
Abstract
DNA methylation constitutes a major obstacle in the reprogramming of cells to pluripotency. Although little is known regarding the molecular mechanisms of DNA demethylation, activation-induced deaminase (AID), which is known to function in antibody diversification, has been implicated in DNA demethylation through a base excision repair (BER)-mediated pathway. Here we comprehensively examine the plausibility of coupled AID-BER demethylation in the generation of induced pluripotent stem cells (iPSCs) and show that AID is dispensable for reprogramming cells into iPSCs. Additionally, the overexpression of AID and other factors involved in AID-coupled DNA demethylation does not increase the efficiency of reprogramming. Moreover, BER is not likely to play a role in this process. Our results indicate that the reactivation of key genes governing the pluripotency circuitry occurs through a mechanism that is independent of deamination-coupled demethylation.
Introduction
E
Accordingly, our study focused on understanding the mechanism of DNA demethylation during the reprogramming of somatic cells into iPSCs. By generating iPSCs from either AID-deficient or shRNA-mediated AID-depleted fibroblast cells, we show that AID is dispensable for iPSC generation and the maintenance of pluripotency in vitro. Our results suggest that AID-coupled DNA demethylation does not occur during factor-mediated reprogramming. Furthermore, other factors implicated in AID-dependent demethylation do not appear to have a crucial role in the establishment of pluripotency.
Materials and Methods
Cell culture
Mouse embryonic fibroblasts (MEFs) were derived from day 13.5 embryos from OG2 transgenic mice, a mouse strain that carries GFP under the control of the Oct4 promoter (Oct4-GFP) and from AID deficient (KO) transgenic mice [16]. MEFs were cultured in FibroGRO Low Serum (FGRO) medium (Millipore, Billerica, MA). iPSCs and mouse ESCs (mESCs) were maintained on a mitomycin C-inactivated MEF feeder (Millipore) layer in mESC medium consisting of 85% KnockOut Dulbecco's modified Eagle's medium (KO DMEM; Gibco, Invitrogen, Carlsbad, CA), 15% Knockout Serum Replacement (KOSR) (Gibco), 1 mM L-glutamine, 0.1 mM nonessential amino acids, 0.1 mM 2-mercaptoethanol, and 1,000 U/mL ESGRO (all from Millipore) or in 2i medium [17].
Retroviral infection and iPSC generation
The pMXs-Oct4, pMXs-Sox2, and pMXs-Klf4 plasmids were obtained from Addgene (Cambridge, MA). The other genes used in this study were purchased from Korea Gene Bank (Daejeon, Korea) and were cloned into the retroviral pMX vector linked to an HA-tag at the C terminus using the PacI and XhoI restriction sites. The clones were verified by sequencing and the expression of viral constructs was confirmed by immunostaining (Supplementary Fig. S2; Supplementary Data are available online at
The virus-containing supernatants were collected at 48 h after transfection and filtered through a 0.45-μm Millex-HV (Millipore) filter. The viral supernatants were concentrated using Retroconcentin (SBI; Mountain View, CA) according to the manufacturer's instructions. Low-passage MEFs (within three passages) were seeded at a density of 1×105 cells per well in a six-well plate 12 h before infection. The infections were performed in FGRO media supplemented with 4 mg/mL polybrene (Millipore). After an overnight incubation, the cells were washed with PBS, and 3 mL iSF1 [18] medium was added. iPSC colonies were isolated at 8 to 10 days postretroviral transduction based on the expression of Oct4-GFP and/or ESC morphology.
Fluorescence-activated cell sorting
Cells were dissociated using Accutase (Millipore) and resuspended into single-cell suspensions in PBS solution containing 2% FBS (FACS buffer). Oct4-GFP+ cells were analyzed using a flow cytometer (FACSCalibur; BD Bioscience, Heidelberg, Germany), and the data were analyzed using CellQuest software (BD Bioscience).
