fig. 6A). This finding indicates that the major role of the core components of the Ccr4-Not complex with or without Trim28 is suppression of genes related to developmental processes. We also found that GO terms related to transcription were closed up among genes downregulated specifically by Trim 28. Next, the upregulated genes (Supplementary Figs. S7B and S8, and Supplementary Table S1) were subjected to the same analyses. However, unlike the downregulated genes, GSEA and comparison of average gene expression levels revealed that genes upregulated by core components of the Ccr4-Not complex and/or Trim28 in partial iPSCs did not show any prominent expression in genuine iPSCs compared with that in partial iPSCs (Supplementary Fig. S8B, C). This result strongly opposed the idea that upregulation of these genes affected conversion of partial iPSCs to genuine iPSCs. Accordingly, GO classification analyses yielded only a few significant terms (37 terms among downregulated genes vs. two terms among upregulated genes) (Fig. 6B). This observation suggests that the genes upregulated by Cnots and/or Trim28 in partial iPSCs are not well-ordered gene sets for specific biological outcomes, but rather disordered gatherings. Therefore, the alteration of expression levels of these genes may represent indirect regulation by Cnots and/or Trim28. Taken together, our data suggest that core components of the Ccr4-Not complex may participate in the transition from partial to genuine iPSCs by repressing rather than activating gene expression and that the major targets of regulation are genes related to developmental and morphogenetic processes.
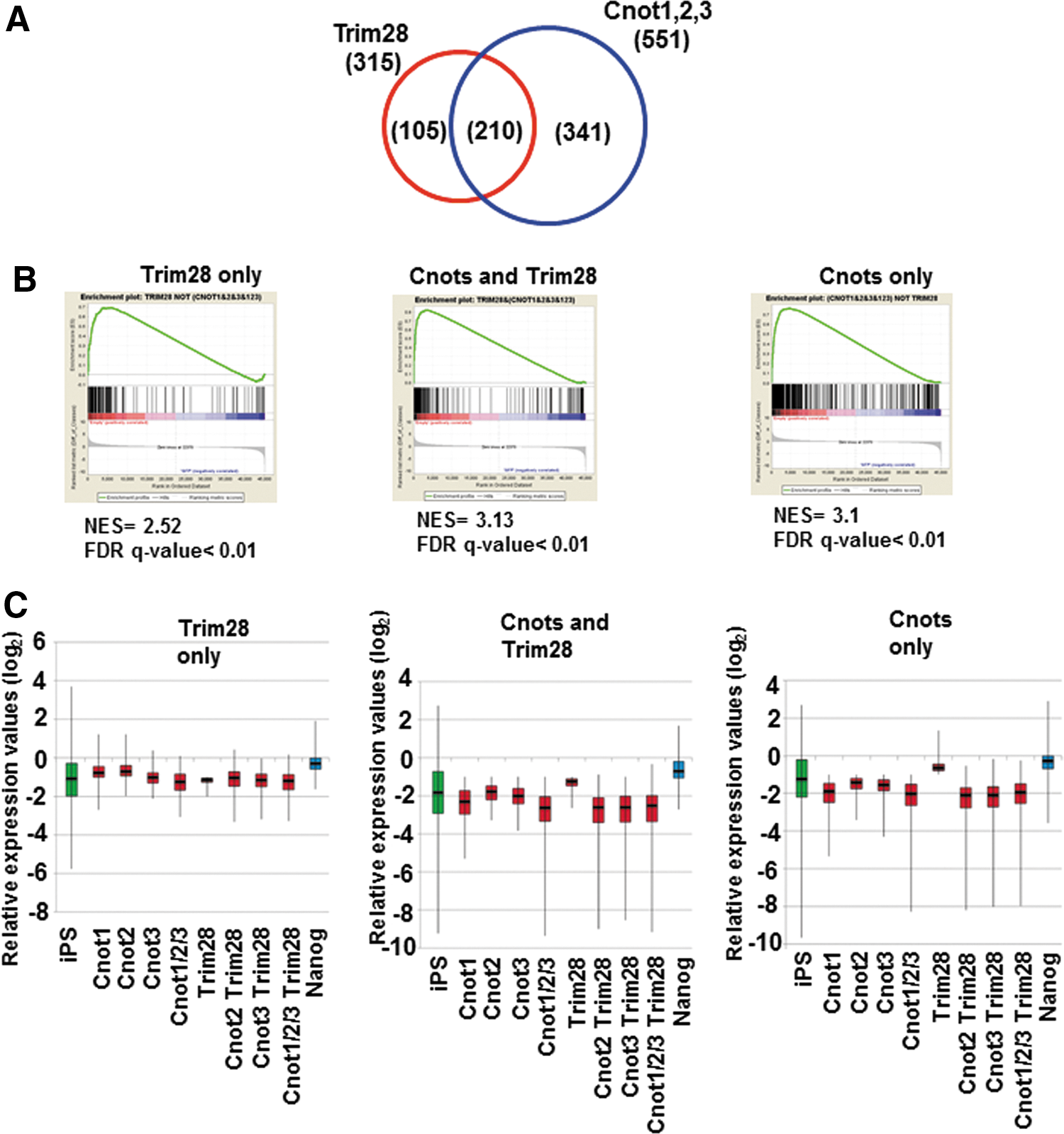
Characterization of genes downregulated by Ccr4-Not complex components and/or Trim28.