Alkaline phosphatase staining
The alkaline phosphatase (AP) staining was performed using Alkaline Phosphatase Staining Kit II (Stemgent, San Diego, CA) according to the manufacturer's instructions. The cells were photographed using a Nikon Eclipse Ti camera (Nikon, Japan).
Immunofluorescence
iPSCs and MEFs were fixed with 4% paraformaldehyde and permeabilized with 0.1% Triton X-100 in phosphate-buffered saline (PBS; Invitrogen) for 30 min at room temperature. The cells were then washed thrice with wash buffer (0.03% Triton X-100 in PBS). The fixed samples were blocked for 1 h with 5% normal goat serum in wash buffer, followed by incubation for 24 h at 4°C with anti-Nanog and anti-HA (Santa Cruz, Dallas, TX) primary antibodies diluted with wash buffer. The samples were washed with wash buffer, and then incubated with TRITC-conjugated secondary antibody (Molecular Probes, Inc., Eugene, OR) for 2 h. The slides were counterstained with DAPI (Vector Laboratories, Burlingame, CA) to stain the cell nuclei. The slides were then photographed using an LSM 510 META confocal microscope (Carl Zeiss, Inc., Oberkochen, Germany).
Real-time PCR
Total RNA was extracted from the cells using RNeasy Mini Kit (QIAGEN, Valencia, CA) according to the manufacturer's instructions, and cDNA synthesis was performed using SuperScript® Vilo cDNA Synthesis Kit (Invitrogen). qRT-PCR was performed in triplicate with SYBR Green I Master Mix (Roche, Mannheim, Germany). The primer sequences used in these assays are listed in Supplementary Table S1.
Bisulfite sequencing analysis
Genomic DNA was extracted using DNeasy Blood & Tissue Kit (QIAGEN). The DNA samples were treated with sodium bisulfite to convert all unmethylated cytosine residues into uracil residues using EpiTect Bisulfite Kit (QIAGEN). The promoter regions of the Oct4 and Nanog genes were amplified by PCR. A 3-μL aliquot of the first-round PCR product was used in the second round of PCR. The PCR products were subcloned into the TA cloning vector (pGEM-T Easy Vector; Promega), and the individual clones were sequenced (Macrogen, Inc., Seoul, Korea). The primer sequences are listed in Supplementary Table S2.
Teratoma formation and immunohistochemical analyses
iPSCs (3×106 cells) were injected into the testicular region of 5-week-old NOD/SCID mice (The Jackson Laboratory, Bar Harbor, ME). The resulting teratomas were explanted 8 weeks later. The teratoma samples were histologically examined after hematoxylin and eosin (H & E) staining of the gut epithelium and the following stains: PAS stain for secretory epithelium, Alcian blue stain for cartilage, and Masson's trichrome stain for muscle fibers. The images were analyzed using inverted microscopy (Nikon).
Enzyme-linked immunosorbent assay
Heat-denatured genomic DNA samples were immobilized on a 96-well plate using Reacti-Bind DNA coating solution (Thermo Scientific, Rockford, IL). After blocking with 5% dry milk in PBS containing 0.05% Tween (PBST), the samples were incubated overnight with an anti-5hmC or anti-5mC antibody (Active motif; 1:5000). The samples were washed with PBST, and then incubated with an HRP-conjugated anti-rabbit lgG antibody (Santa Cruz; 1:1000) for 2 h. The chemiluminescent signals were developed using a TMB substrate (Thermo Scientific) and monitored.
GST pull-down
The plasmids encoding GST-tagged Dnmt3a and HA-tagged AID were transfected into 293T cells. At 48 h after transfection, the cells were lysed in RIPA buffer (Pierce Biotechnology, Rockford, IL) containing protease inhibitors (Complete; Roche, Indianapolis, IN), and the insoluble components were removed by centrifugation. The lysates were incubated with glutathione-Sepharose beads (Amersham Pharmacia Biotech AB, Staffanstorp, Sweden) overnight at 4°C, followed by three washes in RIPA buffer. After washing, the precipitated samples were subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis, and the gel was subjected to an immunoblot analysis using an anti-HA (Santa Cruz) antibody.
Results
AID is dispensable for reprogramming and pluripotency
To identify the role AID in transcription factor-mediated reprogramming, we first reprogrammed MEFs derived from AID-deficient mice (Supplementary Fig. S1A). The MEFs were transduced with retroviral constructs encoding the mouse reprogramming factors Oct4, Sox2, Klf4 (3F), and dsRed; given the auxiliary role of c-Myc in iPSC generation, we did not include c-Myc in the reprogramming cocktail. After infection, the medium was switched to a serum-free medium (iSF1) (Fig. 1A). Individual ESC-like colonies with a round shape and a distinct edge that were dsRed negative were selected between days 10 and 12 postinfection and maintained under standard mESC culture conditions. Six stably expanded 3F AID KO iPSC lines were morphologically indistinguishable from ESCs and maintained strong AP expression (Fig. 1B). Moreover, these iPSCs expressed the typical pluripotency markers Oct4, Nanog, and Rex, as revealed by qRT-PCR (Fig. 1C). A bisulfite analysis of the methylation patterns of the Oct4 and Nanog promoters showed the proper demethylation state in the AID KO iPSCs, whereas the parental MEFs had strong methylation marks, indicating that pluripotency had been established (Fig. 1D).

Reprogramming of activation-induced deaminase (AID) KnockOut (KO) mouse embryonic fibroblast (MEF) cells by OSK.
To verify whether AID KO iPSC pluripotency can be maintained in vitro, the cells were cultured under chemically defined conditions that included GSK3β and MEK1/2 inhibitors (2i condition). The AID KO iPSCs exhibited stable growth and displayed a domed, round-shaped, and tightly packed morphology over 20 passages (Fig. 2A), and an immunocytochemical analysis confirmed that these cells expressed the pluripotency marker Nanog (Fig. 2B). In addition, the pluripotency of the AID KO iPSCs was verified by an in vivo differentiation assay. After subcutaneous injection, the iPSCs formed teratomas with tissues of all germ derivatives, including