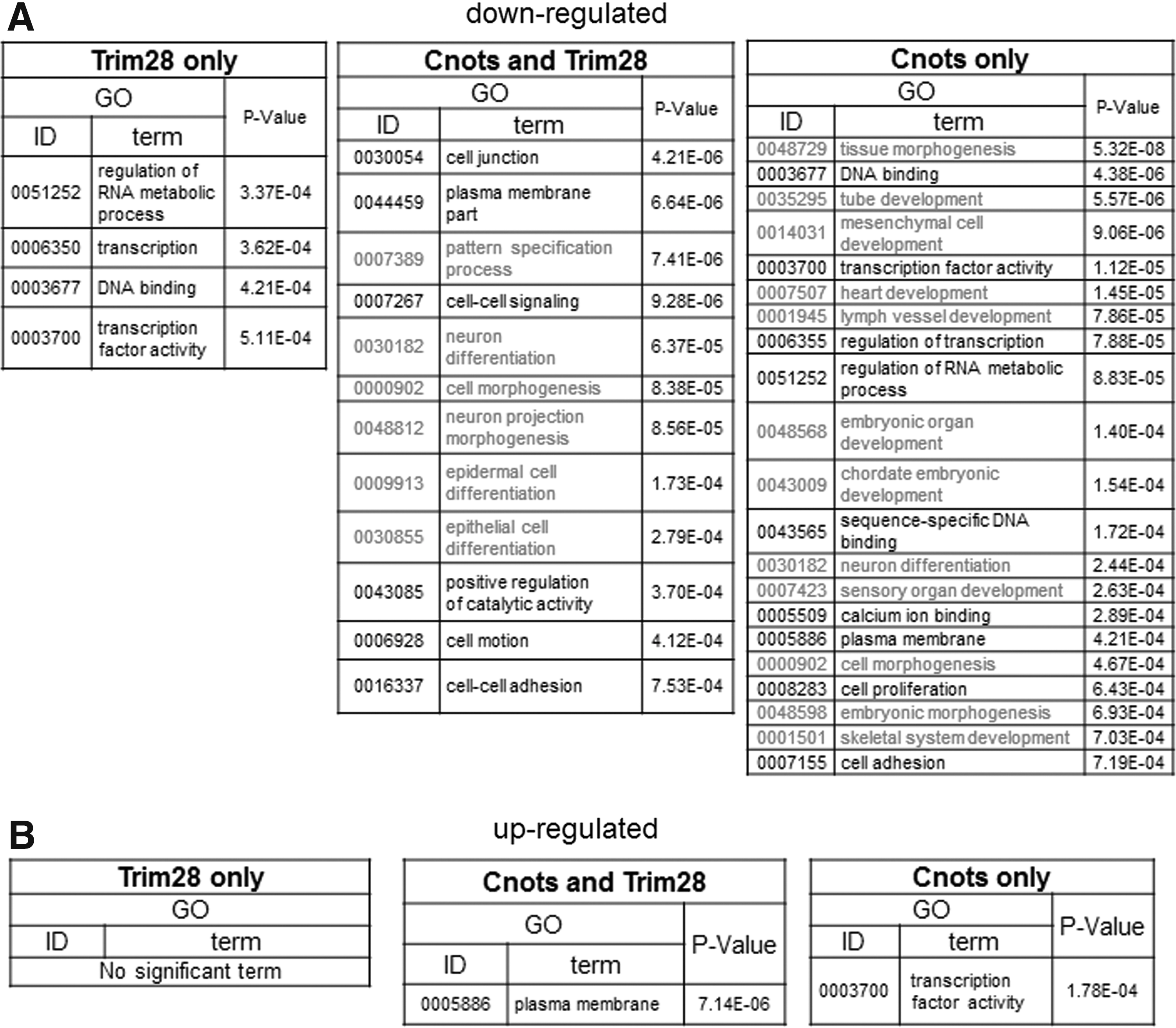
Gene ontology (GO) terms associated with gene sets showing differential expression caused by forced expression of Ccr4-Not complex components and/or Trim28.