gut epithelium (endoderm), muscle (mesoderm), and secretory epithelium (ectoderm; Fig. 2C). Furthermore, ELISA assays revealed that the global levels of 5mC and 5hmC in the AID KO iPSCs were similar to 3F wild-type iPSCs, which were previously established and maintained under the same conditions (Fig. 2D and Supplementary Fig. S1B). These analyses revealed that AID is dispensable for the in vitro maintenance of pluripotency.

Characterization of AID KO iPSCs.
Deamination-coupled DNA demethylation is not involved in iPSC generation
We next examined whether the ectopic expression of AID and two other APOBEC family genes, Apobec1 and Apobec3, promote iPSC generation. MEFs carrying GFP under the control of the Oct4 promoter (OG2 MEFs) were transduced with 3F accompanied by pMX AID, Apobec1, Apobec3, or dsRed (Fig. 3A). The expression of the viral constructs was confirmed by immunostaining (Supplementary Fig. S2). The analysis of the reprogramming efficiency showed that the addition of AID, Apobec1, or Apobec3 did not induce an increase in the reprogramming efficiency, as measured by AP staining (Fig. 3B) and FACS (Fig. 3C and Supplementary Fig. S3)
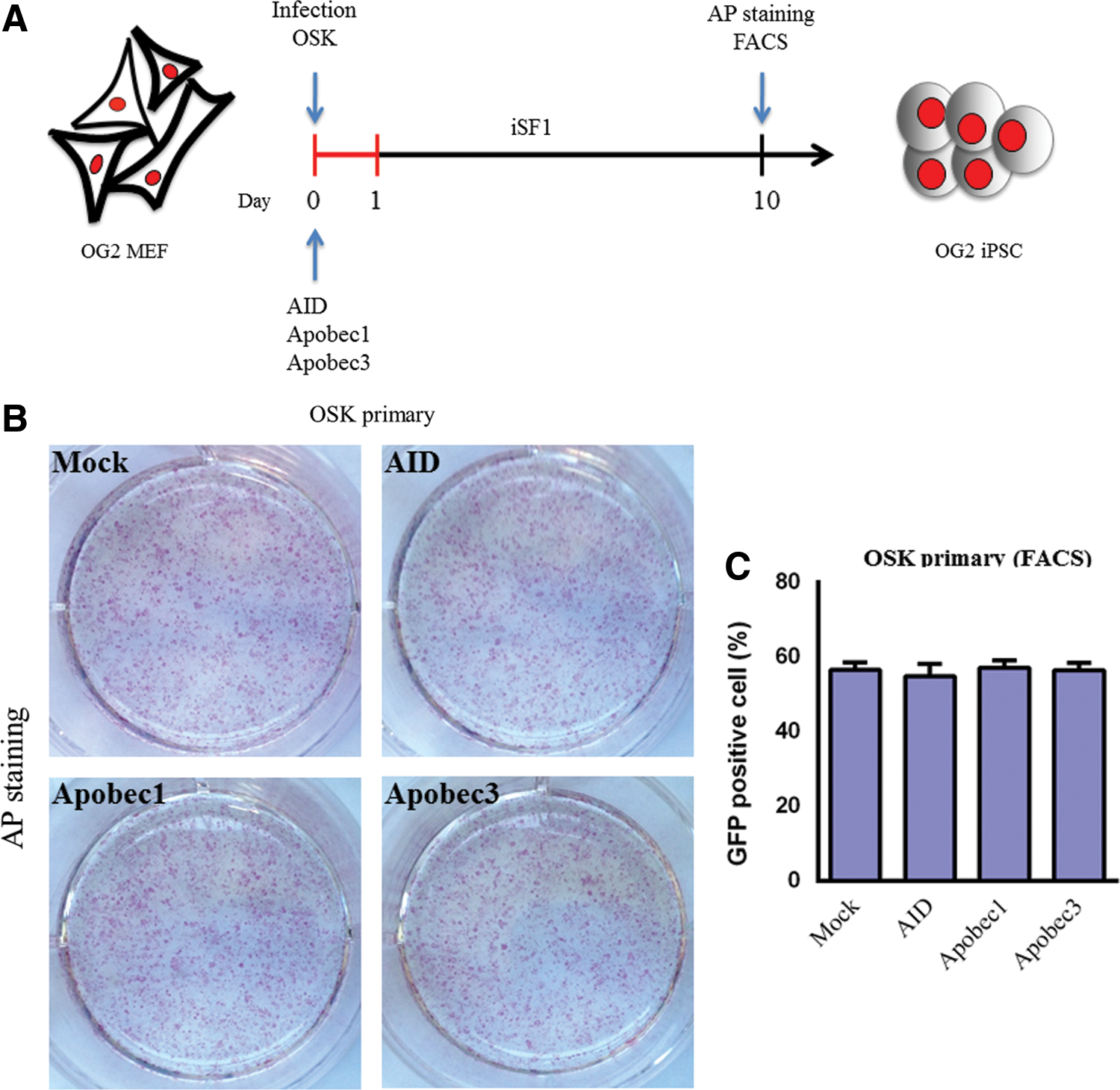
AID overexpression does not affect reprogramming efficiency.
The proposed model for the involvement of AID in DNA demethylation requires the cooperative action of other factors, such as TDG, Gadd45a, and MBD4. Thus, we systematically tested whether the exogenous expression of these genes combined with AID expression increased the overall efficiency of reprogramming (Fig. 4A). However, different combinations of these factors with AID and OSK resulted in no significant changes in iPSC colony formation (Fig. 4B) or GFP-positive cell number when compared to the control cells (Supplementary Fig. S4). These results suggested that AID may not play a role in iPSC generation or that endogenous AID activity is sufficient to obtain pluripotency.

Overexpression of AID partners in iPSC generation.
It has been reported that the acute loss of AID prevents the initiation of reprogramming during iPSC generation when FBS-based culture conditions are used [19]; indeed, it is well known that culture conditions can greatly affect the fate of reprogramming [20]. To assess whether the use of a serum-free system can overcome AID dependence in reprogramming, we employed retroviral-based shRNAs to stably silence endogenous AID during reprogramming. 3F were introduced into OG2 MEF cells together with either AID shRNA or control shRNA (scr shRNA), and the cells were cultured under serum-free conditions (Fig. 5A). The functionality of the shRNAs in downregulating AID expression was assessed in MEFs. AID expression was unaffected by the control shRNA, whereas the AID shRNAs resulted in 60–90% decrease in AID expression, as measured by qRT-PCR (Supplementary Fig. S5A). Intriguingly, when the MEF cells were transduced with 3F plus AID shRNA, iPSCs with ESC-like dome shape morphology and clear boundaries appeared without compromising the reprogramming kinetics (Fig. 5B). We consistently obtained comparable iPSC generation efficiency when compared to the scr shRNA, as assessed by either AP staining or GFP-positive cell number (Fig. 5C and Supplementary Fig. S5B). We characterized the iPSC colonies generated under the acute loss of AID and found that these cells were pluripotent and exhibited normal Oct4 