Discussion
It was originally thought that the partial iPSC state is an absolute barrier that somatic cells must overcome for conversion to genuine iPSCs [6,7]. However, recent investigation by Polo et al. [5] revealed that partial iPSCs do not resemble any of the intermediate states of cells en route to becoming genuine iPSCs, suggesting that these cells have strayed from the normal reprogramming route at an early point and become immortalized. Therefore, to raise the efficiency of iPSC induction by avoidance of partial iPSC generation, two methods can be considered to prevent straying from the normal reprogramming route to partial iPSCs or correct partial iPSCs back to the normal reprogramming route. The use of L-Myc instead of c-Myc as one of the reprogramming factors appears to correspond to the former type of invention [25]. Glis1 has been also shown to effectively lower the probability of partial iPSC generation [26]. Despite these rather effective former-type inventions, no efficient method for conversion of partial to genuine iPSCs has been devised, except for exposure to the 2i condition. However, our genome-wide screening with the piggyBac vector identified the Cnot2 gene encoding one of the core components of the Ccr4-Not complex as a candidate for supporting such conversion. Subsequent knockdown studies revealed that not only Cnot2 but also other core components of the Ccr4-Not complex, that is, Cnot1 and Cnot3, were also crucially involved in the transition. However, forced expression of either one of these genes or their combinatorial expression including that of Trim28 showed no or only a marginal effect corresponding to about 1% of the effect obtained under the 2i condition. These results were not compatible with the fact that piggyBac vector-mediated activation of the Cnot2 gene was sufficient for the production of genuine iPSCs from partial iPSCs. One possible explanation for this discrepancy is that partial iPSCs converted to genuine iPSCs during the functional screening were rare among the partial iPSC population, and factors other than Cnot2 required for the transition were almost fully expressed, making the cells highly competent for conversion to genuine iPSCs. Another unexpected finding was that expression of either Cnot2 or Cnot3 alone supported more efficient transition from partial to genuine iPSCs in collaboration with Trim28 compared with that of simultaneous expression of Cnot1, Cnot2, Cnot3, and Trim28. Compared with individual expression of either Cnot2 or Cnot3, our western blot data revealed that larger amounts of Cnot2 and Cnot3 when all three core components of the Ccr4-Not complex, that is, Cnot1, Cnot2, and Cnot3, were expressed together. The most likely explanation for this finding is that Cnot2 and Cnot3 are more stable in the Ccr4-Not core complex compared with that of their free forms. If this is indeed the case, our results indicate that there is an inverse relationship between the formation of the Ccr4-Not core complex and the promotion of the transition from partial to genuine iPSCs. More specifically, our data suggest that Cnot2 and Cnot3 may be able to support the transition only in their free forms. Although we do not have any evidence to support this hypothesis at present, we assume that generation of Cnot2 or Cnot3 mutants that are defective for complex formation may resolve this issue. Our microarray data and subsequent GO classification analyses suggested that the major role of the core components of the Ccr4-Not complex with or without Trim28 in the transition of partial to genuine iPSCs is to suppress the expression of genes related to developmental processes. Currently, we do not know how these core components perform this function. It is known that the expression of developmental genes is largely repressed in ESCs by a specialized chromosomal architecture termed as the poised bivalent chromatin structure that harbors both transcriptionally active (histone 3-lysine 4 trimethylation) and repressive (histone 3-lysine 27 trimethylation) epigenetic marks [27 –30]. Interestingly, a substantial number of genes that should be regulated by the bivalent chromatin structure in ESCs do not bear a complete bivalent architecture in partial iPSCs [7]. Moreover, Cnot3 and Trim28 are substantially colocalized with bivalent genes in ESCs [11,23]. Therefore, it is tempting to speculate that expression of the core components of the Ccr4-Not complex and/or Trim28 participates in construction of the bivalent chromatin structure in partial iPSCs, leading to the downregulation of developmental genes. The roles of the core components of the Ccr4-Not complex in suppressing the expression of developmental genes have been demonstrated previously with ESCs [31] in which impairment of the function of the complex leads to differentiation of ESCs toward trophectodermal cell lineages. Because our data did not indicate any specificity of the genes involved in differentiation toward trophectodermal cells, it is assumed that the common role of the complex is to suppress development/differentiation-associated genes, but sets of genes subjected to the repression are cell type specific.
In summary, as depicted in Fig. 7, we demonstrate here that core components of the Ccr4-Not complex with or without Trim28 are strongly involved in the transition of partial to genuine iPSCs mainly through the suppression of genes related to development. We hope that these findings will provide a clue to fully understanding the entire process of this transition at the molecular level and eventually lead to increasing the induction efficiency and quality of iPSCs.

Summary of requirements for transition from partial to genuine iPSCs. To convert partial iPSCs to genuine iPSCs, at least three major events must occur, that is, repression of genes related to development/differentiation, silencing of retroviral DNAs [32 –34], and acquisition of endogenous pluripotency gene expression [6 –8]. Our present study indicates that Ccr4-Not complex components and Trim28 at least in part contribute to one of these events, namely repression of developmental genes.
Footnotes
Acknowledgments
We thank Dr. Austin Smith for the piggyBac vector and Tomoko Okuda for technical assistance. This study was performed as a part of the Core Research for Evolutional Science and Technology (CREST) Agency. This study was supported in part by the Ministry of Education, Culture, Sports, Science and Technology (MEXT), Japan, and primarily by a Grant-in-Aid for the Support Project of Strategic Research Center in Private Universities to the Saitama Medical University Research Center for Genomic Medicine. A.O. is a recipient of grants from the Japan Society for the Promotion of Science (JSPS) KAKENHI (grant numbers 25293082 and 25670147).
Author Disclosure Statement
The authors declare that they have no conflict of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.