and Nanog promoter methylation patterns (data not shown), suggesting that the demethylation process occurred through an AID-independent mechanism. Additionally, these data indicate that the culture conditions may influence the factor requirements for reprogramming.
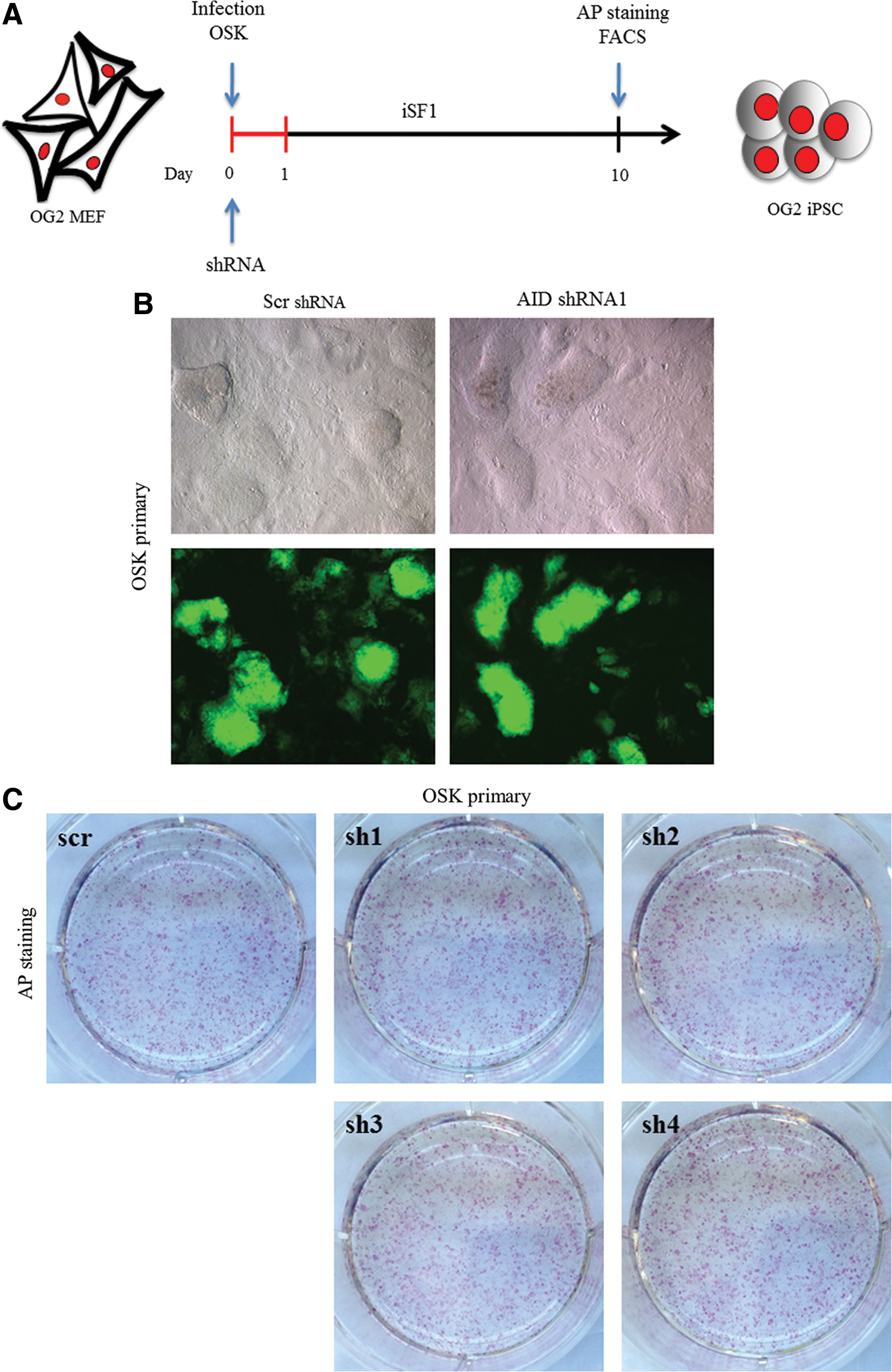
Acute loss of AID does not compromise reprogramming efficiency.
The BER pathway is not associated with the establishment of pluripotency during reprogramming
Lastly, we used chemical inhibitors to examine the potential involvement of the BER pathway, which is indispensable for AID-dependent demethylation (Fig. 6A). Consistent with a recent report [21], the addition of ABT-888, an inhibitor of poly (ADP-ribose) polymerase 1 (PARP1), into the reprogramming medium completely abolished iPSC generation. In contrast, inhibiting APE1, the enzyme that generates the apurinic/apyrimidinic sites to which PARP1 binds, with CRT0044876 did not prevent colony formation (Fig. 6B, C). Both enzymes are critical to the BER pathway, and these two inhibitors block DNA demethylation in mouse zygotes [22]. PARP1 also acts on chromatin remodeling complexes [23,24]. Taken together, these results suggested that the BER pathway was not required for the demethylation of pluripotency genes during iPSC generation. Moreover, it is likely that the PARP1 function is associated with its role in chromatin remodeling.
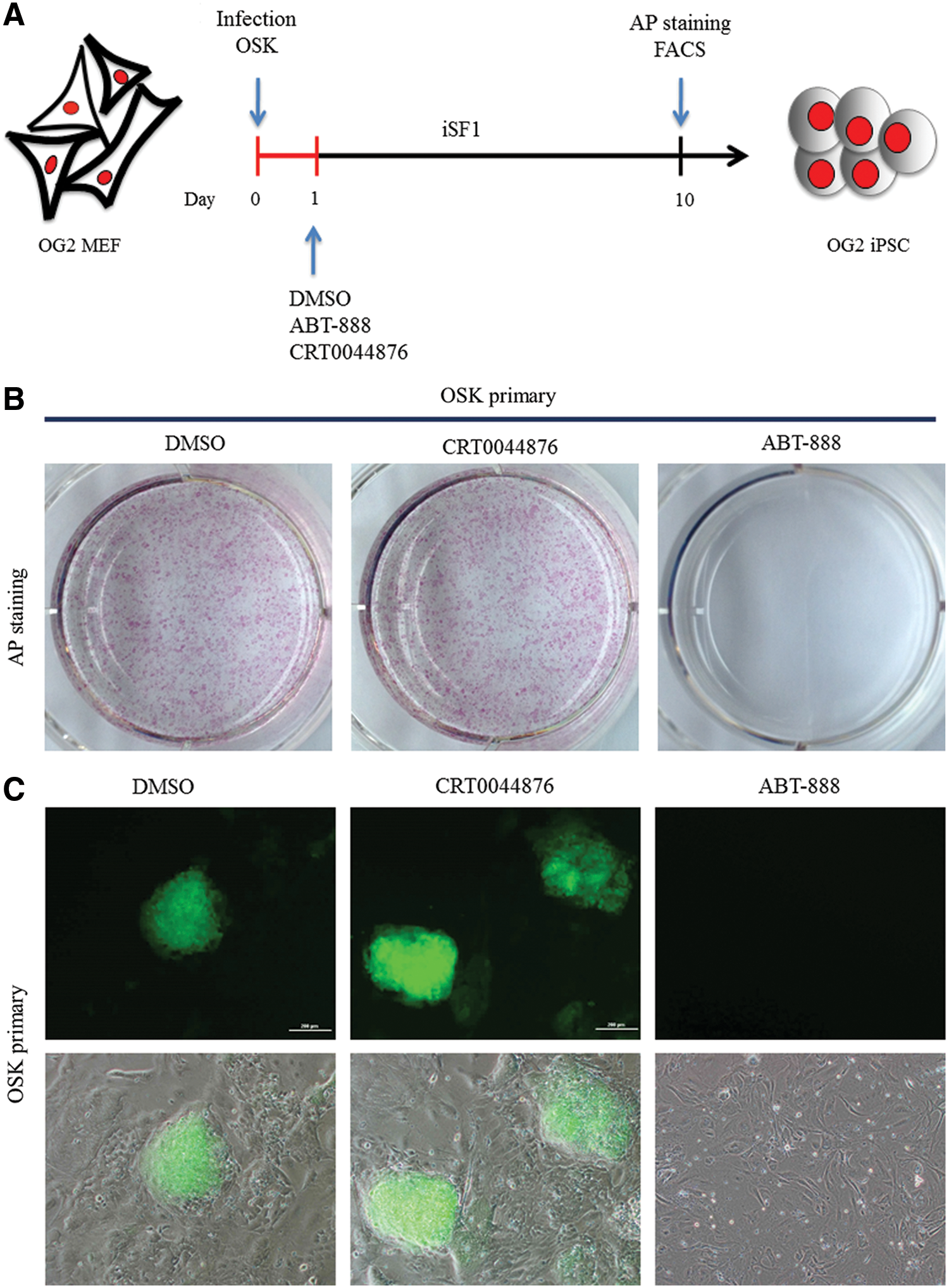
Inhibition of PARP1 and APE1 in iPSC reprogramming.
Discussion
Several studies have suggested that DNA methylation is a major barrier to reprogramming cells that must be removed for the reactivation of pluripotency genes [6,10,25,26]. However, this mechanism remains very controversial. AID, one of the most studied genes in this field, has been proposed to participate in active DNA demethylation [9,17]. In this study, we examined this hypothesis during somatic cell reprogramming through the use of iPSC generation methods to comprehensively identify the effects of AID and other implicated cofactors involved in the deamination-dependent DNA demethylation pathway. To achieve this goal, we generated iPSCs from AID KO MEFs and either AID-overexpressing or shRNA-mediated AID-depleted wild-type MEFs and demonstrated that MEF reprogramming by OSK was independent of AID function. Our results showed that AID KO iPSCs were pluripotent and capable of differentiating into derivatives of all three germ layers (Fig. 2C). In a recent study, a comprehensive methylome analysis of activated B cells in which AID is primarily expressed revealed that AID did not play an active role in the regulation of DNA methylation [26]. Consistent with this report, we found that AID KO iPSCs maintained the DNA methylation and hydroxymethylation patterns similar to wild-type iPSCs (Fig. 2D).
Despite the reports linking AID to active DNA demethylation in vivo and in vitro [11,17,20], AID-deficient mice are viable and fertile [27]. Surani and colleagues demonstrated that AID expression was not detectable in the germ line between E8.5 and E11.5, a time point which the genome-wide loss of 5mC occurs [22]. Moreover, a recent report showed that AID was unlikely to be involved in the demethylation of pluripotency genes during cell fusion-mediated reprogramming [26], which is in conflict with the observations of Blau and colleagues [19]. In our experiments, we observed that the overexpression of AID did not promote iPSC formation. Additionally, the overexpression of Apobec1 and Apobec3, which might compensate for AID activity in AID KO mice [19], showed similar results.
Although Bhutani et al. suggested there are mechanistic similarities between cell fusion and iPSC generation with respect to AID function [19], the time point requiring AID in factor-mediated reprogramming does not coincide with DNA demethylation. It was also stated that the catalytic function of AID was crucial for the initiation of iPSC generation; however, the reactivation of key pluripotency genes, a hallmark of proper demethylation, is a late event [28]. Thus, it appears that the demethylation of the pluripotency genes might not be attributable to AID. Furthermore, the overexpression of TDG and Gadd45a, which form a ternary complex with AID, did not result in increased colony formation in our reprogramming system. Deamination-based DNA demethylation requires the sequential action of BER enzymes. APE1 cleaves the DNA backbone to create a single-strand DNA nick after the removal of a T by glycosylase; PARP1 recognizes this site, and its binding then recruits and activates multi-protein complexes that perform the repair process [10]. However, chemical interventions targeting PARP1 and APE1 revealed that only PARP1 function was essential in iPSC generation. PARP1 is involved in both DNA repair and chromatin remodeling [9,23,29], and Doege et al. showed that PARP1 was rapidly induced in fibroblasts after the initiation of the iPSC generation process and was essential for reprogramming [30]. Our observations suggested that PARP1's role in reprogramming was not due to its function in DNA repair and that the BER pathway might not have an active role in the induction of pluripotency during iPSC reprogramming. Indeed, if the BER pathway was crucial for this process, the inhibition of APE1, which leads to the production of PARP1's substrate, would prevent iPSC generation. Additionally, TDG was shown to participate in active DNA demethylation and is required for normal mammalian development [31] though is dispensable for the generation of iPSCs [32].
An alternative scenario places AID at the center of active DNA demethylation and suggests a Tet-induced oxidation-deamination mechanism [15]. In this scenario, 5mC is oxidized to 5hmC by TET proteins and deaminated to 5-hydroxyuracil (5hmU) by AID; this 5hmU residue is then removed by glycosylase and is ultimately replaced by cytosine through the BER pathway. Although this mechanism has been demonstrated in transfected human cells and in adult mouse brain cells, there is considerable controversy regarding this model. For example, a recent study has revealed that AID has no deamination activity on 5hmC in vitro [26], and further studies have shown that 5hmC is not a simple intermediate product of active DNA demethylation [33 –37].
Because AID can produce off-target DNA damage, its activity is tightly controlled by several mechanisms that affect AID subcellular localization [38]: if AID initiates DNA demethylation, nuclear transport would be the first barrier to overcome. However, several studies have reported that AID remains cytoplasmic even when overexpressed [38 –40]. It has also been shown that Dnmt3a interacts with TDG and re-methylates C residues that are generated through the BER pathway, thereby restoring the deamination of mC [41], and we demonstrated that AID also interacts with Dnmt3a in vitro (Supplementary Fig. S6). Taken together, we suggest that there is a high possibility that Dnmt3a might be a safeguard against the AID-mediated transition from mC to T to ensure the maintenance of a normal methylation status.
In summary, our study provides direct evidence that the AID-mediated DNA demethylation pathway does not play an active role in reprogramming of MEFs into iPSC generation. Our data suggest that passive DNA demethylation is a more appropriate model in transcription factor-mediated reprogramming. Further understanding of the molecular mechanisms underlying reprogramming will be useful in generating fully pluripotent iPSCs that can differentiate into desirable cell types of therapeutic importance.
Footnotes
Acknowledgments
This research was supported by the Bio & Medical Technology Development Program of the National Research Foundation (NRF), which is funded by the Korean government (MEST) (no. 2012-0006107).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.